

Summary
Estudos de genética em leveduras podem ser empregadas para investigar as funções de celular e molecular dos genes humanos no metabolismo do DNA celular. Métodos são descritos para a caracterização genética do ser humano
Abstract
Compreender o papel das proteínas de reparo de DNA humano em vias de genética é um grande desafio para muitos pesquisadores. Estudos genéticos em sistemas de mamíferos têm sido limitados devido à falta de ferramentas disponíveis, incluindo linhas de células mutantes definido genético, sistemas de expressão regulamentar, e apropriado marcadores selecionáveis. Para contornar essas dificuldades, os sistemas de modelo genético em eucariotos inferiores tornaram-se uma opção atraente para o estudo de proteínas funcionalmente DNA conservado reparação e caminhos. Nós desenvolvemos um sistema de levedura modelo para estudar as funções mal definidas genética da síndrome de Werner helicase-nuclease (
Protocol
1. As cepas de levedura
Cepas com tipo selvagem SGS1 TOP3 (WT; W303-1A, genótipo, MAT uma ade2-1 canl-100 his3-11, 15 leu2-3, 112 TRPL-l ura3-1) [2], um mutante sgs1 (W1292-3C ; MAT genótipo uma SUP4-o:: URA3 sgs1-25 ade2-1 CAN1-100 his3-11, 15 leu2-3, 112 trp1-1 ura3-1 rAd5-535) e um sgs1 Top3 mutante (W1058-11C, genótipo, MAT um SUP4-o:: URA3 sgs1-25 Top3-2:: HIS3 ade2-1 CAN1-100 his3-11, 15 leu2-3, 112 trp1-1 ura3-1 rAd5-535) foram caracterizados [3] e foram gentilmente cedidas pelo Dr. Rodney Rothstein (Columbia University).
2. Plasmídeo DNA construções
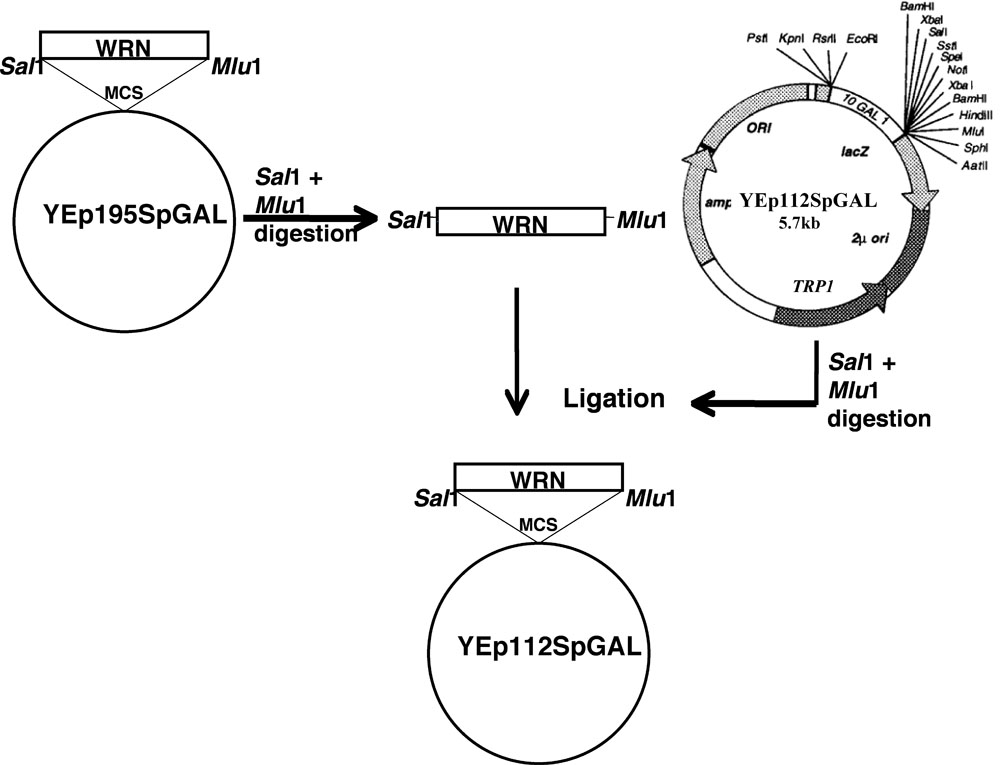
O gene WRN foi clonado em YEp112SpGAL seguindo o esquema como discutido.
