



Overview
资料来源:蒂尔德·安徒森1号,罗尔夫·卢德1
1临床科学隆德系,感染医学系,生物医学中心,隆德大学,221 00 隆德,瑞典
感染原核生物的病毒,称为噬菌体或简称噬菌体,在20世纪初由Twort(1)和d'Hérelle(2)独立发现。自那时以来,噬菌体因其治疗价值(3)及其对人类(4)以及全球生态系统(5)的影响而得到广泛认可。目前的关切促使人们重新关注使用噬菌体作为现代抗生素替代治疗传染病的方法(6)。基本上,所有的噬菌体研究都依赖于纯化和量化病毒的能力,也称为病毒性小子。最初描述在1952年,这是斑块测定的目的(7)。几十年后,多项技术进步,斑块测定仍然是确定病毒性牙点(8)的最可靠的方法之一。
噬菌体通过将遗传物质注入宿主细胞,劫持机器来生产新的噬菌体颗粒,并最终导致宿主通过细胞解释释放许多后代病毒。由于其微小的大小,噬菌体不能只用光显微镜观察;因此,扫描电子显微镜是必需的(图1)。此外,噬菌体不能像细菌一样在营养琼脂板上培养,因为它们需要宿主细胞来捕食。
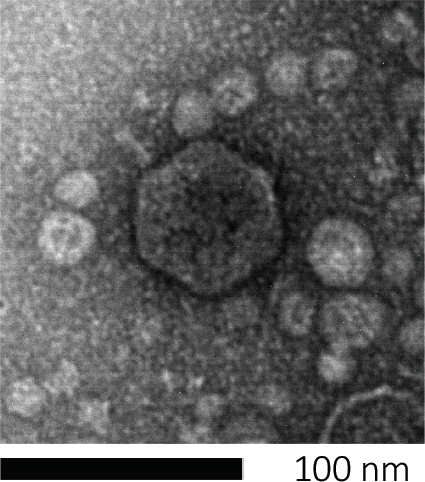
图1:噬菌体形态,这里以大肠杆菌噬菌体为例子,可以用扫描电子显微镜进行研究。大多数噬菌体属于考多病毒(尾噬菌体)。这种特殊的噬菌体有一个很短的尾巴结构和一个食道头,把它放在波多病毒家族。
斑块测定(图2)基于将宿主细胞(优先用于对数相生长)并入培养基。这创造了一个密集,浑浊的细菌层,能够维持病毒生长。分离的噬菌体随后可以感染、在一个细胞内复制和分莱。每个分莱细胞,多个相邻的细胞立即被感染。在几个周期中,在原本浑浊的板块中可以观察到一个清晰的区域(斑块),表明最初是单个噬菌体粒子的存在。因此,样品每体积(即PFU/mL)的斑块形成单位数量可以从产生的斑块数量中确定。
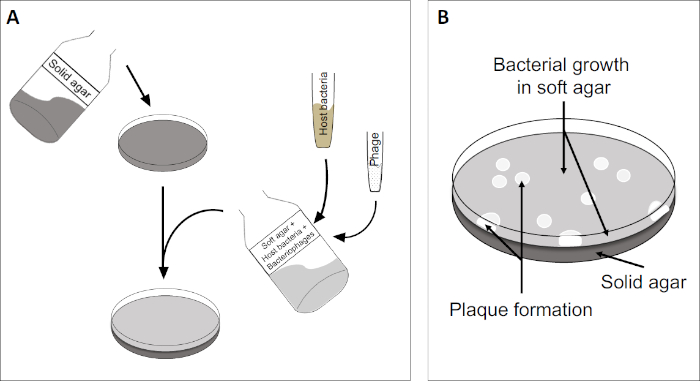
图2:检测斑块形成单元(PFU)是确定样品中噬菌体数量的常见方法。(A)无菌培养皿的基底覆盖着适当的固体营养介质,然后是软培养基、易感宿主细胞的混合物和原噬菌体样本的稀释。请注意,在某些情况下,噬菌体悬浮液也可以均匀地分布在已经凝固的软琼脂表面。(B)宿主细菌的生长在琼脂层形成细胞草坪。噬菌体复制产生由宿主细胞解致引起的透明区域或斑块。
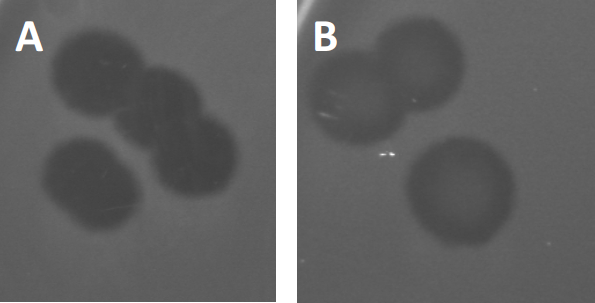
图3:PFU测试结果显示,噬菌体产生的多斑块。由于易感宿主细胞的赖清,斑块可被视为细菌草坪上的清除区,要么具有(A)完全清除,要么(B)由产生耐药细菌(或可能是由耐温噬菌体)引起的部分再生致源循环)。
除了以前描述的溶血生长外,某些温带噬菌体可以采用所谓的溶源性生命周期。在地合体中,病毒通过将其遗传物质纳入宿主细胞(9)的基因组而假定为潜伏状态,这通常对进一步的噬菌体感染具有抵抗力。这有时通过斑块的轻微云彩(图3B)来揭示。然而,值得注意的是,斑块也可能显得模糊,由于细菌的再生长,已经进化了抵抗噬菌体,独立于以前的噬菌体感染。
病毒可以附着或吸附,只有有限的宿主细菌(10)。宿主范围受到细胞内抗病毒策略(如CRISPR-Cas系统(11)的进一步限制。细菌亚群对特定噬菌体表现出的抗药性/敏感性历来被用来将细菌菌株分类为不同的噬菌体类型(图4)。虽然这种方法的有效性现已被新的测序技术所超越,但噬菌体类型仍然可以提供有关细菌-噬菌体相互作用的宝贵信息,例如,促进为临床使用设计噬菌体鸡尾酒.
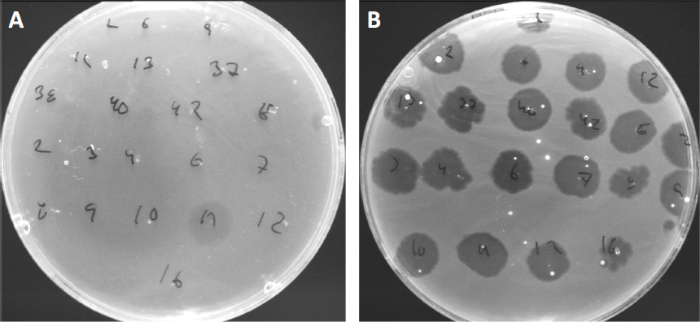
