

Summary
신화는 모델 유기체 Saccharomyces cerevisiae의 표현 단백질 사이의 과도 안정적인 상호 작용의 민감한 감지 수 있습니다. 그것은 성공적으로 높은 처리량 방법으로 상호 작용하는 파트너를 식별하기 위해 외인성 및 효모 필수 막 단백질 연구에 적용되었습니다.
Abstract
통합 멤브레인 단백질의 기본적인 생물 학적 및 임상 중요성은 전체 길이 transmembrane 단백질에 대한 단백질 - 단백질 상호 작용 (PPI)의 높은 처리량을 확인을위한 효모 기반 시스템의 개발을하라는 메시지가 나타납니다. 이를 위해 우리 실험실 분할 - ubiquitin 기반 막 효모 두 하이브리드 (신화) 시스템을 개발했습니다. 이 기술을 사용 일시적 안정 단백질 상호 작용의 민감한 검출을 허용
Protocol
1. 참고 자료
단백질 - 단백질 상호 작용 (PPIs)는 모든 세포 공정을 적용에 관련된 기본 빌딩 블록입니다. 따라서, 그것은 모든 상호 작용이 긴밀이 생물 학적 균형의 변화는 일반적으로 질환과 암 세포 변환의 역할을 같이 세포 항상성을 유지하기 위해 규제하는 것이 필수적입니다. 그들은 복잡한 신호 폭포를 시작하고, 관련된 문제와 같은 의료 분야의 최근 의의되었습니다 의약품 등 다양한 분자의 수출입을 모두 중재 수있는 멤브레인 관련 단백질은 단백질의 가장 생물학적으로 중요한 클래스 사이 아르 약물 저항은 점점 일반지고 있습니다. 이 단백질 클래스의 복잡도에 대한 통찰력을 확보하는 것은 그들의 상호 작용하는 파트너의 확인이 필요합니다. 같은 파트너를 발견하는 것은 도전 입증되었습니다, 자주 그것이 각 막 - 바운드 단백질에 최적화된해야 거친 조건을 필요로 [1].
통합 멤브레인 단백질의 기본적인 생물 학적 및 임상 중요성은 전체 길이 transmembrane 단백질에 대한 PPI의 높은 처리량을 확인을위한 효모 기반 시스템의 개발을하라는 메시지가 나타납니다. 이를 위해, 우리는 분할 - ubiquitin 기반 막 효모 두 하이브리드 (신화) 시스템 [2-4]을 개발했습니다. 이 도구는 일시적인 안정 단백질 상호 작용의 민감한 감지 수 있습니다. 그것은 성공적으로 모델 생물 Saccharomyces cerevisiae의 표현 외인성 및 내생 단백질 [3-7]. 연구에 적용되었습니다 신화는 ubiquitin 두 moieties로 분리 수있는 관찰의 활용 :. C - 터미널 절반 (C UB)과 N - 터미널 절반 (NubI) 생체내 연구에 표시된 것을 위해 높은 친화력으로 인해 저절로 reconstitute이 moieties 한 다른 (Figure. 1A). 그러나, ubiquitin의 N - 터미널 절반으로 글리신 포인트 돌연변이에 이소 류신 13 소개하면 (N UB G로 불리는 조각을 생산) 방지이 자연 재 협회 [8] (Figure.의 1B).
우리는 신화 시스템 (Figure. 1C 2)에서이 원칙을 사용합니다. 간단히, 통합 멤브레인 미끼 단백질은 대장균 DNA 결합 단백질 LexA 구성된 인공 전사 인자와 헤르페스 심플 렉스 바이러스 VP16의 정품 인증 도메인에 연결된 C UB의 잔기에 융합이다. 할까요은 NubG 잔기로 cDNA 또는 게놈 DNA 파생된 조각의 융합에 의해 생성됩니다. 효모 호스트의 미끼 먹이와 단백질 사이의 상호 작용은 cytosolic deubiquitinating 효소 (덥스) 및 전사 인자의 proteolytic 릴리스에 의해 이후의 인식과 함께, 전체 길이 '의사 ubiquitin'분자의 reconstitution로 연결됩니다. 전사 인자는 다음 세포의 핵을 입력하고 미끼와 먹이 상호 작용을 나타내는 것입니다 선택적 미디어에 효모 종자의 성장을 허용하는 리포터 유전자 시스템 (일반적으로 HIS3, lacZ 및 ADE2 유전자의 표현을 포함하는), 활성화할 수 있습니다 [ 2-4].
통합 멤브레인 단백질 상호 작용의 식별 및 특성은 완전히 그들의 기능을 이해할 수 있도록 정보를 제공합니다. 우리가보다 정확하게 이해하고 필수적인 멤브레인 단백질과 상호 작용하는 단백질의 역할을 해부하다, 우리는이 단백질을 조절에 관련된 동적 상호 작용에 대한 통찰력을 얻고 치료 가능성이있을 수 있습니다 소설 대상을 발견할 수 있습니다.
2. 미끼 및 적절한 신화 시스템의 선택
- 이전 신화 분석을 수행하는 단백질은 세포의 cytosol에서의 N - 및 / 또는 C - 말단을 가지고 있는지 확인하십시오. 그것은 전사 인자 릴리스에 필요한 deubiquitinating 효소가 cytosol에있는 이후 C UB - LexA - VP16 태그가 같은 터미 너스에서 단백질을 융합하는 것이 필수적이다 [4].
