

Summary
MİT, geçici ve istikrarlı model organizma Saccharomyces cerevisiae ifade proteinler arasındaki etkileşimleri hassas algılama sağlar. Bu, yüksek kapasiteli bir şekilde etkileşim ortakları belirlemek amacıyla eksojen ve maya integral membran proteinleri çalışma başarıyla tatbik edilmiştir.
Abstract
Integral membran proteinlerinin temel biyolojik ve klinik önemi tam uzunlukta transmembran proteinler için protein-protein etkileşimleri (ÜFE), yüksek verimli bir tanımlama için bir maya-tabanlı sistem geliştirme istenir. Bu amaçla, bizim laboratuvar split-ubikuitin Membran Maya İki Hybrid (MİT) sistemi geliştirdi. Bu teknoloji kullanarak geçici ve istikrarlı bir protein etkileşimleri hassas algılama sağlar
Protocol
1. Temel Bilgiler
Protein-protein etkileşimleri (PPI) tüm hücresel süreçleri yöneten temel yapı taşlarıdır. Sonuç olarak, tüm etkileşimleri sıkıca bu biyolojik denge içinde bir değişim yaygın hastalığı ve kanser hücresi dönüşümün önemli bir rol oynar gibi, hücresel homeostazı korumak amacıyla düzenlenmiş olması esastır. Karmaşık sinyalleşme kaskadlar başlatmak ve arabuluculuk ile ilgili problemleri gibi sağlık alanında son önemi olmuştur ilaçlar da dahil olmak üzere çeşitli moleküller, ithalat ve ihracat gibi Membran ilişkili proteinler, proteinlerin biyolojik olarak en önemli sınıf arasında yer almaktadır ilaç direnci giderek daha yaygın hale gelmiştir. Bu proteinin sınıf karmaşıklığı içgörü kazanmak kendi etkileşim ortakları belirlenmesini gerektirir. Bu ortakların keşfederek zorlu kanıtlanmıştır gibi, sık sık her membrana bağlı bir protein için optimize edilmiş olmalıdır sert koşullar gerektirir [1].
Integral membran proteinlerinin temel biyolojik ve klinik önemi tam uzunlukta transmembran proteinler ÜFE yüksek verimli bir tanımlama için bir maya-tabanlı sistem geliştirme istenir. Bu amaçla, split-ubikuitin Membran Maya İki Hybrid (MİT) sistemi [2-4] geliştirdi. Bu araç, geçici ve istikrarlı bir protein etkileşimleri hassas bir algılama sağlar. Bu model organizma Saccharomyces cerevisiae ifade eksojen ve endojen proteinler [3-7]. Başarıyla çalışma tatbik edilmiştir MİT ubikuitin iki moieties ayrılmış olabilir gözlem yararlanır: C-terminal yarı (C ub) ve N-terminal yarısı (NubI) in vivo çalışmalar göstermiştir için yüksek afinite nedeniyle kendiliğinden sulandırmak bu moieties biri diğerine (Figure. 1a). Ancak, bir izolösin 13 ubikuitin N-terminal yarı glisin nokta mutasyonu (N ub G olarak adlandırılan bir parçası üreten) önler, bu kendiliğinden yeniden dernek [8] (Figure. 1b) .
Biz MİT sistemi (Figure. 1c ve 2) bu prensibi kullanır. Kısaca, integral membran yem protein Escherichia coli DNA-bağlayıcı protein Lexa oluşan yapay bir transkripsiyon faktörü ve herpes simpleks virüsünün VP16 aktivasyon etki ile bağlantılı bir C ub benzer parçaları birleşir. Avlayan NubG benzer parçaları cDNA veya genomik DNA elde edilen parçalarının füzyonu tarafından üretilir. Sitozolik deubiquitinating enzimler (Dubs) ve transkripsiyon faktörü proteolitik serbest sonraki tanıma, bir maya konak yem ve av proteinler arasında bir etkileşim, bir tam uzunlukta 'pseudo-ubikuitin' molekülünün sulandırıldıktan yol açar. Transkripsiyon faktörü daha sonra hücrenin çekirdeğinde girin ve yem ve av etkileşimi göstergesidir selektif ortam üzerine maya suşları büyüme sağlayan bir muhabir gen sistemi (genellikle HIS3 lacZ ve ADE2 genleri içeren ifade), aktive [ 2-4].
Integral membran protein etkileşimleri belirlenmesi ve tanımlanması, kendi işlevlerini tam olarak anlamalarına yardımcı olmak için bilgi sağlayacaktır. Daha doğrusu anlamak ve integral membran proteinleri ile etkileşime proteinlerin rolleri teşrih olarak, biz bu proteinlerin düzenlenmesinde yer alan dinamik bir etkileşim içine anlayış kazanmak ve terapötik bir potansiyele sahip yeni hedeflere keşfetmek.
2. Yem ve Uygun MİT Sistem Seçimi
- Önce MİT analizlerinin yapılması, protein, hücrenin sitoplazmada N-ve / veya C-terminalindeki olduğunu doğrulayın. C ub-Lexa-VP16 etiketi, transkripsiyon faktörü serbest bırakılması için gerekli deubiquitinating enzimler sitoplazmada bulunmaktadır beri böyle bir terminus protein erimiş olması şarttır [4].
