



Overview
资料来源:亚历山大·金1,托尼娅·科尔皮茨1
1波士顿大学医学院微生物学系,国家新兴感染疾病实验室,波士顿,马萨诸塞州
1946年,莱德伯格和塔图姆首次发现,结合是细菌之间的一种水平基因转移形式,依赖于两个细菌细胞之间的直接物理接触(1)。与其他形式的基因转移(如转化或转导)不同,结合是一个自然发生的过程,DNA以单向的方式从供体细胞分泌到受体细胞。这种方向性和这一过程增加细菌遗传多样性的能力,使结合作为细菌"交配"的一种形式的声誉,这被认为极大地促进了最近抗生素耐药性的上升细菌(2,3)。通过使用选择性压力,例如使用抗生素,结合纵用于实验室设置,使其成为细菌之间水平基因转移的有力工具,在某些情况下,从细菌到酵母、植物和动物单元格 (4)。除了在实验室中的应用,通过结合转移真核酸细菌基因是DNA转移的一个令人兴奋的途径,具有多种可能的生物技术应用和自然发生的影响(5)。
结合被认为是通过"两步机制"(6)工作。首先,在转移任何DNA之前,供体细胞必须与受体进行细胞对细胞的直接接触。这个过程在革兰氏阴性细菌中具有最佳特征,其中研究最多的是大肠杆菌。细胞与细胞接触是通过在捐赠者身上存在一个复杂的细胞外细丝网络而建立的,这种结合因子由称为F(生育)因子(7,8)的可转移基因编码。除了在供体和受体之间建立接触外,几种蛋白质通过性皮球输送到受体细胞质,在两个细胞之间形成IV型分泌系统(T4SS)导管,这是第二步的必要结构。共和,DNA转移(6)。通过将性皮鲁斯的这种功能与DNA的滚动循环复制相结合,供体细胞能够通过"芽和泵"模型(6)将DNA以可转位元素的形式(如质粒或转位子)转移给受体。在这种情况下,"射击"是通过T4SS将具有链接DNA的先导蛋白输送到受体细胞,而"泵送"是DNA向受体的主动传输,这个过程依赖于T4SS,并通过耦合蛋白(6)催化。这个过程中使用的机械由转移序列(oriT)的起源组成,必须由CIS和转基因中的DNA提供,该DNA编码松弛酶、配偶配对形成复合体和IV型耦合蛋白,以及可以存在于cis或转(9) 中。这种松弛酶在oriT序列中分裂尼克位点,并共价附着在转移链的5'端,以产生松弛体,一种单链DNA-松弛酶复合物与其他辅助蛋白(9)。一旦形成,松弛体通过IV型耦合蛋白连接到交配对形成复合体,它允许SsDNA-松弛酶复合物通过T4SS(10)转移到受体细胞中。一旦进入受体的细胞质,DNA可以集成到受体基因组中,或者以质粒的形式单独存在,其中任何一种都允许其基因的表达。
在本实验中,使用广泛使用的偶联供体菌株大肠杆菌WM3064将对受体大肠杆菌J53的抗中青霉素的基因编码转移。 虽然革兰氏阴性细菌的两种菌株对四环素具有抗药性,但只有供体菌株WM3064具有氨基青霉素抗性基因,编码为pWD2-oriT穿梭载体中,对二氨基甲酸(DAP)(DAP)具有辅助营养性。该实验包括两个主要步骤,一是制备供体和受体菌株,然后是通过结合将安培林抗性基因从供体转移到受体(图1)。
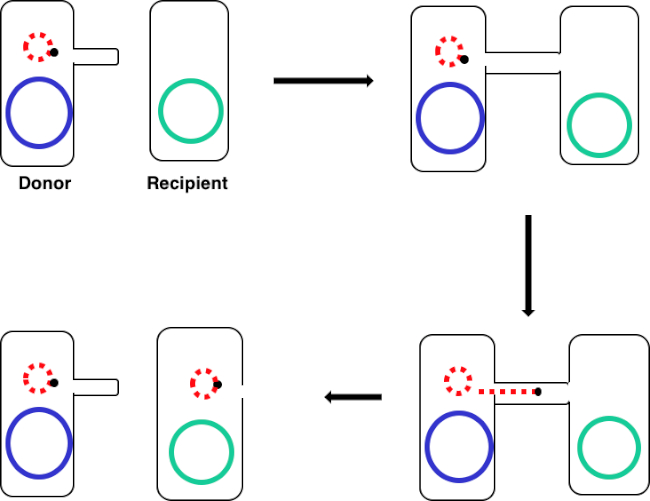
