

Summary
我々はハイスループットな方法で、複数の細胞プロセスと構造を可視化し定量化するために高含有量の画像で自動化された細胞の培養を組み合わせる方法を説明します。このような方法は、ゲノムのさらなる機能アノテーションの助けだけでなく、病気の遺伝子ネットワークや潜在的な創薬ターゲットを識別することができます。
Abstract
ゲノムの機能アノテーション、分子ネットワークと新たな創薬ターゲットの同定の構築は、急務1〜4の問題として対処する必要がある重要な課題です。複数の相補的な"オミクス"アプローチは、多くの神経変性疾患の根底にある遺伝的危険因子と発症メカニズムの手がかりを提供していますが、ほとんどの調査結果は、まだ機能して検証5が必要です。例えば、パーキンソン病(PD)、疾患のリスク因子として同定された多くの新しい遺伝子座が、根本的な原因となる変異(s)または病原性のメカニズムのための最近のゲノムワイド関連研究、6、7を知られていない。それぞれ関連付けられている地域がいくつかの遺伝子を持つことができるので、従来の細胞生物学の手法を用いて疾患と関連する表現型上の遺伝子のそれぞれの機能評価は、時間がかかりすぎるだろう。
分子ネットワークを理解する必要もある、そのリンクそれらが引き起こす表現型への遺伝的変異。それは、病気の表現型が中断されている複数の相互作用の結果であることが期待される。伝統的な分子の方法を使用してこれらのネットワークの再構築は時間がかかるだろう。また、個々のコンポーネントの独立した研究、還元的なアプローチから、ネットワークの予測は、おそらくネットワークの複雑さ8を過小評価される。この過小評価は、部分的に、望ましくない、または毒性の副作用による薬剤の承認の低い成功率を説明することができます。 HT / HC携帯スクリーニングのアプローチを使用して、そしてこれらの経路の中で重要なノードを識別する疾患関連パスウェイのネットワークの視点を得ることは、治療的介入に適している標的の同定につながる可能性があります。
ハイスループットスクリーニング(HTS)は9月12日 、これらの問題に対処するための理想的な方法論です。しかし、従来の方法は、簡単化を使用する一次元の全ウェルの細胞アッセイであったSTICは、複雑な生物学的プロセスのために読み出し。彼らは、同時にそのような形態の特性13、14における軸索輸送障害や変化などの神経変性疾患で見られる多くの表現型を定量化することができませんでした。このアプローチは、細胞のプロセスまたは病原性細胞のサブセットで発生するイベントの動的な性質を調査するために使用することができませんでした。いずれかの多次元表現型に移行しなければならないような機能を定量化するには、高含有量スクリーニング(HCS)4、15-17と呼ばれる。 HCSは、HTSに比べさまざまな摂動に対する細胞応答の詳細な表現を提供する同時に複数のプロセスのセルベースの定量化、です。
HCSは、HTS 18、19に比べて多くの利点がありますが、神経細胞モデルにおけるハイスループット(HT)高含有量(HC)画面を実施することにより、高コスト、環境変動や人為的ミスに問題があります。 "フェノーム"スケールでの細胞応答を検出するためにHCイメージングいずれかを使用すると、感度と再現性を向上させながら、変化およびエラーを削減する必要があります。
ここに我々は正確かつ信頼性の高い神経細胞モデルで自動化された細胞培養20およびHCイメージング法を用いてshRNAの画面を実施する方法を説明します。我々は変異が常染色体劣性パーキンソニズム21を引き起こす一つの特定のタンパク質、DJ1、用モジュレーターを同定するためにこの方法論をどのように使用したかを説明します。
HTの方法でHCイメージングの汎用性を組み合わせることで、それは正確に表現型の茄多を定量化することが可能です。これは、その後だけでなく、病気の発症に関与する経路は、潜在的な治療標的を同定、ゲノムの我々の理解を進めるために利用することができます。
Protocol
1。自動化された接着細胞培養のプロセス(図1)20
