



Overview
ソース: アレクサンダー S. ゴールド1, トーニャ M. コルピッツ1
1ボストン大学医学部微生物学科、国立新興感染症研究所、ボストン、マサチューセッツ州
1946年にレダーバーグとテイタムによって最初に発見された共役は、2つの細菌細胞間の直接的な物理的接触に依存する細菌間の水平遺伝子伝達の一形態である(1)。形質転換やトランスダクションなどの他の形態の遺伝子導入とは異なり、結合はDNAがドナー細胞からレシピエント細胞に一方向的に分泌される自然発生プロセスです。この方向性と細菌の遺伝的多様性を高めるこのプロセスの能力は、抗生物質耐性の最近の上昇に大きく貢献していると考えられている細菌「交配」の一形態としての評判を共役に与えました。バクテリア(2、3)。選択的圧力(例えば抗生物質の使用)を用いることで、実験室での使用のためにコンジュゲーションが操作され、細菌間の水平遺伝子移動、場合によっては細菌から酵母、植物、動物への強力なツールとなっています。セル (4)。実験室での応用とは別に、結合による細菌・ユーカリオテ遺伝子の伝達は、多くの可能なバイオテクノロジーアプリケーションと自然に起こる影響を持つDNA伝達のエキサイティングな手段です(5)。
コンジュゲーションは「2段階機構」(6)によって働くと考えられている。まず、DNAを移送する前に、ドナー細胞はレシピエントと直接細胞間接触を行わなければならない。このプロセスは、グラム陰性細菌で最もよく特徴付けられており、その中で最も研究されているのは大腸菌である。細胞間接触は、性ピラスとして知られているドナー上の細胞外フィラメントの複雑なネットワークの存在によって確立され、F(不妊)因子(7,8)として知られる転写可能な遺伝子によってコードされる共役元素である。ドナーとレシピエントとの接触を確立することに加えて、いくつかのタンパク質は、レシピエント細胞質に性ピラスを介して輸送され、2つの細胞間のIV分泌系(T4SS)導管を形成し、第2工程に必要な構造である。DNA伝達(6)。この性ピルスの機能とDNAの転がり円複製を組み合わせることにより、ドナー細胞は、プラスミドまたはトランスポゾンなどのトランスポーザブル要素の形でDNAを「シュートアンドポンプ」モデル(6)によってレシピエントに移すことができる。この場合、「シューティング」は、パイロットタンパク質の輸送であり、連結DNAを用いて、T4SSによってレシピエント細胞に送られ、「ポンピング」はレシピエントへのDNAの活性輸送であり、T4SSに依存し、結合タンパク質によって触媒されるプロセスである(6)。このプロセスで使用される機械は、リラクサーゼ、合体対形成複合体、およびIV型結合タンパク質をコードするシスおよびトランス遺伝子のDNAによって提供されなければならない転移配列(oriT)の起源から構成され、シスまたはトランス(9)に存在することができます。このリラクサーゼは、oriT配列内のnic部位を切断し、転移した鎖の5'末端に生有して、他の補助タンパク質との一本鎖DNAリラクサーゼ複合体であるリラクソームを産生する(9)。いったん形成されると、リラクソームは交配対形成複合体に接続し、IV型結合タンパク質を介して、SsDNA-relaxase複合体をT4SS(10)によってレシピエント細胞に移植することを可能にする。レシピエントの細胞質に入ると、DNAはレシピエントゲノムに統合することも、プラスミドの形で別々に存在することも、その遺伝子の発現を可能にする。
