

Summary
Vi beskriver en metode til at kombinere automatiseret celle dyrkning med høj-indhold billeddannelse til at visualisere og kvantificere flere cellulære processer og strukturer, i en høj-throughput måde. Sådanne metoder kan hjælpe i den videre funktionelle annotation af genomer samt identificere sygdomsgenet netværk og potentielle lægemiddelkandidater.
Abstract
Den funktionelle annotation af genomer, opførelse af molekylære netværk og nye lægemiddel target identifikation, er vigtige udfordringer, som skal løses som et meget presserende 1-4. Flere komplementære 'omik tilgange har givet fingerpeg om den genetiske risikofaktorer og patogene mekanismer bag talrige neurodegenerative sygdomme, men de fleste fund stadig kræver en funktionel validering 5. For eksempel er en nylig genom bred forening undersøgelse for Parkinsons sygdom (PD), identificeret mange nye loci som risikofaktorer for sygdommen, men den underliggende sygdomsfremkaldende variant (er) eller sygdomsfremkaldende mekanisme ikke er kendt 6, 7. Da hver associeret regionen kan indeholde flere gener, ville den funktionelle evaluering af hver af de gener på fænotyper forbundet med sygdommen, ved hjælp af traditionelle cellebiologi teknikker tager for lang tid.
Der er også behov for at forstå de molekylære netværk, der forbindergenetiske mutationer til fænotyper de forårsager. Det forventes, at sygdommen fænotyper er resultatet af flere interaktioner, der er blevet forstyrret. Rekonstruktion af disse netværk ved hjælp af traditionelle molekylære metoder ville være tidskrævende. Desuden vil netværket forudsigelser fra uafhængige undersøgelser af de enkelte komponenter, reduktionisme tilgang, sandsynligvis undervurderer kompleksitet i nettet 8. Denne undervurdering kan til dels forklare den lave succesrate for godkendelse af lægemidlet på grund af uønskede eller toksiske bivirkninger. Opnåelse af et netværk perspektiv af sygdomsrelaterede veje ved hjælp af HT / HC cellulære screening nærmer sig, og påpege de vigtigste knudepunkter inden for disse veje, kan føre til identifikation af mål, der er mere egnet til terapeutisk intervention.
High-throughput screening (HTS) er en ideel metode til at løse disse spørgsmål 9-12. men traditionelle metoder blev endimensionale hel-såvel celle assays, der anvendes forenklingSTIC aflæsninger for komplekse biologiske processer. De var ude af stand til samtidigt at kvantificere de mange fænotyper observeret i neurodegenerative sygdomme såsom aksonal transport underskud eller ændringer i morfologi egenskaber 13, 14. Denne fremgangsmåde kunne ikke bruges til at undersøge den dynamiske karakter af cellulære processer eller sygdomsfremkaldende hændelser, der opstår i en delmængde af celler. For at kvantificere sådanne funktioner man skal flytte til multi-dimensional fænotyper betegnes høj-indhold screening (HCS) 4, 15-17. HCS er den celle-baserede kvantificering af flere processer samtidigt, hvilket giver en mere detaljeret fremstilling af det cellulære reaktion på forskellige forstyrrelser i forhold til HTS.
HCS har mange fordele frem for HTS 18, 19, men at gennemføre en high-throughput (HT)-højt-indhold (HC) skærmen i neuronal modeller er problematisk på grund af høje omkostninger, miljømæssige variation og menneskelige fejl. For at afsløre cellulære svar på en "phenomics 'skalaved hjælp af HC billeddannelse man nødt til at reducere variation og fejl, og samtidig øge følsomhed og reproducerbarhed.
Heri vi beskriver en metode til præcist og pålideligt gennemføre shRNA skærme ved hjælp af automatiserede celle dyrkning 20 og HC billeddannelse i neuronal cellulære modeller. Vi beskriver, hvordan vi har brugt denne metode til at identificere modulatorer for et bestemt protein, DJ1, som når muteret forårsager autosomal recessive parkinsonisme 21.
Ved at kombinere den alsidighed af HC billeddannelse med HT metoder, er det muligt præcist at kvantificere en overflod af fænotyper. Dette kan efterfølgende bruges til at fremme vores forståelse af genomet, de veje der er involveret i sygdommen patogenese samt identificere potentielle terapeutiske mål.
