

Summary
可在酵母的遗传研究调查的人类基因在细胞DNA的代谢的分子和细胞的功能。方法描述为人类的遗传特征
Abstract
了解人类DNA修复蛋白的遗传途径的角色,是一个严峻的挑战,许多研究者。在哺乳动物系统中的遗传研究已经有限的,由于缺乏现成的工具,包括定义的突变基因的细胞株,表达系统的监管,并适当的可选标记。为了规避这些困难,在低等真核生物的遗传模型系统功能保守的DNA修复蛋白和途径的研究已成为有吸引力的选择。我们已经开发了一个模型酵母系统来研究,沃纳综合征的解旋酶,核酸定义不清的基因功能(
Protocol
1。酵母菌株
菌株与野生型SGS1 TOP3(WT; W303 - 1A,基因型, 共同商定一个 ADE2 1 CANL - 100 HIS3 - 11,15 LEU2 - 3,112 trpl,升URA3 - 1)[2],一个sgs1突变(W1292 - 3C ;基因型共同商定 一个 SUP4 - Ø::URA3 sgs1 - 25 ADE2 1 CAN1 - 100 HIS3 - 11,15 LEU2 - 3,112 TRP1 - 1 URA3 - 1 RAD5 - 535)和一个sgs1 TOP3突变(W1058 - 11C,基因型,MAT 一个 SUP4:URA3 sgs1 - 25 TOP3 - 2::HIS3 ADE2 - 1 CAN1 - 100 HIS3 - 11,15 LEU2 - 3,112 TRP1 - 1 URA3 - 1 RAD5 - 535)已被特点[3],并分别请提供罗德尼罗斯坦博士(哥伦比亚大学)。
2。质粒DNA的建设
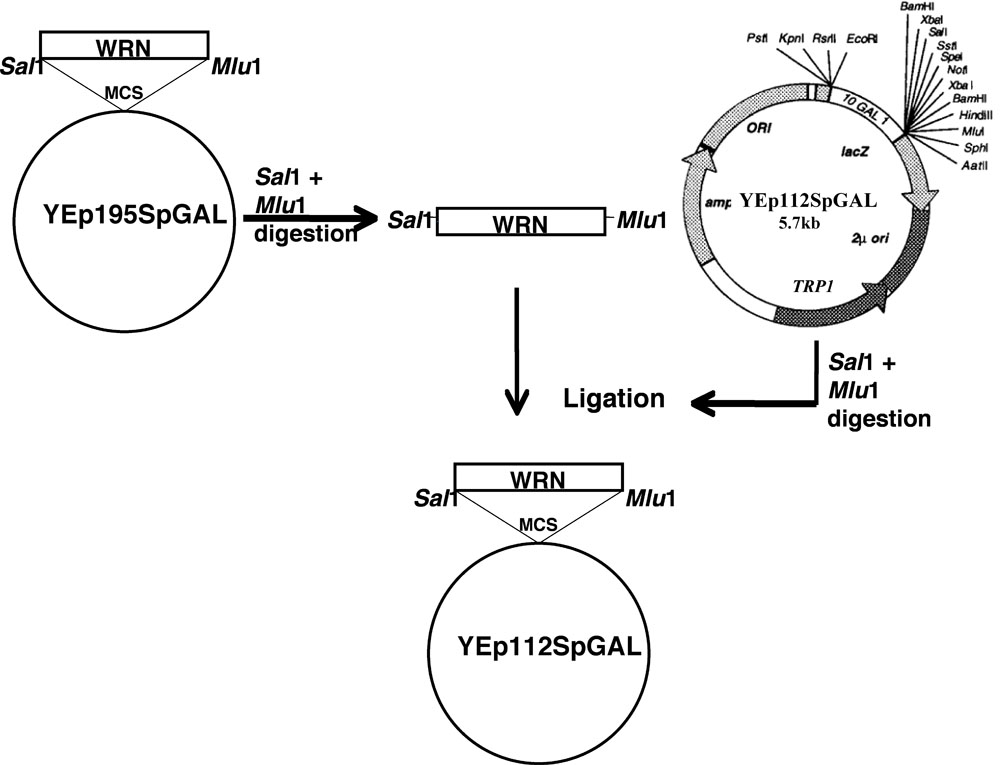
以下所讨论的计划是WRN基因的克隆在YEp112SpGAL 。
- 定点突变WRN(E84A核酸外切酶的死,K577M解旋死了,R834C,和K1016A)在质粒YEp195SpGAL WRN [4]使用快速转换二XL定点突变试剂盒(Stratagene公司)的诱变引物和一个标准的协议Lofstrand实验室(马里兰州盖瑟斯堡)。
- 从各自的YEp195SpGAL WRN结构双酶切后用 Sal I和MLU一凝胶纯化DNA片段编码WRN及其相关变种
- 凝胶纯化的片段,然后克隆到萨尔我MLU我的网站,2微米的多拷贝质粒含有一个TRP1选择标记,构建一个半乳糖苷酶诱导启动子的控制下 YEp112SpGAL WRN或WRN的变种,如在图1所示的矢量YEp112SpGAL 。
- 取得的构造,分别命名为YEp112SpGAL - WRN,YEp112SpGAL WRN E84A,YEp112SpGAL WRN K577M,YEp112SpGAL WRN K1016A和YEp112SpGAL WRN R834C 。
3。转型
酵母培养物生长,使用标准的协议和被执行的转换,使用醋酸锂为基础的协议Gietz等[5]。简单地说,下面的步骤进行。
- 上述菌株培养于5 ml YPD(酵母提取物,蛋白胨,葡萄糖)培养基过夜。 reinoculated于50 ml YPD培养基培养,并获准直到OD 600增长1.0 - 1.2达成。
- 收获培养物,用20毫升水冲洗,并在1 100毫米醋酸锂(LiAc)毫升悬浮。细胞培养在30℃10分钟。
- 细胞沉淀悬浮于100毫米LiAc到终体积为500μL,这是足以做十转换。分装50μL的细胞悬液,离心收集细胞沉淀。