この実験では、広く使用されている共役ドナー株大腸菌WM3064を、レシピエント株大腸菌J53に対するアンピシリン耐性をコードする遺伝子コードを伝達するために用いられた。グラム陰性菌の両方の株はテトラサイクリンに耐性であったが、ドナー株WM3064のみがアンピシリン耐性の遺伝子を持ち、pWD2-oriTシャトルベクターでコードされ、ジアミノピメル酸(DAP)に対して下方栄養性であった(11-13)。この実験は、ドナーおよびレシピエント株の調製、続いてコンジュゲーションによるドナーからレシピエントへのアンピシリン耐性遺伝子の移植の2つの主要なステップからなる(図1)。
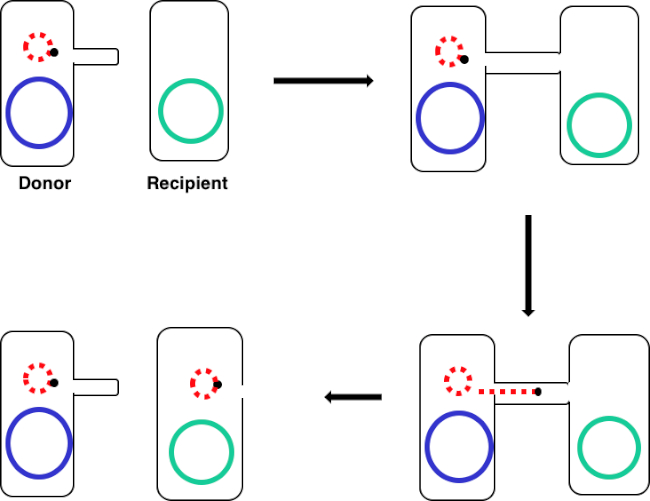
図 1: コンジュゲーション回路図。この回路図は、プラスミドの正常な転移を示し、転移可能なDNA要素の一例に過ぎず、共役を用いてドナー細胞からレシピエント細胞へ送り込む。性ピラスを介してドナー細胞によってレシピエント細胞と接触すると、プラスミドは転がり円複製によって複製し、2つの細胞を結合する多タンパク質複合体を通って移動し、レシピエント細胞に新しい全長プラスミドを形成する。
ドナー細胞とレシピエント細胞の混合物をインキュベートすることにより、テトラサイクリンおよびDAPの存在下でこれらの細胞を連続的にめっきし、これはアンピシリン耐性遺伝子の正常な転写を可能にした。テトラサイクリンとアンピシリンの存在下でこの混合物から増殖した細胞を連続的にめっきし、DAPの欠如とアンピシリン耐性遺伝子を得ていない可能性のある任意のレシピエント細胞の欠如のためにすべてのドナー細胞を除去し、厳密にレシピエントJ53株を得たアンピシリン耐性を獲得した細菌(図2)。一旦行われると、アンピシリン耐性遺伝子の正常な転写がPCRによって確認された。大腸菌のJ53株はpWD2-oriTを含み、アンピシリンに対して耐性があり、この耐性をコードする遺伝子はPCRによって検出可能であった。しかし、もし失敗した場合、アンピシリン耐性遺伝子の検出はなく、アンピシリンはJ53株に対する有効な抗生物質として依然として機能するだろう。
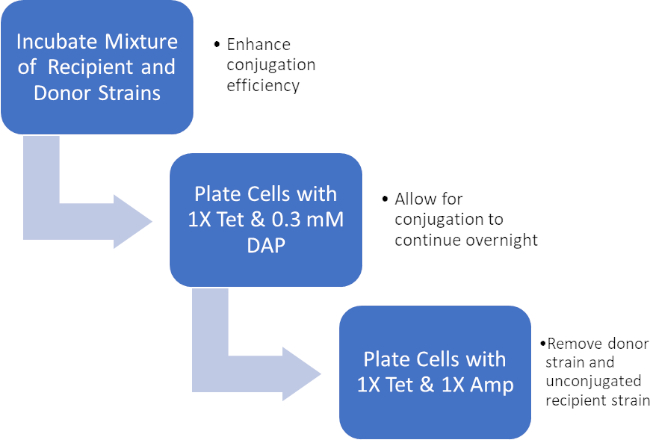
図 2: プロトコルの回路図。この回路図は、提示されたプロトコルの概要を示しています。
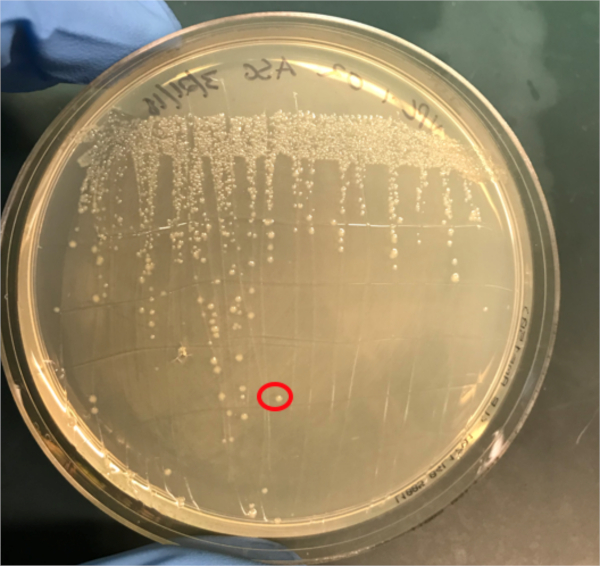
図3A:PCRによるコンジュゲーションの成功の確認A)共役および陰性対照試料の冷凍庫ストックを寒天プレート上にストリークアウトし、DNA分離のためにコロニー(赤色)を選択した。
Procedure
1. セットアップ
- ルリア・ベルタニ培地(LB)のオートクレーブ約1L。この無菌LBは、0.3mMジアミノピメル酸(DAP)を含むLBの約5mLを作るために使用されます。