图4:不同细菌菌株的噬菌体敏感性。软琼脂板与可爱痤疮菌株(A)AD27和(B)AD35,被发现与21种不同的C.痤疮噬菌体。只有噬菌体11能够感染和杀死AD27,而AD35菌株对所有噬菌体表现出敏感性。这种技术,称为噬菌体类型,可用于根据噬菌体易感性将细菌种类和菌株分成不同的亚群。
Procedure
1. 设置
- 在开始任何涉及微生物的工作之前,请确保工作空间已消毒(例如,用 70% 乙醇擦拭)。始终穿着实验室外套和手套,保持长发绑在背上,并确保任何伤口都特别好保护。
- 完成后,对所有表面进行消毒,彻底清洗/消毒手和手腕。
2. 议定书
- LB 介质准备
注意:根据宿主细菌菌株和噬菌体的不同,不同的液体介质可能更适合于宿主细菌菌株的初始培养,或者不同的固体介质可能更适合后续生长。莱索根汤 (LB) 用于此协议中的肉汤和琼脂。- 混合 4 g LB 在 200 mL 蒸馏水中,在三重,为 LB 肉汤, 固体底部琼脂, 和软顶部琼脂.所有解决方案都应在容器中制备,其容量应为最终体积的两倍,以防止高压灭菌时溢出。
注意:如果以三重分法执行测定,则准备将 LB 底部琼脂的量翻倍。 - 根据需要,使用 NaOH 或 HCl 将所有三种解决方案的 pH 量调整为 7.4。
- 在底部琼脂瓶中加入 3 g 的琼脂能量,为固体琼脂制作 1.5% 的琼脂溶液
- 在顶部琼脂瓶中加入 1.2 g 的琼脂能量,使软琼脂的琼脂溶液为 0.6%。
- 将装有半紧固盖的瓶子放入设定为 121°C 的高压灭菌器中 20 分钟,对溶液进行消毒。运行完成后立即关闭盖子,以防止污染。
- 当 LB 介质温度达到约 45-50°C 时,在 200 mL LB 溶液中加入 450 μL 无菌 1 M CaCl2,最终浓度为 2.25 mM。
- 将固体琼脂介质的15 mL等分倒入无菌培养皿中(避免晃动以防止发泡),让琼脂在室温下凝固数小时或过夜(图4A)。板材可在4°C上下倒存储数天。
- 混合 4 g LB 在 200 mL 蒸馏水中,在三重,为 LB 肉汤, 固体底部琼脂, 和软顶部琼脂.所有解决方案都应在容器中制备,其容量应为最终体积的两倍,以防止高压灭菌时溢出。
- 宿主细胞的培养
- 在测定前一天,将10μL的大肠杆菌培养剂加入10 mL的LB汤中。
- 在摇动的培养箱中,在37°C下以160rpm在160rpm下孵育细菌。
- 在测定的早晨,将0.5 mL的过夜培养添加到10 mL的新鲜LB中。
注意:如果要在三次测定中执行测定,请准备此区域性的两倍。 - 在摇动的培养箱中以160rpm在37°C下孵育,直到细菌培养处于对数阶段生长。这可以通过 0.5-0.7 的 OD600 分光度来确定。
- 将培养物保持在室温下,直到细菌被添加到顶部 LB 琼脂。
- 噬菌体10倍的连续稀释
- 将 180μL 的 LB 肉汤添加到 a 第一行的 7 口井中,以 96 孔板
注意:建议对三联产品进行稀释,以提高其统计可靠性。为此,在板的第二行和第三行准备噬菌体的额外稀释剂。 - 小心地涡涡原噬菌体样品,以确保均匀性,并将20μL转移到第一口井中。
- 通过移液和向下混合样品。
- 将产生的悬浮液的 20 μL 转移到第二口井中。
- 继续连续稀释,将第二口井中的20μL溶液转移到第三口井,等等,直到第六口井,留下第七口和负对照,不会添加噬菌体悬浮液。这将创建 10-1 -10-6的稀释范围。
- 将 180μL 的 LB 肉汤添加到 a 第一行的 7 口井中,以 96 孔板
- 电镀