- 다음 적합 신화의 두 가지 주요 변종 중 어떤 결정합니다. 비 - 네이티브 효모 단백질에 대한 전통적인 신화 (tMYTH)이 baits가 플라스미드로부터 ectopically overexpressed 아르 궁극적으로 사용할 수 있습니다. 기본 효모 단백질에 대한 통합 신화 (iMYTH)는 선택의 방법입니다. iMYTH에서 baits은 endogenously들은 기본 발기인의 통제하에 그들을 떠나, C UB - LexA - VP16 태그가 추가됩니다. 이것은 baits의 야생 형 표현 수준이 이러한 잘못된 반응의 증가 숫자 [3]과 같은 단백질 overexpression와 관련된 문제를 제거하는 데 도움이 같은 유리한 것입니다. 초기 미끼 건설 및 사용 선택적 매체를 제외하고, 신화의 두 형태는 본질적으로 동일한 방식으로 수행됩니다. 지혜로움에 대해, 우리는이 변종은, 원칙적으로, 거의 모든 유기체에서 막 단백질과 함께 사용하고 있습니다 수 있듯이, 주로이 보고서에 tMYTH의 사용에 초점을 맞출 것이다따라서 더 널리 적용.
3. 베이트 생성 및 검증
- 필요한 미디어 및 솔루션
- 무균 ddH 2 O는 121 ° C에서 30 분 15 Psi를 autoclaving하여 준비 하였다.
- 3 - 아미노 - 1 ,2,4 - triazole (3 - AT) ddH에서 1M 주식 솔루션으로 준비 방법 2 O. 0.2 μm의 필터를 통해 통과하여 소독.
- 1% W / V 효모 추출물, 2% W / V 펩톤, 2퍼센트 W / V 포도당과 ddH 2 O.에서 준비 100 μm의의 아데닌으로 구성된 YPAD 성장 미디어 121 ° C, 30 분 15 PSI에 autoclaving하여 소독.
- 10X 아미노산 / 염기 자료 믹스 : 전체 믹스 1.0 MM 아데닌, 1.8 당연 유라실, 1.0 MM 아르기닌, 1.0 당연 히스티딘 2.3 MM 이소 류신, 7.6 MM 류신, 1.6 MM 라이신, 메티오닌 10.1 MM, 3.0 MM 페닐알라닌, 16.8 MM를 포함 트레오닌, 2.0 MM 트립토판, 1.7 MM의 티로신과 12.8 MM의 발린은 ddH 2 O에서 준비했습니다. 에 대한 드롭 아웃이 필요한 아미노산 (들) 및 / 또는 뉴클레오 티드베이스 (들)를 생략. 121 ° C, 30 분 15 PSI에 autoclaving하여 소독.
- 합성 드롭 아웃 (SD) 0.67 % W / V 효모 질소베이스 (아미노산 않고 있지만 암모늄 황산 포함), W / V 포도당 2 % 2w / V 한천 %와 1X 아미노산 / 염기 혼합으로 구성된 성장 미디어 ddH 2 O.에서 준비 두 액체와 고체 (2 % 한천을 포함) SD - 류신 미디어를 준비합니다. 또한 고체 SD - 트립토판 - 류신과 SD - 트립토판 - 류신 - 아데닌 - 히스티딘 미디어를 준비합니다. 121 ° C, 30 분 15 PSI에 autoclaving하여 소독. 100x15 mm 배양 접시에 고체 미디어를 하거라.
- 3 - AT를 포함 합성 드롭 아웃 (SD) 성장 미디어. 설명한대로 SD - 트립토판 - 류신 - 아데닌 - 히스티딘 미디어를 준비하지만, 25, 50, 75 및 100 mm의 농도 3 - AT 포함. 의 적절한 금액을 추가 3 - AT가 (그러나 아직 확정되지 않은) autoclaved 및 냉각 후 언론에 1M 멸균 주식 솔루션에서. 100x15 mm 배양 접시에 부어.
- 40 %로 구성된 PEG / 리튬 아세테이트 혼합 W / V PEG - 3350, 120 MM의 리튬 아세테이트 및 167 μg / ddH 2 O.에서 준비 ML 살몽 정자 DNA (종류 III 나트륨 소금) 이 혼합물의 불임은 멸균 물 및 솔루션에서 준비를 보장하기 위해 (즉, 50 % PEG - 3350을 autoclaved 살균 ddH 2 O에서 준비 1M 리튬 아세테이트 2 MG / ML 연어 정자 DNA 유형 III의 나트륨 소금 autoclaved).
- 효소와 PCR를 실시를위한 시약.
- 상업 miniprep 키트.
- 소다 석회 유리 비즈 (0.5 mm).
- 플라스미드 전파 (예 : DH5α, XL10 골드) 및 박테리아 번식 및 플라스미드 선택에 적합한 표준 미디어에 적합한 능력 대장균 세포.
- 특정 효모 종자로 프로토콜에 설명된 plasmids와 primers.
- 갑 복구에 의해 tMYTH Baits의 생성
- 미끼는 태그와 표현에 적합한 벡터로 복제해야합니다. tMYTH 벡터의 다양한 미끼 건설에 사용하기 위해 현재 수 있습니다. 같은 pCMBV4, pAMBV4 및 pTMBV4 같은 벡터 허용 제어 각각 CYC1 (약한), ADH1 (강한)과 TEF1 (강한) 발기인, 아래의 C - 말기 태그 baits 건설 (미끼 - C UB - LexA - VP16) . N - 말기 태그 baits (LexA - VP16 - C UB - 미끼)가 같은 각각 TEF1 및 CYC1 발기인의 통제를 받고 pTLB - 1 pBT3 - N, 같은 벡터를 사용하여 생성할 수 있습니다. 벡터 선택 미끼에 따라 차이가 있으며, 경험적으로 결정되어야합니다. 다른 경우 베이트 overexpression 실제로 잘못된 반응의 증가 숫자로 이어지는, 해로운 수 있습니다 반면 어떤 경우에는 높은 미끼 표현은 상호 작용을 검출하기 위해 필요합니다.