- 细胞沉淀中添加了以下顺序:250μL50%聚乙二醇(PEG)36μL,25μL2 mg / ml的单链DNA(鲑鱼精子DNA),5毫升的质粒DNA(100纳克1米LiAc 1000纳克),和50μl水。细胞沉淀,振荡1分钟。
- 细胞,然后培养于30 ° C为30分钟,热休克在42 ° C为20分钟。
- 收获细胞,并在1毫升水中悬浮。由于质粒YEp112SpGAL包含TRP作为一个可选的标记,所有窝藏突变株的构建将TRP精通。因此,选择SC(合成完整)缺乏葡萄糖媒体色氨酸和孵化30板电镀暂停转化° C为3-4天,直到出现的殖民地。
4。 WRN增长缓慢的表型恢复的遗传分析转化sgs1 TOP3应变。
1。数分析:
- 为了研究上生长的野生型的父母(W303 - 1A),sgs1,或sgs1 TOP3株,与YEp112SpGAL或YEp112SpGAL WRN到SC减去色氨酸板,内含2%葡萄糖挑染相应株 WRN表达的影响( GLU)或2%半乳糖(GAL)。作为对照,sgs1 TOP3 / YEp112SpGAL - SGS1被列入。板在30 ° C孵育2天。
- 要确定的影响WRN蛋白的表达水平较低的增长速度sgs1 TOP3应变能力WRN,sgs1 TOP3应变转化与YEp112SpGAL,YEp112SpGAL WRN,或YEp112SpGAL - SGS1挑染到SC减去色氨酸板含有不同浓度的GAL低至0.005%,高达2%。板,然后在30 ° C孵育2天。
- 为了评估WRN的功能要求, 恢复sgs1 TOP3背景增长缓慢的表型,sgs1 TOP3 YEp112SpGAL WRN(E84A,K577M,R834C,K1016A)转化是streaked到SC减去色氨酸板包含在指定的浓度GLU或GAL。对于控件,sgs1 TOP3应变YEp112SpGAL,YEp112SpGAL WRN或YEp112Sp的GAL - SGS1转化。板,然后在30 ° C孵育2天。
2。液体培养分析:
- 要评估转化sgs1 TOP3液体培养菌株的生长,酵母细胞生长在SC RAF减去色氨酸在30 ° C过夜。文化reinoculated在SC RAF减去色氨酸和生长在30 ° C年初数期( 外径 600〜0.5)。文化,然后在SC媒体含有2%的GAL和增长 0.05外径reinoculated其次,在指定的时间间隔测量吸光度。
5。 WRN羟基脲或methylmethane磺酸敏感性的遗传分析转化酵母菌株。
- 为了确定sgs1或sgs1 TOP3株,株转化与YEp112SpGAL或YEp112SpGAL WRN MMS和胡灵敏度WRN表达的影响,生长在SC RAF减去色氨酸在30 ° C
- 文化reinoculated在SC RAF减去色氨酸和生长在30 ° C至年初数期(外径600〜0.6至0.8)。
- 这些菌株十倍连续稀释准备使用缺乏色氨酸的SC培养基。从文化的大幅增长,5 × 10 6细胞和连续稀释10,000倍。四,每个稀释度微升到SC GAL减去色氨酸板含有胡或MMS表示浓度被发现。在30 ° C孵育板
- 作为对照,野生型菌株(W303 - 1A)转化YEp112SpGAL或YEp112SpGAL WRN,或sgs1 TOP3 / YEp112SpGAL SGS1治疗如上所述。