- Site dirigido mutações do WRN (E84A exonuclease mortos, K577M helicase mortos, R834C e K1016A) no plasmídeo YEp195SpGAL-WRN [4] foram construídos utilizando primers mutagênicos e um protocolo padrão de Quickchange II XL-site dirigido mutagênese kit (Stratagene) por Lofstrand labs (Gaithersburg, MD).
- Fragmentos de DNA de codificação de WRN e suas variantes associadas foram purificados do gel respectivos YEp195SpGAL-WRN construções após a digestão dupla com Sal I e I. Mlu
- Fragmentos de gel purificados foram então clonados no I Sal locais Mlu I do vector YEp112SpGAL, a 2 m de cópia multi-plasmídeo contendo um marcador TRP1 selecionáveis para a construção de WRN YEp112SpGAL ou variantes WRN sob o controle de um promotor induzível-gal, como mostrado na Figura 1 .
- As construções assim obtidos foram nomeados como YEp112SpGAL-WRN, YEp112SpGAL-WRN E84A, YEp112SpGAL-WRN K577M, YEp112SpGAL-WRN K1016A e YEp112SpGAL-WRN R834C.
3. Transformação
Culturas de leveduras foram cultivadas usando o protocolo padrão e transformações foram realizadas usando um acetato com base em protocolo de lítio por Gietz et al [5]. Resumidamente, os passos seguintes foram realizados.
- Culturas das cepas acima mencionadas foram cultivadas durante a noite em 5 ml YPD (extrato de levedura-peptona-dextrose) médio. Culturas foram reinoculados em 50 ml meio YPD e foram autorizados a crescer até que uma OD 600 de 1,0-1,2 foi alcançado.
- Culturas foram colhidas, lavadas com 20 ml de água, e ressuspenso em 1 ml de acetato de lítio de 100 mM (LiAc). As células foram incubadas a 30 ° C por 10 min.
- Pellet celular foi ressuspenso em 100 mM LiAc para um volume final de 500 mL, que é o suficiente para fazer dez transformações. 50 ul da suspensão de células foi aliquotado e pellet celular foi coletado por centrifugação.
- Para o pellet celular foi adicionado na seguinte ordem: 250 mL polietilenoglicol de 50% (PEG), 36 mL de 1 M LiAc, 25 mL de 2 mg / ml ssDNA (DNA de esperma de salmão), 5 ml de DNA plasmídeo (100 ng 1000 ng), e 50 mL de água. Pellet celular foi agitadas durante 1 min.
- As células foram então incubadas a 30 ° C por 30 min seguido por um choque térmico a 42 ° C por 20 min.
- As células foram colhidas e ressuspenso em 1 ml de água. Desde o YEp112SpGAL plasmídeo contém trp como marcador de selecção, todas as cepas mutantes abrigar a construção seria trp proficiente. Portanto, transformantes foram selecionados por plaqueamento da suspensão em SC (Synthetic completa) media glucose falta Trp e incubação das placas a 30 ° C por 3-4 dias até as colônias aparecer.
4. Análise genética da restauração retardar o crescimento no fenótipo transformado WRN sgs1 tensão Top3.
1. Análise Streak:
- Para examinar o efeito de expressão WRN sobre o crescimento do tipo selvagem parental (W303-1A), sgs1, ou sgs1 Top3 cepas, as cepas correspondente com YEp112SpGAL ou YEp112SpGAL-WRN eram listrados em SC Trp menos placas contendo um ou 2% de glicose ( glu) ou 2% galactose (gal). Como controle, sgs1 Top3 / YEp112SpGAL-SGS1 foi incluído. Placas foram incubadas a 30 ° C por 2 dias.
