

Summary
このアッセイは、交連軸索の向きを変更、シグナル伝達分子、ここに骨形成タンパク質7(BMP7)の能力を評価する。胚の背側脊髄の外植片は、COS細胞の分泌候補増殖因子の集合体に隣接して培養する。外植片の中で成長しているReoriented交連軸索は、免疫組織化学により可視化。
Abstract
脊椎動物の脊髄1の背側交連軸索は軸索ガイダンスシグナルを識別するための貴重なモデルシステムとなっている。ここで、我々は、in vitroアッセイ、交連軸索の方向2に外因性と内因性シグナルの効果を研究するために広く用いられている"方向転換アッセイ"、 で説明します。このアッセイは、ジェーンドッド、トーマスJessellとアンドリューラムズデンの研究室で多数の人々によって開発されました(詳細については、確認応答を参照)、このアッセイのバージョンがでchemorepellent BMPを含む、主要な軸索ガイダンス分子の再配向の活動を示すために使用された屋根板3,4と脊髄での床の板でNetrin1 5とソニックヘッジホッグ(Shh)6 chemoattractive活動。
脊髄の背側三分の二の2-3のセグメントを構成する外植片は、胎生(E)11ラットから切除し、三次元コラーゲンゲル7で培養される。 E11背側脊髄外植片は、糖タンパク質、Tag1に8の彼らの軸索の表現によって識別することができる新たに生まれた交連ニューロンが含まれています。文化30〜40時間かけて、交連軸索の軌跡は、 生体内で見られるのと同様のタイムコースでこれらの背側外植片にrecapitulatedている。この軸索軌道は、テストの組織または背側外植片の側縁の一つと接触して候補シグナル伝達分子を発現するCOS細胞の集合のどちらかを置くことによって挑戦することができます。追加された組織の近傍に延在する交連軸索は、内因性の屋根板と異所性の側面組織からの信号の両方の影響を受けて成長します。交連軸索は、これらの状況下でreorientedされる程度を定量化することができる。このアッセイを使用して、それは両方が交連軸索3,4と同様交連軌道9を指示するためにこの信号のための必要性を新しい方向に向けるために特定の信号の十分性を調べることが可能です。
Protocol
パート1:ハンギングドロップを使用してトランスフェクトしたCOS細胞の集合体の調製
- 種子のCOS - 7(COS)35ミリメートル培養皿中の細胞。彼らは、製造業者のプロトコルに従ってLipofectamine2000を用いて細胞に発現プラスミドの80%コンフルエント、トランスフェ1μgのに到達したとき。
- 吊り滴を準備するには、トランスフェクション培地を吸引し、1Xリン酸緩衝溶液(PBS)を1mlでトランスフェクトしたCOS細胞をすすいでください。 15分間酵素無細胞解離培地の0.5ミリリットルを持つ細胞を扱う。反応を停止するのOpti - MEM + 1Xペニシリン/ストレプトマイシン/グルタミン(P / S / G)+ 10%ウシ胎児血清(FBS)の溶液の1mlを追加します。
- 培養皿の表面からそれらを削除し、15ミリリットルコニカルチューブに移すために細胞をひいて粉にする。細胞をペレットに2000Kで2分間スピン。のOpti - MEM + 1 × P / S / G + 10%FBSの100μlの上清とペレットを再懸濁しますを削除します。
- 皿の底の上に蓋と場所反転、35培養皿のふたの内側に複数の20μlのドロップを特定し、細胞集合体になるまで数時間、37℃のインキュベーターに残す。
パート2:背側脊髄の外植片の調製
- 母親の子宮からE11のラット胚を摘出し、必要になるまでL15培地には氷上に置きます。
- シャープタングステン針で、すぐに前肢芽の下の領域から、それぞれの胚の幹から4〜6セグメントのセクションを削除します。プラスチック製のピペットを使用して、4ウェルNunc社製皿の1つのウェルに組織片を収集し、氷上に置きます。
- すべての胚のセグメントが解剖されているときは、1ミリリットルL15のソリューション+ヌンク料理のほか秒で1mgのディスパーゼに作品を転送するためにプラスチック製のピペットを使用してください。 5分よりもはや室温でインキュベートする。以上ディスパーゼで組織片をインキュベートしないでください。
- インキュベーションの間に、ペトリ皿と混在させる渦にL15の約10ミリリットルに熱不活性化正常ヤギ血清(HIGS)の0.5ミリリットルを追加します。第3のウェルNunc社製シャーレの中にこの溶液を1mlを移し、インキュベーションが終わったときに、この溶液中に組織片を転送する。氷上に置きます。注:組織片が時間氷上に"残り"が許可されている場合、後続の解離が容易になる。