- 細胞培養プレートの入力のための自動細胞培養システムを準備します。負荷の消耗品(例えば、ピペットチップ、細胞培養プレート、アッセイプレート)のグラフィカルユーザーインタフェース(GUI)を使用してシステムに。十分な細胞培養培地があることを確認、リン酸塩は、ロボットシステムでは生理食塩水(PBS)とトリプシンをバッファ。
- SH - SY5Y神経芽細胞腫細胞株のプレート当たり2 × 10 6細胞、と手動でシードtwo omnitrayプレート。 10%ウシ胎児血清(FBS)とのOpti - MEMで細胞を維持する。 GUIを使用して細胞培養ロボットにプレートを置く。細胞は37℃、5%CO 2でインキュベートされます。
- 文化のプロトコルが20を開始する必要があるどのセルを選択します。一つは、接着細胞の培養プロセス22からマウスフィーダー細胞23またはサスペンションセルの培養で培養し、胚性幹(ES)細胞の増殖を選択できます。LS 22。
- 接着細胞培養のプロトコール(図1)を選択し、接着細胞株固有のパラメータファイルが合流しきい値(細胞が含まれているomnitray板の面積)は70%に設定されているように調整されていることを確認。 2分に総トリプシン処理の時間を設定します。
- プレート当たり2 × 10 6細胞の播種細胞数で、新しいomnitrayプレートを準備するためにロボットに指示します。
- 自動化された接着細胞培養のプロトコール(図1)を用いて細胞培養系への入力omnitrayプレート。このプロトコルは、次のステップが含まれます:彼らが事前に定義された合流のしきい値に達するまで、プレートをインキュベートし、撮像される。細胞は5測定値内で合流しきい値に到達しない場合、プレートはシステムから削除されます。ユーザー定義の合流のしきい値に達すると、細胞は、洗浄しトリプシン処理し、カウントされます。細胞のあらかじめ定義された番号は、新しいomnitrayプレートに添加し、細胞に十分な数がある場合、指定された麻痺しているアッセイプレートのERは、デッキに運ばれ、細胞の定義数は、各ウェルに分配されています。アッセイプレートは、直接、さらなる処理のためのシステムから統合された顕微鏡または出力を用いて画像化することができます。
- ウェルあたり5,000細胞の総数と、omnitrayプレートあたり4アッセイプレートを準備するためにシステムに指示します。
2。アッセイプレートへのshRNAのウイルス産生とメッキ(所要時間:6日間)
- アンピシリン100μg/ mlの(Sigma - Aldrich社)を含むLuria -ベルターニ媒体のメディアの2ミリリットルで一晩shRNAベクター(オープンバイオシステムズ、TRC1)を含む細菌のグリセロールストックを成長する。
- 製造者のプロトコール(プロメガウィザードMagneSil TFX)以下のプラスミドを抽出します。
- RNAiのコンソーシアムハイスループットレンチウイルス生産(96ウェルプレート)プロトコル24を使用してウイルスを生成。形質導入のために使用されるウイルス粒子は複製欠損しているため、レンチウイルスとの作業は、比較的安全です、分割遺伝子の包装戦略は、その生産に使用されます。しかし、レンチウイルスで作業する場合、追加の生物学的安全性手順では、自分と他人25〜リスクを最小限にするために必要です。すべての実験はMLIIまたはBSL2の安全レベルの実験室で行われなければならない。レンチウイルス粒子と接触しているすべてのプラスチックは(ピペット、プラスチック製の皿、メディア)廃棄前に24時間漂白剤とインキュベートしてください。
- プラスミドpLKO.1 GFPを(Sigma - Aldrich)を用いてGFP陽性細胞の割合を決定することにより、レンチウイルスの感染の多重度(MOI)を計算します。
- MOI 3でアッセイプレートにプレートのレンチウイルス、。
3。レンチウイルス形質導入およびSH - SY5Y細胞(所要時間:6日間)の神経分化
- 細胞をアッセイプレート(ステップ1.7参照)に追加されます。自動細胞培養システムへのshRNAレンチウイルスを含むアッセイ用プレートをロードする。