- 用名称、日期和简短的样品描述(包括噬菌体样品稀释因子)标记培养皿的底座(以前在步骤 2.1.7 中准备)。在测定前一小时在37°C培养箱中预热培养皿。
- 融化凝固的软琼脂介质(通常使用微波进行;凝固的琼脂在85°C下熔化),并让它接近45°C。加热介质可置于 45°C 水浴中,约 1 小时,达到适当的温度。它应该足够热,以保持液体形式,但足够凉爽,不杀死添加的细菌。
- 将 4 mL 细菌培养物(从步骤 2.2 起)与 35 mL LB 软琼脂混合(在 45°C 下)。旋转均匀分布细胞,但避免晃动,以防止发泡 (图4A)。
- 为每个连续稀释步骤标记一个无菌试管,为控制样品标记一个无菌试管,总共标记 7 个试管。将细菌培养剂/软琼脂混合物的5 mL等分从步骤2.4.3放入7管中。通过步骤 2.4.6 快速工作,因为这种小体积的基于琼脂的介质将在室温下迅速凝固。
- 将 100 μL 的控制样本(从步骤 2.3)添加到控制试管中,并小心旋转。丢弃用过的移液器尖端,并将相同的体积从每个序列稀释的噬菌体样品(步骤 2.3)传输到各自的试管,旋转混合。
- 立即将 5 mL 混合物转移到贴有标签的预热固体琼脂板上(图4A)。轻轻旋转板,甚至分散混合物。
注意:如果要在三重中执行测定,请重复步骤 2.4.3-6 两次。 - 用实验室薄膜密封每个板,并在室温下(约15分钟)使两层正确凝固,然后再将其从37°C培养箱上侧放置,刺激细菌和噬菌体的生长,持续24小时或直到斑块发育,它通常需要大约1-5天的斑块出现(图4B),但时间差异很大,取决于孵育条件,培养和细菌物种。
3. 数据分析和结果
-
计数斑块
- 确保标有"控制"的板中看不到斑块,因为这将表示病毒污染。
- 从标有10-6的板开始,含有最稀释的噬菌体样本。在不取下盖子的情况下对斑块进行计数,在进行标记时用笔标记它们,以指示哪些斑块已计数。
- 计算剩余的盘子。有些盘子的斑块可能太少或太多,无法计数。使用带有 10-150 块板的板进行进一步分析。
-
计算 PFU
- 将斑块数量除以稀释系数(例如,最稀释样品的10-6),以获得100μL噬菌体混合物中的斑块形成单位(PFU)数量。
注意:如果以三联形式执行测定,则使用三个板的平均斑块数。 - 要确定原始样品的浓度(以PFU/mL为单位),再乘以10的额外稀释系数,因为只镀了100μL的样品。(
 即)
即) - 计算具有 10 到 150 块斑块的所有稀释液的 PFU/mL 的平均值,以获得更可靠的结果。
- 将斑块数量除以稀释系数(例如,最稀释样品的10-6),以获得100μL噬菌体混合物中的斑块形成单位(PFU)数量。
 即)
即)噬菌体,也称为噬菌体,是专门感染细菌的病毒,我们可以确认它们的存在,并使用一种称为斑块测定的工具对其进行量化。噬菌体首先附着在细菌细胞壁上并注射其遗传物质,从而感染其易感宿主。然后,他们劫持细胞的生物合成机械来复制他们的DNA并产生许多后代噬菌体颗粒,然后通过解说和杀死宿主细胞释放这些粒子。
这种溶性活性是广泛使用的噬菌体枚举技术的基础,称为斑块测定或双琼脂层测定。在这里,噬菌体混合物首先在含有低浓度琼脂的熔融营养汤中制备。混合物中使用的所有细菌都应在生长的日志阶段保持活力并积极分裂,这将确保大部分细菌是活的,并能够在斑块周围形成密集的草坪。接下来,这种熔融细菌-噬菌体琼脂混合物分布在更固体,浓缩的琼脂营养介质,已经凝固在培养皿。在室温下孵育时,低浓度的琼脂-噬菌体汤也会凝固成柔软的琼脂覆盖物。