Protocol
1. Automatiseret Tilhænger cellekultur processen (figur 1) 20
- Forbered automatiserede cellekultur-system for input af cellekultur plader. Load hjælpematerialer (f.eks pipettespidser, cellekultur plader, assay plader) i systemet ved hjælp af den grafiske brugergrænseflade (GUI). Sørg for tilstrækkelig cellekulturmedier, fosfat-bufferet saltvand (PBS) og trypsin i robotsystem.
- Manuelt seed to omnitray plader med 2 x 10 6 celler pr plade, af SH-SY5Y neuroblastomer cellelinje. Bevar celler i Opti-MEM med 10% føtalt bovint serum (FBS). Sæt pladerne i cellekultur robotten ved hjælp af den grafiske brugergrænseflade. Cellerne vil blive inkuberes ved 37 ° C og 5% CO 2.
- Vælg, hvilke cellekultur protokol skal iværksættes 20. Man kan vælge fra en Tilhænger celledyrkningsprocessen 22, dyrkning og udvidelse af embryonale stamceller (ES) celler på musen feeder celler 23 eller dyrkning af suspension CELls 22.
- Vælg den klæbende cellekultur-protokollen (figur 1) og sikre, klæbende cellelinie specifikke parameter filer er tilpasset, så den sammenløbet tærskel (det område af omnitray pladen, der indeholder celler) er sat til 70%. Set samlet trypsinization tid til to minutter.
- Instruer robotten til at forberede nye omnitray plader, med en seeding celle nummer af 2 x 10 6 celler pr plade.
- Input omnitray plader i cellekultur systemet ved hjælp af automatiserede klæbende cellekultur-protokollen (figur 1). Denne protokol omfatter følgende trin: plader inkuberes og filmede, indtil de når de foruddefinerede sammenløbet tærskel. Hvis celler ikke når sammenløbet tærsklen inden for 5 behandlinger, er pladerne fjernet fra systemet. Da de nåede den brugerdefinerede sammenløbet tærskel, der er celler vasket, trypsinized og tælles. En præ-defineret antal celler tilsættes til nye omnitray plader, og hvis der er et tilstrækkeligt antal celler, en bestemt følelsesløseis af test plader transporteres til dæk og et defineret antal celler dispenseres i hver brønd. Assay plader kan direkte filmede med den integrerede mikroskop eller output fra systemet til videre forarbejdning.
- Instruere systemet til at forberede 4-analysen plader pr omnitray plade, med en total på 5.000 celler per brønd.
2. shRNA virus produktion og plettering i assay plader (påkrævet Tid: 6 dage)
- Grow bakteriel glycerol lagre indeholder shRNA vektorer (Open Biosystems, TRC1) natten over i 2 ml Luria-Bertani medium medier, der indeholder 100 mg / ml ampicilin (Sigma-Aldrich).
- Uddrag plasmider følge producentens protokol (Promega Wizard MagneSil TFX).
- Producere virus ved hjælp af RNAi Consortium Højkapacitetsforskning lentiviral Produktion (96 brønds plade) Protokol 24. Arbejde med lentivirus er forholdsvis sikker, fordi den virus partikler, der anvendes til transduktion er replikationskompetente mangelfulde og split-genet emballage strategier bruges til deres produktion. Men når man arbejder med lentivirus, yderligere biosikkerhed procedurer er nødvendige for at minimere risikoen for sig selv og andre 25. Alle forsøg skal udføres i en MLII eller BSL2 sikkerhedsniveau laboratorium. Alle plast (pipetter, plast retter, medier), der har været i kontakt med lentivirus partikler bør inkuberes med blegemiddel i 24 timer før bortskaffelse.
- Beregn mangfoldigheden af infektion (MOI) af lentivirus ved at bestemme den procentdel af GFP positive celler ved hjælp af pLKO.1 GFP plasmid (Sigma-Aldrich).
- Plate lentivirus ind assay plader, med en MOI på 3.
3. Lentiviral transduktion og neuronal differentiering af SH-SY5Y celler (påkrævet Tid: 6 dage)
- Celler føjes til analysen pladerne (se trin 1.7). Load-analysen plader indeholder shRNA lentivirus i den automatiske cellekultur-system.
- Efter 24 timer, medier påanalysen plader vil blive ændret til Opti-MEM indeholdende 0,5% FBS og 0,1 μM retinsyre at starte differentiering processen. Differentiering af SH-SY5Y celler giver mulighed for visualisering af neuritic strukturer og synkroniserer celledeling.