6。 WRN细胞周期分布改变sgs1 TOP3应变。
- 为了研究细胞周期分布的WRN转化sgs1 TOP3应变,转化YEp112SpGAL,YEp112SpGAL WRN,或YEp112SpGAL - SGS1 sgs1 TOP3的文化,以及矢量转化野生型菌株(W303 - 1A)的资深大律师皇家空军的成长零下30 ° TRP在彗星为通宵。
- 文化reinoculated在外径0.05和增长TIL年初数期(外径0.5 -0.6 600)。WRN诱导表达增加2%终浓度的GAL 。文化,然后让其生长在30 ° 6小时彗星
- 文化当时的DAPI(4',6 diamidino - 2 -苯基吲哚)染色处理。
- 文化收获离心10分钟在室温(RT)在5800 XG。
- 洗涤细胞一次1X磷酸盐缓冲液(PBS),在室温为20分钟,然后被固定在70%的乙醇。
- 固定细胞1 × PBS洗两次,最后悬浮在1X PBS。
- 细胞分别置于幻灯片,并在Vectashield安装,安装用DAPI(1微克/毫升)(矢量,美国)的媒介。一个AXIOVERT 200米显微镜(蔡司镜头100X)和复合微分干涉对比(DIC)的细胞研究。荧光图像分析使用AxioVision,3.0版程序(蔡司)。
7。免疫印迹分析。
1。要确定的WRN蛋白的表达,或转化 sgs1 或sgs1 TOP3株是 WRN变种,准备下面的方法,通过immunblotting分析蛋白质裂解液。
- SC RAF减去色氨酸转化sgs1或sgs1 TOP3株生长在30℃过夜。
- 文化reinoculated和孵育30 ° C直到年初数期(外径600 0.5 - 0.8),然后加入2%终浓度的GAL诱导。文化,然后让其生长在30 ° 6小时彗星
- 离心收集细胞(3毫升),用PBS冲洗。在碱性裂解液悬浮细胞沉淀[50毫米的NaOH,pH为10.5,2毫米EDTA,1毫米苯甲基磺酰氟(PMSF),2%SDS,10%甘油,5%2 - 巯基乙醇和蛋白酶抑制剂(罗氏分子生化)]煮沸5分钟,离心澄清,并与1 M盐酸中和。
- 从细胞裂解液相当于量的8-16%SDS聚丙烯酰胺凝胶蛋白解决。
2。要确定不同的半乳糖浓度的影响下,WRN蛋白的表达水平,文化如下处理。
- TOP3 sgs1转化株生长在30 ° C过夜SC RAF减去色氨酸。
- 文化是reinoculated,并在30 ° C直到年初数期( 外径 600 0.5 - 0.8),进而诱发加入GA孵育l至2%终浓度。文化,然后让其生长在30 ° 6小时彗星
- 离心收集细胞,重悬于缓冲液中(50毫米的Tris - CL,pH值7.5,1%去氧胆酸钠,1%TRITON X - 100,0.1%十二烷基硫酸钠[SDS])含有蛋白酶抑制剂(PMSF 0.05%,0.05微克/ ml的亮肽素)。
- 玻璃珠(425至600微米)的混合物,涡旋破碎细胞。
- 在15000 XG和由此产生的匀浆,离心5 min,收集上清。
- Bradford法蛋白在裂解馏分估计。
- 从不同的裂解液组分的蛋白质浓度相同的是8-16%SDS聚丙烯酰胺凝胶上解决。
3。由西方印迹使用一个WRN鼠单克隆抗体针对一个抗原决定簇的指示,在纯化的C -末端片段WRN WRN或WRN突变蛋白的表达[6](1:1000,春谷实验室)。