- Sonra, iki büyük MİT varyantları hangi uygundur karar verir. Yerli olmayan maya proteinleri için, geleneksel MİT (tMYTH), kullanılan yemler bir plazmid ektopik aşırı eksprese eden olabilir. Doğal maya proteinleri için, entegre MİT (iMYTH) tercih edilen bir yöntemdir. IMYTH olarak yemler içsel olarak kendi ana organizatörü kontrol altında bırakarak, C ub-Lexa-VP16 etiketiyle etiketlenir. Bu yemler yaban tip ekspresyon seviyesi artan bir sayıda yanlış pozitif [3] gibi protein aşırı ekspresyonu ile ilişkili sorunları ortadan kaldırmak yardımcı olarak avantajlıdır. Ilk yem inşaat ve kullanılan seçici besiyerleri hariç olmak üzere, MİT her iki türlü bir temelde aynı şekilde yürütülmektedir. Netliği için, biz bu varyant, prensip olarak, hemen hemen her organizma membran proteinleri ile kullanılan ve olması gibi, ağırlıklı olarak bu raporda tMYTH kullanımı üzerinde durulacakböylece daha yaygın olarak uygulanabilir.
3. Yem Üretimi ve Doğrulama
- Gerekli Medya ve Çözümler
- Steril GKD 2 O, 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak hazırlanmıştır .
- 3-amino-1 ,2,4-triazol (3-AT) GKD 1M bir stok solüsyonu olarak hazırlanan çözüm 2 O. 0.2 mikron filtre geçişi ile sterilize edin.
- % 1 w / v maya özü,% 2 w / v pepton,% 2 w / v glikoz ve GKD 2 O. hazırlanan 100 mcM adenin oluşan YPAD Büyüme Medya 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak sterilize edin.
- 10x Amino Asit / Nükleotid Temel Karması: tam karışımı 1.0 mM Adenin, 1.8 mM Urasil, 1.0 mM Arginin, 1.0 mM histidin, 2.3 mM Isoleucine, 7.6 mM Lösin, 1.6 mM Lizin, Metionin 10.1 mM, 3.0 mM Fenilalanin, 16,8 mM treonin, 2.0 mM Triptofan, 1.7 mM Tirozin ve 12.8 mM Valin, GKD 2 O hazırlanmıştır. Için damla-out gerekli amino asit (ler) ve / veya nükleotid baz (ler) çıkarırsanız. 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak sterilize edin.
- Sentetik Drop-Out (SD) 0.67% w / v maya azot tabanı (amino asitler olmadan ama amonyum sülfat), w / v şekeri% 2, 2 w / v agar ve% 1x Amino Asit / Nükleotid Mix oluşan Büyüme Medya, GKD 2 O. hazırlanan Hem sıvı ve katı (% 2 agar içeren) SD-Leucine ortamı hazırlayın. Ayrıca katı SD-Triptofan-Leucine ve SD-Triptofan-Leucine-Adenin-Histidin medya hazırlar. 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak sterilize edin. Katı ortam 100x15 mm Petri kaplarına dökün.
- 3-AT içerenler Sentetik Drop-Out (SD) Büyüme Medya açıklandığı gibi SD-Triptofan-Leucine-Adenin-Histidin ortam hazırlayın, ancak, 25, 50, 75 ve 100 mM konsantrasyonlarda 3-AT içeren . Uygun miktarda ekle 3-AT (ama henüz katılaşmış değil) otoklavlanabilir ve soğutulduktan sonra medyaya 1M steril stok solüsyonu. 100x15 mm Petri kaplarına dökün.
- PEG / Lityum Asetat Mix oluşan% 40 w / v PEG-3350, 120 mM Lityum asetat ve 167 mg / ml GKD 2 O. hazırlanan Somon Sperm DNA (Tip III Sodyum Tuz) Bu karışımı steril steril su ve çözümler hazırlamak sağlamak için (yani,% 50 PEG-3350 otoklava steril GKD 2 O hazırlanan 1M Lityum asetat ve 2 mg / ml Somon Sperm DNA tip III sodyum tuzu otoklava).
- Enzim ve PCR yürütülmesi için reaktifler.
- Ticari miniprep kiti.
- Soda kireç cam boncuklar (0.5 mm).
- Plazmid yayılımı (örneğin DH5α, XL10 altın) ve bakteriyel yayılımı ve plazmid seçimi için uygun standart ortam için uygun olan Yetkili Escherichia coli hücrelerinde .
- Özel maya suşları, protokolde açıklandığı gibi plazmid ve astarlar.
- Boşluk Onarımı tMYTH yemler Üretimi
- Etiketleme ve ifade etmek için uygun bir vektör içine yem klonlanmış olmalıdır. TMYTH vektörleri çeşitli yem yapımında kullanmak için şu anda mevcuttur. PCMBV4 pAMBV4 ve pTMBV4 gibi vektörler izin sırasıyla CYC1 (zayıf), ADH1 (güçlü) ve TEF1 (çok güçlü) destekçileri, kontrol altında, C-terminal etiketli yemler inşaatı (DİYECEK-C ub-Lexa-VP16 ) . N-terminal etiketli yemler (Lexa VP16-C ub-DİYECEK) sırasıyla pTLB TEF1 ve CYC1 yararlanıcı kontrol altında-1 ve pBT3-N, gibi vektörler kullanarak oluşturulabilir. Vektör seçimi yem bağlıdır ve ampirik olarak tespit edilmelidir. Aslında diğer durumlarda yem aşırı ekspresyonu artan bir sayıda yanlış pozitif yol açan zararlı olabilir, bazı durumlarda daha yüksek yem ifade, etkileşimleri tespit etmek için gerekli.