图1:共通示意图。此原理图显示了质粒(仅可转位DNA元素的一个示例)从供体细胞使用结合从受体细胞成功转移。当供体细胞通过性皮鲁斯与受体细胞接触时,质粒通过滚动循环复制复制,通过连接两个细胞的多蛋白复合体移动,并在受体细胞中形成新的全长质粒。
通过孵育供体和受体细胞的混合物,然后在四环素和DAP的存在下连续电镀这些细胞,从而成功转移了阿霉素抗性基因。在四环素和安平存在的情况下,从这种混合物中生长的连续电镀细胞,由于缺少DAP和可能尚未获得环青素抗基因的任何受体细胞,去除了所有供体细胞,产生严格受体J53菌株获得对青霉素的细菌(图2)。一旦进行,通过PCR证实了青霉素抗性基因的成功转移。由于结合成功,大肠杆菌的J53菌株含有pWD2-oriT,对阿霉素具有抗药性,PCR可检测到这种抗性的基因编码。然而,如果不成功,将没有检测到阿霉素耐药性基因,而阿霉素仍将作为对抗J53菌株的有效抗生素。
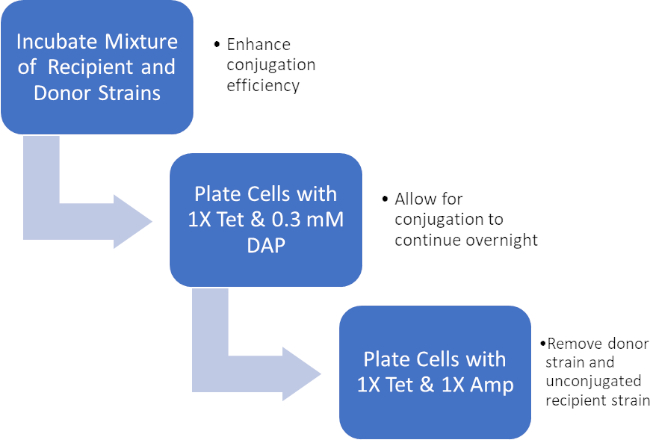
图2:协议示意图。此原理图显示了所呈现协议的概述。
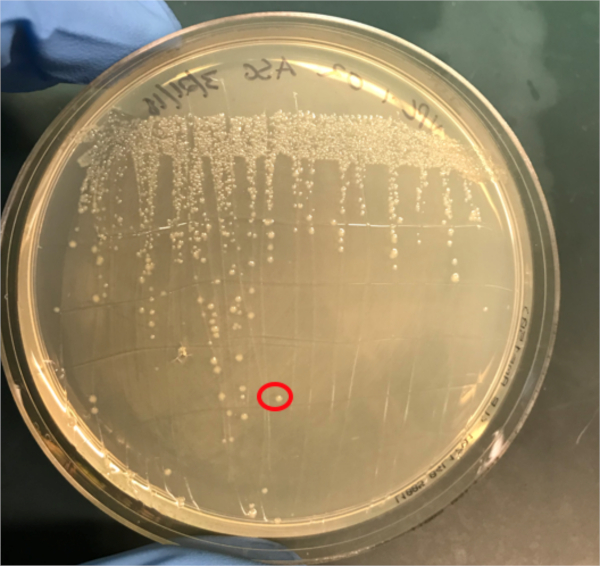
图3A:PCR成功结合的确认。A) 在琼脂板上条纹的偶联和阴性对照样本的冷冻库,并选择一个菌群(红色)进行DNA分离。
Procedure
1. 设置
- 高压灭菌器约 1L 卢里亚-贝尔塔尼介质 (LB)。这种无菌 LB 将用于制造约 5 mL 的 LB,其中包含 0.3 mM 二氨基酸 (DAP)。
- 收集以下板:LB 琼脂板,1X Tet 和 0.3 mM DAP,仅 1X Tet 的 LB 琼脂板,仅 1X 置时/特的 LB 琼脂板。
- 确保一些甘油和一盒预消毒的塑料移液器吸头在手附近。
- 在开始任何涉及微生物的工作之前,使用 70% 乙醇对工作空间进行消毒。务必佩戴必要的个人防护装备,包括实验室外套和手套。
- 完成后,用70%乙醇消毒所有表面和手套,然后洗手。
2. 供体和受助人应变准备
- 准备5 mL细菌培养物的供体和受体菌株,并在37°C过夜,在220rpm下曝气和摇动。供体菌株应在 LB 中生长,具有 0.3 mM DAP。
- 将两种培养物的 1 mL 旋转(±3000 rpm 5 分钟),并使用 PBS 清洗细胞。
- 在500μL的磷酸盐缓冲盐水(PBS)中重新悬浮细胞。
3. 合并
- 将50μL的受体细胞与50μL的供体细胞结合在微离心管中,并通过移液混合。
- 在37°C下孵育细胞混合物1小时。这将提高在下一个电镀步骤之前和期间的联结效率。
- 将100 μL的细胞混合物移至琼脂板上,带1X Tet和0.3 mM DAP。不要铺在盘子上。
- 将受体细胞培养的移液100 μL移至琼脂板上,具有1X Tet和0.3 mM DAP。不要铺在盘子上。
- 在37°C下孵育两个板过夜。
- 用无菌细胞刮刀刮起结合细胞混合物和受体细胞培养。将细胞转移到无菌微离心管中,并将细胞重新悬浮在1 mL的PBS中。
- 涡旋细胞,轻轻旋转它们(约3000rpm5分钟)。