4。其次是基层抗体孵育孵化与二级(辣根过氧化物酶标记)抗体(1:5000)。杂交的ECL Plus西方的检测系统,每个制造商的协议进行了处理。
5。对于定量免疫印迹分析,提高纯化的重组凝胶后的印迹的量化标准线性曲线可以产生包括标签全长WRN蛋白的浓度。此外,细胞从细胞或酵母裂解液样品20微克的量相当于提取物被装上凝胶估计 WRN浓度。
6。 ImageQuant然后进行分析,生成凝胶和标准的线性曲线。
7。 WRN蛋白的浓度在不同的半乳糖苷酶浓度,使用标准的线性曲线产生上述估计。
8。代表性的成果
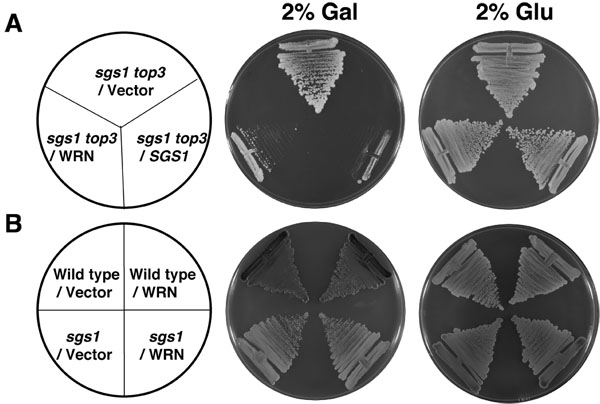
在这项研究中使用的所有菌株色氨酸营养缺陷型。因此,sgs1,sgs1 TOP3和野生型W303 - 1A与YEp112SpGAL或YEp112SpGAL WRN转化株,其成长能力存在SC减去色氨酸媒体的基础上选定的。要研究WRN表达转化sgs1 TOP3突变细胞生长的影响,文化挑染到含有2%的半乳糖苷酶SC减去色氨酸媒体诱发 WRN表达的板块。 TOP3 decatenates交织Sgs1解旋酶生成的复制过程中[1,2] DNA分子,因此,在TOP3的情况下,扭转应力是不能解除导致增长缓慢和超重组。 sgs1遗传功能是抑制增长缓慢的一个TOP3突变表型。如果WRN可替代SGS1与遗传相互作用TOP3,TOP3 sgs1 TOP3增长缓慢的表型恢复将预期。
如图2a所示,是WRN转化sgs1 TOP3应变向量相比增长明显缓慢转化sgs1 TOP3应变。sgs1 TOP3转化的YEp112SpGAL SGS1质粒作为阳性对照(图2A),这表明,野生型表示Sgs1 sgs1 TOP3突变基因互补的生长表型。同样在半乳糖苷酶缺乏增长的转变sgs1 TOP3株为2天(图2A)所示。 WRN表达对父母的野生型(W3031A) 或 sgs1株(图2B)增长没有影响,表明细胞生长的影响,WRN具体sgs1 TOP3突变背景观察。进行的遗传研究表明,WRN解旋酶,但没有外切酶活性恢复 TOP3增长的表型( 图 3B) WRN变种。中WRN自然发生错义多态性与解旋酶的活性的干扰,取消其恢复 TOP3增长缓慢,表型的能力。
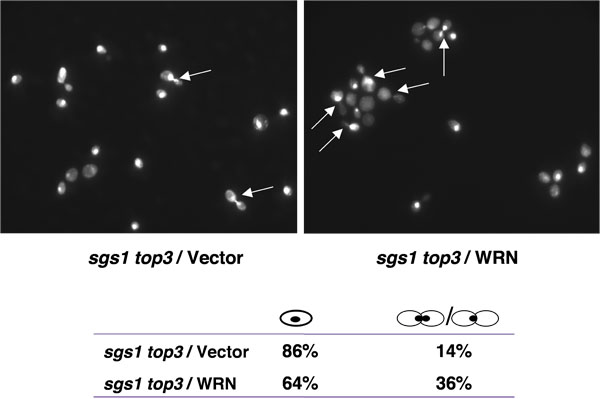
TOP3突变株在细胞周期的后期S/G2阶段延迟[1],可能占到其增长缓慢的一个特点。 TOP3背景SGS1基因突变,抑制在细胞周期的S/G2阶段的延迟。如果WRN表达可以恢复在S/G2阶段sgs1 TOP3的细胞周期的延迟,不可分割的核出芽细胞升高的人口将预计sgs1表达WRN TOP3突变的细胞。正如图4所示,大芽生细胞的百分比明显高于sgs1 TOP3 比sgs1 TOP3 /矢量/ WRN,建议恢复S/G2 DE打下TOP3特点。
sgs1 TOP3双突变是不太敏感的MMS或胡比TOP3单一突变[2]。自sgs1 TOP3 WRN受影响的细胞的生长,我们未来methylmethane的磺酸(MMS,烷化剂)和羟基脲(HU,复制抑制剂)。sgs1 TOP3 / WRN显示这两个药物的敏感性与sgs1 TOP3的敏感性研究其效果/ SGS1(图5)。 WRN变种的遗传分析表明,WRN解旋酶/ ATP酶,但不 WRN外切酶活性,其灵敏度MMS和胡锦涛的影响。

图1。 YEp112SpGAL载体克隆WRN / WRN变种。原理演示 ,请点击这里为图1的放大版本。