- Para determinar a capacidade de WRN para afetar a taxa de crescimento da cepa sgs1 Top3 no menor WRN níveis de expressão da proteína, o sgs1 Top3 estirpe transformada com YEp112SpGAL, YEp112SpGAL-WRN, ou YEp112SpGAL-SGS1 foi semeada SC Trp menos placas contendo concentrações variadas de gal de tão baixo quanto 0,005% para tão alto quanto 2%. As placas foram então incubadas a 30 ° C por 2 dias.
- Para avaliar os requisitos funcionais de WRN para restaurar o fenótipo de crescimento lento no Top3 sgs1 fundo, sgs1 Top3 transformada com YEp112SpGAL-WRN (E84A, K577M, R834C e K1016A) foram streaked em SC menos Trp placas contendo um ou outro ou glu gal nas concentrações indicadas. Para controles, sgs1 tensão Top3 transformada com YEp112SpGAL, YEp112SpGAL-WRN ou YEp112Sp GAL-SGS1 foi incluído. As placas foram então incubadas a 30 ° C por 2 dias.
2. Análise de cultura líquida:
- Para avaliar o crescimento de cepas transformada sgs1 Top3 na cultura líquido, células de levedura foram cultivadas em SC raf menos Trp a 30 ° C durante a noite. Culturas foram reintroduzidas in vitro na SC raf menos Trp e cresceram a 30 ° C para a fase log precoce (OD 600 de ~ 0,5). Culturas foram então reinoculados no OD 0,05 em media SC gal contendo 2% eo crescimento foi seguido por medição de absorbância em intervalos de tempo indicado.
5. Análise genética de hidroxiuréia ou methylmethane-sulfonato de sensibilidade em cepas de leveduras transformadas WRN.
- Para determinar o efeito da expressão da sensibilidade WRN MMS e HU de sgs1 ou sgs1 Top3 cepas, estirpes transformadas com YEp112SpGAL ou YEp112SpGAL-WRN foram cultivadas em SC raf menos Trp a 30 ° C.
- Culturas foram reintroduzidas in vitro na SC raf menos Trp e cresceram a 30 ° C para a fase log precoce (OD 600 de ~ 0,6-0,8).
- Dez diluições dessas cepas foram preparados utilizando SC médias falta Trp. De uma cultura em crescimento exponencial, 5 x 10 6 células foram retiradas e diluídas serialmente até 10.000 vezes. Quatro microlitros de cada diluição foi flagrada em SC gal placas menos Trp contendo concentrações indicadas do HU ou MMS. Placas foram incubadas a 30 ° C.
- Como controle, do tipo selvagem cepa parental (W303-1A) transformada com YEp112SpGAL ou YEp112SpGAL-WRN, ou sgs1 Top3 / YEp112SpGAL-SGS1 foi tratado como descrito acima.
6. Distribuição do ciclo celular de WRN transformado sgs1 tensão Top3.
- Para estudar a distribuição do ciclo celular de WRN transformado sgs1 tensão Top3, culturas de sgs1 Top3 transformada com YEp112SpGAL, YEp112SpGAL-WRN, ou YEp112SpGAL-SGS1, eo vetor-transformadas cepa selvagem parental (W303-1A) foram cultivadas em SC raf trp menos a 30 ° C para pernoite.
- Culturas foram reinoculados no OD 0,05 e permitiu a crescer até início de fase log (OD 600 de 0,5 -0,6). Expressão WRN foi induzida pela adição de gal a 2% de concentração final. Culturas foram, então, autorizado a crescer a 30 ° C por 6 h.
- Culturas foram então processados para DAPI (4 ', 6-diamidino-2-phenylindole) coloração.
- Culturas foram colhidas por centrifugação a 5800 xg por 10 min à temperatura ambiente (RT).
- Células foram lavadas uma vez com 1X tampão fosfato (PBS) e foram fixados em etanol 70% por 20 min em temperatura ambiente.
- Células fixadas foram lavadas duas vezes com PBS 1 X e, finalmente ressuspenso em PBS 1X.