パート3:プライミングコラーゲン
- 360μlコラーゲンに40μlの10倍の最小必須培地を追加して、チューブを軽く叩いたり、簡単にボルテックスで迅速かつ十分に混ぜる。 picofugeで急速にスピンダウン。溶液は黄色になります。 可能な限り氷の上に置かないでください。
- (下記の注を参照)少しオレンジコラーゲン溶液を回すのに十分な0.8M炭酸水素ナトリウム(NaHCO 3を )追加する。再び、フリックで急速に、徹底的にミックス、そしてpicofugeで簡単にスピンダウン。解決策は、混合後のピンク色のままの場合、あなたはあまりにも多くのNaHCO 3を追加し、コラーゲンの新しいチューブを再び開始する必要があります。
- この時点で、コラーゲン溶液が準備されている。それは氷の上に液体のままですが、室温時に持って約5分(ピンクを回す)内に固化される。
- 4ウェルNunc社製ディッシュの各ウェルにおけるコラーゲンのスポット20μlの。あなたのピペットの先端を使用して、小さな"パッド"を形成するコラーゲンを広げ。コラーゲンは常温で設定してみましょう。
注:のNaHCO 3の正確な量は、コラーゲンのバッチごとに滴定する必要があります。 (11μl)、低起動し、その溶液はオレンジ色のわずかな日陰になるまで0.51μlの段階を追加。 "右の"量は、通常、コラーゲンのピンクを回すというのNaHCO 3の最小量よりも少ない1ULです。 NaHCO 3の量は直線的に拡大または縮小していません。
パート4:背側脊髄の外植片とコラーゲンマトリックスにおけるCOSの細胞凝集体とそれらの位置の解剖
- 新鮮なシャープタングステン針と細かいピンセット(#5または#55)を使用して、脊髄を囲む中胚葉を削除します。
- タングステン針で床の板に平行に切断、慎重に脊髄の腹側五分の一を削除する。
- エッジはようにきれいに可能な限りカットしていることを確認し、背側脊髄外植片を二等分する。必要に応じて、エッジを再トリム。約に引っ張らガラスキャピラリーチューブを取り付けた口のピペットを使用して、別のコラーゲンのパッドにそれぞれ背側脊髄外植片を移す。直径0.5ミリメートル。
- 背側脊髄外植片の外側縁とほぼ同じ幅の正方形にトランスフェクトしたCOS細胞の集合体をカット。
- 口のピペットを使用して、背側脊髄外植片とコラーゲンのパッド上にこれらの四角形の1つを転送する。
- 口のピペットで、口のピペットを使用して余分な液体をオフに吸引除去する。吸引以上しないでください。
- パッドの上に下塗りコラーゲンの4μlを適用する、と、タングステン針を用いて、直接組織に触れることなく、外植片を介してコラーゲンを移動する。外植片を周囲のコラーゲンでタングステン針を移動させることにより、東洋植ので、COS細胞の集合体は背側脊髄外植片の側縁部に隣接していること。
- コラーゲンが設定された後、外植片はコラーゲンで完全に覆われていることを保証するためにコラーゲンの4μlのアプリケーションを繰り返す。
- 組織培養フードの中で30〜40時間のOpti - MEM + 1xP/S/Gと文化の0.5ミリリットルを追加します。
- 氷の上にプレートを保つ、1xPBSで0.5ミリリットルを4%パラホルムアルデヒドで45分間外植片を修正。
- 1xPBSのブロッキング溶液+ 0.1%のTriton - X100 +1%HIGS(PBTN)の0.5ミリリットルで2回洗浄。 4のブロックは、℃で時間の少なくともカップルのために、一晩のインキュベーションは免疫組織化学によってプロセスして望ましいとあります。
パート5:免疫組織化学による交連軸索の可視化
- PBTNでブロッキング後、鉗子と48ウェルディッシュの場所で4ウェルプレートの外植片を切り取る。
- 各ウェルに一次抗体の200μlのを追加し、4℃で一晩おきます℃、交連軸索を可視化するために、マウス抗Tag1に抗体の午前1時06ソリューションを使用してください。シグナル伝達分子がタグ付けされたエピトープされている場合、トランスフェクションが成功したかどうかを判断するために、また、テストされているシグナル伝達分子のどちらかに対する一次抗体、または関連するエピトープが含まれています。
- 4でPBTNの5 × 1時間℃で各サンプルを洗う
- 各ウェルに二次抗体の200μlのを追加し、4℃で一晩おきます℃、抗Tag1に抗体を一次抗体として使用された場合、FITCまたはCy3のいずれかに結合したヤギ抗マウスIgM二次抗体を使用してください。 FITCまたはCy3標識に結合され、光に敏感な二次抗体を使用している場合、この時点でからホイルで覆われた48ウェルディッシュを保つ。