图2。WRN在sgs1 TOP3表达恢复增长缓慢,表型的TOP3。 小组,sgs1 TOP3应变YEp112SpGAL或YEp112SpGAL WRN转化SC -色氨酸板,内含2%或2%GLU GAL条纹。作为对照sgs1 TOP3应变改造YEp112SpGAL SGS1的是挑染两个板块。板在30 ° C孵育2天,然后拍照。 小组B,野生型菌株W303 - 1A或sgs1应变YEp112SpGAL或YEp112SpGAL WRN转化SC -色氨酸板块挑染或者在含2%葡萄糖或2% GAL 。板在30℃孵育4天,然后拍照。板块组成,分别在A和B组小组。请点击这里看大图图2。

图3。WRN ATP酶/解旋酶,但没有外切酶活性,需要恢复增长缓慢,表型的TOP3 sgs1 TOP3背景 。sgs1 TOP3应变转化与ATP酶/解旋死(YEp195SpGAL WRN K577M),外切酶的死(YEp195SpGAL WRN E84A),RQC突变(YEp195SpGAL WRN K1016A),多态突变(YEp195SpGAL - WRN R834C)挑染SC -色氨酸要么是2%的葡萄糖(C组 )或2% 加仑(B组 )的板块。板在30 ° C孵育2天,然后拍照。板块组成,在小组答:请点击这里图3看到一个更大的版本。

图4。WRN表达诱导S/G2逮捕sgs1 TOP3细胞。sgs1 TOP3 YEp112SpGAL,YEp112SpGAL WRN,或YEp112SpGAL SGS1转化株对数增长的文化,和矢量转化野生型菌株在半乳糖苷酶浓度为2 %诱导6小时文化收获,加工材料和方法中所述的DAPI染色和使用AXIOVERT 200米显微镜(蔡司镜头100X)观察。显示的是YEp112SpGAL(左上角)转化sgs1 TOP3与YEp112SpGAL WRN(右上)的DAPI染色。箭头显示一心一意核细胞。 G1(单细胞)和S/G2(芽生细胞)的细胞分布在下部面板上显示。图4看到一个更大的版本, 请点击这里。

图5。对数增长sgs1 TOP3 YEp112SpGAL,YEp112SpGAL WRN,死亡外切酶(YEp112SpGAL WRN E84A),ATP酶/解旋死(YEp112SpGAL WRN K577M),转化的应变文化 WRN表达对MMS和胡锦涛sgs1 TOP3应变灵敏度 。 RQC突变(YEp112SpGAL WRN K1016A),多态突变(YEp112SpGAL - WRN R834C),YEp112SpGAL SGS1和载体转化野生型株父母发现一个到SC -色氨酸含葡萄糖或半乳糖苷酶和MMS或胡板10倍的系列稀释在指定的浓度。培养板在30 ° C为2天(控制盘),第4天(MMS和胡板),然后拍照。请9/1639fig5.jpg“>点击这里看大图图5。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