- 上で24時間後に、メディアアッセイプレートは、分化のプロセスを開始するために0.5%FBS、0.1μMレチノイン酸を含むのOpti - MEMに変更されます。 SH - SY5Y細胞の分化は、神経突起構造の可視化を可能にし、細胞分裂を同期させます。
- 5日間分化のメディアでアッセイプレートのインキュベーションを続ける。これは、標的遺伝子の発現の最大のノックダウンを保証します。
- 5日目に、ミトコンドリアにDJ1の転座を刺激する24時間アッセイプレートの半分に50μMのH 2 O 2を加える。
- 6日目に、ウェル当たり200nMの最終濃度で、細胞にMitoTrackerによりのCmxROS(Invitrogen)を追加し、37℃で30分間。
- 一つは、直接HCイメージャまたはプレートがさらなる処理のためのシステムからエクスポートすることができますを使用して画像板にシステムに指示することができます。
4。アッセイプレートの自動免疫染色(所要時間:2日間)
画質はparamounです高感度かつ信頼性の高いHCSを実施するためのT。不正確なピペッティングによる細胞の単分子層へのダメージは、画質の悪さや再現不可能な結果につながることができます。細胞層の損傷を最小限にするために、免疫染色は、ロボットステーションを用いて行った。プロシージャは、以前に26を説明してきたものと似ていますが、スループットを向上させ、消耗品使用量を減らすためにカスタマイズされています。
- 4%パラフォルムアルデヒド37に予め温めておいた100μlの℃で細胞を固定室温で20分間インキュベートする。
- 5分間、3回PBS200μlで細胞を洗浄。
- PBSで10分間0.1%トリトン(PBST)を含む200μlのアッセイプレートをインキュベートする。
- 5分間、3回PBS200μlで細胞を洗浄。
- 室温で1時間ブロック緩衝液200μl(5%FBSとPBST)でアッセイプレートをインキュベートする。
- 5分間、3回PBS200μlで細胞を洗浄。
- follでインキュベート一晩4により一次抗体° C:
- ヤギDJ1 N20(サンタクルス、5μg/ ml)を
- ウサギβ- IIIチューブリン(Sigma - Aldrich社、1μg/ mlの)
- 次の日に、5分間、3回PBS200μlで細胞を洗浄。
- 室温で1時間、次の二次抗体でアッセイプレートをインキュベートする。
- AlexaFluor 488ロバ抗ヤギ(Invitrogen社、2μg/ ml)を
- AlexaFluor 647ヤギ抗ウサギ(Invitrogen社、2μg/ mlの
- 5分間、3回PBS200μlで細胞を洗浄。
- 10分間、ヘキスト(1μg/ mlのInvitrogen)で培養する。
- 5分間、3回のために200μlのPBSで細胞を洗浄。
- 4℃店舗プレートは° Cまで、それらのイメージを作成することができます。
5。高いコンテンツの画像取得と画像解析(所要時間:5日間)
- 20倍対物レンズを使用しても毎秒30フィールドの全体像。 FIとDJ1を可視化するTCフィルターセット、TRITCフィルターセット、Cy5にフィルタセットとUVフィルターセット(図3)を使用して核をもつβ- IIIチューブリンとミトコンドリア。
- ミトコンドリア内MitoTrackerにより信号の平均強度を決定するためにコンパートメント解析のBioapplication(セロミクス、ThermoFisher)を使って画像を分析する。 (図4B、F)。
- DJ1とミトコンドリア間の平均のオーバーラップ係数を決定するために、セロミクスColocalisationのbioapplication(セロミクス、ThermoFisher)を使って画像を分析する。次のように関心領域(ROI)を定義します。ROI A - 核(図4A、E)、ROI B - ミトコンドリア(図4 B、F)。唯一の細胞質の分析を確実にするためにROI BからROI Aを除外する。ターゲット地域ターゲット地域II(図4C、G)などのIとDJ1としてミトコンドリアを定義します。