- Fortsæt inkubation af analysen plader i differentieringen medierne i 5 dage. Dette sikrer maksimal Knockdown af target genekspression.
- På dag 5, tilsættes 50 μM H 2 O 2 til halvdelen af analysen plader i 24 timer for at stimulere translokation af DJ1 til mitokondrierne.
- På dag 6, tilføje Mitotracker CmxROS (Invitrogen) til cellerne, i en endelig koncentration på 200 nM per brønd og inkuberes ved 37 ° C i 30 minutter.
- Man kan instruere systemet til billede plader direkte ved hjælp af HC imager eller plader kan eksporteres fra systemet til videre forarbejdning.
4. Automatiseret immunfarvning af test plader (påkrævet Tid: 2 dage)
Billedkvaliteten er paramount for at gennemføre en følsom og pålidelig HCS. Skader på cellerne på grund af unøjagtige pipettering kan føre til dårlig billedkvalitet og irreproducible resultater. For at minimere cellelag skade, blev immunfarvning udført ved hjælp af en robot-station. Proceduren svarer til en, der tidligere har været beskrevet 26, men er blevet tilpasset for at øge kapacitet og reducere forbrugsvarer brug.
- Fix celler med 100 μl på 4% paraformaldehyd forhaand opvarmes til 37 ° C. Inkuber i 20 minutter ved stuetemperatur.
- Vask cellerne med 200 μl af PBS i 5 minutter, 3 gange.
- Inkubér analysen plader med 200 μl af PBS indeholdende 0,1% Triton (PBST) i 10 minutter.
- Vask cellerne med 200 μl af PBS i 5 minutter, 3 gange.
- Inkubér analysen plader med 200 μl af blok buffer (PBST med 5% FBS) i 1 time ved stuetemperatur.
- Vask cellerne med 200 μl af PBS i 5 minutter, 3 gange.
- Inkubér med Follpå grund af primære antistoffer natten over ved 4 ° C:
- Ged DJ1 N20 (Santa Cruz, 5 mg / ml)
- Rabbit β-III tubulin (Sigma-Aldrich, 1 mg / ml)
- Den følgende dag, vaske celler med 200 μl af PBS i 5 minutter, 3 gange.
- Inkubér analysen plader med følgende sekundære antistoffer 1 time ved stuetemperatur:
- AlexaFluor 488 æsel anti-ged (Invitrogen, 2 mg / ml)
- AlexaFluor 647 ged anti-kanin (Invitrogen, 2 mg / ml
- Vask cellerne med 200 μl af PBS i 5 minutter, 3 gange.
- Inkuber celler med Hoechst (Invitrogen, 1 mg / ml) i 10 minutter.
- Vask cellerne med 200 μl PBS i 5 minutter, 3 gange.
- Store plader ved 4 ° C, indtil de kan blive afbildet.
5. Højt indhold Image Acquisition og image analyse (Tidsforbrug: 5 dage)
- Billede i alt 30 felter pr godt bruge 20x objektivlinsen. Visualiser DJ1 med FITC filter indstillet, mitochondrier med TRITC filter sæt, β-III tubulin med Cy5 filter sæt og kernerne ved hjælp af UV-filter sæt (Figur 3).
- Analyser billeder ved hjælp af kompartment analyse Bioapplication (Cellomics, ThermoFisher) for at afgøre den gennemsnitlige intensitet af de Mitotracker signalet i mitokondrierne. (Figur 4B, F).
- For at bestemme den gennemsnitlige overlap koefficient mellem DJ1 og mitokondrier, analysere billeder ved hjælp af Cellomics Colocalisation bioapplication (Cellomics, ThermoFisher). Definer områder af interesse (ROI) som følger: ROI A - nucleus (Figur 4A, E), ROI B - mitokondrierne (Figur 4 B, F). Udeluk ROI A fra ROI B for at sikre analyse af kun cytoplasma. Definer mitokondrierne, som målregionen I og DJ1 som målregion II (figur 4C, G).
- Analyser billeder ved hjælp af Neuronal Profiling bioapplication (Cellomics, Thermofisher) at spore den gennemsnitlige længde af de neurites fra β-III tubulin farvning (Figur4D, H).
- Billede plader ved hjælp af Operaen LX automatiserede konfokal læseren (Perkin-Elmer). Billede i alt 30 felter pr godt bruge 60x objektiv med vand fordybelse. Visualiser mitochondrier med de 561 nm laser og kerner med UV excitation.