- 제한은 적절한 제한 사이트 (S)에서 선택한 플라스미드를 소화. 절단은 C UB - LexA - VP16 태그 (업스트림 C - 터미널 태그 또는 N - 터미널 태그에 대한 하류에 대한 태그의)의 인근에서만 발생한다. 예를 들어, pAMBV4 벡터를 사용하는 경우, SfiI는 이상적인 선택입니다. 사용을위한 준비까지 -20 ° C에서 소화 플라스미드를 저장합니다.
- 관심의 유전자의 증폭 및 복제에 대한 설계 primers. 최종 목표 유전자의 첫 번째 18-20 세포핵과 일치해야합니다 5 '3 동안 전달 프리머의 끝은 제한 사이트의 상류 약 35-40 세포핵과 일치해야'을 선택하십시오. 5 최종 대상 유전자의 마지막 18-20 세포핵의 반대로 보완을 (일치 '리버스 프리머의 끝 3, 제한 사이트의 하류 약 35-40 세포핵의 반대로 보완과 일치해야합니다'는 정지 코돈을 생략 경우 C UB - LexA - VP16 태그)은 C - 말단에 배치되고있다. 여부에 따라 N - 또는 C - 터미널 태그가 수행되고, 대상 유전자가 C UB - LexA - VP16 태그로 프레임에 복제되었는지 등 앞으로 또는 역방향 프라이머의 35-40 세포핵을 선택합니다.
- 위의 primers를 사용하여 PCR하여 관심있는 유전자를 증폭. PCR 매개 변수는 특정 효소와 사용되는 특정 primers에 따라 달라집니다.
- leu2 돌연변이 (예 : BY4741)을 베어링 적절한 효모 실험실 스트레인에 소화 플라스미드와 함께 PCR 제품을 변환. 신화의 기자 스트레인 (예 : THY.AP4 또는 L40)를 사용하지만, 필요하지 않습니다,이 시점에서 효모의 목적으로 간격 복구가 상동 재조합이 발생할 수있는 환경으로서 단순히 수있다. 변환은 다음과 같이 진행되어야합니다
- 무균 YPAD 미디어의 5 ML로 선택한 효모 변형의 단일 식민지를 예방하고 30 밤새 품어 ° C 일정한 잡고 (200 RPM).
- ~ 0.15의 OD600 신선한 YPAD 미디어 50 ML에 야간 문화를 희석하고 30 ° C를 품어 (200 RPM) 잡고. ~ 0.6의 OD600에 도달할 때까지 약 3~4시간에 대한 성장.
- 5 분 700xg에서 세포를 원심하여 표면에 뜨는를 제거합니다.
- 세포 25 ML 살균 ddH 2 O의 펠릿 5 분 700xg에서 원심 분리기를 Resuspend.
- 뜨는을 제거하고 1 ML 살균 ddH 2 O.에서 세포 펠릿을 resuspend
- microfuge 튜브에 세포 100 μL, 300 μL PEG / 리튬 아세테이트 믹스, 그리고 소화 플라스미드 (50 fmol) 및 PCR 제품 (250-500 fmol) 추가합니다.
- 30 분 30 ° C에서 알을 품다.
- 1 시간 42 ° C에서 열 충격.
- 5 분 3000xg에서 원심 분리기 및 뜨는을 제거하십시오.
- 세포 200 μL 살균 ddH 2 O의 펠렛과 접시 고체, SD - 류신 선택적 미디어에 전체 볼륨을 Resuspend. 2-4일 30 ° C에서 성장.
- 30 ° C 하루에서 5 ML SD - 류신 액체 미디어의 변형 변형의 단일 식민지 철 좀 들어라.
- 5 분 700xg에 세포를 원심 분리기 및 뜨는을 제거하십시오.
- 상업적 miniprep 키트를 사용하여 셀 여럿 미끼 플라스미드 DNA를 분리. 하나 수정과 표준 프로토콜을 따르십시오. 충분한 효모 세포 용해를 보장하기 위해 5 분 동안 적극적으로 초기 resuspension과 와동 후 펠렛으로 0.5 mm 소다 석회 유리 구슬의 작은 볼륨을 추가합니다. 그러면 정상적으로 상용 프로토콜 진행합니다.
- 관할 E.으로 분리된 효모의 DNA를 변형 대장균은 적어도 1x10 7 셀 / μg의 DNA의 변환 효율 플라스미드 전파 (예 : DH5α, XL10 골드)에 적합한 변형. 미끼 plasmids이 kanamycin를 사용하기위한 선택하실 수 있습니다.
- 변형 E.에서 수확 플라스미드 DNA 표준 DNA 분리 방법 또는 상업적 키트를 사용하여 대장균.
- 순서에 의해 플라스미드 미끼의 적절한 구조를 확인합니다.
- 검증된 미끼가 적절한 신화의 기자 스트레인 (예 : THY.AP4, L40)로 구성 변환. 방금 설명한 효모 변환 프로토콜은 소화 플라스미드 및 PCR 제품의 대신에 미끼 플라스미드 DNA를 대체, 사용할 수 있습니다.
- 베이트 확인 - 적절한 현지화
- 사용하기 전에, 미끼 종자들은 제대로 효모 막에 현지되는 것을 보장하기 위해 분석됩니다. iMYTH을 사용하는 경우,이 지방화는 태그 미끼의 속성에 따라 구체적으로 따라 달라집니다. tMYTH 들어, 미끼 plasmids은 일반적으로 신호 순서 (예 : Matα) 플라즈마 막에 표현 단백질을 연출.을 포함 현지화은 형광 현미경을 사용하여 결정됩니다. 미끼 태그 순서에 YFP 분자의 포함은 (예 : C UB - YFP - LexA - VP16) 라이브 세포의 직접적인 시각화를 허용, 가장 간단한 가장 직접적인 방법이며, 일반적으로 iMYTH에서 사용됩니다. 또는 LexA 또는 태그의 VP16 구성 요소에 대한 항체를 사용하여 표준 immunofluorescence 방식을 사용할 수 있습니다.
