

Summary
Whole-cell patch-clamp enregistrements de dendrites auditives fibres nerveuses au niveau des synapses du ruban des cellules ciliées internes de la cochlée de mammifère.
Abstract
La synapse afférente entre les cellules ciliées internes (IHC) et la fibre du nerf auditif donne une place électrophysiologiquement accessible pour l'enregistrement de l'activité postsynaptique de la synapse un seul ruban 1-4. Synapses ruban de cellules sensorielles libération du neurotransmetteur en continu, dont le taux est modulé en réponse aux changements notés dans le IHC potentiel de membrane 5. Synapses à ruban ont été montré pour fonctionner par voie de communiqué multivésiculaires, où de multiples vésicules peuvent être libérés simultanément pour évoquer excitateurs courants postsynaptiques (EPSCs) de différentes amplitudes 1, 4, 6-11. Ni le rôle du ruban présynaptique, ni le mécanisme sous-jacent communiqué multivésiculaires est actuellement bien compris.
L'IHC est innervée par 10-20 fibres du nerf auditif, et tous les contacts de fibre l'IHC avec un seul amyéliniques se termine pour former une synapse seul ruban. La petite taille de la CSI boutons afférentes contactant (environ 1 m de diamètre) permet des enregistrements avec une résolution temporelle exceptionnelle à faire. Par ailleurs, la technique peut être adaptée à enregistrer à partir de deux cellules pré-et post-synaptiques simultanément, permettant ainsi la fonction de transfert à la synapse à être étudiés directement 2. Cette méthode offre donc un moyen par lequel les aspects fondamentaux de la neurotransmission peut être étudiée, d'une libération multivésiculaires à la fonction insaisissable du ruban dans les cellules sensorielles.
Protocol
1. Solutions
- Des solutions peuvent être préparées à l'avance. Solutions extracellulaires peuvent être conservés à 4 ° C pendant jusqu'à un mois. Aliquotes de la solution interne ne peut être conservé congelé (-20 ° C). S'assurer que les solutions sont à température ambiante avant de commencer les expériences.
- Solution de dissection et une solution d'enregistrement extracellulaire (mM): 5,8 KCl, 144 NaCl, 0,9 MgCl 2, 1,3 CaCl 2; 0,7 NaH 2 PO 4; 5,6 glucose; HEPES 10; une pyruvate de Na, pH 7,4 (NaOH), à 300 mOsm. Pour l'IHC Ca 2 + l'isolement actuel de la solution extracellulaire peut être modifié comme indiqué: 5,8 KCl, 114 NaCl, 0,9 MgCl 2, 1,3 CaCl 2; 0,7 NaH 2 PO 4; 5,6 glucose; HEPES 10; 30 cl de TEA; pH 7,4 (NaOH ); 300 mOsm.
- 1-2 uM tétrodotoxine (TTX) peut être ajouté à la solution extracellulaire de bloquer les canaux sodiques voltage dépendants et d'isoler les courants postsynaptiques excitateurs ou potentiels (EPSCs ou EPSPS).
- Solution d'enregistrement intracellulaire (mm): 20 KCl, 110 K-méthanesulfonate; 5 MgCl 2, 0,1 CaCl 2, 5 EGTA; HEPES 5, 5 Na 2 ATP; 0,3 Na 2 GTP; 5 Na 2 phoshocreatine; pH 7,2 (KOH), 290 mOsm; ou 135 KCl; 3,5 MgCl 2, 0,1 CaCl 2, 5 EGTA; HEPES 5, 4 Na 2 ATP; 0,2 Na 2 GTP; pH 7,2 (KOH), 290 mOsm. Solution intracellulaire de Ca 2 + IHC isolement actuel: 135 CsMeSO3; 13 cl TEA; 3,5 MgCl 2, 2,5 Na 2 ATP; une EGTA; pH 7,2 (CsOH); 290 mOsm. Cette solution pourrait bloquer une quantité significative des conductances beaucoup plus K +, Ca 2 + en laissant courant restant.
2. Faire Supports pour tissus Disséqué
- Lamelles de modification utilisé pour maintenir la préparation en place pendant les enregistrements peuvent également être préparés à l'avance.