- Analyser billeder ved hjælp af Spot-Edge-Ridge (SER) tekstur funktioner algoritme. Den SER-Ridge filteret transmitterer intensitet i pixels danne ridge-lignende mønstre. Jo mere fragmenteret mitokondrierne, jo højere SER-Ridge score (figur 8).
6. Data normalisering og analyse
- Importer data fra billedanalyse software i BioConductor CellHTS2 pakke til R-software miljø (R version 2.11.1, BioConductor version 2.6).
- Logaritmen basen (2) omdanne data, før per plade median baseret normalisering 27, 28. Må ikke anvendes variansen reguleringen pr plade.
- At identificere modifikatorer af en fænotype, bruge en to-vejs ANOVA mellem Forskelnt behandlingsgrupper dvs Scrambled inficerede ubehandlede celler vs scrambled toksin behandlede celler versus target-genet ubehandlede celler versus target gen behandlede celler (figur 5-7).
7. Repræsentative resultater
Mutationer i DJ1 give anledning til tidlig debut-recessive parkinsonisme 21, men det er uklart, hvordan tabet af DJ1 giver anledning til sygdommen fænotype. Det er kendt, at celler mangelfuld af DJ1 er mere modtagelige for oxidativt stress-induceret celledød og som svar på oxidativt stress, DJ1 translocates fra cytoplasmaet til mitokondrier 29, 30. Ved at konstruere HC assays til at overvåge disse fænotyper, kan vi identificere gener, der regulerer eller berører fænotyper forbundet med DJ1. Denne tilgang kan hjælpe dechifrere de veje, inden for hvilken DJ1 funktioner og som kunne være involveret i sygdommen patogenese.
Eksempel på en epistatic interaktion med DJ1 (Figur 5): Knockdown af DJ1 i udsatte cellertil toksin resulterer i et større tab af cellernes levedygtighed (BAR-B: image-B) sammenlignet med celler inficeret med røræg lentivirus (BAR-A: image-A). Knockdown af target-genet A har en lignende effekt, der blev observeret i celler med en DJ1 Knockdown (BAR-C: image-C). Knockdown af både DJ1 og målrette genet A resulterer i et væsentligt større tab af cellernes levedygtighed end tabet af enten gen alene (BAR-D: image-D). Dette tyder på en epistatic samspil mellem DJ1 og målrette gen A.
Eksempel på et gen, der regulerer DJ1 translokationen (Figur 6): Når celler bliver udsat for et giftstof, DJ1 translocates fra cytoplasmaet til mitokondrierne, som er kvantificeret ved en højere overlap koefficient mellem DJ1 og mitokondrier (BAR-A: image En versus BAR-C: image C). I celler, hvor målet genet B er blevet bragt til tavshed, mindre DJ1 translocates til mitokondrier, når cellerne udsættes for toksinet. Dette antyder, at målet genet B er involveret i transport af DJ1 til mitokondrierne. (BAR-B: billede B og BAR-D: image D)
Eksempel på et gen, der er involveret i neuronal udvækst (Figur 7): Knockdown af target-genet C i vildtype SH-SY5Y celler resulterer i en betydelig stigning i neurite længde (BAR-B: image-B) sammenlignet med celler inficeret med lentivirus udtrykke røræg shRNA (BAR-A: billede A). Denne effekt er tabt i cellerne inkuberes med toksin (BAR-C og D).
Eksempel på et gen, der er involveret i mitokondrie morfologi (Figur 8): Infektion af vildtype SH-SY5Y celler med shRNA målrettet genet D resulterer i et fald i mitokondrie SER-Ridge segmentering værdi (fig. 8, Billede C og D) i forhold til celler inficeret med røræg lentivirus (Figur 8, Billede A og B).
