

Summary
Des études génétiques chez la levure peut être utilisée pour étudier les fonctions cellulaires et moléculaires de gènes humains dans le métabolisme de l'ADN cellulaire. Les méthodes sont décrites pour la caractérisation génétique de l'humain
Abstract
Comprendre les rôles des protéines humaines dans les voies de réparation de l'ADN génétique est un formidable défi pour beaucoup de chercheurs. Des études génétiques dans les systèmes mammifères ont été limités en raison du manque d'outils disponibles tels que définis mutant génétique des lignées cellulaires, les systèmes de régulation de l'expression, et des marqueurs de sélection appropriés. Pour contourner ces difficultés, les systèmes de modèle génétique chez les eucaryotes inférieurs sont devenus un choix attrayant pour l'étude des protéines fonctionnellement conservé réparation de l'ADN et les voies. Nous avons développé un système de levure modèle pour étudier les fonctions mal définies génétique du syndrome de Werner hélicase-nucléase (
Protocol
1. Les souches de levure
Les souches de type sauvage SGS1 TOP3 (WT; W303-1A, génotype, MAT a ade2-1 canl-100 his3-11, 15 leu2-3, 112 trpl-l ura3-1) [2], un mutant sgs1 (W1292-3C ; MAT génotype d'une SUP4-o:: URA3 sgs1-25 ade2-1 CAN1-100 his3-11, 15 leu2-3, 112 trp1-1 ura3-1 rAd5-535) et un sgs1 top3 mutant (W1058-11C, génotype, MAT une SUP4-o:: URA3 sgs1-25 top3-2:: HIS3 ade2-1 CAN1-100 his3-11, 15 leu2-3, 112 trp1-1 ura3-1 rAd5-535) ont été caractérisées [3] et ont été aimablement fournies par le Dr Rodney Rothstein (Columbia University).
2. Constructions d'ADN plasmidique
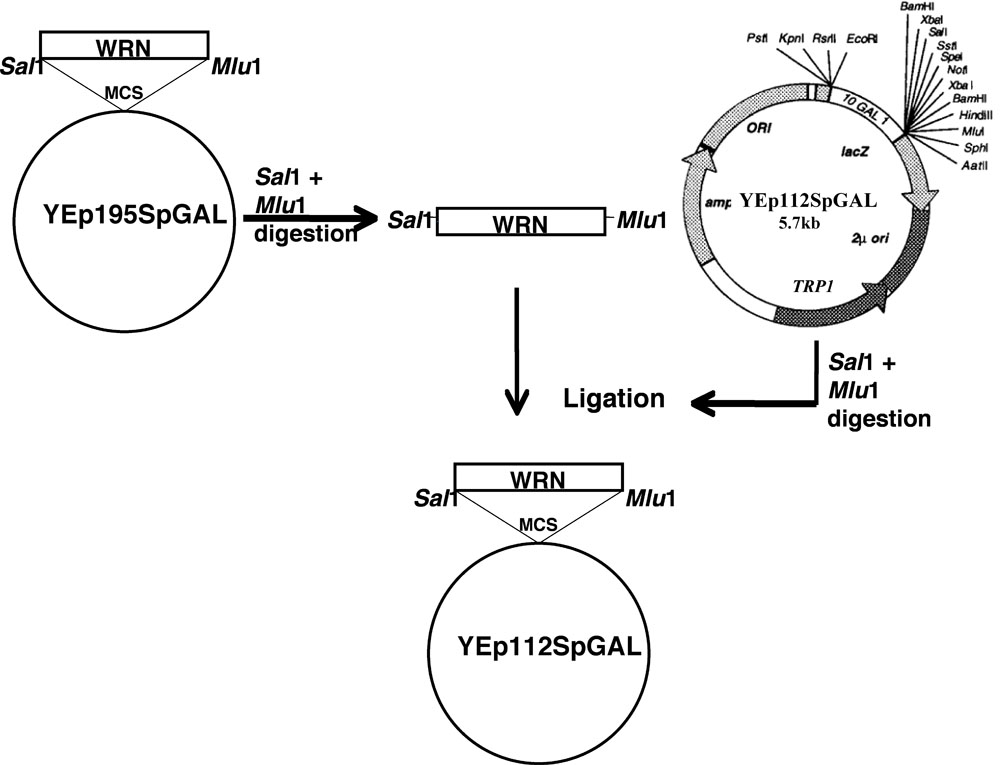
Le gène WRN a été cloné en YEp112SpGAL suivant le schéma tel que discuté.