- Kısıtlama uygun kısıtlama site (ler) seçilen plazmid sindiremez. Dilinim C ub-Lexa-VP16 tag (etiket C-terminal etiketleme veya N-terminal etiketleme sonra upstream) hemen yakınında meydana gelmelidir. Örneğin, pAMBV4 vektör kullanırken, SfiI ideal bir seçimdir. Kullanıma hazır olana kadar, -20 ° C'de sindirilir plazmid saklayın.
- Tasarım ilgi gen amplifikasyonu ve klonlama için primerler. Sonunda hedef gen ilk 18-20 nükleotid eşleşmelidir 5 '3 İleri Primer sonunda kısıtlama sitesi upstream yaklaşık 35-40 nükleotidler eşleşmelidir'. 5 ucu ters tamamlayıcı hedef genin son 18-20 nükleotidlerin (eşleştirme 'Ters Primer sonu 3 ile kısıtlama sitesinde alt yaklaşık 35-40 nükleotidlerin ters tamamlayıcı eşleşmelidir' stop kodon atlayarak eğer C ub-Lexa-VP16 tag) C-terminalindeki konuyor. Bağlı olsun N-ya da C-terminali etiketleme yapılıyorsa, hedef gen C ub-Lexa-VP16 etiketiyle çerçeve klonlanmış olduğunu İleri veya Ters astar 35-40 nükleotidler seçin.
- Yukarıdaki primerler kullanılarak PCR ile ilgi gen yükseltin. PCR parametreler belirli enzim ve kullanılan spesifik primerler bağlıdır.
- PCR ürünü leu2 mutasyon (örneğin BY4741) taşıyan uygun bir maya laboratuar gerginlik içine sindirilmiş plazmid ile birlikte dönüşümü. MİT muhabiri suşu (örneğin THY.AP4 veya L40) kullanılan ancak gerekli değildir, bu noktada maya amaç olarak sadece boşluğu onarım homolog rekombinasyon oluşabilir olduğu bir ortam olarak hizmet vermeye olabilir. Dönüşüm şu şekilde yapılmalıdır:
- Seçilen maya suşu tek bir koloni 5 ml steril YPAD medya İnokülasyon ve 30 gece boyunca inkübe ° C sabit sallayarak (200 rpm).
- ~ 0.15 OD600 50 ml taze YPAD medya gecelik kültürü seyreltilir ve 30 ° C'de sallayarak (200 rpm). ~ 0.6 OD600 ulaşana kadar yaklaşık 3-4 saat için büyütün.
- 5 dakika 700xg hücreleri Santrifüj ve supernatant çıkarın.
- 25 ml steril GKD 2 O hücre pelet ve 5 dakika santrifüjlenir 700xg tekrar
- Süpernatantı alın ve 1 ml steril GKD 2 O. hücre pelletini tekrar süspansiyon
- Bir mikrofuge'de tüp, hücrelerin 100 mcL, 300 mcL PEG / Lityum asetat karışımı, ve sindirilmiş plazmid (50 fmol) ve PCR ürünü (250-500 fmol) ekleyin.
- 30 ° C'de 30 dakika inkübe edin.
- 1 saat süreyle 42 ° C'de ısı şoku.
- 3000xg az 5 dakika süreyle santrifüj ve süpernatantı kaldırmak.
- 200 mcL steril GKD 2 O hücre pelet ve plaka, katı, SD-Leucine seçici ortama tüm hacim tekrar 2-4 gün süreyle 30 ° C'de büyütün.
- 30 ° C gecede 5 ml SD-Leucine sıvı ortam dönüştürülmüş suşu tek bir koloni büyüyorlar.
- 5 dakika için 700xg hücreleri Santrifüj ve supernatant çıkarın.
- , Herhangi bir ticari miniprep kiti kullanılarak hücre pelet yem plazmid DNA izole edin. Bir değişiklik ile standart bir protokol izleyin. Yeterli maya hücre parçalama sağlamak amacıyla, 5 dakika boyunca şiddetle ilk tabanda ve vorteks, sonra pelet 0,5 mm soda kireç cam boncuklar küçük hacimli ekleyin. Sonra normal olarak ticari protokol ile devam edin.
- Yetkili E. izole maya DNA Dönüşümü coli plazmid yayılma bir dönüşüm verimliliği ile en az 1x10 7 hücre / mg DNA (örneğin DH5α, XL10 Altın) uygun süzün. Kanamisin kullanarak yem plazmid için seçilebilir unutmayın.
- Hasat plazmid DNA dönüştürülmüş E. standart bir DNA izolasyon yöntemi veya ticari kit kullanılarak coli.
- Sıralama ile plazmid yem uygun inşaat olun.
- Doğrulanmış yem, uygun bir MİT muhabiri suş (örneğin THY.AP4, L40) içine inşa Transform. Açıklanan maya dönüşüm protokolü sindirilir plazmid ve PCR ürün yerine yem plazmid DNA yerine kullanılabilir.
- Bait Doğrulama - Uygun Yerelleştirme
- Kullanmadan önce, yem suşları düzgün maya membran lokalize olduğunu sağlamak için analiz edilir. IMYTH kullanırken, bu lokalizasyon etiketli yem özellikleri üzerine bağlı olacaktır. TMYTH için, yem plazmidler genellikle bir sinyal dizisi (örneğin Matα) plazma membranına ifade protein yönetmenlik. Yerelleştirme floresan mikroskopi kullanılarak belirlenir. YFP yem etiketi sırayla bir molekülün İçerme (yani C ub-YFP-Lexa VP16), canlı hücreleri doğrudan görselleştirme sağlayan basit ve en doğrudan bir yaklaşım ve iMYTH yaygın olarak kullanılır. Alternatif olarak, Lexa veya etiketi VP16 bileşenleri karşı antikor kullanarak standart immunofloresan yaklaşım kullanılabilir.