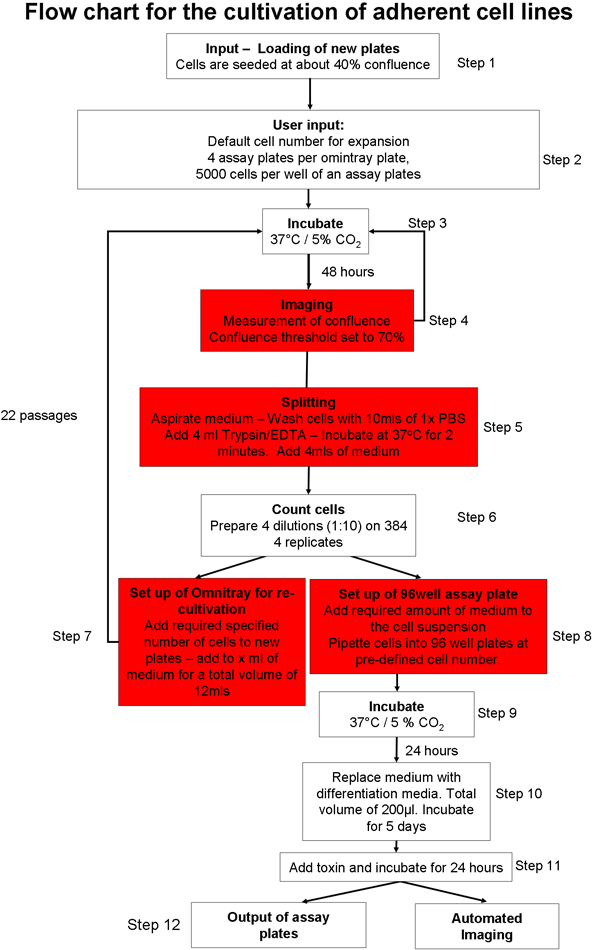
Figur 1 Oversigt over automatiserede cellekultur protokol.:

Figur 2 Skematisk oversigt over den screening, billedanalyse og statistiske metoder, der anvendes under sorterings-processen:. I) Cellerne dyrkes indtil de er sammenflydende og efterfølgende belagt i analysen plader, der indeholder shRNA lentivirus. Celler er differentieret i 5 dage og toksin føjes derefter til pladerne i 24 timer. Assay plader er output fra systemet og immunostained. Mængden af tid, det tager for hver af de processer, er angivet i parentes. ii) Data er erhvervet ved hjælp af et HC imager (celle levedygtighed, protein translokation og neuronal udvækst) og en automatiseret konfokal Imager (mitokondrie morfologi). Data eksporteres til CellHTS2 pakke inden for R, base (2) log transformeret og normaliseret. To-vejs ANOVA bruges til at identificere signifikante interaktioner mellem de forskellige variabler.
Figur 3. Composite billeder af celler overtaget af HC billeddannelse. A) Ubehandlet celler, B) Celler behandlet med H 2 O 2. DJ1 er mærket med grønt, mitokondrierne i rødt og kerner i blå. Neurite farvning ikke er fremhævet.

Figur 4. Kvantificering af flere cellulære funktioner indhentet fra en HCS. AD) Ubehandlet SH-SY5Y celler. EH) H 2 O 2 behandlet SH-SY5Y celler. A, E) Kerner segmentering og definition af ROI A, B, F) Identifikation og kvantificering af mitokondrier, ROI B, C, G) Identifikation af DJ1, Target kanal II, D, G) Identifikation og beregning af den gennemsnitlige neurite længde. Celler tæt på kanten af billedet, er udelukket fra analysen. Inset billeder er billeder inden analyse.

Figur 6. Identifikation af et gen, der regulerer DJ1 translokation (bogstaver på barer svarer til bogstaver på billeder). Billeder fra A til D er SH-SY5Y celler mærket til DJ1 (grøn), mitochondrier (rød) og kerner (blå), som blev brugt til kvantificering af DJ1 translokation til mitokondrierne.

Figur 7. Identifikation af et gen, der er involveret i neuronal udvækst (bogstaver på barer svarer til bogstaver på billeder). Billeder fra A til D er SH-SY5Y celler mærket til β-III tubulin (grøn) og kerner (blå), som blev brugt til kvantificering af neurite længde.

Figur 8. Identifikation af et gen, der er involveret i reguleringen af mitokondrie morfologi. Billede A og C er sammensatte billeder af SH-SY5Y celler inficeret med røræg shRNA eller en shRNA rettet mod gen D hhv. Mitokondrier er farvet røde, mens kerner er farvet i blåt. Billede B og D er visualiseringer af SER-Ridge kvantificering.