- 各サンプル5 4で× 1時間PBTNの各℃を洗う
- 3よくうつ病のスライドに植を転送する、Vectashield媒体とカバーガラスに余分な液体、マウントを取り外します。 4℃または-20℃のいずれかで光金庫に保管
パート6:植の代表画像
成功した実験では、外植片とCOS集計は互いに隣接して保持され、コラーゲンのセットと離れ離れになるではない。軸索の最低限の繁茂があるでしょう。重要な軸索の増殖は、背側脊髄外植片は、長すぎるために培養されたことを示します。外植片からの成長はblebsは存在しません。組織は、培養液に直接接触する場合ブレブ形成が発生します。
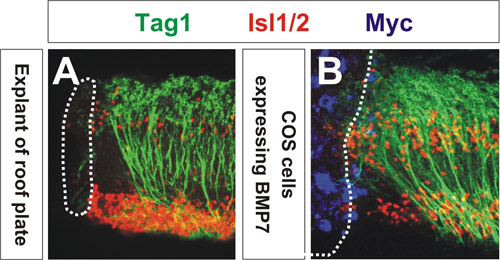
軸索の向きを変更するシグナル伝達分子の能力は、共焦点顕微鏡を用いて評価する。軸索の再配向の程度は、COS細胞の集合体3に最も近い軸索の方向転換の平均角度を測定することによって定量することができます。図1に示すように、COS細胞はmycタグを発現BMP7(青)は、屋根板の外植片が交連軸索(図をはじくための方法に似て離れてBMP7のソース(図1B)からTag1に+交連軸索(緑)を再設定する1A)。コントロールベクターを発現COS細胞は交連軸索3,4の方向には影響を与えません。これらの外植片は、運動ニューロンと背側介在を飾る転写因子ISL1 / 2(赤)、に対する抗体で標識した。この結果は、BMPファミリーのメンバーは、屋根板chemorepellent 3の活性を仲介する最初の兆候だ。

図1:してくださいこちらをクリックして図1の拡大バージョンを参照すること。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
このアッセイを行うに成功したかどうかを判断する重要な要因が最初に、組織があまりにも長時間ディスパーゼで処理されるべきではないされ、そのような治療は、組織が非常に粘着性と生存率の減少となることになります。二、コラーゲンは完全にプライムし、可能な限り氷上で保存する必要があります。それが設定を開始する場合は、処理することはますます困難になる、ピンクターンすなわち。三つは、タングステンの針は常に非常にシャープに保つ必要があります。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
このプロトコルは、ジェーンドッド、トーマスJessellとアンドリューラムズデンの研究室で開発されました。コンラートバスラー、アンCalof、トーマスエドランド、フィルハミルトン、Domna Karagogeos、アリエルルイス私Altabaと俊哉山田を含む多くの人々は、どのようにコラーゲンの脊髄の文化の外植片に決定。 Marysia Placzekとマルクテッシェ-ラヴィーンは、軸索ガイダンスの分子を識別する手段として、生体外植片からで軸索の成長を使用してのテクニックを開拓してきました。バトラーの実験室での作業は、NIH / NINDSからダイムの月とR01 NS063999からの補助金によってサポートされています。
Materials
| Name | Company | Catalog Number | Comments |
| Type 1 rat tail collagen | BD Biosciences | 354236 | |
| COS-7 cells | ATCC | CRL-1651 | |
| 10x Minimal Essential Medium | Invitrogen | 11430-030 | |
| Opti-MEM | Invitrogen | 51985-034 | |
| L15 | Invitrogen | 11415-064 | |
| Lipofectamine 2000 | Invitrogen | 1166-8019 | |
| Fetal bovine serum | Mediatech, Inc. | 35-016-CV | |
| Cell dissociation solution | EMD Millipore | S-004-C | |
| Trypsin 0.25% EDTA | Invitrogen | 2520-0056 | |
| Sodium Bicarbonate | Sigma-Aldrich | S6297 | |
| Triton X-100 | Sigma-Aldrich | X100 | |
| Pen/Strep/Glutamate | Invitrogen | 10378-016 | |
| Paraformaldhyde | Baker/VWR | JTS898-7 | |