- Bait Doğrulama - N ub G / Test Kontrol
- Yem kez uygun lokalizasyon kurulmuş olup, yem tek başına ya da olmayan etkileşim avlayan varlığı (yani yem kendini-aktive olmadığını doğrulamak) muhabiri sistemini aktive etmediğini sağlamak için gerekli. Bu yem etkileşim (pozitif) ve etkileşimli olmayan (negatif) kontrol avlayan dönüşür ve büyüme seçici medya değerlendirilir N ub G / I Testi, kullanılarak yapılır. Bir yem seçici medya büyümek zorundadır in pozitif kontrol varlığı, ve MİT kullanmak için uygun olmak için, negatif kontrol varlığı büyümek değil.
- Plazmid kontrol yırtıcı 100-200 ng ile test edilecek yem gerginlik dönüştürerek başlayın. Daha önce açıklanan maya dönüşüm protokolü YPAD yerine SD-Leucine medya yerine ve nihai kaplama adım için SD-Triptofan-Leucine ortamını kullanarak, kullanılabilir. Aşağıdaki kontrol av yapıları yaygın olarak kullanılan:
- İleti1-N UB (Pozitif Kontrol)
- İleti1-N ub G (Negatif Kontrol)
- pFUR4-N UB (Pozitif Kontrol)
- pFUR4-N ub G (Negatif Kontrol)
- OST1 oligosaccharyl kompleksinin bir bileşeni olan ve endoplazmik retikulum zarı [9] lokalize FUR4 urasil permease ve plazma membran lokalize iken [10]. Bu proteinler genellikle non-etkileşim avlayan olarak kullanmak için ilk seçim olmasına rağmen, onların uygunluğu vaka bazında bir durumda değişecektir. Yem gerçekten bu kontroller ile etkileşim tahmin etmesi halinde, alternatif avlayan seçilmesi gerekir. N ub ubikuitin N-terminus yabani tip ve kendiliğinden C ub bağımsız C ub ve N ub erimiş olan proteinler arasında bir etkileşim ile etkileşim olduğunu hatırlayın. N ub G avlayan (N ub ve C ub spontan ilişki engeller Glycine mutasyon Isoleucine taşıyan 13), negatif kontrol olarak hizmet Böylece N ub beslenmez, pozitif kontrol oluşturmaktadır .
- 100 mcL steril GKD 2 O. dönüştürülmüş her yem tek koloniler süspanse edin
- Çözülmelere 1 / 10, 1 / 100 ve 1 / 1000 seri olarak üretmek için yeniden süspanse hücreler steril GKD 2 O seyreltin.
- Konsantrasyonlarının bir dizi olan ve olmayan 3-AT, SD-Triptofan-Leucine ve SD-Triptofan-Leucine-Adenin-Histidin medya üzerine seyreltilmemiş ve seyreltilmiş hücreleri 5 mcL hacimleri Nokta. 3-AT HIS3 muhabiri gen rekabetçi bir inhibitörü olarak davranır ve seçim sürecinin darlığı arttırmak için hizmet vermektedir. Zayıf orta kendini-aktive yemler inhibe non-spesifik büyüme için bazı durumlarda yararlı olabilir.
- Lekeler kuru ve daha sonra 2-4 gün süreyle 30 ° C'de inkübe plakalar için izin verin.
- Tüm transformants başarıyla plazmid av dönüştürülmüş olduğunu belirten SD-Triptofan-Leucine plakalar üzerinde büyümek gerekir. Olmayan yemler kendini etkinleştirmek N ub G beslenen yapıları yırtıcı değil, N ub ile dönüştürülmüş sadece SD-Triptofan-Lösin-Adenin-Histidin medya büyüyecek. Ne konsantrasyon Not 3-AT (eğer varsa) bu tarama sırasında kullanılmak üzere ihtiyaç olarak medya gereklidir.
4. Tarama
- Gerekli Medya ve Çözümler
- Steril GKD 2 O, 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak hazırlanmıştır .
- GKD 2 O hazırlanan ve 121 otoklavlanarak steril% 0.9 NaCl Çözüm ° C, 30 dakika boyunca 15 psi.
- GKD 2 O. 493 mM sodyum fosfat dibazik ve 250 mM sodyum fosfat tekbazlı oluşan Sodyum Fosfat Çözüm 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak sterilize edin.
- X-Gal (5-bromo-4-kloro-3-indolyl-β-D-galactopyranoside) Çözüm, N, N-dimetil formamid 100 mg / mL stok çözelti olarak hazırlanmıştır .
- GKD 2 O. hazırlanan% 2 w / v maya özü,% 4 w / v pepton, adenin% 4 w / v glikoz ve 100 mcM içeren 2xYPAD Büyüme Medya 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak sterilize edin.
- Daha önce açıklandığı gibi hazırlanmış Sentetik Dropout (SD) Büyüme Medya, . Sıvı SD-Leucine ve katı SD-Triptofan-Leucine hazırlayın. Katı ortam hem 100x15 mm Petri kaplarına dökün. N ub G / I kontrol testi belirlenen konsantrasyonda 3-AT içeren 150 mm yuvarlak plakalar, her ekran için 16 tabak, gerekirse, katı SD-Triptofan-Leucine-Adenin-Histidin ortam hazırlayın.