- Appliquez une goutte de Sylgard (Dow Corning, Midland, MI) vers le bord d'une lamelle de verre circulaire (8-12 mm). Placez l'extrémité d'une tige épaisse insectes fines (FST, réf 26002-10) sur la lamelle. Tenez la goupille d'insectes étroitement contre la vitre avec une pince et tenir près d'une bobine chauffée à régler le Sylgard.
3. Fabrication d'électrodes
- Préparer électrodes fraîche sur chaque journée expérimentale et de stocker dans une boîte hermétique. Fabriquer de dix à vingt électrodes pour chaque préparation.
- Sélectionnez électrode de verre, nous utilisons 1 mm de verre borosilicate capillaires (1B100F-4 de WPI, Sarasota, Floride).
- Conception d'un programme sur un extracteur multi-étapes pour tirer des électrodes avec un diamètre de la pointe d'environ 2-3 um (~ 6 MQ dans les solutions décrites ci-dessus). Nous utilisons une Sutter P-87 multi-étapes extracteur horizontale ou verticale Narishige PC-10 extracteur.
- Soigneusement manteau de la tige de l'électrode avec Sylgard aussi proche de la pointe que possible. Cela diminue la capacité d'enregistrement de pipette et le bruit minimise.
- Incendie-polonais électrodes en utilisant un microforge avec un filament chauffé tel que celui disponible à partir WPI. Électrodes polies au feu doivent avoir un diamètre de l'extrémité extérieure d'environ 1 um (10-15 MQ des solutions décrites ci-dessus). L'épaisseur de la paroi pipette est environ 1 / 3 um. Le diamètre extérieur a environ la taille de l'Bouton afférentes à patcher.
- En cas de mise en œuvre des enregistrements simultanés: IHC pipette doit être construit de la même manière (même le verre, le même programme en tirant), avec la distinction que le polissage d'incendie devraient laisser un pourboire de plus grand diamètre d'environ 3 um (6 - 8 MQ).
4. Configuration de l'Expérience
- Remplissez chambres reliées au système de perfusion par gravité avec une solution extracellulaire et des solutions de test contenant des médicaments ou des toxines d'intérêt.
- Set de perfusion de telle sorte que le volume de bain est d'environ 2 ml, constamment perfuser à un taux de l'ordre de 1,5 ml min-1.
- Si différentes solutions sont à appliquer lors de l'expérience avec un système de perfusion locale, remplir les réservoirs avec des solutions. Exécuter des solutions à travers le système et s'assurer qu'il n'ya pas de bulles d'air.
5. Dissection et préparation des échantillons
- Le film accompagne illustre la dissection de l'organe de Corti pendant trois semaines chez des rats âgés (Sprague-Dawley, Charles River), qui est plus difficile que celle des premiers stades postnataux. Avantages et inconvénients de l'enregistrement à partir de rats d'âges différents sont examinés dans la discussion.
- Profondément anesthésier le rat par inhalation d'isoflurane. Lorsque les réflexes de retrait sont absents et les réflexes cornéens sont gravement déprimés, décapiter. Ces procédures ont été approuvées par les soins de la Johns Hopkins University et animale Comité utilisation.
- Retirez le museau et la peau de la tête décapitée. Couper en deux et retirer la tête du cerveau afin d'exposer l'os temporal.
- Retirez les deux os temporaux et les placer dans de la vaisselle propre dissection solution standard contenant extracellulaire.
- Retirerl'os encapsulant l'os temporal afin d'exposer la cochlée. Tenir l'os temporal solidement à la base avec une paire de pinces. Identifier les fenêtres ronde et ovale. Orienter la cochlée afin que la fenêtre ovale et le côté en spirale de la hausse face à la cochlée et éliminent l'os excès autour de la cochlée.
- Retirer l'os encapsulant la cochlée afin d'exposer l'épithélium sensoriel, en prenant soin de protéger la bobine apicale, qui sera utilisé pour l'expérience. Utilisez une deuxième paire de pinces fines carte à puce de l'os directement à partir de la cochlée, en commençant par la zone qui est plus transparent que le reste de l'os. Voici l'os est plus mince et plus facile à enlever. Continuez à enlever l'os autour de la bobine d'apicale.
- Utiliser des micro-dissection des ciseaux pour couper à travers la columelle sous la tour apical. Puis débrancher le tour apical des spires inférieures de la cochlée.