- 次のプレートを収集する:1Xテトと0.3 mM DAPのLB寒天プレート、1XテトのみのLB寒天プレート、および1Xアンプ/テットのみのLB寒天プレート。
- いくつかのグリセロールと殺菌前のプラスチックピペットの先端の箱が手元に近いであることを確認してください。
- 微生物を含む任意の作業を開始する前に、70%のエタノールでワークスペースを殺菌します。ラボコートや手袋など、必要な個人用保護具は必ず着用してください。
- 終了後、70%のエタノールですべての表面と手袋を殺菌し、手を洗います。
2. ドナーおよびレシピエント株の調製
- ドナーおよびレシピエント株の5mL細菌培養物を調作し、通気と220rpmで振盪して37°Cで一晩成長させる。ドナー株は0.3 mM DAPでLBで成長する必要があります。
- 両方の培養物の1 mL(5分間〜3000 rpm)をスピンダウンし、PBSで細胞を洗浄します。
- リン酸緩衝生理食生の500 μLで細胞を再中断する(PBS)。
3. コンジュゲーション
- 50 μLのレシピエント細胞と50μLのドナー細胞をマイクロ遠心管に組み合わせ、ピペッティングで混合します。
- 細胞混合物を37°Cで1時間インキュベートします。これは次のめっきステップの前および間に共役の効率を高める。
- セル混合物のピペット100 μLを1X Tetおよび0.3 mM DAPの寒天板上に置いた。皿の上に広がらないで下さい。
- 1X Tetおよび0.3 mM DAPを用いて寒天プレート上にレシピエント細胞培養のピペット100 μL。皿の上に広がらないで下さい。
- 両方のプレートを37°Cで一晩インキュベートします。
- 生殖細胞スクレーパーで共役細胞混合物とレシピエント細胞培養を掻き取る。無菌マイクロ遠心チューブに細胞を移し、PBSの1 mLで細胞を再懸濁する。
- 細胞を渦にし、穏やかにスピンダウンします(~3000 rpm 5分間)。
- PBSの1 mLで細胞を再中断する。
- 1Xアンプ/テットのみでLB寒天プレート上のコンジュゲーション反応細胞混合物をプレートします。このプレートでは、結合によってアンプ耐性遺伝子を獲得したレシピエント菌のみが増殖することが期待されます。
- レシピエントセルをLB寒天プレートに1X Tetのみでプレートします。このプレートでは、アンプ耐性遺伝子を持たない未結合レシピエント細菌のみが増殖することが期待されます。
- プレートを37°Cで一晩インキュベートします。
- 両方のプレートから単一のコロニーを選択し、メディアの5 mL(220 rpmで通気で37°C)で一晩培養を成長させるためにそれらを使用します。
4. DNA分離
- DNAミニプレップにより総培養量の4.5mLを用いて、以前に調製した培養物からDNAを分離する。
- これを行うには、ヌクレアーゼフリー水の35 μLを使用してDNAを溶出します。
- 純DNAは約1.8の吸光度比(A260/280)を生成します。
- 各培養の残りの0.5 mLを使用して、細菌培養と100%グリセロールの1:1混合物を作ることによってグリセロールストックを調製します。
- 冷凍庫の在庫は-80 °C.