- Sentetik Dropout (SD) Medya + 5-Bromo-4-kloro-3-Indoyl-β-D-Galactopyranoside (X-gal). Hazırlayın SD-Triptofan-Lösin-Adenin-Histidin daha önce açıklandığı gibi agar içeren medya. Otoklav sonrası 1/10th hacmi o 3-AT (gerekirse), soğuk eklemek için izinf steril Sodyum Fosfat çözümü. Sonra, son bir 80 mg / ml konsantrasyonu X-Gal Çözüm ekleyin. İyice karıştırın ve 150 mm yuvarlak levhalar içine dökünüz.
- PEG / Lityum Asetat Çözüm II% 40 içeren PEG-3350, 100 mM Lityum asetat, 1 mM EDTA ve 10 mM Tris, pH 7.5. Steril GKD 2 O ve çözümleri kullanarak bu çözüm hazırlayın (örneğin otoklava% 50 PEG-3350, 1 M Lityum Asetat, 100 mM Tris, pH 7.5 ve 500 mM EDTA pH 8.0).
- Lityum asetat / 110 mM Lityum asetat, 11 mM Tris, pH 7.5 ve 1.1 mM EDTA içeren Tris EDTA Çözüm. Bu çözüm steril GKD 2 O ve çözümleri (örneğin otoklava 1M Lityum Asetat, 100 mM Tris, pH 7.5 ve 500 mM EDTA pH 8.0) kullanarak hazırlayın.
- GKD 2 O. hazırlanan 100 mM Tris, pH 7.5 ve 10 mM EDTA oluşan 10x Tris EDTA Çözüm 121 ° C, 30 dakika boyunca 15 psi otoklavlanarak sterilize edin.
- Steril GKD 2 O. hazırlanan 2 mg / ml sperm Somon DNA Tip III sodyum tuzu içeren tek zincirli Taşıyıcı DNA (ssDNA) Çözüm
- Ticari miniprep kiti.
- Soda kireç cam boncuklar (0.5 mm).
- Plazmid yayılımı (örneğin DH5α, XL10 altın) ve bakteriyel yayılımı ve plazmid seçimi için uygun standart ortam için uygun olan Yetkili Escherichia coli hücrelerinde .
- Protokolde açıklandığı gibi özel maya suşları ve plazmid
- Büyük Ölçekli Dönüşüm
- MİT muhabiri suşu SD-Leucine medya 5 ml yem içeren tek bir koloni aşılamak ve 30 gece inkübe ° C sallayarak (200 rpm).
- Bir OD600 200 ml SD-Leucine medya gecelik kültürü sulandırınız 30 = 0.15 ve inkübe ° C sallayarak (200 rpm). OD600 kadar büyümeye = 0.6 - 0.7 (yaklaşık 4-5 saat).
- Hedef OD600 ulaşıldığında kısa bir süre önce, bir kısım ssDNA çözüm Çözülme. Kaynatın, 100 ° C buz üzerinde serin sonra 5 dakika. Bir kez tekrarlayın.
- Hedef OD600 ulaşıldığında, 5 dakika (200 ml, 4x50 ml vidalı kapaklı santrifüj tüplerine arasında kültür bölme) 700xg santrifüj yoluyla hücreler hasat.
- 30 ml steril GKD 2 O ve kısaca vorteks örnek her pelet yıkayın. 5 dakika 700xg santrifüjleyin.
- Süpernatantı atın ve 1 ml Lityum asetat / Tris EDTA çözeltisi her pelet tekrar süspansiyon haline getirin. 1,5 mL steril bir mikrofuge'de tüp ve 5 dakika santrifüjlenir 700xg aktarın.
- Süpernatantı atın ve Lityum asetat / Tris EDTA çözeltisi 600 mcL her pelet tekrar süspansiyon haline getirin.
- 4x15 ml vidalı kapaklı santrifüj tüplerine aşağıdakileri ekleyin:
- 2.5 mL PEG / Lityum Asetat Çözüm II
- 600 mcL yeniden süspanse hücreleri
- 100 mcL ssDNA çözüm
- 7 av kütüphane DNA mikrogram
- Kütüphaneler, N ub G ile N-veya C-terminalindeki ya tagged ve cDNA ve genomik kaynakları çeşitli hazırlanan avlayan içeren (piyasada www.dualsystems.com ). Kullanılan özel kütüphanesi seçilen yem ve deneysel amaçlara bağlı bir case-by-case temelinde tespit edilmelidir.
- Vorteks tüpleri 1 dakika sonra 45 dakika boyunca 30 ° C su banyosunda inkübe edin tam karıştırma sağlamak için. Her 15 dakikada bir kısa bir süre karıştırın.
- Her bir tüp 160 mcL dimetil sülfoksit (DMSO) tüpleri tersini hemen ekleyin ve karıştırın.
- 42 ° C su banyosunda 20 dakika inkübe edin.
- 700xg az 5 dakika santrifüj yoluyla transformants toplayın.
- Süpernatantı atın. 3 ml 2xYPAD her pelet tabanda transformants kurtarın. Birlikte tek bir 50 ml vidalı kapaklı santrifüj tüpüne Havuzu tüm örnekler.
- Hücre kurtarma için 90 dakika süreyle 30 ° C'de inkübe edin.
- 700xg az 5 dakika Santrifüj ve supernatant atın.
- 4.9 ml steril% 0.9 NaCl hücre pelletleri tekrar
- Yeniden süspanse hücrelerinin 100 mcL kadar steril% 0.9 NaCl içinde 10 kat seri dilüsyonları hazırlamak 1000x 10x.