在这里,细菌细胞可以从底层获得额外的营养,并应迅速繁殖,产生细菌的汇流草坪。然而,由于噬菌体颗粒也存在于软层中,这些颗粒会感染和复制细菌内的遗传物质,最终导致细胞变热,从而释放多个后代。这些噬菌体后代然后感染邻近的细胞,因为细菌-噬菌层的半固态限制其运动到距离较远的宿主细胞。这种感染和莱沙循环持续多轮,在局部地区杀死大量细菌。邻近细胞被破坏的效果是产生一个圆形的透明区域,称为斑块,肉眼可以看到,有效地放大噬菌体的细菌溶酶活性,并使其枚举。
培养皿上的斑块数量称为斑块形成单位,或PFUs,如果初始噬菌体浓度足够稀释,应直接对应于原始样品中感染性噬菌体颗粒的数量。该技术还可用于表斑块形态的表征,帮助识别噬菌体类型,或分离噬菌体突变体。在本实验中,您将学习如何以大肠杆菌的T7噬菌体为例,为枚举噬菌体执行斑块测定。
首先,确定适合培养宿主细菌细胞和噬菌体的介质。在这里,乳原肉,或LB培养,用于培养大肠杆菌和T7噬菌体。接下来,取三个干净的玻璃瓶,并标记他们与媒体,名称,然后第一个作为LB-Broth,第二个作为LB-底部阿加,第三作为LB-Top阿加。现在,在三组中称量四克预配方的 LB 粉末,然后将一套称重干燥介质转移到每个瓶子中。将200毫升水加入第一瓶。使用磁性搅拌棒混合内容物。
然后,使用pH计和不断搅拌,通过添加氢氧化钠或盐酸,使最终pH达到7.4。对其余两个瓶子也重复加水和pH调整。现在,称量三克的琼脂粉,并添加到第二瓶,使一个1.5%的底部琼脂。最后,称量1。2 克琼脂,并添加到第三瓶,使 0.6% LB 顶部琼脂。瓶中的汤条件不需要添加琼脂。将瓶子半紧封盖,然后通过在121摄氏度的高压灭菌器对介质进行消毒20分钟。完成后,从高压灭菌器中取出介质瓶,并立即拧紧瓶盖以将其完全关闭,以防止污染。将 LB-Broth 和 LB-Top Agar 介质放在长凳上,以便日后使用。将 LB-底部 Agar 放在预设约 45 摄氏度的水浴中冷却。
当 LB-底部琼脂达到约 45 摄氏度时,将其转移到工作台上。接下来,使用 70 % 异醇对工作区进行消毒。接下来,在熔融底部琼脂中加入450微升无菌一摩尔氯化钙,最终浓度为2.25毫摩尔。轻轻旋转瓶子混合。然后,设置七个干净的培养皿。将每道菜的底部贴上介质名称和准备日期的标签。然后,将 15 毫升的底部琼脂倒入七个培养皿中。让板在室温下设置几个小时或过夜。一旦设置,培养板可以存储在四摄氏度,如果需要几天,倒置,以尽量减少冷凝。在测定前一小时将培养皿从4摄氏度的冰箱转移到37摄氏度的培养箱。
在预成型前一天,应培养大肠杆菌。在这里,10微升大肠杆菌培养被接种成10毫升的LB-Broth。将细菌放在160RPM下设置为37摄氏度的摇动培养箱中过夜。然后,在测定当天,从培养箱中取出细菌培养物。播种新鲜10毫升的新鲜LB汤与0.5毫升的过夜文化。将这些细胞置于160RPM下,生长成37摄氏度的摇动培养箱。接下来,使用分光光度计检查此培养体何时达到对数相增长,光学密度为 0.5 到 0.7。一旦OD达到这个水平,通过将细胞培养转移到工作台来停止孵育。它们现在可用于噬菌体覆盖测定。
噬菌体在不同噬菌体类型和样品之间可能呈指数变化。因此,为了有效地计数它们,它们应该被稀释,以产生广泛的噬菌体浓度。在测定当天,采用10倍稀释技术,产生一系列从十分之一到百万分之一浓度不等的噬菌体稀释剂。为了获得统计显著性和准确性的数据,在三元化中执行连续稀释。
接下来,使用微波熔化凝固的 LB-top 琼脂。然后,将其放入预设在 45 摄氏度的水浴中一小时。一小时后,从孵化器收集含有底部琼脂层的培养皿。用噬菌体浓度和测定日期标记盘子。然后,设置七个干净的试管。用串行噬菌体稀释编号标记每个试管,并指定一个作为控制。