| Dispase | Roche Group | 10241750001 | |
| mouse anti Tag1 IgM (4D7) | Developmental Studies Hybridoma Bank | ||
| mouse anti Myc IgG (9E10) | Santa Cruz Biotechnology, Inc. | sc-40 | |
| goat anti mouse IgM FITC | Jackson | 115-095-075 | |
| goat anti mouse Fcy Cy3 | Jackson | 115-165-071 | |
| Goat serum | Invitrogen | 16210064 | |
| Vectashield | Vector Laboratories | H-1000 | |
| 4 well Nunclon dish | Nalge Nunc international | 62407-068 | |
| 3 well depression slide | VWR international | 48339-009 | |
| 48 well dish | Falcon BD | 62406-195 | |
| Aspirator tuve assembly | FHC, Inc. | 30-32-0 | |
| #55 Dumont forceps | Fine Science Tools | 11252-20 | |
| #5 Dumont forcept | Fine Science Tools | 11255-20 | |
| Needle Holder | Fine Science Tools | 26016-12 |
References
- Altman, J., Bayer, S. A. The development of the rat spinal cord. , (1984).
- Placzek, M. Tissue recombinations in collagen gels. Methods in molecular biology. , Springer. Clifton, N.J. 461 vol 325-335 (2008).
- Augsburger, A., Schuchardt, A. BMPs as mediators of roof plate repulsion of commissural neurons. Neuron. 24, 127-141 (1999).
- Butler, S. J., Dodd, J. A role for BMP heterodimers in roof plate-mediated repulsion of commissural axons. Neuron. 38, 389-401 (2003).
- Kennedy, T. E., Serafini, T. Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell. 78, 425-435 (1994).
- Charron, F., Stein, E. The morphogen sonic hedgehog is an axonal chemoattractant that collaborates with netrin-1 in midline axon guidance. Cell. 113, 11-23 (2003).
- Lumsden, A. G., Davies, A. M. Chemotropic effect of specific target epithelium in the developing mammalian nervous system. Nature. 323, 538-539 (1986).
- Dodd, J., Morton, S. B. Spatial regulation of axonal glycoprotein expression on subsets of embryonic spinal neurons. Neuron. 1, 105-116 (1988).
- Yamauchi, K., Phan, K. D., Butler, S. J. BMP type I receptor complexes have distinct activities mediating cell fate and axon guidance decisions. Development. 135, 1119-1128 (2008).

{kind=link}