- As células foram colocadas em slide e montado em Vectashield meio de montagem com DAPI (1 mg / ml) (Vector, EUA). As células foram examinadas com um microscópio M Axiovert 200 (Zeiss, lentes de 100x) e contraste de interferência diferencial composto (DIC). Imagens de fluorescência foram analisados usando AxioVision, versão 3.0 do programa (Zeiss).
7. Blot análises ocidentais.
1. Para determinar a expressão da proteína de WRN ou as variantes WRN em sgs1 transformado ou sgs1 Top3 cepas, lisados foram preparados pelo método a seguir e proteínas analisadas por immunblotting.
- Transformado sgs1 ou sgs1 Top3 cepas foram cultivadas em SC raf menos Trp a 30 ° C para pernoite.
- Culturas foram reinoculados e incubados a 30 ° C até a fase log precoce (OD 600 de 0,5 - 0,8) e, em seguida, induzida pela adição de gal a 2% de concentração final. Culturas foram, então, autorizado a crescer a 30 ° C por 6 h.
- Células (3 ml) foram coletadas por centrifugação e lavadas com PBS. Pellet celular foi ressuspenso em tampão de lise alcalina [50 mM NaOH, pH 10,5, 2 mM EDTA, 1 mM de flúor fenilmetilsulfonil (PMSF), 2% SDS, glicerol 10%, 5% 2-mercaptoetanol e inibidores da protease (Bioquímicos Roche Molecular)] , fervido por 5 min, clarificadas por centrifugação, e neutralizados com 1 M HCl.
- Proteínas a partir de quantidades equivalentes de lisado celular foram resolvidos em géis de poliacrilamida 8-16% SDS.
2. Para determinar o nível de expressão da proteína WRN sob a influência de diferentes concentrações de galactose, as culturas foram processadas como se segue.
- Transformado sgs1 Top3 cepas foram cultivadas em SC raf menos Trp a 30 ° C para pernoite.
- Culturas foram reinoculados e incubados a 30 ° C até a fase log precoce (OD 600 de 0,5 - 0,8) e, em seguida, induzida pela adição de gal para 2% de concentração final. Culturas foram, então, autorizado a crescer a 30 ° C por 6 h.
- As células foram colhidas por centrifugação e ressuspendidas em tampão (50 mM Tris-Cl, pH 7,5, desoxicolato de sódio a 1%, 1% de triton X-100, sulfato de sódio a 0,1% dodecil [SDS]) contendo inibidores da protease (PMSF 0,05%, 0,05 mg / ml leupeptin).
- Contas de vidro (425-600 mm) foram adicionados à mistura, e as células foram quebradas por vórtex.
- O homogenato resultante foi centrifugado por 5 min a 15.000 xg eo sobrenadante foi coletado.
- Proteína nas frações lisado foi estimada pelo método de Bradford.
- Concentração igual de proteínas de diferentes frações lisado foi resolvido em gel de poliacrilamida 8-16% SDS.
3. Expressão de proteínas WRN ou WRN mutante foi determinada por Western blot utilizando um anticorpo monoclonal de rato WRN dirigido contra um epitopo em um fragmento purificado C-terminal de WRN [6] (1:1000, Spring Valley Labs).
4. Incubação do anticorpo primário foi seguido de incubação com um anticorpo (peroxidase conjugada) secundário (1:5000). Borrões foram processados com um sistema de detecção ECL Plus ocidental por protocolo do fabricante.
5. Para a análise quantitativa blot ocidentais, aumentando as concentrações de recombinantes purificadas His-tag proteína WRN full-length foram incluídos em gel de tal forma que sobre a quantificação dos blot uma curva padrão linear poderia ser gerada. Além disso, extrair células de quantidade equivalente de células ou 20 mg das amostras de lisado de levedura foram carregados em gel para estimar a concentração de WRN.