- β- IIIチューブリンの染色(図から神経突起の平均長さを追跡するために神経プロファイリングbioapplication(セロミクス、Thermofisher)を使って画像を分析する4D、H)。
- オペラLX自動化された共焦点リーダー(パーキンエルマー)を用いて画像のプレート。水の浸漬で60倍対物レンズを使用してもあたり30フィールドの全体像。 UV励起と561 nmのレーザー、核とミトコンドリア可視化する。
- スポットエッジリッジ(SER)テクスチャ機能のアルゴリズムを使用して画像を分析する。 SER -リッジフィルタでは、尾根のようなパターンを形成するピクセルの輝度を送信します。より断片化されたミトコンドリア、高いSER -リッジスコア(図8)。
6。データの正規化と解析
- Rのソフトウェア環境のBioConductor CellHTS2パッケージ(Rのバージョン2.11.1、BioConductorのバージョン2.6)に画像解析ソフトウェアからデータをインポートします。
- を底とする対数(2)あたりのプレートの中央値ベースの正規化27、28の前にデータを変換する。プレートごとの分散の調整を適用しないでください。
- 表現型の修飾子を識別するために、differeの間に2つ配置ANOVAを使用してくださいNT治療群すなわち感染した未処理の細胞をスクランブル対毒素処理細胞対標的遺伝子未処理の細胞対標的遺伝子処理した細胞を(図5-7)スクランブル。
7。代表的な結果
DJ1内の変異は早期発症、劣性パーキンソニズム21に生じさせるが、それはDJ1の損失は、病気の表現型を生じさせる方法は不明である。それは、DJ1の欠損細胞はより多くの酸化ストレス誘発性細胞死に感受性と酸化ストレス、ミトコンドリア〜29細胞質からDJ1移行し、30に対応していることが知られている。これらの表現型を監視するためにHCのアッセイを構築することによって、我々は、DJ1に関連する表現型を調節するか影響を与える遺伝子を識別することができます。このアプローチは、解読が経路内のDJ1の機能とその疾患の病因に関与することができますすることができます。
DJ1とエピスタシス相互作用(図5)の例:暴露された細胞のDJ1のノックダウン細胞の生存率の大きな損失で毒素の結果に(BAR - B:画像- B)(:画像- BAR -)スクランブルレンチウイルスを感染させた細胞と比較して。標的遺伝子のノックダウンは、DJ1ノックダウン(:画像- C BAR - C)を持つ細胞で観察されたのと同様の効果があります。 DJ1と標的遺伝子の単独遺伝子の損失(:画像- D BAR - D)よりも細胞生存率の有意に高い損失の結果、両方のノックダウン。これはDJ1と標的遺伝子Aの間のエピスタシス相互作用を示唆している
細胞が毒素にさらされ、細胞質からミトコンドリアへのDJ1移行し、DJ1とミトコンドリア(BAR -の間に高いオーバーラップ係数によって定量化される::画像対DJ1転座(図6)を制御する遺伝子の例BAR - C:画像C)。標的遺伝子のBが沈黙している細胞では、以下のミトコンドリアへのDJ1移行して、細胞が毒素にさらされているとき。これは、その標的遺伝子BはミトコンドリアにDJ1の輸送に関与していることを示唆している。 (BAR - B:画像BとBAR - D:画像D)
神経突起(図7)に関与する遺伝子の例:神経突起の長さの大幅な増加で野生型SH - SY5Y細胞の結果の標的遺伝子Cのノックダウン(BAR - B:画像- B)スクランブルを発現するレンチウイルスに感染した細胞に比べてshRNAを(BAR -:画像)。この効果は、毒素(BAR - CおよびD)でインキュベートした細胞では失われている。
ミトコンドリアの形態(図8)に関与する遺伝子の例:に比べてミトコンドリアSER -リッジのセグメンテーションの値(図8、画像CとD)の減少に遺伝子Dの結果をターゲットとしたshRNAと野生型SH - SY5Y細胞の感染細胞は、スクランブルされたレンチウイルス(図8、画像AとB)に感染。