- 베이트 유효성 검사 - N UB G / 제가 테스트 제어
- 미끼의 일단 적절한 현지화가 설립되었습니다, 그것은 미끼가 혼자가 아닌 상호 작용할까요 (예 : 미끼 자기 활성화 있지 않은지 확인합니다)의 존재에있는 기자 시스템을 활성화하지 않도록하는 것이 필요합니다. 이것은 미끼가 상호 작용 (긍정)이 아닌 상호 작용 (음수) 제어할까요로 변환하고, 성장이 선택적 미디어에 부과되는 N UB G / I 시험을 사용하여 수행됩니다. 미끼는 선택적 미디어에 성장해야합니다 IN 긍정적인 컨트롤의 존재, 그리고 신화에 사용하기 위해 적합하기 위하여 대조군의 존재 성장하지 않습니다.
- 플라스미드 제어 먹이의 100-200 NG와 테스트하기 위해 미끼 부담을 변환하여 시작합니다. 이전에 설명한 효모 변환 프로토콜 YPAD 대신에 SD - 류신 미디어를 대체하고 최종 도금 단계에 SD - 트립토판 - 류신 미디어를 사용하여 사용할 수 있습니다. 다음 컨트롤 먹이 구조는 일반적으로 사용됩니다 :
- pOST1 - N UB I (긍정적인 제어)
- pOST1 - N UB G (대조군)
- pFUR4 - N UB I (긍정적인 제어)
- pFUR4 - N UB G (대조군)
- OST1은 oligosaccharyl 복합의 구성 요소이며 endoplasmic reticulum 막 [9]하는 언어이다 FUR4가 유라실의 permease이며 플라즈마 막에 지역화된 상태에서 [10]. 이러한 단백질은 일반적으로이 아닌 상호 작용할까요으로 사용하기위한 우리의 첫번째 선택지만, 그들의 적합성은 케이스별로 사건에 따라 달라질 수 있습니다. 당신의 미끼가 진정으로 이러한 컨트롤 모두와 상호 작용 예측되는 가능성이 경우, 대체할까요 선택한해야합니다. N UB 전 ubiquitin의 N - 말단의 야생 타입 형태이며, 자발적 C UB와 N UB가 융합 아르에 단백질 사이의 상호 작용의 C UB 독립와 상호 작용하는 것을 기억. N UB G할까요은 (N UB 및 C UB의 자발적인 연결을 방지 글리신의 돌연변이에 이소 류신 13 운반) 부정적인 제어 역할을하는 동안 따라서 N UB I의할까요은 긍정적인 컨트롤을 구성합니다.
- 100 μL 무균 ddH 2 O.로 각각 변화 미끼의 한 식민지 Resuspend
- 순차적으로 10분의 1의 dilutions, 1 / 100 1 / 1000 생산 무균 ddH 2 O의 resuspended 세포를 희석.
- 농도 범위에서 3 - AT과 않고 SD - 트립토판 - 류신과 SD - 트립토판 - 류신 - 아데닌 - 히스티딘 미디어에 undiluted 및 희석 세포의 5 μL 볼륨 스팟. 3 - AT HIS3 리포터 유전자의 경쟁 억제제 역할을하고 선택 과정의 엄중을 높이는 역할을합니다. 그것은 약하게 알맞게 스스로 활성화 baits의 억제가 아닌 구체적인 성장을위한 경우에 유용할 수 있습니다.
- 장소는 건조하고 2-4일 30 ° C에서 접시를 품어 수 있습니다.
- 모든 transformants들은 성공적으로 플라스미드 먹이로 변형되었는지 나타내는 SD - 트립토판 - 류신 접시에서 성장한다. 하지 Baits 자기 활성화 제가 N UB G할까요와 구조를 먹이, 그리고 N UB로 변환하는 경우에만 SD - 트립토판 - 류신 - 아데닌 - 히스티딘 미디어에 성장합니다. 어떤 농도 주 3 - AT가이 검사하는 동안 사용해야하므로 (있을 경우) 미디어가 필요합니다.
4. 전형
- 필요한 미디어 및 솔루션
- 무균 ddH 2 O는 121 ° C에서 30 분 15 Psi를 autoclaving하여 준비 하였다.
- 0.9 % NaCl 솔루션 ddH 2 O의 준비 121에서 autoclaving 소독 ° C 30 분 15 PSI.
- ddH 2 O.에서 493 mm까지 나트륨 인산염 이염 250 MM 나트륨 인산염 일염기의 구성된 인산 나트륨 솔루션 121 ° C, 30 분 15 PSI에 autoclaving하여 소독.
- X - 걸 (5 브로모 -4 - 클로로 - 3 - 인돌릴 - β - D - galactopyranoside) N, N - 디메틸 포름 아미드에 100 MG / ML 주식 솔루션으로 준비 솔루션입니다.
- ddH 2 O.에서 준비 2% W / V 효모 추출물, 4% W / V 펩톤, 아데닌 4퍼센트 W / V 포도당과 100 μm의를 포함한 2xYPAD 성장 미디어 121 ° C, 30 분 15 PSI에 autoclaving하여 소독.
- 앞에서 설명한 것처럼 준비 합성 중퇴 (SD) 성장 미디어. 액체 SD - 류신와 고체 SD - 트립토판 - 류신을 준비합니다. 두 100x15 mm 배양 접시에 고체 미디어를 하거라. N UB G / I 제어 시험에서 결정된 농도 3 - AT를 포함 150mm 원형 접시, 각 화면 16 접시, 필요한 경우에 고체 SD - 트립토판 - 류신 - 아데닌 - 히스티딘 미디어를 준비합니다.