- 在 PBS 的 1 mL 中重新悬浮细胞。
- 将结合反应细胞混合物板在LB琼脂板上,仅带1倍安培/Tet。在这个板块上,只有通过结合成功获得Amp抗性基因的受体细菌才能生长。
- 将受体细胞板板到仅带 1X Tet 的 LB 琼脂板上。在这个板块上,只有没有Amp抗性基因的未结合受体细菌才会生长。
- 在37°C下孵育板。
- 从两个板中挑选单个菌落,并在 5 mL 的介质(37 °C,220 rpm 转速下曝气)中使用它们在一夜之间培养。
4. DNA分离
- 通过DNA最小制备,使用总培养体积的4.5 mL从先前制备的培养物中分离DNA。
- 为此,使用35 μL的无核酸酶水洗脱DNA。
- 纯DNA将产生约1.8的吸收比(A260/280)。
- 使用每种培养物的剩余 0.5 mL,通过制造细菌培养物和 100% 甘油的 1:1 混合物来制备甘油库存。
- 将冷冻库存储存在-80°C。
5. PCR确认质粒转移结合
- 准备两个PCR主混合物,每个混合物与一组不同的正向和反向引物,一个针对在阿霉素抗基因内的500基对段,另一个针对内务管理基因中的一段。
- 家政基因引物被设计为在DNA陀螺仪B(14)的细菌基因编码中扩增一段DNA。
- 以下试剂体积用于制备每个主混合物的 90 μL:
7.5 μL 的 10 μM 前引底
7.5 μL 的 10 μM 反向底漆
75 μL 2X PCR 主混合
- 使用15μL主混合物、10 ng模板DNA和无核酸酶水制备以下6种PCR反应,最终体积为25μL。
结合反应DNA和安西林引基
结合反应DNA和家政引体
阴性对照DNA和安西林引基
负控制DNA和家政引种
无DNA和安西林引种
无DNA和内务引种 - 将这些反应转移到预加热至 98 C 的 PCR 机器,并在以下条件下开始热循环:
98 °C 30 秒
98 °C 的 25-35 次循环,5-10 秒,45-72 °C 10 至 30 秒,72°C,每 kb 15-30 秒
72 °C 5-10 分钟
保持在 4°C - 将所有六种PCR反应加载到1%的甘蔗凝胶中,并在±150V下运行约20分钟。
- 使用紫外线照明器可视化 PC 产品。
细菌细胞,如大肠杆菌,能够将遗传信息从细胞转移到细胞。结合不同于DNA转移的其他机制,如转导或转化,因为它需要细胞之间的物理接触。
要继续,结合需要一个表示生育力的供体细胞,或F,因子,没有它的受体细胞,F减细胞。该过程需要两个步骤。第一种是建立直接的细胞对细胞接触。为此,供体细胞产生一种称为性皮毛的细胞外丝状结构。它之所以被命名为这个,因为结合是无性繁殖细菌的交配形式,但应该注意的是,它不是真正的性繁殖,因为没有交换配子,也没有后代形成。
第二步是将DNA输送到受体细胞。性皮鲁斯在两个细胞之间建立接触后,建立了一个称为IV型分泌系统的管道,允许DNA的转移。供体细胞然后开始复制将基于遗传元素(称为OriT或转移源)的遗传元素进行转移的染色体外DNA。新复制的DNA的一端通过DNA蛋白质结合线入导管。随着DNA的进一步复制,它通过通道泵送,由靠近OriT的基因编码的复合蛋白促进。一旦DNA被完全转移,它将形成一个额外的染色体质粒,或者它可能集成到受体细胞的染色体。无论转移的DNA的终点,它编码的基因都会被表达。这种基因表达可用于确认成功的结合。