- Site mutations dirigées sur WRN (E84A exonucléase morts, K577M hélicase morts, R834C, et K1016A) dans le plasmide YEp195SpGAL-WRN [4] ont été construits en utilisant des amorces mutagènes et un protocole standard d'Quickchange II XL kit de mutagenèse dirigée (Stratagene) par les laboratoires Lofstrand (Gaithersburg, MD).
- Fragments d'ADN codant WRN et ses variantes ont été purifiés sur gel associée de l'respectifs YEp195SpGAL-WRN construit après la double digestion avec Sal I et Mlu I.
- Gel des fragments purifiés ont ensuite été cloné dans les sites Sal I du vecteur Mlu YEp112SpGAL, un 2 um multi-copie plasmide contenant un marqueur sélectionnable pour construire TRP1 YEp112SpGAL WRN ou variantes WRN sous le contrôle d'un promoteur inductible par gallon, comme indiqué dans la figure 1 .
- Les constructions ainsi obtenues ont été désignés comme YEp112SpGAL-WRN, YEp112SpGAL-WRN E84A, YEp112SpGAL-WRN K577M, YEp112SpGAL-WRN K1016A et YEp112SpGAL-WRN R834C.
3. Transformation
Les cultures de levure ont été cultivées en utilisant le protocole standard et transformations ont été effectuées en utilisant un à base d'acétate de lithium protocole par Gietz et al [5]. En bref, les étapes suivantes ont été effectuées.
- Les cultures de souches susmentionnées ont été cultivées pendant une nuit dans 5 ml YPD (extrait de levure-peptone-glucose) à moyen terme. Les cultures ont été réinoculées dans 50 ml YPD moyenne et ont été autorisés à croître jusqu'à une DO 600 de 1,0 à 1,2 a été atteint.
- Les cultures ont été récoltées, lavées avec 20 ml d'eau, et remis en suspension dans 1 ml de 100 mM d'acétate de lithium (LiAc). Les cellules ont été incubées à 30 ° C pendant 10 min.
- Culot cellulaire a été resuspendu dans 100 mM à LiAc un volume final de 500 pl, ce qui est suffisant pour faire dix transformations. 50 ul de la suspension cellulaire a été aliquotés et culot cellulaire a été recueilli par centrifugation.
- Pour le culot cellulaire a été ajouté dans l'ordre suivant: 250 Polyéthylène glycol ul de 50% (PEG), 36 pl d'1 M LiAc, 25 pl de 2 mg / ml ADNss (ADN de sperme de saumon), 5 ml d'ADN plasmidique (100 ng 1000 ng), et 50 pi d'eau. Culot cellulaire a été vortexé pendant 1 min.
- Les cellules ont ensuite été incubées à 30 ° C pendant 30 min suivi d'un choc thermique à 42 ° C pendant 20 min.
- Les cellules ont été récoltées et remises en suspension dans 1 ml d'eau. Depuis le YEp112SpGAL plasmide contient trp comme marqueur de sélection, toutes les souches mutantes abriter la construction serait trp compétents. Par conséquent, les transformants ont été sélectionnés par étalement sur la suspension SC (synthétique complète) des médias du glucose manque Trp et incuber les plaques à 30 ° C pendant 3-4 jours jusqu'à ce que les colonies apparaissent.
4. L'analyse génétique de la restauration phénotype de croissance lente WRN transformé sgs1 top3 souche.
1. Analyse Streak:
- Pour examiner l'effet de l'expression WRN sur la croissance de type sauvage parentale (W303-1A), sgs1 ou sgs1 top3 des souches, les souches correspondantes aux YEp112SpGAL ou YEp112SpGAL-WRN ont été étalées sur des plaques LC moins Trp contenant soit 2% de glucose ( galactose glu) ou 2% (gallons). Comme contrôle, sgs1 top3 / YEp112SpGAL-SGS1 a été inclus. Les plaques ont été incubées à 30 ° C pendant 2 jours.