5. PCRによるプラスミド転写の確認
- 2つのPCRマスターミックスを準備し、それぞれが異なる前方プライマーとリバースプライマーのセットを持ち、1つはアンピシリン耐性遺伝子内の500塩基セグメントを標的とし、もう1つはハウスキーピング遺伝子内のセグメントを標的とする。
- ハウスキーピング遺伝子プライマーは、DNAジャイラゼB(14)をコードする細菌遺伝子内のDNAのセグメントを増幅するように設計された。
- 以下の試薬量を使用して、各マスターミックスの90 μLを調作しました。
10 μM 前方プライマーの 7.5 μL
10 μM 逆プライマーの 7.5 μL
2X PCRマスターミックスの75 μL
- 15 μLマスターミックス、10ngのテンプレートDNA、および25 μLの最終容積までのヌクレアーゼフリー水を使用して、以下の6つのPCR反応を調製します。
共役反応DNAとアンピシリンプライマー
コンジュゲーション反応DNAとハウスキーピングプライマー
陰性制御DNAとアンピシリンプライマー
陰性制御DNAとハウスキーピングプライマー
DNAとアンピシリンプライマーなし
DNAとハウスキーピングプライマーなし - ブロックを98°Cに予熱したPCRマシンにこれらの反応を転送し、次の条件下でサーモサイクリングを開始します。
98 °C 30 秒
98 °C の 25 ~ 35 サイクル(5~10 秒)、10 ~ 30 秒の 45 ~72 °C、および 1 kb あたり 15 ~ 30 秒の 72 °C
72 °C 5-10分
4 °Cでホールド - 6つのPCR反応をすべて1%のアガロースゲルにロードし、約20分間〜150Vで実行します。
- UVイルミレータを使用してPC製品を視覚化します。
大腸菌などの細菌細胞は、遺伝情報を細胞から細胞に移すことができる。結合は、細胞間の物理的な接触を必要とするという点で、転写や形質転換などのDNA伝達の他のメカニズムとは異なる。
続行するには、生殖能力を発現するドナー細胞、またはF因子およびそれなしでレシピエント細胞を発現するドナー細胞、Fマイナスセルが必要である。このプロセスには 2 つの手順が必要です。1つ目は、細胞間接触の直接的な確立である。これを行うには、ドナー細胞は、性ピラスと呼ばれる細胞外糸状構造を生成する。これは、コンジュゲーションは無性繁殖細菌の交配の一形態であるので、この名前が付けられていますが、ゲームが交換されず、子孫が形成されないため、それは真の性的生殖ではないことに留意すべきです。
第2のステップは、レシピエント細胞へのDNAの送達である。性ピラスが2つの細胞間の接触を確立した後、タイプIV分泌システムと呼ばれる導管が構築され、DNAの移動を可能にする。次いで、ドナー細胞は、OriTまたは転移の起源として知られている遺伝的要素の存在に基づいて選択される転写される外染色体DNAを複製し始める。新たに複製されたDNAの一端は、DNAタンパク質結合を介して導管に通される。DNAがさらに複製されると、チャネルを通してポンピングされ、OriTの近くに位置する遺伝子によってコードされるタンパク質の複合体によって促進される。DNAが完全に移された後、余分な染色体プラスミドを形成するか、またはレシピエント細胞の染色体に統合されてもよい。転送されたDNAのエンドポイントのいずれにしても、それがコードする遺伝子は発現されます。この遺伝子発現は、成功した結合を確認するために使用することができる。