- SD-Triptofan-Leucine medya ve inkübe üzerine 100x 1000x dilüsyonları 30 Plaka 100 mcL ° C de 2-3 gün için. Bu plakalar kontrol olarak hizmet veren ve dönüşüm verimini hesaplamak için kullanılır..
- Aynı şekilde, büyük (150 mm) SD-Triptofan-Leucine-Adenin-Histidin plakalar üzerine kalan 4.8 mL yeniden süspanse hücre ve plaka bölmek3-4 gün boyunca 30 ° C'de içeren N ub G / I testi belirlenen 3-AT gerekli miktar ve kuluçkaya yatmaktadır.
- Üzerine% 0.9 NaCl ve plaka 5 mcL alikotları 100 mcL tek koloniler süspanse edin (her temsil eden hücreler potansiyel bir etkileşim yem-av çifti içeren) SD-Triptofan-Leucine-Adenin-Histidin + X-Gal medya (ve eğer 3-AT gereklidir). 2-4 gün boyunca büyümeye izin verin. Bu adım, ikinci turda, seçici tarama olarak hizmet veren ve ilk turda elde edilen yanlış pozitif kaldırılmasına yardımcı olur. Sadece koloniler güçlü büyüme ve mavi bir renk ekran, daha fazla analiz için seçilir.
- DNA İzolasyonu ve sıralamaya Prey
- Değişiklik ile daha önce açıklanan bir miniprep protokolünü kullanarak mavi maya kolonileri plazmid DNA izole edin. Sadece av tutma için seçin, ama yem, plazmid için SD-Triptofan medya hücreleri büyümek için emin olun. Çok sayıda hit çıkaran ekranları için, piyasada bulunan yüksek verimlilik miniprep kiti bu noktada yararlı olabilir.
- Bir yetkili E. içine izole maya plazmid DNA Dönüşümü coli plazmid yayılma bir dönüşüm verimliliği ile en az 1x10 7 hücre / mg DNA (örneğin DH5α, XL10 Altın) uygun süzün. Av plazmid ampisilin kullanımı için seçilebilir unutmayın.
- Dönüştürülmüş E. plazmid DNA'sı izole standart bir DNA izolasyon yöntemi veya ticari kit kullanılarak coli. Numune sayısı büyükse Bir kez daha, yüksek verimli miniprep kiti, yararlı olabilir. E. DNA amplifikasyon coli plazmid verimi önemli ölçüde artırır ve DNA sıralama ve daha fazla analiz için yeterli miktarda mevcut olduğunu garanti eder .
- N ub G. içinde dizisi tamamlayıcı bir astar kullanarak izole plazmid Sıralı
- Interactors ön liste monte tüm sıralama verileri derlemek ve analiz. Bu elle yapılması, ya da uygun bir yazılım kullanarak otomatik bir şekilde olabilir.
- Bait Bağımlılık Testi
- Interactors ön montaj sonra etkileşimler tekrar kontrol edin ve etkileşim / yem kimlik bağımsız bir şekilde muhabiri sistemini aktive promiscuous avlayan ortadan kaldırmak için önemlidir. Bu Bait Bağımlılık Testi kullanılarak yapılır. Bu testte, belirlenen interactors orijinal yem gerginlik yanı sıra C ub-Lexa-VP16 etiketi ile kaynaşmış tek bir transmembran etki alanı oluşan bir kontrol yapay yem barındıran bir yük haline dönüşür. Dönüşüm YPAD ve nihai kaplama adım için SD-Triptofan-Leucine katı medya yerine SD-Leucine medyayı kullanarak, standart bir protokol daha önce açıklanan uyarınca yapılır.
- SD-Triptofan-Leucine-Adenin-Histidin + X-Gal medya (ve gerekirse 3-AT dahil olmak üzere) üzerine 5 mcL hacimli steril GKD 2 O ve spot 100 mcL yukarıdaki dönüşümler tek koloniler yeniden süspanse. Tabaklar, sonra 2-4 gün 30 ° C süreyle inkübe İdeal olarak, her av için birden fazla transformants seçilmiş olmalıdır ve orijinal yem ve suni yem aynı plaka üzerine tespit edilmelidir.
- Maya muhabiri sistemi (yani büyüme ve mavi renk) neden aktivasyon promiscuous olarak kabul edilir ve o belirli av interactors listesi çıkarılır yapay yem ve av taşıyan.
- Maya büyüme ve mavi rengi neden yem faiz, ancak yapay yem ile beslenen, bu özel etkileşim onaylar. Ancak, maya av barındıran ve yem faiz büyümek değil, bu av interactors listesi çıkarılır.
- Kalan avların, MİT ekran tespit interactors tam listesini oluşturmaktadır.
5. Başka Çalışmalara
MİT tarama tamamlandıktan sonra tespit etkileşimleri biyolojik önemi doğrulamak ve belirlemek amacıyla, daha fazla analiz yapılmalıdır. Özel çalışmalar yapılması, case-by-case bazında değişir ve bireysel araştırmacı tarafından tespit edilmelidir. Takip çalışmalarının bazı yaygın örnekler yerli organizmanın co-immunoprecipitation deneyler ve silme çalışmaları içerir. Ayrıca, elde edilen verileri hesaplama analiz desenler tespit ve farklı etkileşimler oynayabilir potansiyel önemi ve rolünü belirlemek için yardımcı olmak için yararlı olabilir. Böylece, MİT teknolojisi, membran proteinlerinin kritik fonksiyonel etkileşimlerin belirlenmesi ve anlayışa karşı güçlü bir ilk adım 'olarak hizmet vermektedir. Detaylı takip çalışmaları ve diğer yakın zamanda gelişmiş ve gelişmekte olan te ile birleştiğindechnologies, gizemleri hücrenin kilidini değerli bir aracı olmayı vaat ediyor.