- Utilisez des ciseaux de micro-dissection de nouveau, si nécessaire, pour s'assurer que le tour apical est complètement sectionné. Prenez soin de protéger la bobine apicale, il ne doit pas être tiré ou tendu. Utilisez la pince fine pour amadouer le tour apical du reste de la cochlée.
- Retirer le reste de l'os des deux côtés de la tour apical.
- Retirez délicatement la strie vasculaire, la bande brillante de tissu située à l'extérieur de la région cellules ciliées (Schéma 1). Assurez-vous que pour éviter d'enlever les cellules ciliées sensorielles, qui peuvent facilement se détacher avec la strie vasculaire.
- Utilisez une pince fine pour détacher la membrane tectoriale, le brillant, semi-transparent membrane qui se trouve au-dessus des cellules ciliées sensorielles.
- Maintenant couper l'excès de tissu et d'os et aplatir la préparation avec la pince. Cela est nécessaire, de sorte que le tissu peut être placé uniformément sous une épingle.
- Placer la préparation sous la goupille attachée à une lamelle (préparée précédemment), en prenant soin de positionner la broche loin des cellules de cheveux.
- Utilisez une pince pour le transfert de la lamelle à la chambre d'enregistrement. Assurer le tissu cochléaire est entièrement recouverte d'une goutte de solution extracellulaire tout en transférant la lamelle. Appuyez sur la lamelle fermement sur le fond de verre de la chambre pour s'assurer qu'il ne bouge pas pendant l'enregistrement.
- Commencer immédiatement la perfusion avec une solution extracellulaire, afin d'assurer une meilleure survie de la préparation.
6. Enregistrement
- Repérez la préparation à travers les oculaires du microscope en utilisant les 10x et 40x les objectifs immersion dans l'eau DIC. Orienter la préparation afin que les électrodes d'enregistrement peut approcher CSI orthogonalement à la paroi latérale de l'IHC.
- Si la préparation est enroulée sur, limitant la visibilité de la région basale de CSI, l'utilisation d'une électrode d'enregistrement pour pousser le bord extérieur de la préparation vers le bas contre la lamelle de verre, en prenant soin d'éviter d'appuyer sur les CSI eux-mêmes.
- * Si vous utilisez les jeunes rats, à ce stade de l'épaisse couche de cellules de soutien au-dessus des cellules ciliées peuvent être enlevés avec une pipette de nettoyage (avec un diamètre de pointe de ~ 10-20 um) pour accéder à des cellules sensorielles 1, 12.
- Utilisez le moniteur pour déterminer si le tissu est sain. En utilisant une lentille de grossissement 4X entre microscope et la caméra NC70 Newvicon amplifie encore l'image et des projets une superficie d'environ 4800 um 2 sur le moniteur. Les cellules ciliées devrait être allongée avec des ballots de cheveux intacts. Quand le tissu se détériore, les cellules ciliées gonfler et devenir plus transparent et plus granuleuse.
- Localiser boutons afférentes autour de la base de l'IHC. Boutons sont sphérique ou ellipsoïdale, à environ 1 m de diamètre et sont de couleur claire avec un aspect brillant. La majorité des boutons sont localisés en dessous du niveau du noyau. Lorsque le tissu est malsaine boutons gonflent à environ 4 fois la taille normale et devenir transparente, plutôt que brillant.
- Remplir une électrode d'enregistrement avec une solution intracellulaire, appliquer une pression positive et utiliser le micromanipulateur de manœuvre de l'électrode à la préparation.
- Faire une incision dans la préparation au niveau de la base de l'IHC avec l'électrode. Poussez l'électrode entre la mince couche de cellules de soutien et de la CSI, en utilisant une pression positive pour forger une voie d'accès à la zone à la base de l'IHC. Cette étape est "clé" pour atteindre le bouton afférentes et pour ne pas enregistrer à partir des cellules de soutien que bien ensheath les terminaisons afférentes.
- Retirer délicatement l'électrode et la remplacer par une électrode fraîchement rempli.
- Utiliser une pression positive pour maintenir une extrémité de l'électrode propre, manoeuvre de l'électrode à travers le trou d'accès à la membrane de l'IHC. La pression positive devrait aider à déplacer les cellules de soutien autour de la CSI.