- Pour déterminer la capacité de WRN à affecter le taux de croissance de la souche sgs1 top3 à des niveaux inférieurs d'expression de protéines WRN, le top3 sgs1 souche transformée par YEp112SpGAL, YEp112SpGAL-WRN, ou YEp112SpGAL-SGS1 été étalées sur des plaques LC moins Trp contenant des concentrations variables de gallons par aussi bas que 0,005% à aussi haut que 2%. Les plaques ont été incubées à 30 ° C pendant 2 jours.
- Pour évaluer les besoins fonctionnels de WRN pour restaurer le phénotype de croissance lente dans le top3 des sgs1 fond, sgs1 top3 transformé avec YEp112SpGAL-WRN (E84A, K577M, R834C, et K1016A) ont été streAked sur des plaques LC moins Trp contenant soit glu ou gal aux concentrations indiquées. Pour les contrôles, sgs1 top3 souche transformée par YEp112SpGAL, YEp112SpGAL-WRN ou YEp112Sp GAL-SGS1 a été inclus. Les plaques ont été incubées à 30 ° C pendant 2 jours.
2. L'analyse de la culture liquide:
- Afin d'évaluer la croissance de la transformée sgs1 top3 souches en culture liquide, des cellules de levure ont été cultivées dans SC raf moins Trp à 30 ° C pendant la nuit. Les cultures ont été réinoculées dans SC raf moins Trp et cultivées à 30 ° C à la phase logarithmique précoce (DO 600 de ~ 0,5). Les cultures ont été ensuite réinoculées à une DO de 0,05 dans les médias SC contenant gal 2% et la croissance a été suivie en mesurant l'absorbance à des intervalles de temps indiqués.
5. L'analyse génétique de l'hydroxyurée ou méthylméthane-sulfonate de sensibilité dans WRN transformé des souches de levure.
- Pour déterminer l'effet de l'expression de la sensibilité WRN MMS et des HU sgs1 ou sgs1 top3 des souches, les souches transformées avec YEp112SpGAL ou YEp112SpGAL-WRN ont été cultivées dans SC raf moins Trp à 30 ° C.
- Les cultures ont été réinoculées dans SC raf moins Trp et cultivées à 30 ° C à la phase logarithmique précoce (DO 600 de ~ 0,6 à 0,8).
- Dix-dilutions de ces souches ont été préparées en utilisant un milieu SC manque Trp. Partir d'une culture en croissance exponentielle, 5 x 10 6 cellules ont été prélevées et diluées en série jusqu'à 10.000 fois. Quatre microlitres de chaque dilution a été repéré sur SC plaques gal Trp minus contenant les concentrations indiquées de HU ou MMS. Les plaques ont été incubées à 30 ° C.
- Comme contrôle, de type sauvage souche parentale (W303-1A) transformées avec YEp112SpGAL ou YEp112SpGAL-WRN, ou sgs1 top3 / YEp112SpGAL-SGS1 a été traitée comme décrit ci-dessus.
6. Distribution du cycle cellulaire de WRN transformé sgs1 top3 souche.
- Pour étudier la distribution du cycle cellulaire de WRN transformé sgs1 top3 souche, des cultures de sgs1 top3 transformé avec YEp112SpGAL, YEp112SpGAL-WRN, ou YEp112SpGAL-SGS1, et le vecteur transformé de type sauvage souche parentale (W303-1A) ont été cultivées dans SC raf trp moins à 30 ° C pour la nuit.
- Les cultures ont été réinoculées à une DO de 0,05 et laisser pousser jusqu'à ce début de phase log (DO 600 de 0,5 -0,6). Expression de WRN a été induite en ajoutant gal à la concentration finale de 2%. Les cultures ont été ensuite permis de croître à 30 ° C pendant 6 h.
- Les cultures ont été ensuite traitées pour DAPI (4 ', 6-diamidino-2-phénylindole) coloration.
- Les cultures ont été récoltées par centrifugation à 5800 xg pendant 10 min à température ambiante (RT).
- Les cellules ont été lavées une fois avec 1X tampon phosphate salin (PBS) et ont ensuite été fixés dans l'éthanol 70% pendant 20 min à température ambiante.