Şekil 1. Bölünmüş ubikuitin a. Prensibi C-terminal yarı (C ub) ve N-terminal yarısı (N ub I): Ubiquitin iki moieties ayrılmış olabilir. Bu moieties biri başka bir yüksek afinite nedeniyle kendiliğinden sulandırmak için. b. Glisin bir izolösin pozisyon 13 AN ub I nokta mutasyonu (N ub G), bu kendiliğinden yeniden-dernek engeller . c. MİT sisteminde, C ub yem faiz (B) erimiş ve av N ub G (A) birleşir. AB protein etkileşim pseudo-ubikuitin yeniden oluşturulmasını sağlamak.
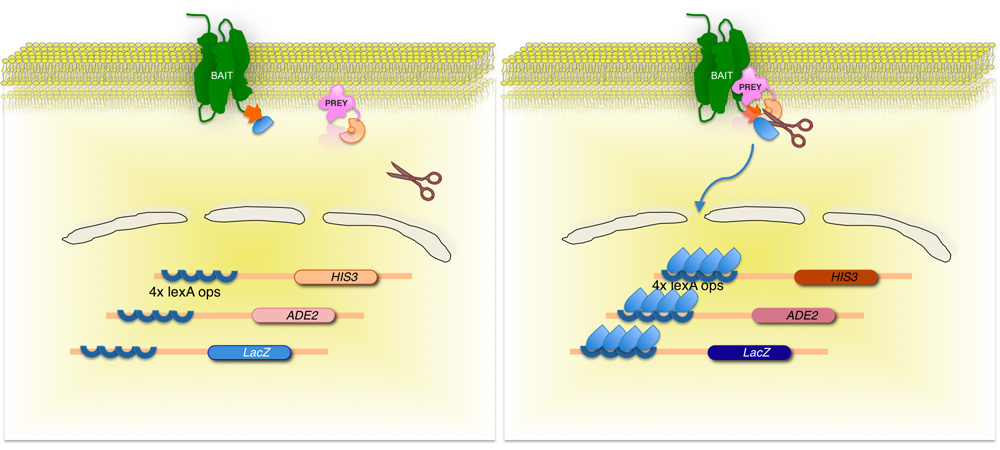
Şekil 2. Split-ubikuitin esaslı membran maya iki hibrid (MİT) sistemi.
Membran protein-faiz (yem), C-terminal ubikuitin maya yarısı (C Ub), bir transkripsiyon faktörü konjuge erimiş . CDNA veya gDNA bir kütüphaneyi kullanarak, kitaplığı (av) tarafından kodlanan her protein ubikuitin benzer parçaları (N Ub G), N-terminus gelen erimiş. Iki protein etkileşim yoksa, transkripsiyon faktörü membran arayüzü (sol panel) kalır. Ancak, proteinler etkileşim halinde, iki ubikuitin moieties ubikuitin spesifik proteazlar tarafından bölünme katılmak. Dilinim muhabiri genlerin ekspresyonu (sağ panel), transkripsiyon faktörü bültenleri

Şekil 3. MİT boru hattı.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
MİT, tam uzunlukta membran proteinleri ve sitozolik veya membrana bağlı ortaklar arasındaki etkileşimlerin belirlenmesi sağlayan ilk yüksek kapasiteli bir sistemdir. Membran proteini organizmalar [3-7] bir dizi çalışma için kullanılır olmuştur. Ancak, protein-faiz MİT ile çalışmak için uygun olduğundan emin olmak için incelenip gerekebilir belirli ayrıntıları vardır.
Birçok membrana bağlı proteinler daha sonra olgun bir protein üretmek için bölünmüş bir sinyal dizisi ile plazma membran yönlendirilir. Bu dizi organizma özgüdür ve sinyal dizisi tanınmayan kalır çünkü bu native sinyal dizisi maya ifade protein faiz yanlış yerelleştirme neden olacağını mümkündür. Bu sorunu aşmak için, biz Çiftleşme faktör alfa (MATα) türetilen maya sinyal dizisi, erimiş olması bu özel proteine tasarlanmıştır. Bu peptid dizisi (MFα-ss sözde) maya plazma membran proteini yeniden lokalize ve daha önemlisi, maya sinyal peptidazlar tarafından bölünmüş. Bu peptid dizisi plazmid pTMBV MFα ve pAMBV-MFα bulundu.
Vurgulanması gereken bir diğer önemli parametre yem ifade seviyeleridir. Yem ifade sürücüler İşverenin bu parametre düzenler. NubG / NubI test ve miktarını kullanarak yem ifade seviyelerini optimize etmek için gerekli olabilir 3-AT yem (yani promiscuously, non-spesifik birçok yırtıcı proteinler ile etkileşime girer) "self-aktive" aşırı ekspresyonu eserler, ortadan kaldırmak için gerekli . Maya proteinleri incelenmesi iMYTH uygulanan en önemli fizyolojik yem konsantrasyonları üretir. Bu durumda, gen-faiz gDNA içinde Cub-TF ile etiketlenir. Alternatif olarak, eksojen proteinler pBT3-STE ve düşük yem ifade CYC1 organizatörü taşıyan pCMBV plazmid ifade edilebilir. Plazmid pTMBV ve pTLB1 liman pAMBV ADH1 organizatörü ise TEF1 organizatörü, yem protein bu sürücü güçlü bir ifadesi hem. Yem protein düzeyleri daha fazla optimize etmek gerekiyorsa, TEF1 organizatörü taşıdığı pTLB-1 plazmid kullanmak gerekli olabilir, ancak sonuçta azalan Lexa DNA bağlayıcı etki alanı dışsal muhabiri gen rehberleri doğru yakınlık azaltabilmek için R156G mutasyona uğramış. kendini etkinleştirme olasılık [5].