当 LB 顶琼脂达到 45 摄氏度时,将其转移到工作台。现在,在200毫升的琼脂中加入450微升的一个摩尔氯化钙,最终浓度为2.25毫摩尔。轻轻旋转瓶子混合。接下来,在无菌锥形管中加入35毫升的LB顶琼脂和4毫升的细菌悬浮液。轻轻旋转以均匀分布细胞,但避免晃动,以防止发泡。
现在,将五毫升的这种细菌顶琼脂混合到七个试管中。然后,将100微升的连续稀释噬菌体样品和控制介质(应为无噬菌体)转移到贴有尊重标签的试管中。轻轻旋转混合物,以确保正确混合。轻轻地将五毫升噬菌体混合物转移到各自的Petri板上。通过轻轻旋转 Petri 板,均匀地将混合物均匀地铺过整个表面。
一旦所有 Petri 板都与混合层分层,通过在室温下孵育 15 分钟,使顶层凝固。完成这些步骤后,使用剩余的两组噬菌体稀释法重复第二套和第三套培养皿的过程。用副膜密封每道菜,并在室温下孵育15分钟。将培养板倒置在适当的温度下 24 小时或直到斑块形成。在这里,板被放置在37摄氏度的培养箱一天,一个刺激生长条件的大肠杆菌和T7噬菌体。
根据细菌种类、孵育条件和介质的选择,在孵育一至五天后会出现斑块。在这里,在37摄氏度的孵育一天后,斑块就可见了。首先检查标有控制的板,并确保这些板块中未形成斑块,因为这将表示病毒污染。要确定原始样品中的噬菌体,请首先从包含最稀释的噬菌体样本的板开始,在不取下盖子的情况下对斑块进行计数,并标记它们以指示哪些已经计数。对每组每个板重复计数。有些盘子的斑块可能太多或太少,无法计数。将 10 到 150 视为理想的斑块计数。
接下来,生成一个表,列出不同稀释和复制的斑块数值。然后,计算包含理想斑块计数数的稀释板的平均斑块数值。在此示例中,这些是 10 到负 3 和 10 到负 4 稀释板中形成的斑块的平均数量。现在,通过将获得的均值斑块值除以相应的噬菌体稀释因子来调整噬菌体稀释系数。在这里,形成到10到负3和10到负4稀释板的平均斑块数,被它们各自的稀释因子除以,以获得100微升噬菌体混合物中的斑块形成单位(PFUs)的数量。要将该值转换为每毫升的 PFU,请将生成的值乘以 10,因为噬菌体覆盖制备步骤中仅使用了 100 微升的噬菌体稀释混合,从而产生 10 的额外稀释系数。最后,计算从不同稀释板中获得的值的平均值。这将给出每毫升 PFUS 的平均数量。PFUs 的数量对应于原始样本中感染性噬菌体颗粒的数量。
Subscription Required. Please recommend JoVE to your librarian.
Applications and Summary
尽管多项技术进步,斑块测定仍然是测定病毒性牙酸盐(作为PFU)的黄金标准,对纯噬菌体种群的分离至关重要。易感宿主细胞在两层琼脂板的上部涂层中培养,形成均匀的床,实现病毒复制。在溶质生命周期中分离的噬菌体感染细胞,在细胞内复制并最终将其解伤的初始事件太小,无法观察。然而,释放的病毒会感染相邻的细胞,随后产生一个清除区,或斑块,表示存在单个PFU。
Subscription Required. Please recommend JoVE to your librarian.
References
- Twort, F. An investigation on the nature of ultra-microscopic viruses. Lancet. 186 (4814): 1241-1243. (1915)