- Les cellules fixées ont été lavées deux fois avec PBS 1 X, et enfin remis en suspension dans du PBS 1X.
- Les cellules ont été placées sur des diapositives et monté dans Vectashield milieu de montage avec DAPI (1 ug / ml) (Vector, USA). Les cellules ont été examinés avec un microscope Axiovert 200 M (Zeiss; objectif 100x) et composite contraste d'interférence différentiel (DIC). Images de fluorescence ont été analysées à l'aide AxioVision, la version 3.0 du programme (Zeiss).
7. Analyses par Western blot.
1. Pour déterminer l'expression des protéines de WRN ou des variantes de WRN sgs1 transformés ou sgs1 top3 des souches, des lysats ont été préparés par la méthode suivante et les protéines analysées par immunblotting.
- Transformé sgs1 ou sgs1 top3 souches ont été cultivées dans SC raf moins Trp à 30 ° C pour la nuit.
- Les cultures ont été réinoculées et incubées à 30 ° C jusqu'à la phase logarithmique précoce (DO 600 de 0,5 - 0,8) et ensuite induite par l'ajout gal à la concentration finale de 2%. Les cultures ont été ensuite permis de croître à 30 ° C pendant 6 h.
- Cellules (3 ml) ont été collectées par centrifugation et lavées avec du PBS. Culot cellulaire a été resuspendu dans un tampon de lyse alcaline [NaOH 50 mM, pH 10,5, 2 mM EDTA, 1 mM de fluorure de phénylméthylsulfonyle (PMSF), 2% SDS, 10% glycérol, 5% 2-mercaptoéthanol et les inhibiteurs de protéase (Roche Molecular Biochemicals)] , bouillir pendant 5 min, clarifiée par centrifugation, et neutralisé avec 1 M HCl.
- Les protéines de quantités équivalentes de lysat cellulaire ont été résolus sur 8-16% des gels de SDS polyacrylamide.
2. Pour déterminer le niveau d'expression de la protéine WRN sous l'influence de différentes concentrations de galactose, les cultures ont été traitées comme suit.
- Transformé sgs1 top3 souches ont été cultivées dans SC raf moins Trp à 30 ° C pour la nuit.
- Les cultures ont été réinoculées et incubées à 30 ° C jusqu'à la phase logarithmique précoce (DO 600 de 0,5 - 0,8) et ensuite induite en ajoutant gal à une concentration finale de 2%. Les cultures ont été ensuite permis de croître à 30 ° C pendant 6 h.
- Les cellules ont été récoltées par centrifugation et remises en suspension dans le tampon (50 mM Tris-Cl, pH 7,5, le désoxycholate de sodium à 1%, 1% Triton X-100, le sulfate de sodium à 0,1% dodécyl [SDS]) contenant des inhibiteurs de protéase (0,05% PMSF, 0,05 pg / ml de leupeptine).
- Les perles de verre (425 à 600 um) ont été ajoutés au mélange, et les cellules ont été brisés par vortex.
- L'homogénat résultant a été centrifugé pendant 5 min à 15000 xg et le surnageant a été recueilli.
- Protéines dans les fractions de lysat a été estimée par la méthode de Bradford.
- Concentration égale de protéines à partir de fractions différentes lysat a été résolu sur 8-16% des gels de SDS polyacrylamide.
3. L'expression de protéines mutantes ou de WRN WRN a été déterminée par Western blot utilisant un anticorps monoclonal de souris WRN dirigé contre un épitope dans un purifiée fragment C-terminal du WRN [6] (1:1000, printemps Labs Valley).
4. D'incubation anticorps primaire a été suivie d'une incubation avec un secondaire (la peroxydase de raifort conjuguée) antibody (1:5000). Les transferts ont été traitées avec un système de détection ECL Plus l'Ouest par le protocole du fabricant.
5. Pour l'analyse quantitative blot de l'Ouest, l'augmentation des concentrations de recombinant purifié étiquette His protéines WRN pleine longueur ont été inclus sur des gels tels que, lors de la quantification de la tache d'une courbe linéaire standard pourraient être générés. En outre, l'extrait de cellule à partir de quantité équivalente de cellules ou de 20 ug d'échantillons lysat de levure ont été chargés sur des gels d'estimer la concentration de WRN.