例えば、ドナー株がアンピシリン耐性を発現し、共役DNAでこれをレシピエント細菌に渡すシナリオを考えてみましょうが、レシピエント株はドナーに存在しないテトラサイクリン耐性遺伝子も有する。この場合、細胞がテトラサイクリンとアンピシリンの両方を含むLB培地にメッキされると、コロニーは正常に共役細菌からのみ成長する必要があり、これは両方の耐性表現型を発現することになる。さらに成功した結合を確認するために、これらのコロニーからのプラスミドDNAを採取することができ、次に、移されたプラスミドに特異的なDNAのセクションをポリメラーゼ連鎖反応、またはPCRを使用して増幅することができる。PCR製品が標準サイズのはしごと一緒に電気泳動ゲル上で実行される場合、既知のサイズのPCR断片がゲル上に表示され、さらに成功した結合を確認する必要があります。この実験では、プラスミドを使用して、ドナー株からテトラサイクリン耐性レシピエント株への結合を介してアンピシリン耐性遺伝子を伝達する。この後、共役を確認するために、共役混合物は、形質転換細菌のみを残して両方の抗生物質を含むプレート上でインキュベートされる。最後に、成功した共役はPCRでさらに確認されます。
手順を開始する前に、ラボコートや手袋を含む適切な個人用保護具を着用してください。次に、70%のエタノールを使用してワークスペースを殺菌し、表面を拭き取ります。
この手順では、アンピシリン耐性遺伝子を大腸菌のWM3064株から大腸菌のJ53株に結合を介して転移する。ドナー株WM3064はテトラサイクリンおよびアンピシリンに対して耐性であり、成長するためにジアミノピメル酸、またはDAPを必要とする。レシピエント株J53はテトラサイクリンに対してのみ耐性であり、DAPが成長する必要はありません。これは、正常に共役細胞がテトラサイクリンとアンピシリンに耐性であるべきであり、DAPなしで成長できることを意味します。
凍結ドナー株グリセロールストックのスクラップでDAPの0.3ミリモルを含むLBの5ミリリットルを接種することにより、ドナー株培養を調製する。次いで、凍結レシピエント株グリセロールストックのスクラップを用いてDAPなしでLBブロスの5ミリリットルを接種してレシピエント株を調製する。これらの培養物を37°Cで一晩成長させ、220 RPMで揺れ動きます。培養物が2のOD 600に成長したら、それぞれから1ミリリットルの培養物を取り除き、これを2つの別々の1.5ミリリットルマイクロ遠心管に入れます。次いで、これらのアリコートを3000RPMで5分間遠心分離し、細菌細胞をペレットする。上清を廃棄し、1X PBSの250マイクロリットルで各ペレットを洗浄します。サンプルを再度遠心分離し、上清を廃棄した後、PBSの500マイクロリットルで各ペレットを再懸濁する。
共役手順を開始するには、まず50マイクロリットルのレシピエント細胞と50マイクロリットルのドナー細胞を1.5ミリリットルのマイクロ遠心分離管に組み合わせ、上下にピペッティングして混ぜます。次に、レシピエント細胞培養のピペット100マイクロリットルをDAPを含む別の1Xテトラサイクリンプレート上に置く。次に、DAPを含む非選択的寒天プレート上にのみ、レシピエント細胞培養の100マイクロリットルをピペッティングして陰性コントロールを調製する。その後、コンジュゲーションと負のコントロールプレートを一晩37°Cでインキュベートします。
翌日、コロニーを採取して生殖細胞スクレーパーを採取し、結合板から細胞を収穫する。次に、コロニーを1X PBSの1ミリリットルを含む無菌1.5ミリリットルマイクロ遠心管に移す。このプロセスを繰り返して、他のプレートから受信者セルを収集します。
この後、混合するサンプルを渦させる。混合した後、チューブを遠心分離機に移し、細胞を穏やかにペレットにする。上清を廃棄し、その後、PBSの1ミリリットルで細胞ペレットを洗浄し、細胞を再懸濁するためにチューブを渦。遠心分離によって再び細胞をペレット。再び上清を廃棄し、PBSの1ミリリットルで両方の細胞ペレットを再懸濁します。さて、無菌ピペット先端を用いて、1Xテトラサイクリンおよび1Xアンピシリンを含有するDAPを含まないLB寒天板上に共役反応細胞混合物の100マイクロリットルをプレートする。1Xテトラサイクリンと1Xアンピシリンを含むDAPを含まない別のLB寒天プレートにPBS中の同じ細胞混合物の100倍希釈の100マイクロリットルを使用してめっき方法を繰り返します。