- 합성 중퇴 (SD) 미디어 + 5 - 브로모 -4 - 클로로 - 3 - Indoyl - β - D - Galactopyranoside (X - 걸.) 준비 SD - 트립토판 - 류신 - 아데닌 - 히스티딘 앞에서 설명한 것처럼 한천을 포함 미디어. autoclaving 후, 1/10th 볼륨 O 다음 (필요한 경우) 3 - AT, 멋진 추가할 수 있습니다F 멸균 인산 나트륨 솔루션입니다. 다음, 80 μg / ML의 최종 농도 X - 걸 솔루션을 추가합니다. 철저하게 믹스와 150mm 원형 접시에 부어.
- PEG / 리튬 아세테이트 솔루션 II가 40 % 포함된 PEG - 3350, 100 MM의 리튬 아세테이트, 1 MM EDTA (에틸렌 다이아 민 테트라 초산) 10 MM 트리스 산도 7.5. 무균 ddH 2 O 및 솔루션을 사용하여이 솔루션을 준비 (예 : autoclaved 50% PEG - 3350, 1 M의 리튬 아세테이트, 100 MM 트리스 산도 7.5 500 MM EDTA (에틸렌 다이아 민 테트라 초산) 산도 8.0).
- 리튬 아세테이트 / 110 MM의 리튬 아세테이트, 11 MM 트리스 산도 7.5 및 1.1 MM EDTA (에틸렌 다이아 민 테트라 초산)를 포함하는 트리스 EDTA (에틸렌 다이아 민 테트라 초산) 솔루션입니다. 무균 ddH 2 O 및 솔루션 (예 : autoclaved 1M 리튬 아세테이트, 100 MM 트리스 산도 7.5 500 MM EDTA (에틸렌 다이아 민 테트라 초산) 산도 8.0)를 사용하여이 솔루션을 준비합니다.
- 100 MM 트리스 산도 7.5 10 MM EDTA (에틸렌 다이아 민 테트라 초산)으로 구성된 10X 트리스 EDTA (에틸렌 다이아 민 테트라 초산) 솔루션 ddH 2 O.에서 준비 121 ° C, 30 분 15 PSI에 autoclaving하여 소독.
- 무균 ddH 2 O.에서 준비 2 MG / ML 살몽 정자의 DNA 유형 III 나트륨 소금을 포함하는 단일 좌초 캐리어 DNA (ssDNA) 솔루션
- 상업 miniprep 키트.
- 소다 석회 유리 비즈 (0.5 mm).
- 플라스미드 전파 (예 : DH5α, XL10 골드) 및 박테리아 번식 및 플라스미드 선택에 적합한 표준 미디어에 적합한 능력 대장균 세포.
- 같은 프로토콜에 설명된 특정 효모 종자와 plasmids
- 대규모 변환
- SD - 류신 미디어의 5 ML에 미끼를 포함하는 신화의 기자 스트레인의 단일 식민지를 예방하고 30 밤새 품어 ° C를 (200 RPM) 잡고.
- OD600 200 ML SD - 류신 미디어에 야간 문화를 희석 30 = 0.15 및 부화 ° (200 RPM) 잡고 C. OD600까지 성장 = 0.6-0.7 (약 4~5시간).
- 대상 OD600에 도달 직전, ssDNA 솔루션의 나누어지는를 녹여. 100 ° C 삶아 얼음에 냉각 후 5 분하십시오. 한 번 반복합니다.
- 대상 OD600에 도달하면, 5 분 (4x50 ML 나사 캡 원심 분리기 튜브 사이에 200 ML 문화를 나눕니다)에 대한 700xg에서 원심 분리를 통해 세포를 수확.
- 30 ML 무균 ddH 2 O와 간단히 와동 샘플을 각 펠릿 씻으십시오. 5 분 700xg에서 원심 분리기.
- 뜨는을 취소하고 1 ML의 리튬 아세테이트 / 트리스 EDTA (에틸렌 다이아 민 테트라 초산) 솔루션의 각 펠렛을 resuspend. 멸균 1.5 ML의 microfuge 관과 5 분 700xg에서 원심 분리기로 전송합니다.
- 뜨는을 취소하고 리튬 아세테이트 / 트리스 EDTA (에틸렌 다이아 민 테트라 초산) 솔루션을 600 μL의 각 펠렛을 resuspend.
- 4x15 ML 나사 캡 원심 튜브에 다음 추가 :
- 2.5 ML PEG / 리튬 아세테이트 솔루션 II
- 600 μL resuspended 세포
- 100 μL ssDNA 용액
- 먹이 라이브러리 DNA 7 μg
- N UB G와 N - 또는 C - 말단 하나의 태그와 cDNA 또는 게놈 다양한 소스로부터 준비할까요을 포함하는 라이브러리 (상업적으로 사용할 수 있습니다 www.dualsystems.com ). 사용되는 특정 라이브러리는 선택된 유인 실험 목표에 의존 사례별로, 결정되어야합니다.
- 소용돌이 1 분 튜브는 45 분 동안 30 ° C waterbath에서 부화 후 철저한 믹싱을 보장하고 있습니다. 15 분 간격으로 간략하게 섞는다.
- 각 튜브 160 μL 디메틸 sulfoxide (DMSO)를 추가하고 튜브 반전에 의해 즉시 섞는다.
- 20 분 동안 42 ° C waterbath에 품어.
- 5 분 700xg에서 원심 분리하여 transformants를 수집합니다.
- 뜨는 폐기하십시오. 3 ML의 2xYPAD 각 펠렛의 resuspension하여 transformants를 복구할 수 있습니다. 한 50 ML 나사 캡 원심 분리기 튜브 함께 풀 모든 샘플을.
- 세포 복구를 위해 90 분간 30 ° C에서 알을 품다.
- 5 분 700xg에서 원심 분리기 및 뜨는 폐기하십시오.
- 4.9 ML 무균 0.9 % NaCl에있는 세포 알약을 Resuspend.