6. Analyse ImageQuant a ensuite été réalisée sur des gels et des standards courbe linéaire a été généré.
7. La concentration de la protéine WRN à des concentrations variant gal a été estimée en utilisant la norme courbe linéaire généré ci-dessus.
8. Les résultats représentatifs
Toutes les souches utilisées dans cette étude sont auxotrophes Trp. Par conséquent, sgs1, sgs1 top3 et de type sauvage W303-1A souches transformées avec YEp112SpGAL ou YEp112SpGAL-WRN ont été sélectionnés sur la base de leur capacité à croître en présence de SC moins Trp médias. Pour examiner l'effet de l'expression WRN sur la croissance de transformer sgs1 top3 des cellules mutantes, les cultures ont été étalées sur des plaques contenant des LC moins Trp médias gal 2% pour induire l'expression WRN. Top3 decatenates entrelacés molécules d'ADN générés par Sgs1 hélicase lors de la réplication [1, 2], par conséquent, en l'absence de Top3, des contraintes de torsion n'est pas soulagée résultant en une croissance lente et d'hyper-recombinaison. La fonction génétique de sgs1 est de supprimer le phénotype de croissance lente d'un mutant top3. Si WRN pouvait se substituer à SGS1 en interaction génétique avec Top3, la restauration de top3 phénotype de croissance lente sgs1 top3 serait attendu.
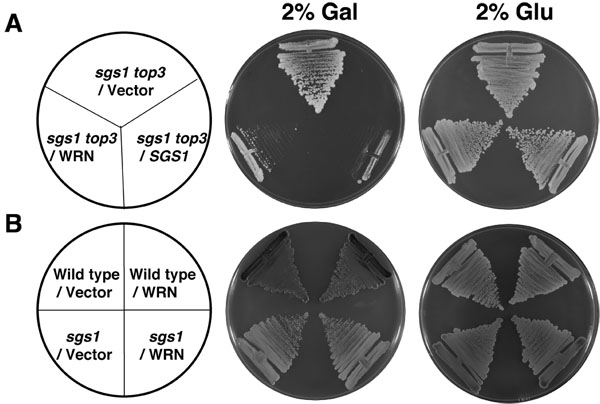
Comme le montre la figure 2A, le WRN transformé sgs1 top3 souche a considérablement augmenté plus lentement par rapport au vecteur transformé sgs1 top3 souche. Sgs1 top3 transformées avec le plasmide YEp112SpGAL-SGS1 a été incluse comme un contrôle positif (figure 2A), démontrant ainsi que du type sauvage Sgs1 exprimé chez le mutant sgs1 top3 été en mesure de compléter le phénotype génétiquement croissance. Le sgs1 transformé le top3 des souches de la même grandi en l'absence de gal comme le montre pendant 2 jours (figure 2A). L'expression de WRN n'a eu aucun effet sur la croissance des souches parentales de type sauvage (W3031A) ou sgs1 (figure 2B), indiquant que l'effet de WRN sur la croissance cellulaire était spécifique à celle observée dans le top3 des sgs1 contexte mutant. Les études génétiques effectuées en utilisant des variantes WRN démontré que WRN hélicase, mais pas d'activité exonucléase a été nécessaire pour la restauration du phénotype de croissance top3 (figure 3B). Un polymorphisme faux-sens naturellement dans WRN qui interfère avec l'activité hélicase abolie sa capacité à restaurer le top3 phénotype de croissance lente.
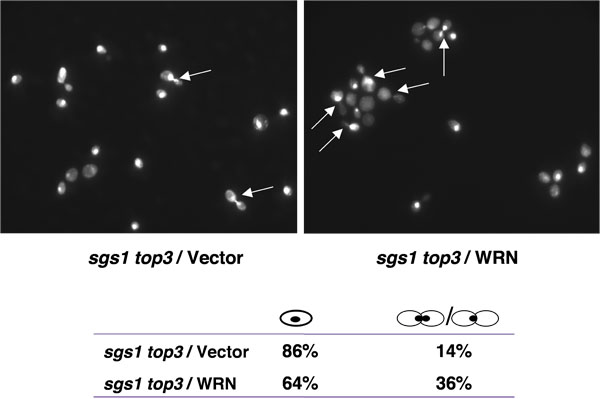
top3 des souches mutantes sont retardés dans la phase S/G2 fin du cycle cellulaire [1], une caractéristique qui peut rendre compte de leur croissance lente. La mutation du gène SGS1 dans le fond top3 supprime le retard dans la phase S/G2 du cycle cellulaire. Si l'expression WRN pourrait rétablir le retard dans la phase S/G2 du cycle cellulaire en sgs1 top3, une population élevée de grandes cellules avec des noyaux bourgeonné indivise serait attendu pour le top3 sgs1 cellules mutantes exprimant WRN. Comme le montre la figure 4, le pourcentage de grandes cellules bourgeonné était plus élevé pour le top3 sgs1 / WRN que sgs1 top3 / vecteur, suggérant la restauration de la de S/G2 jeter caractéristique du top3.