Discussion
Med de faldende omkostninger af HT / HC cellulære screening-systemer, kombineret med tilgængeligheden af kraftfulde genom-dækkende værktøjer til at ændre genfunktion, er HT / HC skærme bliver hverdagskost i den akademiske verden. Den fremgangsmåde er allerede blevet anvendt med succes til forskellige forskningsområder såsom identifikation af lægemiddelkandidater inden for kræft 9, 31-33 og embryonale udvikling 34-36 og endda har potentiale til anvendelse i decifrere de veje, der er involveret i neuropsykiatriske lidelser 37,38. Men gennemførelsen af et sådant system kræver en betydelig investering af tid og kræfter med procesoptimering ofte tager mindst 6 måneder. Alle trin, såsom trypsinization gange, pipettering hastigheder og såning tætheder må justeres, for at sikre, at cellerne er sunde og vokse konsekvent. Forebyggelse af bakteriel forurening er en af de vanskeligste udfordringer automatiseret cellekultur med ugentlig rengøring protokoller i Combination med konstant skylning af alle flydende pejlelinjer med 70% ethanol er nødvendigt for forurening gratis kulturer. Det vil også være nødvendigt at forbedre robotteknologi, således at yderligere instrumenter såsom konfokal mikroskoper til højere opløsning og -80 ° C frysere for sammensatte opbevaring kan integreres.
Der er også begrænsninger, der skal løses for at forbedre den følsomme, hastighed og anvendeligheden af denne metode til at studere gen-netværk og identificere gener involveret i sygdomsfremkaldende molekylære veje.
At gennemføre en HT / HC skærmen og sikre, at pålidelige data indsamles, flere aspekter skal optimeres. Første, pålideligheden af målingen er altafgørende, og er afhængig af robusthed og følsomhed af analysen. For eksempel er de analyser, der er beskrevet ovenfor, velegnet til mindre skærme, men er vanskelige at gennemføre på et genom bred skala, på grund af antallet behandling nødvendige skridt til, før billedet overtagelsen.Således ville man være nødt til at konstruere stabile cellelinier der udtrykker reporter genet, hvilket vil give mulighed for direkte billedbehandling og føre til nedsat variation som følge af den reducerede antallet af behandlinger trin. På nuværende tidspunkt, at designe et forsøg, der præcist afbilder og pålideligt kvantificerer en fænotype af interesse er en stor flaskehals i HC screening proces.
Mange skærme er udført i mammale celler ved hjælp af forskellige RNAi biblioteker, som alle lider af ikke-tilsigtede effekter, begrænset gen silencing effektivitet og ufuldstændig genom dækning. Således biblioteker skal gøres, som er mere specifikke, potent og har bedre dækning. Gøres en indsats for at skabe sådanne Det er håbet sådanne bestræbelser vil forbedre reproducerbarheden af HT / HC scre EN hits.
En begrænsning af de mange store celle baserede skærme er, at de er gennemført i neuroblastomceller fordi de kan være genetisk manipuleret og kulturperler til store numre med relativ lethed. Men relevansen af 'hits' identificeret i ex vivo cellekulturer modeller til in vivo-funktionen er tvivlsom, især da hjernen består af højt specialiserede celletyper, der danner en tæt og kompliceret netværk af synaptiske forbindelser til fungere som en stærkt integreret enhed. Som følge heraf er det almindeligt, at rammer er identificeret ved hjælp af screening metode der er beskrevet ovenfor, er valideret i sekundære skærme ved hjælp af yderligere teknikker og i mere fysiologisk relevante modeller 39. For at forbedre oversættelsen af hits identificeret under HCS, mere repræsentative og avancerede modeller, som f.eks primærelementer og differentierede stamceller eller co-kultur systemer skal udvikles og tilpasses til HT / HC tilgange.
ntent "> Med en kombination af automatiserede celle dyrkning og HC billedbehandling kan man hurtigt få ny indsigt i, hvordan neuroner funktion og afgøre, hvilke veje der er vigtige for udvikling af sygdom. kombinere HCS / HTS-data med oplysninger, der genereres fra andre 'omik tilgange, det vil så være muligt at konstruere en systembiologi overblik over hjernens sygdomme, hvilket vil lette terapeutisk udvikling.Disclosures
Vi har intet at videregive
Acknowledgments
Vi takker Hamilton programmører og specialister til fortsat støtte og Eva Blaas til teknisk bistand. Dette arbejde blev støttet af to NWO Investeringstilskud (911-07-031 og 40-00506-98-10011), The Prinses Beatrix Fonds Wetenschapsprijs 2009 og Neuroscience Campus Amsterdam SJ er støttet af Ti-Pharma: T5-207.
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 9, 580-588 (2009).
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M. High-content analysis in neuroscience. Nat. Rev. Neurosci. 9, 779-788 (2008).
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C. High-throughput in vivo vertebrate screening. Nat. Methods. 7, 634-636 (2010).
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