- Déplacez l'électrode dans les cellules voisines de soutien vers le bouton afférents (comme montré dans le film qui l'accompagne. Une autre approche consiste à aborder le bouton afférent en déplaçant unlongtemps la membrane IHC jusqu'à la pointe de l'électrode touche le bouton). S'assurer que l'extrémité de l'électrode est directement en face de l'Bouton afférents (il ya toujours un petit écart entre le bouton et l'électrode en raison de la pression positive sur la pipette). Boutons offrir plus de résistance au mouvement de la pipette que les cellules de soutien et de la membrane IHC; ils peuvent être «ressenti» par l'expérimentateur.
- Déplacer la pipette de haut en bas et de pousser à faire en sorte que les mouvements Bouton. Ceci indique que la pointe de la pipette et Bouton sont dans le même plan Z.
- Avec l'électrode pressée contre le bouton afférent, simultanément relâcher la pression positive et appliquer le vide pour former un joint GQ. Formation d'un joint sur un bouton GQ afférente est similaire à celle sur une cellule de cheveux. Formation de Seal peut être rapide ou peut se produire lentement.
- Appliquer éclate douceur de l'aspiration à la rupture de la membrane à l'intérieur de l'électrode et entrez la configuration de cellule entière enregistrement.
- Si la cellule est un bouton patché afférentes, petite transitoires capacitifs apparaîtra sur l'impulsion carrée d'essai, contrôlée durant la formation d'étanchéité (voir Figure 1). Nous avons estimé la capacité de la fin afférentes à être de 0,4 à 1,8 pF (voir 3, 4). Si la cellule est patché une IHC, les transitoires seront de l'ordre de 3-5 fois plus grande en fonction de la résistance d'accès.
- Pour confirmer que la cellule est une afférentes bouton, exécutez un protocole avec hyperpolarisante et dépolarisants étapes de tension. Les relations actuelles de tension (IV) pour les fibres afférentes et CSI (souvent à tort quand patché visant une bouton afférent) sont caractéristiques pour les types cellulaires individuels et sont présentés dans la figure 2.
- Surveiller le potentiel de membrane; pour une fibre afférente, il est normalement d'environ -60 à -65 mV.
- Si la cellule n'est pas un bouton afférent, retirez l'électrode et répétez les étapes 8 - 13 avec une électrode fraîchement rempli. Utilisez une nouvelle électrode pour chaque tentative.
- Surveiller la résistance série à travers l'enregistrement en appliquant les mesures de tension pour s'assurer que le joint n'est pas refermer. Si la résistance série augmente, il peut être utile d'appliquer des impulsions douces d'aspiration ou de déplacer l'électrode légèrement en arrière. Résistances en série sont généralement autour de 30 MQ. Lorsque l'on analyse l'activité synaptique, nous supprimons les enregistrements avec des résistances en série de plus de 50 MQ.
- Vous pouvez maintenant enregistrer l'activité synaptique (voir figure 3). Soit la fibre afférente présente une activité spontanée, ou la cellule cheveux ont besoin d'être dépolarisée pour activer la libération du transmetteur. Appliquer une solution extracellulaire avec une concentration élevée de potassium (par exemple 40 mm) sera dépolariser les cellules ciliées et souvent d'activer ou d'augmenter le taux de libération de l'émetteur.
- Pour des enregistrements simultanés de l'IHC et de boutons afférentes 2, la procédure devrait être modifiée de la manière suivante: Procéder par étapes 1 à 5. Remplir une pipette IHC avec la solution correspondante intracellulaire de Ca 2 + isolement actuel et suivez comme indiqué en 6. Laisser la pipette pour l'enregistrement de l'IHC en "position d'attente" proche de l'IHC à être enregistrés à partir. Continuer avec les étapes 6 - 14 pour l'enregistrement afférent. Lorsque la pipette afférente est en configuration cellule entière continuer avec la pipette IHC. Manœuvre de la pipette seconde vers le mur latéral de l'IHC correspondante, en maintenant toujours une pression positive. L'IHC devrait montrer une échancrure sur la paroi latérale au moyen de la pression et il doit être veillé à ce que les cellules de soutien sont séparés d'elle. Étapes 12 à 13 pourraient être appliquées pour l'IHC ainsi. Comme indiqué dans 14, par rapport à la fibre afférente, plus transitoires capacitifs sont une caractéristique d'un enregistrement IHC, en plus de la relation caractéristique IV. Une fois les enregistrements simultanés sont établis, un certain temps devraient être autorisés (3 - 5 min, en fonction de la résistance série) pour les solutions intracellulaire de se laver dans l'IHC. Cela se traduira dans les grands courants de Ca 2 + en raison de bloquer l'augmentation de la beaucoup plus grande conductances K +.