Le sgs1 top3 double mutant est moins sensible aux MMS ou HU que le mutant top3 unique [2]. Comme la croissance des cellules affectées WRN sgs1 top3, nous a ensuite examiné son effet sur la sensibilité à sulfonate méthylméthane (MMS, un agent alkylant) et l'hydroxyurée (HU, un inhibiteur de la réplication). Sgs1 top3 / WRN sensibilité affichée à la fois les médicaments comparables à sgs1 top3 / SGS1 (figure 5). L'analyse génétique des variantes WRN a révélé que WRN hélicase / ATPase, mais pas d'activité exonucléase WRN, était nécessaire pour son effet sur la sensibilité aux MMS et HU.

Figure 1. Présentation schématique pour le clonage WRN / WRN variantes dans le vecteur YEp112SpGAL. S'il vous plaît cliquez ici pour une version agrandie de la figure 1.

Figure 2. Expression de WRN sgs1 top3 restaure le phénotype de croissance lente de top3. Panel A, sgs1 top3 souche transformée par YEp112SpGAL ou YEp112SpGAL WRN ont été striés sur une plaque SC-Trp contenant soit 2% glu ou gal 2%. Comme un contrôle sgs1 top3 souche transformée par YEp112SpGAL SGS1 était striée sur les deux plaques. Les plaques ont été incubées à 30 ° C pendant 2 jours et ensuite photographiées. Groupe B, la souche de type sauvage parentale W303-1A ou de la souche sgs1 transformé avec YEp112SpGAL ou YEp112SpGAL WRN ont été striés sur une plaque SC-Trp soit contenant 2% de glu ou gal 2% . Les plaques ont été incubées à 30 ° C pendant 4 jours et ensuite photographiées. Composition des plaques a été que dans le Panneau de A et B, respectivement Panel. S'il vous plaît cliquez ici pour voir une version agrandie de la figure 2.

Figure 3. WRN ATPase / hélicase, mais pas d'activité exonucléase, est nécessaire pour restaurer le phénotype de croissance lente de top3 dans le top3 sgs1 fond. Sgs1 top3 souche transformée par l'ATPase / hélicase-morts (WRN YEp195SpGAL K577M), exonucléase-morts (YEp195SpGAL- WRN E84A), RQC mutant (YEp195SpGAL-WRN K1016A), ou polymorphes mutants (YEp195SpGAL-WRN R834C) a été rayée sur le SC-Trp plaques contenant soit 2% glu (Groupe C) ou 2% Gal (Groupe B). Les plaques ont été incubées à 30 ° C pendant 2 jours et ensuite photographiées. Composition des plaques a été que dans le Panneau de A. S'il vous plaît cliquez ici pour voir une version agrandie de la figure 3.

Figure 4. Expression de WRN induit S/G2 arrestation en sgs1 top3 cellules. Cultures logarithmique croissante de sgs1 top3 souche transformée par YEp112SpGAL, YEp112SpGAL WRN, ou YEp112SpGAL SGS1, et le vecteur transformé souche sauvage parentale ont été induites à la concentration gal 2% pendant 6 h. Les cultures ont été récoltées, transformées pour la coloration DAPI comme décrit dans Matériels et Méthodes et ont été observées en utilisant un microscope Axiovert 200 M (Zeiss; objectif 100x). Montré est la coloration DAPI du top3 sgs1 transformé avec YEp112SpGAL (en haut à gauche) et avec YEp112SpGAL WRN (en haut à droite). Les flèches indiquent les cellules avec des noyaux indivise. Répartition des cellules en G1 (cellules isolées) et S/G2 (cellules bourgeonné) est indiqué en bas du panneau. S'il vous plaît cliquez ici pour voir une version agrandie de la figure 4.