- resuspended 세포 100 μL를 사용하면 살균까지 0.9 % NaCl의 10 배 시리얼 dilutions를 준비하기 위해 1000x 10X에서.
- 30 SD - 트립토판 - 류신 미디어와 부화에 100x와 1000x dilutions의 플레이트 100 μL ° 2-3 일동안은 C. 이 번호판은 제어 역할과 변화의 효율성을 계산하는 데 사용됩니다.
- 마찬가지로, 대형 (150 ㎜) SD - 트립토판 - 류신 - 아데닌 - 히스티딘 접시에 resuspended 세포와 접시의 나머지 4.8 ML를 나누어3-4일 30 ° C에서 필요에 따라 N UB G / I 시험에서 결정 3 - AT의 금액과 부화를 포함.
- 에 0.9 % NaCl 및 플레이트 5 μL aliquots 100 μL에 하나의 식민지 (각 나타내는 세포가 잠재적인 상호 작용 미끼 - 먹이 쌍을 포함) Resuspend SD - 트립토판 - 류신 - 아데닌 - 히스티딘 + X - 걸 미디어 (및 경우 3 - AT를 포함 필요). 2-4일에 대한 성장을 허용합니다. 이 단계는 선택적 검사의 두 번째 라운드 역할을하고, 초기 라운드에서 얻은 잘못된 반응의 제거에 도움이됩니다. 강력한 성장과 파란색을 표시만이 식민지는 추가 분석을 위해 선택됩니다.
- 프레이의 DNA의 분리 및 시퀀싱
- 이전에 설명한 수정 miniprep 프로토콜을 사용하여 파란색 효모 식민지에서 플라스미드 DNA를 분리. 뿐만 아니라, 먹이의 보존을 선택하지만, 미끼, plasmids에 SD - 트립토판 미디어에 세포를 성장해야합니다. 히트 매우 많은 수의를 생성 화면 들면, 상용 높은 처리량 miniprep 키트는이 시점에서 유리한 수 있습니다.
- 관할 E.에 고립 효모 플라스미드 DNA를 변형 대장균은 적어도 1x10 7 셀 / μg의 DNA의 변환 효율 플라스미드 전파 (예 : DH5α, XL10 골드)에 적합한 변형. 먹이의 plasmids은 암피실린를 사용하기위한 선택하실 수 있습니다.
- 변형 E.로부터 플라스미드 DNA를 분리 표준 DNA 분리 방법 또는 상업적 키트를 사용하여 대장균. 샘플 번호가 큰 경우 다시 한번, 높은 처리량 miniprep 키트는 유용할 수 있습니다. E.에있는 DNA의 증폭 대장균 크게 플라스미드 수율을 증가하고 DNA의 충분한 양의는 시퀀싱 및 추가 분석 모두에있다는 것을 보장합니다.
- N UB G. 이내 순서 보완 프라이머를 사용하여 격리 plasmids 순서
- interactors의 예비 목록을 조립하는 모든 시퀀스 데이터를 컴파일하고 분석할 수 있습니다. 이것은 수동으로 수행하거나, 적절한 소프트웨어를 사용하여 자동화된 방식으로 수 있습니다.
- 베이트 종속성 테스트
- interactors의 예비 목록 조립 후에 그것은 상호 작용을 다시 확인하고 상호 작용 / 미끼 정체성의 독립적인 방식으로 기자 시스템을 활성화 난잡한할까요을 제거하는 것이 중요합니다. 이것은 베이트 종속성 테스트를 사용하여 수행됩니다. 이 시험에서 확인된 interactors의 모든 원래 미끼 스트레인뿐만 아니라 C UB - LexA - VP16 태그에 융합 단일 transmembrane 도메인 구성된 제어 인공 미끼를 숨겨주는 부담으로 다시 변환됩니다. 변환은 YPAD와 최종 도금 단계에 SD - 트립토판 - 류신 고체 매체 대신에 SD - 류신 미디어를 사용하여 표준 프로토콜 이전에 설명한 당로 수행됩니다.
- 무균 ddH 2 O 및 현장 100 μL에 위의 변환에서 하나의 식민지를 Resuspend SD - 트립토판 - 류신 - 아데닌 - 히스티딘 + X - 걸 미디어 (및 필요한 경우 3 - AT 포함)에 5 μL 권. 접시는 다음 30 2-4일 ° C.에 대한 incubated 아르 이상적으로 여러 transformants은 각 먹이를 선정해야하고, 원래의 유인 인공 미끼 모두 같은 접시에 발견해야합니다.
- 기자 시스템 (즉, 성장과 파란색)의 원인이 활성화가 싸구려로 간주되며 특정 먹이가 interactors 목록에서 제거되는 인공 미끼와 먹이를 들고 효모.
- 미끼 - 오브 - 관심지만 인공 미끼와 효모의 성장과 청색 착색을 일으킬할까요,이 특정 상호 작용을 확인합니다. 그러나, 효모는 먹이를 숨겨와 미끼 - 오브 - 관심이 성장하지 않는 경우,이 먹이가 interactors 목록에서 제거됩니다.
- 나머지할까요은 신화 화면에서 확인 interactors의 전체 목록을 구성합니다.
5. 추가 연구
신화 심사가 완료되면, 추가 분석이 감지 상호 작용의 생물 학적 중요성을 확인하고 결정하기 위해 수행되어야합니다. 수행되어야하는 구체적인 연구 사례별로 다양하며, 개인 연구자에 의해 결정되어야합니다. 후속 작업의 몇 가지 일반적인 예는 원래 유기체 공동 immunoprecipitation 실험 및 삭제 연구를 포함합니다. 또한, 획득한 데이터의 전산 분석 패턴을 감지하고, 다양한 상호 작용 재생 수있는 잠재적인 관련 성과 역할을 식별하는 데 도움이 유용할 수 있습니다. 따라서 신화 기술은 막 단백질의 중요한 기능 상호 작용의 인식과 이해를 향해 강력한 첫 단계 '로 역할을합니다. 자세한 후속 연구 및 기타 최근 개발하고 신흥 테과 함께chnologies, 그것은 세포의 신비를 잠금 해제에 유용한 도구가 될 것을 약속드립니다.