- Afin d'effectuer lâche-joint enregistrements extracellulaires, à l'étape 12, plutôt que de faire un joint GQ, faire un joint lâche de 30 à 50 MQ sur le bouton afférent. Ceci peut être réalisé en appliquant moins d'aspiration tout en relâchant la pression positive. Voir Figure 5 pour un exemple d'un enregistrement à partir d'un bouton extracellulaire P21 rat afférentes.
7. Dépannage
- Si les phoques peuvent être formés, mais la transition de la cellule attachée à l'enregistrement de cellules entières ne peuvent pas être atteints, le diamètre interne de la pipette peut être trop étroite.
- Si les joints serrés ne peuvent pas être formé, le diamètre intérieur pipette peut être trop grande et le bouton entier pourrait être aspirée dans l'électrode.
- Si tous les événements synaptiques sont petites et uniformes, l'enregistrement peut être extracellulaire à l'afférentes. Test de retournement des événements au potentiel de membrane positive; intracellulaire enregistréesEPSCs renversera aux potentiels positifs.
- Si résistances en série sont toujours élevés lors patcher terminaisons afférentes, essayez de déplacer la pipette à reculons avant de tenter de percer dans la configuration cellule entière. Cela contribue à éviter le colmatage de pipette et la résistance d'accès élevés.
- Position correcte de la pipette sans crainte. Il est possible de déplacer l'électrode de réduire la résistance série élevée ou obstruer pipette sans "perdre" le sceau sur le afférent. Pendant l'enregistrement simultané création, l'IHC est généralement «poussés» vers le bouton. La position pipette afférents peuvent être corrigées en conséquence.
8. Les résultats représentatifs
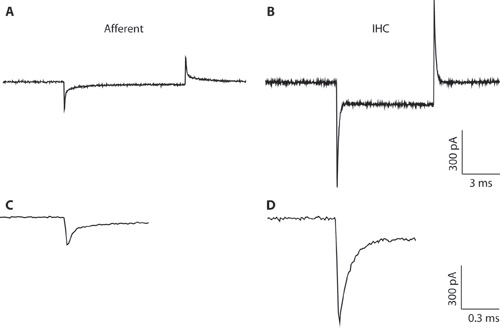
Figure 1. AB. Transitoires typiques enregistrés à partir d'une fibre afférente (A) et IHC (B) en réponse à une mV 10 hyperpolarisante commande de tension d'une tension de tenue de -94 mV. En raison du diamètre pipette étroites et résistance d'accès élevé, l'enregistrement IHC (B) est sous-optimal pour la cellule d'enregistrement toute l'IHC. L'enregistrement est montré ici que pour illustrer la différence entre les transitoires capacitif d'IHC et de fibres afférentes. Cela peut aider à distinguer entre les types cellulaires lors de la formation de la configuration cellule entière. Cellules entières transitoires capacitif d'IHC sont de l'ordre de 5 fois plus grand en amplitude que ceux des fibres afférentes. CD. Transitoires de A à B & montré sur une échelle de temps étendue. C. La décroissance de la réponse afférente peut être adapté aux deux exponentielles. La capacité de la fin afférent a été estimée à partir de la composante rapide.

Figure 2. IV relations à partir d'un bouton afférentes (A) et une IHC (B). Relations IV sont enregistrées à partir d'un potentiel de maintien de -84 mV à pas de tension de -124 mV à + 36 mV par incréments de 10 mV (tension nominale). Les tensions sont indiquées à droite de quelques traces. Ces enregistrements ont été effectués avec 5,8 mM de KCl extracellulaire à température ambiante. Échelle pour les deux: 500 Pa, 200 ms.