6. ImageQuant análise foi então realizada em gel e curva padrão linear foi gerado.
7. Concentração de proteína WRN em concentrações variando gal foi estimado pela curva linear padrão gerado acima.
8. Resultados representante
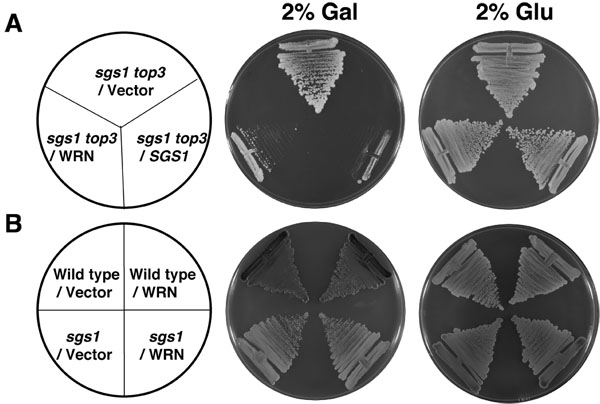
Todas as cepas utilizadas neste estudo são auxotrophs Trp. Portanto, sgs1, sgs1 Top3 e do tipo selvagem W303-1A cepas transformada com YEp112SpGAL ou YEp112SpGAL-WRN foram selecionados com base em sua capacidade de crescer na presença de SC media menos Trp. Para examinar o efeito de expressão WRN sobre o crescimento da transformada sgs1 Top3 de células mutantes, as culturas foram semeadas em placas contendo SC menos Trp mídia com gal 2% de induzir a expressão WRN. Top3 decatenates entrelaçadas moléculas de DNA gerados por Sgs1 helicase durante a replicação [1, 2] e, portanto, na ausência de Top3, stress torção não for aliviada, resultando em crescimento lento e hiper-recombinação. A função genética da sgs1 é suprimir o fenótipo lento crescimento de um mutante Top3. Se WRN poderia substituir SGS1 na interação genética com Top3, a restauração do fenótipo Top3 de crescimento lento em sgs1 Top3 seria de esperar.
Conforme mostrado na Figura 2A, o WRN transformado sgs1 tensão Top3 cresceu significativamente mais lentamente em comparação com o vetor transformado sgs1 tensão Top3. Sgs1 Top3 transformado com o plasmídeo YEp112SpGAL-SGS1 foi incluída como um controle positivo (Figura 2A), demonstrando que o tipo selvagem Sgs1 expressa no Top3 sgs1 mutante foi capaz de complementar o fenótipo geneticamente crescimento. A transformada sgs1 Top3 cepas cresceu da mesma forma, na ausência de gal, como mostrado por 2 dias (Figura 2A). Expressão de WRN não teve efeito sobre o crescimento de cepas parentais de tipo selvagem (W3031A) ou sgs1 (Figura 2B), indicando que o efeito de WRN no crescimento celular foi específico ao observado no Top3 sgs1 fundo mutantes. Estudos genéticos realizados utilizando variantes WRN demonstrou que WRN helicase, mas não a atividade exonuclease era necessária para a restauração do fenótipo Top3 do crescimento (Figura 3B). Um polimorfismo missense ocorrem naturalmente em WRN que interfere na atividade helicase aboliu a sua capacidade de restaurar Top3 fenótipo de crescimento lento.
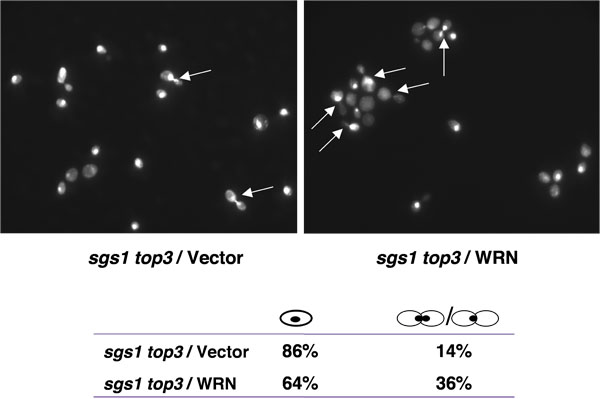
Top3 cepas mutantes estão atrasadas na fase S/G2 tardia do ciclo celular [1], uma característica que pode ser responsável por seu crescimento lento. Mutação do gene SGS1 no fundo Top3 suprime o atraso na fase S/G2 do ciclo celular. Se a expressão WRN poderia restaurar o atraso na fase S/G2 do ciclo celular em sgs1 Top3, uma população elevada de grandes células enxertadas com núcleos indivisa seria esperado para sgs1 Top3 de células mutantes expressando WRN. Conforme mostrado na Figura 4, o percentual de grandes células enxertadas foi maior para sgs1 Top3 WRN / sgs1 Top3 do que vector /, sugerindo a restauração do de S/G2 leigos característica de Top3.