그림 1. 분할 ubiquitin. A.의 원리 C - 터미널 절반 (C UB)과 N - 터미널 절반 (N UB I) : Ubiquitin 두 moieties로 분리 수 있습니다. 한 또 다른 이유에 대한 높은 친화력의 자발적 reconstitute이 moieties. B. 글리신에 이소 류신의 위치 13 UB I 포인트 돌연변이가 (N UB G)이 자발적으로 다시 연결을 방지합니다. C. 신화 시스템에서 C UB는 미끼 - 오브 - 관심 (B)에 융합되고 먹이가 N UB G (A)에 융합이다. AB 단백질 상호 작용은 의사 ubiquitin 재구성.
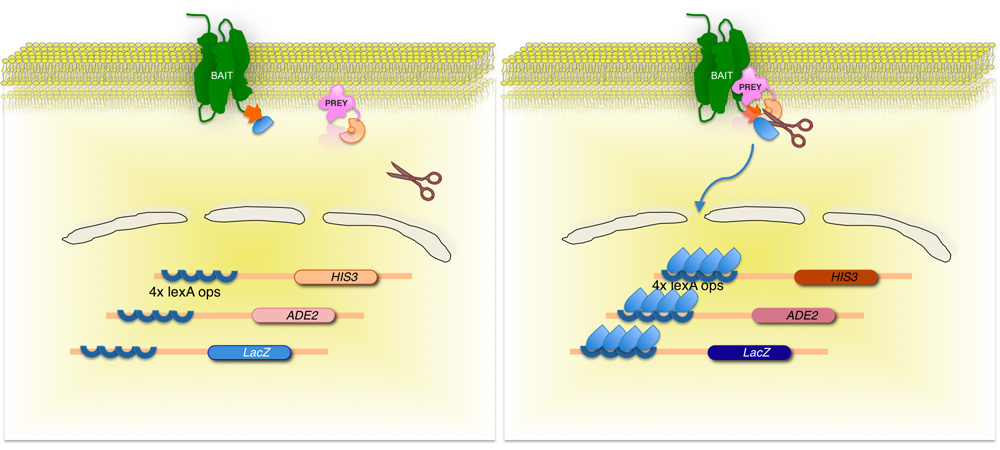
그림 2. 스플릿 - ubiquitin 기반 멤브레인 효모 두 하이브리드 (신화) 시스템.
막 단백질의 - 관심 (미끼)는, ubiquitin 효모의 C - 터미널 절반 (C UB)에 전사 인자에 복합 융합이다. cDNA 또는 gDNA 라이브러리를 사용하여 라이브러리 (먹이)로 인코딩된 각 단백질은 ubiquitin 잔기 (N UB G)의 N - 말단에 해당하는 융합이다. 두 단백질이 상호 작용을하지 않는 경우, 전사 인자는 멤브레인 인터페이스 (왼쪽 패널)에 남아 있습니다. 그러나 단백질이 상호 작용하는 경우, 두 ubiquitin moieties은 ubiquitin 특정 프로 테아제에 의해 절단의 결과로, 가입. 절단은 기자의 유전자의 표현 (오른쪽 패널)의 결과로, 전사 인자 출시

그림 3. 신화 파이프라인.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
신화는 전체 길이 막 단백질과 cytosolic 또는 멤브레인 - 바운드 파트너 간의 상호 작용의 신분을 허용 최초의 높은 처리량 시스템입니다. 그것은 생물 [3-7]의 범위에서 막 단백질을 연구하는 데 사용되었습니다. 단백질의 - 관심이 신화에서 공부 의무가되도록하기 위해 시시콜콜 따지는해야 할 수 있습니다 구체적인 내용은, 그러나,있다.
대부분의 멤브레인 - 바운드 단백질은 이후 성숙 단백질을 생산하기 위해 죽습니다 신호 순서를 통해 플라즈마 막로 이동합니다. 이 시퀀스는 유기체 특정이며 신호 순서는 알 수없는 남아 있기 때문에 그것은 누룩으로 표현 때 원시 신호 순서는 단백질의 - 관심 잘못 지방화를 발생시킬 수 있습니다. 이 문제를 회피하기 위해, 우리는 암컷의 팩터 알파 (MATα)에서 파생된 효모 신호 시퀀스에 융합되는 이러한 특정 단백질을 설계. 이 펩티드 순서 (MFα - SS 소위) 효모 플라스마 막에 단백질을 다시 localizes 그리고 중요한 것은, 효모 신호 peptidases에 의해 죽습니다. 이 펩티드 순서는 plasmids pTMBV - MFα 및 pAMBV - MFα에서 발견된다.
강조해야 할 또 다른 중요한 파라미터는 미끼 표현 수준이다. 미끼의 표현을 드라이브 발기인이 매개 변수를 조절. 이것은 NubG / NubI 테스트의 양을 사용하는 미끼 표현 수준을 최적화해야 할 수 있습니다 3 - AT 미끼가 (즉 promiscuously 많은 불특정 먹이 단백질과 상호 작용) "자기 활성화"입니다 overexpression 유물을 제거하기 위해 필요한 . 효모 단백질을 검사하는 것은 iMYTH가 적용되는 가장 생리학 관련 미끼 농도를 생산하고 있습니다. 이 경우에는 유전자의 - 관심은 gDNA 내에 컵 - TF 태그입니다. 또는 외인성 단백질은 pBT3 - 인트과 낮은 미끼 표현식에서 발생하는 CYC1 모터 들고 pCMBV의 plasmids에서 표현할 수있다. plasmids pTMBV 및 pTLB1 항구 pAMBV이 ADH1 모터가 동시에 TEF1 발기인, 미끼 단백질의 드라이브를 강하게 표현 둘. 미끼 단백질 수준 추가 최적화가 필요한 경우, 그것이 TEF1 모터를 수행하는 pTLB - 1 플라스미드를 사용해야 할 수도 있습니다, 그러나, LexA의 DNA 바인딩 도메인은 궁극적으로 감소, 외인성 기자의 유전자 발기인으로 친화력을 공격하게 R156G에 변이이다 자기 활성화의 가능성 [5].