Un relations. IV à partir d'une fibre afférente au jour postnatal 19. EPSCs sont présents pendant la majorité des étapes de tension; EPSCs inversée positive à 6 mV. Cet enregistrement a été effectué en présence de TTX pour bloquer la tension fermée Na + courants. Remarque lentement le courant vers l'intérieur à l'activation de tensions hyperpolarisante (I h). Ce courant n'est pas présent dans CSI ou de cellules de soutien et fournit une bonne indication que la cellule a enregistré d'une fibre afférente (voir 3). B. Relations IV d'une IHC P19. En raison du diamètre pipette étroites et résistance d'accès élevé, l'enregistrement n'est pas optimale pour la caractérisation de l'IHC courants et les courants sont plus petits que prévu. L'enregistrement est montré ici que pour montrer les relations IV de CSI et de fibres afférentes peuvent être clairement distingués, quand un enregistrement fibre afférente est tentée. Notez le jeûne activant extérieur courants K + à des potentiels positifs (flèche) suivi par rectification retardée courants K + 13.

Figure 3. Exemplar courants synaptiques enregistrés à partir d'une fibre afférente au jour postnatal 21, en présence de 40 mM K + extracellulaire pour augmenter le taux de libération de l'IHC. La température ambiante, avec le TTX appliquée à bloquer la tension fermée Na + courants. Une. Échelle de 200 pA, 5 ms, notez la taille et de forme variable des EPSCs. Pour une description détaillée des caractéristiques de RPEC voir 4. B. Deux EPSCs marqués d'une (#: multiphasique, o: monophasique) représenté à une échelle élargie: Échelle 100 pA, 1 ms.

Figure 4. Enregistrement simultané d'une IHC et contactant bouton afférentes dans un organe de Corti chez le rat excisée, jour postnatal 10 (voir également 2). Un échelon de tension dépolarisants l'IHC provoque la libération de neurotransmetteurs et active dans le bouton EPSCs afférentes trace supérieure:. Protocole de tension pour la dépolarisation IHC. Tenir potentiels: -79 mV, 50 étapes ms à -29 mV oligo-Orient:. L-type Ca 2 + courants enregistrés par l'IHC montrent typiquement l'inactivation peu et d'activer à des potentiels négatifs traces du bas:. Courants synaptiques dans la fibre afférente en réponse à la dépolarisation IHC. Notez la dépression synaptique lors d'une dépolarisation de 50 ms IHC.

Figure 5. Une. Enregistrement Exemplar extracellulaire à partir d'un bouton afférentes au jour postnatal 21. Cela a été enregistrée à température, avec 5,8 mM K + extracellulaire. Cet enregistrement a un signal typique de bruit pour un enregistrement dans une préparation d'un rat trois semaines vieilles. B. Forme d'onde moyenne pour les événements extracellulaires enregistrées à partir d'un bouton P20 afférentes. C'est la forme d'onde moyenne de 10 272 événements.

Schéma 1. Vue en coupe transversale à travers un tour de la cochlée de rat illustrant la relation anatomique entre les cellules ciliées internes et externes, les ganglions de la spirale, la membrane tectoriale et strie vasculaire.
Discussion
L'étape cruciale dans cette procédure est la dissection. Si le tissu est étiré ou endommagés lors de la dissection, les fibres afférentes ne survivra pas. Tissu de jeunes rats est plus élastique et indulgent. Nous trouvons que les jours postnataux 10 à 11 sont plus faciles à décortiquer et des expériences ont des taux de réussite plus élevé. Un degré significatif de la maturation cochléaire survient après la naissance, avec les rats commencent à entendre de partout jour postnatal 12 14. Par conséquent, à l'âge où la dissection est plus facile, les synapses peuvent ne pas être en pleine maturité 4.
La dissection décrite ici pour le rat est essentiellement le même pour les souris, la principale différence étant la plus petite taille de la cochlée de souris. Cette technique permet aux propriétés des synapses ruban pour être examinés dans des souris transgénique modifié 15. D'autres modifications à cette technique sont: l'ajout d'un colorant fluorescent à la solution intracellulaire aux fibres étiquette 3; enregistrements jumelé avec la cellule ciliée interne pré-et postsynaptiques bouton afférents, permettant à la fonction de transfert entre pré et post-synaptiques des cellules à déterminer 2 et lâche- joint enregistrements extracellulaires à boutons afférentes pour éviter la perte de l'intégrité cellulaire. La configuration d'enregistrement extracellulaire est plus facile à atteindre que la configuration cellule entière et les expériences sont généralement plus durables.