例如,假设供体菌株表达环青素耐药性,并在结合的DNA中将其传递给受体细菌,但受体菌株也有四环素抗性基因不存在于供体中。在这种情况下,当细胞在含有四环素和氨基青霉素的LB介质上镀时,菌落应只从成功的结合细菌中生长,这将表达两种抗性表型。为了进一步确认成功的结合,可以采集这些菌落的质粒DNA,然后利用聚合酶链反应或PCR放大与转移质粒特有的一段DNA。当 PCR 产品在电泳凝胶上运行,同时与标准尺寸的阶梯一起运行时,已知尺寸的 PCR 片段应在凝胶上可见,从而进一步确认成功结合。在这个实验中,将用质粒将一个质粒通过结合将环青素抗性基因从供体菌株转移到四环素抗受体菌株。在此之后,为了确认结合,结合混合物将孵育在含有两种抗生素的板上,只留下转化的细菌。最后,使用PCR进一步确认成功的结合。
在开始手术之前,请穿上适当的个人防护装备,包括实验室外套和手套。接下来,使用 70% 乙醇对工作空间进行消毒,以擦拭表面。
在此过程中,阿霉素抗性基因将通过联结从大肠杆菌的WM3064菌株转移到大肠杆菌的J53菌株。供体菌株WM3064对四环素和氨基青霉素具有抗药性,它需要二氨基甲酸(DAP)生长。受体应变 J53 仅对四环素具有抗性,并且不需要 DAP 生长。这意味着成功的结合细胞应耐四环素和环氧西林,并且无需DAP即可生长。
通过接种含有0.3毫摩尔DAP的5毫升LB,用冷冻的供体菌株甘油库存的废料来制备供体菌株培养。然后,通过接种五毫升的LB汤而不用DAP与冷冻受体菌株甘油库存的废料来准备受体菌株。在37摄氏度的温度下生长,在摇动的培养箱中以220RPM的转速进行曝气和摇动。一旦培养物发展到两个的OD 600,从每个培养物中取出一毫升的培养物,然后放入两个新的单独的1.5毫升微离心管中。然后,在3000RPM下将这些等分物离心5分钟,以颗粒细菌细胞。丢弃上清液,用 250 微升 1X PBS 洗净每个颗粒。再次将样品离心,在丢弃上清液后,将每粒颗粒重新悬浮在 500 微升 PBS 中。
为了开始结合过程,首先将50微升的受体细胞与50微升的供体细胞结合在1.5毫升微离心管中,然后通过轻轻上下移液混合。接下来,移液器 100 微升的受体细胞培养液到另一个包含 DAP 的 1X 四环素板上。接下来,通过将受体细胞培养的 100 微升移液至包含 DAP 的非选择性琼脂板上,准备负控制。然后,在37摄氏度的温度下孵育共和负控制板过夜。
第二天,采取无菌细胞刮刀,并通过收集菌落从结合板收获细胞。然后,将菌落转移到含有一毫升1X PBS的无菌1.5毫升微离心管中。重复此过程以从另一个板收集接收细胞。