신화의 성공을 위해 중요한 역할을 또 다른 요인은 심사 과정에 사용되는 선택의 라이브러리입니다. 이것은 내생 미끼 표현 프로필에 따라 달라집니다. 예를 들어, 미끼는 특정 조직으로 표현하고, 따라서이 특정 조직에서 구축하는 라이브러리를 사용하는 것이 중요있을 수 있습니다. 이것은 생리학 관련 상호 작용이 감지되도록합니다.
신화 시스템은 공부하기 어려운되었습니다 단백질의 클래스에 대한 정보의 풍부를 제공하는 간단하고 신속한 도구입니다. 이러한 식별 상호 작용은 막 단백질의 전체 생물 학적 기능의 해설에 도움이됩니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
이고르 Stagljar는 DualSystems 생명 공학, 스위스의 공동 창립자이다.
Acknowledgments
우리는이 원고의 중요한 읽기위한 돈 에드먼즈 감사하고 싶습니다. Stagljar 연구소는 혁신 (CFI), 보건 연구 (CIHR)의 캐나다 연구소, 심장 및 뇌졸중 재단, 캐나다 암 협회와 노바티스에 대한 캐나다 재단의 기금에 의해 지원됩니다.
Materials
| Name | Company | Catalog Number | Comments |
| Polyethenlene Glycol (PEG3350) | BioShop Canada | PEG335 | |
| Lithium Acetate Bihydrate | BioShop Canada | LIA001 | |
| X-Gal (5-Bromo-4-Chloro-3-Indolyl-b-D-galactopyranoside) | BioShop Canada | XGA001 | |
| N`,N-dimethyl formamide | BioShop Canada | DMF 451 | |
| 3-amino-1,2,4-triazole (3-AT) | BioShop Canada | ATT124 | |
| Sodium phosphate dibasic | BioShop Canada | SPD307 | |
| Sodium phosphate monobasic | Fisher Scientific | BP329-500 | |
| Salmon Sperm DNA | VWR international | CA80601-120 | |
| D-Glucose | BioShop Canada | GLU501 | |
| LB Broth LENOX | BioShop Canada | LBL405 | |
| Yeast Nitrogen Base | BioShop Canada | YNB406 | |
| Yeast Extract | BioShop Canada | YEX401 | |
| Peptone | BD Biosciences | 211677 | |
| Bio-Tryptone | BioShop Canada | TRP402 | |
| Adenine Sulphate | BioShop Canada | ADS201 | |
| L-Uracil | BioShop Canada | URA241 | |
| L-Threonine | BioShop Canada | THR002 | |
| L-Histidine | BioShop Canada | HIS200 | |
| L-Methionine | BioShop Canada | MET222 | |
| L-Valine | BioShop Canada | VAL201 | |
| L-Phenylalanine | BioShop Canada | PHA302 | |
| L-Isoleucine | BioShop Canada | ISO910 | |
| L-Tyrosine | BioShop Canada | TYR333 | |
| L-Leucine | BioShop Canada | LEU222 | |
| L-Arginine | BioShop Canada | ARG006 | |
| L-Tryptophane | Fisher Scientific | BP395-100 | |
| L-Lysine | BioShop Canada | LYS101 | |
| L-Alanine | Fisher Scientific | BP369-100 | |
| Agar | BioShop Canada | AGR001 | |
| Soda Lime Galss Beads | Biospec Products | 11079105 | |
| Sodium Chloride | BioShop Canada | SLD002 |
References
- Stagljar, I., Fields, S. Analysis of membrane protein interactions using yeast-based technologies. Trends Biochem Sci. 27 (11), 559-563 (2002).
- Iyer, K. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins. Sci STKE. 275, pl3-pl3 (2005).
- Paumi, C. M. Mapping protein-protein interactions for the yeast ABC transporter Ycf1p by integrated split-ubiquitin membrane yeast two-hybrid analysis. Mol Cell. 26 (1), 15-25 (2007).
- Stagljar, I. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc Natl Acad Sci U S A. 95 (9), 5187-5192 (1998).
- Gisler, S. M. Monitoring protein-protein interactions between the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeast two-hybrid system. Mol Cell Proteomics. 7 (7), 1362-1377 (2008).
- Scheper, W. Coordination of N-glycosylation and protein translocation across the endoplasmic reticulum membrane by Sss1 protein. J Biol Chem. 278 (39), 37998-38003 (2003).
- Thaminy, S. Identification of novel ErbB3-interacting factors using the split-ubiquitin membrane yeast two-hybrid system. Genome Res. 13 (7), 1744-1753 (2003).
- Johnsson, N., Varshavsky, A. Split ubiquitin as a sensor of protein interactions in vivo. Proc Natl Acad Sci U S A. 91 (22), 10340-10344 (1994).
- Kelleher, D. J., Gilmore, R. The Saccharomyces cerevisiae oligosaccharyltransferase is a protein complex composed of Wbp1p, Swp1p, and four additional polypeptides. J Biol Chem. 269 (17), 12908-12917 (1994).
- Chevallier, M. R. Cloning and transcriptional control of a eucaryotic permease gene. Mol Cell Biol. 2 (8), 977-984 (1982).