利用酵母作为模型系统的优势之一是在定义的DNA复制和修复的途径,酵母和人类之间的保守突变体的可用性。进一步选择窝藏的特定基因的转化是很容易的和可靠的实验室菌株营养缺陷型突变体,并与营养缺陷型标记载体,都是现成的。使用这些载体的基因产物的表达可诱导启动子的控制(例如,半乳糖苷酶诱导启动子,等)置于受。考虑到这些优势,我们开发的酵母为基础的模型系统研究沃纳综合症在一个保守的途径,是潜在的人类和酵母之间有缺陷的人类WRN基因的功能要求。我们使用的描述方法,WRN能作用于一个影响TOP3相关表型的遗传途径,提供了第一手的证据。我们的观察提示WRN的功能与人体细胞Top3α相互作用在细胞DNA复制或重组的可能性进一步调查。可以想象,在博莱霉素受损情况,WRN可部分替代对博莱霉素通过其蛋白与拓扑异构酶合作。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
完全支持这项工作是由美国国立卫生研究院,国家老龄问题研究所的院内研究计划。我们感谢SGS1表达质粒的酵母菌株和布拉德约翰逊博士(美国宾夕法尼亚大学医学院,费城,宾夕法尼亚)的博士罗德尼罗斯坦(哥伦比亚大学)。
References
- Aggarwal, M., Brosh, R. M. Jr WRN helicase defective in the premature aging disorder Werner Syndrome genetically interacts with Topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3. Aging. 1, 219-233 (2009).
- Gangloff, S., McDonald, J. P., Bendixen, C., Arthur, L., Rothstein, R. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol Cell Biol. 14, 8391-8398 (1994).
- Shor, E., Gangloff, S., Wagner, M., Weinstein, J., Price, G., Rothstein, R. Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae. Genetics. 162, 647-662 (2002).
- Sharma, S., Sommers, J. A., Brosh, R. M. Jr In vivo function of the conserved non-catalytic domain of Werner syndrome helicase in DNA replication. Hum Mol Genet. 13, 2247-2261 (2004).
- Gietz, R. D., Schiestl, R. H., Willems, A. R., Woods, R. A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 11, 355-360 (1995).
- von Kobbe, C., Thoma, N. H., Czyzewski, B. K., Pavletich, N. P., Bohr, V. A. Werner syndrome protein contains three structure specific DNA binding domains. J Biol Chem. 278, 52997-53006 (2003).

{kind=link}
{kind=link}
{kind=link}
{kind=link}