O sgs1 Top3 duplo mutante é menos sensível a MMS ou HU do que o mutante Top3 único [2]. Desde WRN o crescimento de células afetadas do sgs1 Top3, nós próximos examinaram seu efeito sobre a sensibilidade à methylmethane sulfonato (MMS, um agente alquilante) e hidroxiuréia (HU, um inibidor da replicação). Sgs1 sensibilidade Top3 / WRN exibido para ambas as drogas comparável ao sgs1 Top3 / SGS1 (Figura 5). Análise genética de variantes WRN revelou que WRN helicase / ATPase, mas não a atividade exonuclease WRN, era necessária para seu efeito sobre a sensibilidade ao MMS e HU.

Figura 1. Apresentação esquemática para clonagem WRN / WRN variantes no vetor YEp112SpGAL. Por favor, clique aqui para uma versão ampliada da figura 1.

Figura 2. WRN expressão em sgs1 Top3 restaura o fenótipo de crescimento lento Top3. Painel A, sgs1 Top3 estirpe transformada com YEp112SpGAL ou WRN YEp112SpGAL foram semeado em uma placa SC-Trp contendo um ou outro glu 2% ou 2% gal. Como um controle de tensão sgs1 Top3 transformada com YEp112SpGAL SGS1 foi semeado em ambas as placas. Placas foram incubadas a 30 ° C por 2 dias e depois fotografadas. Painel B, cepa selvagem W303-1A pais ou tensão sgs1 transformada com YEp112SpGAL ou WRN YEp112SpGAL foram semeado em uma placa SC-Trp ou contendo glu 2% ou 2% gal . Placas foram incubadas a 30 ° C por 4 dias e depois fotografadas. Composição das placas foi como no Painel A e Painel B, respectivamente. Por favor, clique aqui para ver uma versão ampliada da figura 2.

Figura 3. WRN ATPase / helicase, mas não a atividade exonuclease, é necessário para restaurar o fenótipo de crescimento lento no Top3 sgs1 fundo Top3. Sgs1 tensão Top3 transformada com mortos-helicase ATPase / (YEp195SpGAL WRN K577M), exonuclease-morto (YEp195SpGAL- WRN E84A), RQC mutante (YEp195SpGAL-WRN K1016A), ou polimórficos mutante (YEp195SpGAL-WRN R834C) foi semeado em SC-Trp placas contendo um ou outro glu 2% (Painel C) ou 2% gal (Painel B). Placas foram incubadas a 30 ° C por 2 dias e depois fotografadas. Composição das placas foi como no Painel de A. Por favor, clique aqui para ver uma versão ampliada da figura 3.

Figura 4. Expressão WRN induz S/G2 prisão em células sgs1 Top3. Culturas logaritmicamente crescente de tensão sgs1 Top3 transformada com YEp112SpGAL, WRN YEp112SpGAL, ou YEp112SpGAL SGS1, eo vetor-transformadas cepa selvagem dos pais foram induzidos na concentração de 2% gal por 6 h. Culturas foram colhidas, processadas para coloração DAPI, conforme descrito em Materiais e Métodos e foram observadas utilizando microscópio Axiovert M 200 (Zeiss, lentes de 100x). Mostrada é a coloração DAPI do Top3 sgs1 transformada com YEp112SpGAL (superior esquerdo) e com YEp112SpGAL WRN (superior direito). Setas indicam células com núcleos indivisível. Distribuição das células no G1 (células individuais) e S/G2 (células enxertadas) é mostrado na parte inferior do painel. Por favor, clique aqui para ver uma versão ampliada da figura 4.