MİT başarı için önemli bir rol oynayan diğer bir faktör, tarama işlemi için kullanılan seçim bir kütüphanedir. Bu endojen yem ifade profilleri bağlıdır. Örneğin, yem belirli dokularda ifade ve bu nedenle bu özel doku inşa edilmiş bir kütüphane kullanmak için önemlidir olabilir. Bu fizyolojik ilgili etkileşimleri tespit sağlayacaktır.
MİT sistemi, çalışmak için zor olmuştur proteinlerin bir sınıfı hakkında bilgi bir bolluk sağlayan basit ve hızlı bir araçtır. Bu tespit etkileşimleri, membran proteinlerinin tam bir biyolojik fonksiyonu aydınlatılmasında yardımcı olabilir.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Igor Stagljar DualSystems Biotech, İsviçre kurucularından.
Acknowledgments
Biz Şafak Edmonds Bu yazının eleştirel bir okuma için teşekkür etmek istiyorum. Stagljar laboratuvar Kanada Yenilik (CFI), Kanada Sağlık Araştırmaları (CIHR) Enstitüsü, Kalp ve İnme Vakfı, Kanada Kanser Derneği ve Novartis Vakfı fonları tarafından desteklenmektedir.
Materials
| Name | Company | Catalog Number | Comments |
| Polyethenlene Glycol (PEG3350) | BioShop Canada | PEG335 | |
| Lithium Acetate Bihydrate | BioShop Canada | LIA001 | |
| X-Gal (5-Bromo-4-Chloro-3-Indolyl-b-D-galactopyranoside) | BioShop Canada | XGA001 | |
| N`,N-dimethyl formamide | BioShop Canada | DMF 451 | |
| 3-amino-1,2,4-triazole (3-AT) | BioShop Canada | ATT124 | |
| Sodium phosphate dibasic | BioShop Canada | SPD307 | |
| Sodium phosphate monobasic | Fisher Scientific | BP329-500 | |
| Salmon Sperm DNA | VWR international | CA80601-120 | |
| D-Glucose | BioShop Canada | GLU501 | |
| LB Broth LENOX | BioShop Canada | LBL405 | |
| Yeast Nitrogen Base | BioShop Canada | YNB406 | |
| Yeast Extract | BioShop Canada | YEX401 | |
| Peptone | BD Biosciences | 211677 | |
| Bio-Tryptone | BioShop Canada | TRP402 | |
| Adenine Sulphate | BioShop Canada | ADS201 | |
| L-Uracil | BioShop Canada | URA241 | |
| L-Threonine | BioShop Canada | THR002 | |
| L-Histidine | BioShop Canada | HIS200 | |
| L-Methionine | BioShop Canada | MET222 | |
| L-Valine | BioShop Canada | VAL201 | |
| L-Phenylalanine | BioShop Canada | PHA302 | |
| L-Isoleucine | BioShop Canada | ISO910 | |
| L-Tyrosine | BioShop Canada | TYR333 | |
| L-Leucine | BioShop Canada | LEU222 | |
| L-Arginine | BioShop Canada | ARG006 | |
| L-Tryptophane | Fisher Scientific | BP395-100 | |
| L-Lysine | BioShop Canada | LYS101 | |
| L-Alanine | Fisher Scientific | BP369-100 | |
| Agar | BioShop Canada | AGR001 | |
| Soda Lime Galss Beads | Biospec Products | 11079105 | |
| Sodium Chloride | BioShop Canada | SLD002 |
References
- Stagljar, I., Fields, S. Analysis of membrane protein interactions using yeast-based technologies. Trends Biochem Sci. 27 (11), 559-563 (2002).
- Iyer, K. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins. Sci STKE. 275, pl3-pl3 (2005).
- Paumi, C. M. Mapping protein-protein interactions for the yeast ABC transporter Ycf1p by integrated split-ubiquitin membrane yeast two-hybrid analysis. Mol Cell. 26 (1), 15-25 (2007).
- Stagljar, I. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc Natl Acad Sci U S A. 95 (9), 5187-5192 (1998).
- Gisler, S. M. Monitoring protein-protein interactions between the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeast two-hybrid system. Mol Cell Proteomics. 7 (7), 1362-1377 (2008).
- Scheper, W. Coordination of N-glycosylation and protein translocation across the endoplasmic reticulum membrane by Sss1 protein. J Biol Chem. 278 (39), 37998-38003 (2003).
- Thaminy, S. Identification of novel ErbB3-interacting factors using the split-ubiquitin membrane yeast two-hybrid system. Genome Res. 13 (7), 1744-1753 (2003).
- Johnsson, N., Varshavsky, A. Split ubiquitin as a sensor of protein interactions in vivo. Proc Natl Acad Sci U S A. 91 (22), 10340-10344 (1994).
- Kelleher, D. J., Gilmore, R. The Saccharomyces cerevisiae oligosaccharyltransferase is a protein complex composed of Wbp1p, Swp1p, and four additional polypeptides. J Biol Chem. 269 (17), 12908-12917 (1994).
- Chevallier, M. R. Cloning and transcriptional control of a eucaryotic permease gene. Mol Cell Biol. 2 (8), 977-984 (1982).