Figure 5. Effet de l'expression de la sensibilité WRN MMS et des HU sgs1 top3 souche. Logarithmique croissante des cultures de sgs1 top3 souche transformée par YEp112SpGAL, YEp112SpGAL WRN, exonucléase-morts (YEp112SpGAL-E84A WRN), l'ATPase / hélicase-morts (WRN YEp112SpGAL K577M), RQC mutant (YEp112SpGAL-WRN K1016A), polymorphes mutants (YEp112SpGAL-WRN R834C), YEp112SpGAL SGS1 et vecteur transformé de type sauvage des souches parentales ont été repérés dans un dilutions dix fois de série sur SC-Trp plaques contenant Glu ou gal et soit MMS ou HU aux concentrations indiquées. Les plaques ont été incubées à 30 ° C pendant 2 jours (plaques de contrôle) et 4 jours (MMS et des plaques HU) et ensuite photographiées. S'il vous plaît9/1639fig5.jpg "> cliquez ici pour voir une version agrandie de la figure 5.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Une des forces de l'utilisation de la levure comme système modèle est la disponibilité de mutants dans la réplication d'ADN définies et les voies de réparation qui sont conservés entre les levures et les humains. En outre la sélection des transformants abritant les gènes spécifiques est facile et fiable que les souches de laboratoire sont des mutants auxotrophes et les vecteurs avec des marqueurs auxotrophes sont facilement disponibles. L'utilisation de ces vecteurs de l'expression des produits géniques peuvent être réglementées en les plaçant sous le contrôle d'un promoteur inductible (par exemple, promoteur inductible gal, etc.) Compte tenu de ces avantages, nous avons développé un système modèle à base de levure pour étudier les besoins fonctionnels du gène WRN humaine défectueux dans le syndrome de Werner dans une voie qui est potentiellement conservée entre l'homme et la levure. En utilisant les approches décrites, nous avons fourni la première preuve que WRN peut fonctionner dans une voie génétique qui affecte le top3 des phénotypes liés. Nos observations prompte enquête plus approfondie sur la possibilité que WRN interagit fonctionnellement avec Top3α dans les cellules humaines lors de la réplication d'ADN cellulaire ou de la recombinaison. Il est concevable que, dans un état BLM-affaiblies, WRN peut partiellement se substituer à BLM grâce à son partenariat avec une protéine de la topoisomérase.
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
Ce travail a été soutenu dans son intégralité par le programme de recherche intra-muros de l'Institut des NIH, National sur le Vieillissement. Nous remercions le Dr Rodney Rothstein (Columbia University) pour les souches de levure et le Dr Brad Johnson (University of Pennsylvania School of Medicine, Philadelphie, Pennsylvanie) pour l'expression SGS1 plasmide.
References
- Aggarwal, M., Brosh, R. M. Jr WRN helicase defective in the premature aging disorder Werner Syndrome genetically interacts with Topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3. Aging. 1, 219-233 (2009).
- Gangloff, S., McDonald, J. P., Bendixen, C., Arthur, L., Rothstein, R. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol Cell Biol. 14, 8391-8398 (1994).
- Shor, E., Gangloff, S., Wagner, M., Weinstein, J., Price, G., Rothstein, R. Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae. Genetics. 162, 647-662 (2002).
- Sharma, S., Sommers, J. A., Brosh, R. M. Jr In vivo function of the conserved non-catalytic domain of Werner syndrome helicase in DNA replication. Hum Mol Genet. 13, 2247-2261 (2004).
- Gietz, R. D., Schiestl, R. H., Willems, A. R., Woods, R. A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 11, 355-360 (1995).
- von Kobbe, C., Thoma, N. H., Czyzewski, B. K., Pavletich, N. P., Bohr, V. A. Werner syndrome protein contains three structure specific DNA binding domains. J Biol Chem. 278, 52997-53006 (2003).

{kind=link}
{kind=link}
{kind=link}
{kind=link}