Figura 5. Efeito da expressão da sensibilidade WRN MMS e HU de sgs1 tensão Top3. Logaritmicamente crescente culturas de sgs1 tensão Top3 transformada com YEp112SpGAL, YEp112SpGAL WRN, exonuclease-morto (YEp112SpGAL-WRN E84A), ATPase / helicase-morto (YEp112SpGAL WRN K577M), RQC mutante (YEp112SpGAL-WRN K1016A), polimórfico mutante (YEp112SpGAL-WRN R834C), YEp112SpGAL SGS1 e vetor transformado tipo selvagem estirpes parentais foram vistos em um diluições em dez vezes de série em SC-Trp placas contendo glu ou gal e ou MMS ou HU nas concentrações indicadas. Placas foram incubadas a 30 ° C por 2 dias (placas de controle) e 4 dias (MMS e placas HU) e, em seguida, fotografados. Por favor9/1639fig5.jpg "> clique aqui para ver uma versão ampliada da figura 5.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Um dos pontos fortes da utilização de leveduras como um sistema modelo é a disponibilidade de mutantes na replicação do DNA definido e vias de reparação que são conservados entre leveduras e humanos. Uma outra seleção de transformantes abrigar os genes específicos é fácil e confiável como as cepas de laboratório são mutantes auxotróficos e vetores com marcadores auxotróficos estão prontamente disponíveis. Usando esses vetores a expressão de produtos de genes pode ser regulada, colocando-os sob o controle de um promotor induzível (por exemplo, gal promotor induzível, etc.) Considerando estas vantagens, nós desenvolvemos um sistema de levedura modelo baseado para estudar os requisitos funcionais do gene WRN humanos defeituosos em Síndrome de Werner em um caminho que é potencialmente conservada entre o humano eo fermento. Usando as abordagens descritas, que forneceu a primeira evidência de que WRN podem funcionar em um caminho genético que afeta Top3 relacionados com fenótipos. Nossas observações prompt de uma investigação mais aprofundada da possibilidade de que WRN funcionalmente interage com Top3α em células humanas durante a replicação do DNA celular ou recombinação. Concebível, em uma condição BLM-prejudicada, WRN podem substituir parcialmente para BLM através da sua parceria proteína com uma topoisomerase.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Este trabalho foi financiado integralmente pelo programa de Pesquisa Intramural do, NIH National Institute on Aging. Agradecemos ao Dr. Rodney Rothstein (Columbia University) para as cepas de leveduras e Brad Dr. Johnson (University of Pennsylvania School of Medicine, na Filadélfia, Pensilvânia) para a expressão SGS1 plasmídeo.
References
- Aggarwal, M., Brosh, R. M. Jr WRN helicase defective in the premature aging disorder Werner Syndrome genetically interacts with Topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3. Aging. 1, 219-233 (2009).
- Gangloff, S., McDonald, J. P., Bendixen, C., Arthur, L., Rothstein, R. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol Cell Biol. 14, 8391-8398 (1994).
- Shor, E., Gangloff, S., Wagner, M., Weinstein, J., Price, G., Rothstein, R. Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae. Genetics. 162, 647-662 (2002).
- Sharma, S., Sommers, J. A., Brosh, R. M. Jr In vivo function of the conserved non-catalytic domain of Werner syndrome helicase in DNA replication. Hum Mol Genet. 13, 2247-2261 (2004).
- Gietz, R. D., Schiestl, R. H., Willems, A. R., Woods, R. A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 11, 355-360 (1995).
- von Kobbe, C., Thoma, N. H., Czyzewski, B. K., Pavletich, N. P., Bohr, V. A. Werner syndrome protein contains three structure specific DNA binding domains. J Biol Chem. 278, 52997-53006 (2003).

{kind=link}
{kind=link}
{kind=link}
{kind=link}