- d'Hérelle, F. An invisible antagonist microbe of dysentery bacillus. Comptes Rendus Hebdomadaires Des Seances De L Academie Des Sciences. 165: 373-375. (1917)
- Cisek AA, Dąbrowska I, Gregorczyk KP, Wyżewski Z. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages. Current Microbiology. 74 (2):277-283. (2017)
- Mirzaei MK, Maurice CF. Ménage à trois in the human gut: interactions between host, bacteria and phages. Nature Reviews Microbiology. 15 (7):397. (2017)
- Breitbart M, Bonnain C, Malki K, Sawaya NA. Phage puppet masters of the marine microbial realm. Nature Microbiology. 3 (7):754-766. (2018)
- Leung CY, Weitz JS. Modeling the synergistic elimination of bacteria by phage and the innate immune system. Journal of Theoretical Biology. 429:241-252. (2017)
- Dulbecco R. Production of Plaques in Monolayer Tissue Cultures by Single Particles of an Animal Virus. Proceedings of the National Academy of Sciences of the United States of America. 38 (8):747-752. (1952)
- Juarez D, Long KC, Aguilar P, Kochel TJ, Halsey ES. Assessment of plaque assay methods for alphaviruses. J Virol Methods. 187 (1):185-9. (2013)
- Clokie MRJ, Millard AD, Letarov AV, Heaphy S. 2011. Phages in nature. Bacteriophage. 1 (1):31-45. (2011)
- Moldovan R, Chapman-McQuiston E, Wu XL. On kinetics of phage adsorption. Biophys J. 93 (1):303-15. (2007)
- Garneau JE, Dupuis M-È, Villion M, Romero DA, Barrangou R, Boyaval P, Fremaux C, Horvath P, Magadán AH, Moineau S.. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 468 (7320):67. (2010)