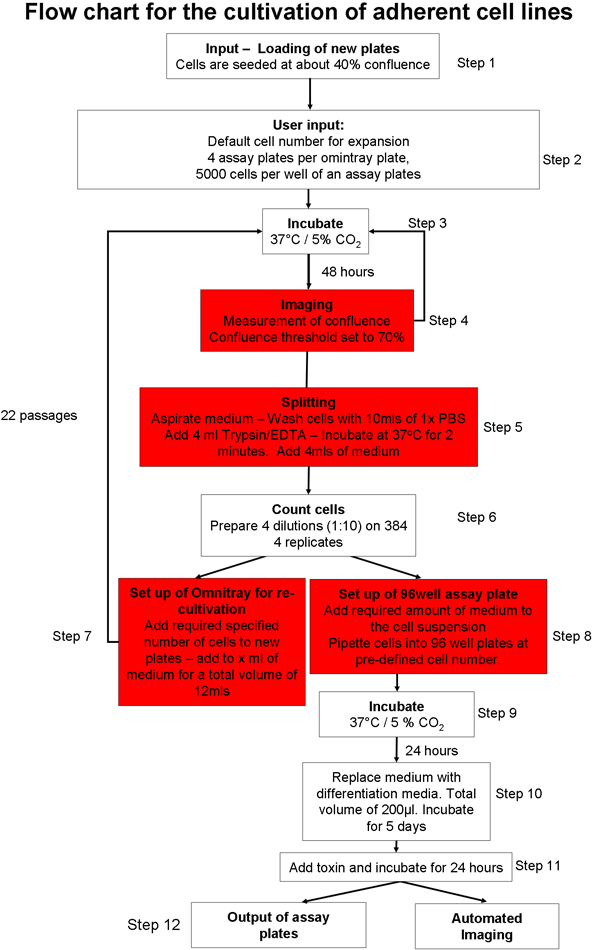
図1自動化された細胞培養プロトコルの概要。。

図2の回路のスクリーニングプロセスの概要を、画像解析やスクリーニングの過程で使用される統計的手法:。彼らはコンフルエントとしたshRNAレンチウイルスを含むアッセイ用プレートに続いてメッキができるまで、i)の細胞を培養している。細胞を5日間分化され、毒素は、24時間プレートに追加されます。アッセイプレートは、システムから出力され、免疫染色。の各プロセスに要する時間の量は、括弧内に示されます。 ⅱ)データは、HCイメージャ(細胞の生存、タンパク質の移行と神経突起伸長)と自動化された共焦点撮像装置(ミトコンドリアの形態)を使用して取得しています。データは、ベースは(2)ログが変換され、正規化、R内CellHTS2パッケージにエクスポートされます。双方向分散分析は、異なる変数の間に有意な相互作用を識別するために使用されます。
図3。HCイメージングによって取得された細胞の合成画像。 A)未処理細胞は、B)細胞は、H 2 O 2で処理した。 DJ1は緑、赤と青の核内ミトコンドリアにラベルが付いています。神経突起の染色が強調表示されていません。

図4。HCSから得られるいくつかの細胞機能の定量化。 AD)未処理SH - SY5Y細胞。 EH)H 2 O 2処理されたSH - SY5Y細胞。 ROI Aの、E)原子核のセグメンテーションと定義、ミトコンドリア、ROI BのB、F)の同定と定量、C、G)DJ1の同定、ターゲットチャンネルII、D、G)の平均神経突起の長さの同定と計算。画像のエッジに近い細胞を解析から除外されます。はめ込み画像は、分析前のイメージです。

図6。DJ1転座を制御する遺伝子の同定(棒上の文字は、画像上のレタリングに対応)。画像はDを介してミトコンドリアへDJ1転座の定量化のために使われたミトコンドリアDJ1(緑)、(赤)と核(青)用のラベルSH - SY5Y細胞である。

図7。神経突起伸長に関与する遺伝子の同定(棒上の文字は、画像上のレタリングに対応)。 A〜Dの画像神経突起の長さの定量化のために使用されたβ- IIIチューブリン(緑)と核(青)用のラベルSH - SY5Y細胞である。

図8。ミトコンドリアの形態調節に関与する遺伝子の同定。画像AとCはそれぞれスクランブルshRNAまたはshRNAを標的遺伝子Dに感染したSH - SY5Y細胞の合成画像です。核は青色で着色されている間、ミトコンドリアは赤色に着色される。画像BとDは、SER -リッジの定量化の可視化です。
Discussion
強力なゲノムワイドな遺伝子機能を変更するためのツールの可用性と結合すれば、HT / HC携帯スクリーニングシステムの減少コスト、と、HT / HCの画面は、学界で一般的になりつつあります。アプローチは、すでに正常にこのような癌9、31から33と胚発生34から36までの創薬ターゲットの同定などの研究の多様な分野に適用され、さらには精神神経疾患37,38に関与する経路を解読するには、アプリケーションの可能性を秘めている。しかし、そのようなシステムの実装は、プロセスの最適化は頻繁に6ヶ月以上を取ると時間と労力の大幅な投資が必要です。このようなトリプシン処理の時間、ピペッティングの速度と播種密度などのすべてのステップは、細胞が健康であることを保証し、一貫して成長する、調整する必要があります。細菌汚染の予防には、C言語で毎週洗浄プロトコルで自動化された細胞培養が直面する最も困難な課題の一つです。コンタミネーションフリーの文化のために必要であることを70%エタノールですべての液体ベアリングラインの洗浄定数とombination。それはまた、より高い分解能と-80のための共焦点顕微鏡などの追加楽器は、化合物の貯蔵℃の冷凍庫を統合できるように、ロボット工学を改善することが必要となります。
遺伝子ネットワークを研究し、病原性の分子経路に関与する遺伝子を同定するために、この方法の感度、速度およびユーティリティを向上させるために対処する必要があるの制限もあります。