最後に、1Xテトラサイクリンのみを有する単一のLB寒天プレート上に陰性対照細胞混合物のピペット100マイクロリットル。摂氏37度で一晩インキュベーションした後、コロニーが見えるはずです。無菌ピペット先端を使用して、共役反応プレートから単一のコロニーを選択し、両方の抗生物質を含む選択的LB培地の5ミリリットルを含むチューブに追加します。次いで、レシピエント細胞板から単一のコロニーを選択してコロニー分離を繰り返す。220 RPMで通気で摂氏37度で一晩これらの文化を成長させます。
翌日、70%のエタノールでベンチトップを拭き取り、インキュベーターからプレートを取り出します。DNAミニ準備キットを使用して、4からDNAを分離します。メーカーの指示に従って各培養の5ミリリットル。DNAミニ準備を完了した後、ヌクレアーゼフリー水の35マイクロリットルを使用してDNAを溶出します。最後に、残りの 0 を使用します。各培養物の5ミリリットルは、1対1の希釈のために100%グリセロールの0.5ミリリットルを追加することにより、1ミリリットルのグリセロールストックを調製する。必要になるまで保存のために、これらのアリコートをマイナス80度に置きます。
PCRによるコンジュゲーションの成功を確認するには、まずマイクロ遠心管に75マイクロリットルの2X PCRマスターミックスを加えてPCRマスターミックスを準備します。次いで、10マイクロモルフォワードプライマーとプラスミドからアンピシリン耐性遺伝子を増幅するように設計された10マイクロモルリバースプライマーのそれぞれに7.5マイクロリットルを加えます。次に、マイクロ遠心チューブに75マイクロリットルの2X PCRマスターミックスを追加し、10マイクロモルフォワードプライマーと10マイクロモルリバースプライマーのそれぞれを追加して、2番目のPCRマスターミックスを準備し、この場合はDNAギラゼ B.
次に、最初のマスターミックスの15マイクロリットルをPCRチューブに追加し、10ナノグラム、テンプレート実験DNAの約2マイクロリットルを同じチューブに追加します。ヌクレルリースフリー水で25マイクロリットルの最終容積まで反応を持ち込みます。これらの手順を繰り返して残りの 5 つの反応を生成し、チューブにここに示すコンポーネントが含まれるようにします。次に、これらの反応をブロックを98°Cに予熱したサーモサイクラーに移し、プログラムを開始します。PCRが完了したら、機械からチューブを取り外します。次に、各反応の2マイクロリットルを装填染料の2マイクロリットルと分子量マーカーの4マイクロリットルを1%のアガロースゲルの連続したウェルに積み込む。ゲルを150ボルトで20分間動かします。最後に、UVイルミレータを使用してゲルを視覚化します。
本実験では、コンジュゲーションを介したアンピシリン耐性遺伝子の正常な転移をPCRを介して確認した。ここで、約500塩基対サイズのバンドは、共役DNAおよびアンピシリンプライマーを含むウェル内で観察されるべきであるが、この例では2つである。ハウスキーピング遺伝子であるDNAジャイラゼBは、それぞれ共役DNAとレシピエント細胞DNAを持つウェル3と5にロードされた。これらのウェルで観察されたバンドは、DNAテンプレートが存在し、PCRが成功したことを確認するための陽性対照として機能します。レシピエント細胞DNAおよびアンピシリンプライマー対に対する反応を含むウェルにおいてバンドが観察されるべきではないが、この例では、レシピエント細胞がアンピシリン耐性ではないため、ウェル4である。さらに、テンプレートDNAを欠く反応ではバンドが観察されなくてはならない、ここで6と7をウェル。これらの条件が満たされれば、アンピシリン耐性遺伝子の正常な転移が確認され、大腸菌のWM3064株から大腸菌のJ53株にアンピシリン耐性を付与する。
Subscription Required. Please recommend JoVE to your librarian.