之后,涡旋样品混合。混合后,将管子转移到离心机,轻轻颗粒细胞。丢弃上清液,然后在一毫升的PBS中清洗细胞颗粒,并涡旋管以重新悬浮细胞。通过离心再次将细胞压过。再次丢弃上清液,并将两个细胞颗粒重新悬浮在一毫升的PBS中。现在,使用无菌移液器尖端,将100微升的偶联反应细胞混合物板到不含1X四环素和1X环二环素的LB琼脂板上。重复电镀方法,使用PBS中同一细胞混合物的100微升稀释到另一个LB琼脂板上,不含1X四环素和1X环素。
最后,将负控制细胞混合物的移液器100微升放在单个LB琼胶板上,仅带1X四环素。在摄氏37度过夜孵育后,群落应可见。使用无菌移液器尖端,从结合反应板中挑选单个菌落,并将其添加到含有五毫升选择性LB培养素的管中,其中含有两种抗生素。然后,通过从受体细胞板中选择单个菌落来重复菌落隔离。在 37 摄氏度下一夜之间生长这些培养物,在 220 RPM 下进行曝气。
第二天,用70%乙醇擦拭台面,从培养箱中取出盘子。使用DNA迷你准备试剂盒从4分离DNA。根据制造商的说明,每种培养的5毫升。完成DNA迷你制备后,使用35微升无核酸酶水洗脱DNA。最后,使用剩余的 0。5毫升每种培养物,通过添加0.5毫升100%甘油来制备一毫升甘油,进行一对一稀释。将这些等分置于零下 80 摄氏度,以便存放,直到需要为止。
为了确认PCR成功结合,首先通过在微离心管中加入75微升的2XPCR主混合物来制备PCR主混合物。然后,加入7.5微升各10微摩尔前引底和10微摩尔反向引基,旨在放大质粒中的环青素抗性基因。接下来,通过将 75 微升的 2X PCR 主混合物添加到微离心管中,然后加入 10 微摩尔前引物和 10 微摩尔反向引物,用于扩增内务基因,从而分别加入 7.5 微升,从而制备第二个 PCR 主混合物,在此例中为 DNA陀螺B。
现在,将15微升的第一个主混合物加入PCR管中,然后加入10毫微克,大约两微升的模板实验DNA到同一管中。使用无核酸酶水将反应调至25微升的最终体积。重复这些步骤以产生其余五个反应,以便管包含此处所示的组件。现在,将这些反应转移到热循环器与块预热到98摄氏度,然后启动程序。PCR 完成后,从机器上拆下管子。然后,将每个反应的两微升与两个微升的加载染料和四微升的分子量标记物混合到1%的甘蔗凝胶的连续井中。将凝胶设置为在 150 伏电压下运行 20 分钟。最后,使用紫外线照明器可视化凝胶。
在实验中,通过PCR证实了阿霉素抗性基因通过结合的成功转移。在这里,在含有结合DNA和安培霉素引种的井中应观察到大约500个碱基对大小的带,在本例中,井中应观察两个。内务管理基因,DNA陀螺B,分别被加载到井三和五与结合的DNA和受体细胞DNA。在这些井中观察到的带子作为正对照,确保DNA模板存在,PCR成功。在包含受体细胞DNA和环霉素引种对反应的孔中不应观察到带,本例中为4,因为受体细胞不耐环西林。此外,在缺乏模板DNA的反应中不应观察到带,此处为6和7井。如果满足这些条件,这将确认环霉素抗性基因的成功转移,使环酸素从WM3064大肠杆菌菌株转移到大肠杆菌的J53菌株。
Subscription Required. Please recommend JoVE to your librarian.