HT / HC画面を実施し、信頼性の高いデータが収集されることを保証するために、いくつかの側面は、最適化する必要がある。最初に、測定の信頼性が最優先事項であり、アッセイの堅牢性と感度に依存している。例えば、上記のアッセイでは、画像取得の前に必要な数の処理ステップのために小さい画面に適していますが、ゲノム規模での実装が困難です。このように、一つはダイレクトイメージングを可能と処理ステップ数の減少による減少変化につながるレポーター遺伝子を発現する安定細胞株を構築する必要があります。現時点では、正確に描かれ、確実に関心の表現型を定量アッセイを設計することはHCのスクリーニングプロセスの主要なボトルネックです。
多くの画面は、オフターゲット効果、限られた遺伝子サイレンシングの効率性と不完全なゲノムカバー率に苦しむすべてが別のRNAiライブラリーを使用して、哺乳動物細胞で行われている。したがって、ライブラリは強力な、より具体的であり、より良いカバレッジを持つ行う必要がある。努力はそのような作成が進められているそれはそのような努力がHT / HC screの再現性を向上させることが期待されます ENがヒット。
それらは遺伝的に操作し、比較的容易に大量に培養することができるので、多くの大規模なセルベースの画面の制限は、それらが神経芽細胞腫の細胞で行われていることです。しかしながら、 生体機能のためにex vivoでの細胞培養モデルで識別される"ヒット"の妥当性は、脳が高度に統合されたユニットとして機能するシナプス結合の密と複雑なネットワークを形成し、専門性の高い種類の細胞で構成され、特にとして、疑問である。結果として、それは上述のスクリーニング手法を用いて同定された、追加の技術を使用して、セカンダリ画面にし、より生理的な関連モデル39における検証されてヒットするのが一般的である。 HCS中に同定されたヒット曲の翻訳を改善するために、そのような初代培養細胞や、分化幹細胞または共培養システムなど、より代表的で洗練されたモデルは、HT / HC的なアプローチのために開発され、適応させる必要がある。
自動細胞培養とHCイメージングつの組み合わせは、急速に神経細胞がどのように機能するか新たな洞察を獲得し、経路は病気の開発に重要であるかを判断できるとntent">。それは、他の"オミクス"アプローチから生成された情報をHCS / HTSデータを予定組み合わせるその後、このように治療法の開発を促進、脳疾患のシステム生物学の概要を構築することが可能である。Disclosures
我々は、開示することは何もない
Acknowledgments
我々は、継続的な支援と技術支援のためのエヴァBlaas用ハミルトンのプログラマーや専門家に感謝。この作品は、2つのNWOの投資助成金(911-07-031と40-00506-98-10011)、PrinsesベアトリクスフォンWetenschapsprijs 2009年と神経科学キャンパスアムステルダムによってサポートされていました。SJは、Ti -ファーマによってサポートされています:T5 - 207。
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 9, 580-588 (2009).
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M. High-content analysis in neuroscience. Nat. Rev. Neurosci. 9, 779-788 (2008).
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C. High-throughput in vivo vertebrate screening. Nat. Methods. 7, 634-636 (2010).
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