Disclosures
Aucun conflit d'intérêt déclaré.
Acknowledgments
Ce travail a été soutenu par une surdité Research Foundation Research Grant de EY et NIDCD DC006476 à EG et par NIDCD DC005211 au Centre pour l'ouïe et l'équilibre, l'Université Johns Hopkins. Oeuvre d'auteur Tim Phelps, Johns Hopkins University.
LG a écrit le manuscrit initial, EY et LG a filmé la dissection et l'enregistrement. Tous les auteurs ont fourni des chiffres exemplaire et a contribué à la rédaction du manuscrit.
Materials
| Name | Company | Catalog Number | Comments |
| Air table | TMC | ||
| Gibraltar Stage with xy-table | Burleigh | ||
| Axioscope FS2 upright microscope DIC optics Green filter | Carl Zeiss, Inc. | ||
| Newvicon camera with controller | Dage | ||
| Monitor | Dage | ||
| Multiclamp 700B (or similar) | Molecular Devices | ||
| Digidata 1322A (or similar) | Molecular Devices | ||
| Manipulator MP285 | Sutter Instrument Co. | ||
| 6-channel valve application system for local perfusion (used with hand made perfusion pipettes) | Warner Instruments | ||
| PC with acquisition software (PClamp) | Molecular Devices | ||
| Above is the equipment in our electrophysiological setups used for recording from afferent terminals. | |||
References
- Glowatzki, E., Fuchs, P. A. Transmitter release at the hair cell ribbon synapse. Nat Neurosci. 5 (2), 147-154 (2002).
- Goutman, J. D., Glowatzki, E. Time course and calcium dependence of transmitter release at a single ribbon synapse. Proc Natl Acad Sci U S A. 104 (41), 16341-16346 (2007).
- Yi, E., Roux, I., Glowatzki, E. Dendritic HCN channels shape excitatory postsynaptic potentials at the inner hair cell afferent synapse in the mammalian cochlea. J Neurophysiol. 103 (5), 2532-2543 (2010).
- Grant, L., Yi, E., Glowatzki, E. Two modes of release shape the postsynaptic response at the inner hair cell ribbon synapse. J Neurosci. 30 (12), 4210-4220 (2010).
- LoGiudice, L., Matthews, G. The role of ribbons at sensory synapses. Neuroscientist. 15 (4), 380-391 (2009).
- Singer, J. H. Coordinated multivesicular release at a mammalian ribbon synapse. Nat Neurosci. 7 (8), 826-833 (2004).
- Keen, E. C., Hudspeth, A. J. Transfer characteristics of the hair cell's afferent synapse. Proc Natl Acad Sci U S A. 103 (14), 5537-5542 (2006).
- Li, G. L. The unitary event underlying multiquantal EPSCs at a hair cell's ribbon synapse. J Neurosci. 29 (23), 7558-7568 (2009).
- Singer, J. H., Diamond, J. S. Vesicle depletion and synaptic depression at a mammalian ribbon synapse. J Neurophysiol. 95 (5), 3191-3198 (2006).
- Suryanarayanan, A., Slaughter, M. M. Synaptic transmission mediated by internal calcium stores in rod photoreceptors. J Neurosci. 26 (6), 1759-1766 (2006).
- Neef, A. Probing the mechanism of exocytosis at the hair cell ribbon synapse. J Neurosci. 27 (47), 12933-12944 (2007).
- Tritsch, N. X. The origin of spontaneous activity in the developing auditory system. Nature. 450 (7166), 50-55 (2007).
- Kros, C. J., Ruppersberg, J. P. Expression of a potassium current in inner hair cells during development of hearing in mice. Nature. 394 (6690), 281-284 (1998).
- Muller, M. Developmental changes of frequency representation in the rat cochlea. Hear Res. 56 (1-2), 1-7 (1991).
- Seal, R. P. Sensorineural deafness and seizures in mice lacking vesicular glutamate transporter 3. Neuron. 57 (2), 263-275 (2008).