Results
如果结合成功,将在装载PCR反应1的井中观察到500基对大小的带PCR产物(图3B中#2井),而在装载PCR反应3的孔中不会观察到带子(图3B中#4)。该带的存在证实了环青素抗性基因的成功转移,从而赋予对大肠杆菌J53菌株的环酸素耐药性。

图3B:PCR成功结合的确认。B) PCR分析使用从选定菌群分离的DNA进行。每口井的内容如下:1)DNA阶梯,2)结合DNA和安西林引物,3)结合DNA和内务引物,4)阴性对照DNA和安平素引物,5)负控制DNA和内务引物,6)无DNA和ampicillin引种,和7)无DNA和阴性对照引种。来自PCR反应1(井2)的+500基对带PCR产物的存在,以及PCR反应3(井4)中缺乏本产品的存在,证实了成功的结合。
Subscription Required. Please recommend JoVE to your librarian.
Applications and Summary
结合是一个自然发生的水平基因转移过程,依赖于供体细胞和受体细胞的直接细胞对细胞接触。这个过程在各种细菌之间共享,并在细菌进化中发挥了作用,最显著的是抗生素耐药性。在实验室中,结合可用作基因转移的有效方法,与其他技术相比,其破坏性要小得多。在实验室之外,通过结合将DNA从细菌转移到真核生物的能力为基因治疗提供了一条令人兴奋的新途径,并理解了这些自然发生的基因转移的影响,例如细菌感染和癌症,是一个迅速兴起的研究领域。
Subscription Required. Please recommend JoVE to your librarian.
References
- Lederberg J, Tatum, E.L. Gene recombination in Escherichia coli Nature. 1946;158:558.
- Holmes R.K. J, M.G. Genetics: Exchange of Genetic Information. 4th Edition ed. Baron S, editor. Galveston, TX: University of Texas Medical Branch at Galveston; 1996.
- Cruz F, Davies, J. Horizontal gene transfer and the origin of species: lessons from bacteria. Trends in Microbiology. 2000;8:128-33.
- Llosa M, Cruz, F. Bacterial conjugation: a potential tool for genomic engineering. Ressearch in Microbiology. 2005;156:1-6.
- Lacroix B, Citovsky, V. Transfer of DNA from Bacteria to Eukaryotes. mBio. 2016;7(4):1-9.
- Llosa M, et al. Bacterial conjugation: a two-step mechanism for
- DNA transport. Molecular Microbiology. 2002;45:1-8.
- Grohmann E, Muth, G., Espinosa, M. Conjugative Plasmid Transfer in Gram-Positive Bacteria. Microbiology and Molecular Biology Reviews. 2003;67:277-301.
- Firth N, Ippen-Ihler, K, Skurray, RA. Structure and function of the F factor and mechanism of conjugation. Escherichia coli and salmonella: cellular and molecular biology. 1996;2:2377-401.
- Smillie C, Garcillan-Barcia MP, Francia MV, Rocha EPC, De La Cruz F. Mobility of Plasmids. Microbiology and Molecular Biology Reviews. 2010;74(3):434-52.
- Cascales E. Definition of a Bacterial Type IV Secretion Pathway for a DNA Substrate. 2004;304(5674):1170-3.
- Wang P, Yu Z, Li B, Cai X, Zeng Z, Chen X, et al. Development of an efficient conjugation-based genetic manipulation system for Pseudoalteromonas. Microbial Cell Factories. 2015;14(1):11.
- Yi H, Cho YJ, Yong D, Chun J. Genome Sequence of Escherichia coli J53, a Reference Strain for Genetic Studies. Journal of Bacteriology. 2012;194(14):3742-3.
- Baumann RLB, E. H.; Wiseman, J. S.; Vaal, M.; Nichols, J. S. Inhibition of Escherichia coli Growth and Diaminopimelic Acid Epimerase by 3-Chlorodiaminopimelic Acid. Antimicrobial Agents and Chemotherapy 1988;32:1119-23.
- Rocha D, Santos, CS, Pacheco LG. Bacterial reference genes for gene expression studies by RT-qPCR: survey and analysis. Antonie Van Leeuwenhoek. 2015;108:685-93.