Results
コンジュゲーションが成功した場合、PCR反応1がロードされたウェルで500塩基対サイズのバンドPCR産物が観察され(図3Bではウェル#2)、PCR反応3がロードされたウェルにはバンドは観察されない(図3B#4)。このバンドの存在は、アンピシリン耐性遺伝子の正常な転移を確認し、それによって大腸菌のJ53株に対するアンピシリン耐性を付与する。

図3B:PCRによるコンジュゲーションの成功の確認B)選択コロニーから単離したDNAを用いてPCR分析を行った。各ウェルの内容は以下の通りです:1)DNAはしご、2)コンジュゲーションDNAとアンピシリンプライマー、3)コンジュゲーションDNAとハウスキーピングプライマー、4)陰性制御DNAとアンピシリンプライマー、5)陰性制御DNAとハウスキーピングプライマー、6)DNAなしアンピシリンプライマー、および7)DNAおよび陰性制御プライマーなし。PCR反応1(ウェル2)から約500塩基対バンドPCR産物の存在、およびPCR反応3(ウェル4)からの本産物の欠如は、正常な結合を確認する。
Subscription Required. Please recommend JoVE to your librarian.
Applications and Summary
結合は、ドナー細胞とレシピエント細胞の直接細胞間接触に依存する水平遺伝子導入の自然発生過程である。このプロセスは、細菌のすべての種類の間で共有され、細菌の進化、最も顕著な抗生物質耐性に役立ってきました。研究室では、他の技術と比較してはるかに破壊的ではない遺伝子導入の効果的な方法として活用できます。研究室の外では、共役を介して細菌から真核生物にDNAを伝達する能力は、遺伝子治療のエキサイティングな新しい道を提供し、これらの自然に存在する遺伝子移植の意味を理解する、例えば、間の関係細菌感染症と癌は、研究の急速に新興分野です。
Subscription Required. Please recommend JoVE to your librarian.
References
- Lederberg J, Tatum, E.L. Gene recombination in Escherichia coli Nature. 1946;158:558.
- Holmes R.K. J, M.G. Genetics: Exchange of Genetic Information. 4th Edition ed. Baron S, editor. Galveston, TX: University of Texas Medical Branch at Galveston; 1996.
- Cruz F, Davies, J. Horizontal gene transfer and the origin of species: lessons from bacteria. Trends in Microbiology. 2000;8:128-33.
- Llosa M, Cruz, F. Bacterial conjugation: a potential tool for genomic engineering. Ressearch in Microbiology. 2005;156:1-6.
- Lacroix B, Citovsky, V. Transfer of DNA from Bacteria to Eukaryotes. mBio. 2016;7(4):1-9.
- Llosa M, et al. Bacterial conjugation: a two-step mechanism for
- DNA transport. Molecular Microbiology. 2002;45:1-8.
- Grohmann E, Muth, G., Espinosa, M. Conjugative Plasmid Transfer in Gram-Positive Bacteria. Microbiology and Molecular Biology Reviews. 2003;67:277-301.
- Firth N, Ippen-Ihler, K, Skurray, RA. Structure and function of the F factor and mechanism of conjugation. Escherichia coli and salmonella: cellular and molecular biology. 1996;2:2377-401.
- Smillie C, Garcillan-Barcia MP, Francia MV, Rocha EPC, De La Cruz F. Mobility of Plasmids. Microbiology and Molecular Biology Reviews. 2010;74(3):434-52.
- Cascales E. Definition of a Bacterial Type IV Secretion Pathway for a DNA Substrate. 2004;304(5674):1170-3.
- Wang P, Yu Z, Li B, Cai X, Zeng Z, Chen X, et al. Development of an efficient conjugation-based genetic manipulation system for Pseudoalteromonas. Microbial Cell Factories. 2015;14(1):11.
- Yi H, Cho YJ, Yong D, Chun J. Genome Sequence of Escherichia coli J53, a Reference Strain for Genetic Studies. Journal of Bacteriology. 2012;194(14):3742-3.
- Baumann RLB, E. H.; Wiseman, J. S.; Vaal, M.; Nichols, J. S. Inhibition of Escherichia coli Growth and Diaminopimelic Acid Epimerase by 3-Chlorodiaminopimelic Acid. Antimicrobial Agents and Chemotherapy 1988;32:1119-23.
- Rocha D, Santos, CS, Pacheco LG. Bacterial reference genes for gene expression studies by RT-qPCR: survey and analysis. Antonie Van Leeuwenhoek. 2015;108:685-93.