

Summary
We beschrijven een methodiek combineert geautomatiseerde cel kweken met een hoog-content imaging te visualiseren en te kwantificeren meerdere cellulaire processen en structuren, in een high-throughput manier. Dergelijke methoden kunnen helpen bij de verdere functionele annotatie van genomen en te identificeren ziektegen netwerken en potentiële drug targets.
Abstract
De functionele annotatie van genomen, de bouw van de moleculaire netwerken en nieuwe drug target identificatie, zijn belangrijke uitdagingen die moeten worden aangepakt als een zaak van grote urgentie 1-4. Meerdere complementaire 'omics' benaderingen hebben aanwijzingen met betrekking tot de genetische risicofactoren en pathogene mechanismen die ten grondslag liggen aan tal van neurodegeneratieve ziekten, maar de meeste bevindingen moeten nog functionele validering 5. Bijvoorbeeld, is een recent genoom associatie studie voor de ziekte van Parkinson (PD), ook tal van nieuwe loci als risicofactoren voor de ziekte, maar de onderliggende veroorzakende variant (s) of pathogene mechanisme niet bekend is zes, zeven. Aangezien elke regio geassocieerd kan bevatten meerdere genen, zou de functionele evaluatie van elk van de genen op fenotypes geassocieerd met de ziekte, met behulp van traditionele celbiologische technieken te lang duren.
Er is ook een noodzaak om de moleculaire netwerken te begrijpen die verwijzengenetische mutaties aan de fenotypes die ze veroorzaken. De verwachting is dat de ziekte fenotypes zijn het gevolg van meerdere interacties die zijn verstoord. Reconstructie van deze netwerken met behulp van traditionele moleculaire methoden zou tijdrovend zijn. Bovendien zal het netwerk voorspellingen van onafhankelijke studies van de individuele componenten, het reductionisme aanpak, waarschijnlijk een onderschatting van de complexiteit van het netwerk 8. Deze onderschatting kan voor een deel te verklaren het lage slagingspercentage van de goedkeuring van geneesmiddelen als gevolg van ongewenste of toxische bijwerkingen. Het verkrijgen van een netwerk perspectief van ziekte gerelateerde paden met behulp van HT / HC-cellulaire screening benaderingen, en het identificeren van de belangrijkste knooppunten binnen deze paden, zou kunnen leiden tot de identificatie van targets die meer geschikt zijn voor therapeutische interventie.
High-throughput screening (HTS) is een ideale methode om 9-12 aanpakken van deze problemen. maar de traditionele methodes werden eendimensionale whole-cell assays goed, dat vereenvoudigd wordt gebruiktstic uitlezingen voor complexe biologische processen. Ze waren niet in staat om gelijktijdig kwantificeren van de vele fenotypes waargenomen in neurodegeneratieve ziekten zoals axonaal transport tekorten of wijzigingen in morfologie eigenschappen van 13, 14. Deze aanpak kan niet worden gebruikt om het dynamische karakter van cellulaire processen of pathogene gebeurtenissen die zich voordoen in een subset van cellen te onderzoeken. Te kwantificeren dergelijke functies moet men om te verhuizen naar multi-dimensionale fenotypes genoemd high-content screening (HCS) 4, 15-17. HCS is de cel-gebaseerde kwantificering van meerdere processen tegelijk, die een meer gedetailleerde weergave van de cellulaire respons op verschillende verstoringen biedt ten opzichte van HTS.
HCS heeft veel voordelen ten opzichte van HTS 18, 19, maar het uitvoeren van een high-throughput (HT)-high-inhoud (HC) scherm in neuronale modellen is problematisch vanwege de hoge kosten, milieu-variatie en menselijke fouten. Om de cellulaire respons te detecteren op een 'phenomics' schaalmet behulp van HC beeldvorming moet men variatie en fouten te verminderen, terwijl het verhogen van de gevoeligheid en reproduceerbaarheid.
Hierin beschrijven we een methode om nauwkeurig en betrouwbaar gedrag shRNA schermen met behulp van geautomatiseerde cel-kweken van 20 en HC imaging in neuronale cellulaire modellen. Beschrijven we hoe we deze methodologie gebruikt om modulatoren te identificeren voor een bepaald eiwit, DJ1, die, wanneer gemuteerd veroorzaakt autosomaal recessief parkinsonisme 21.
De combinatie van de veelzijdigheid van de HC-beeldvorming met HT methoden, is het mogelijk om nauwkeurig te kwantificeren een overvloed aan fenotypes. Dit kan vervolgens worden gebruikt om ons begrip van het genoom, de routes die betrokken zijn bij de ziekte pathogenese vooraf evenals het identificeren van potentiële therapeutische doelen.
Protocol
1. Geautomatiseerde hechtende celcultuur proces (figuur 1) 20
- Bereid de geautomatiseerde celkweek systeem voor de invoer van celkweek platen. Belasting verbruiksgoederen (bijv. pipetpunten, celkweek platen, assay platen) in het systeem met behulp van de grafische user interface (GUI). Zorg dat er voldoende media voor celkweek, fosfaat gebufferde zoutoplossing (PBS) en trypsine in het robotsysteem.
- Handmatig zaad twee omnitray platen met 2 x 10 6 cellen per plaat, van de SH-SY5Y neuroblastoma cellijn. Behouden cellen in Opti-MEM met 10% foetaal bovine serum (FBS). Leg de platen in de celkweek robot met behulp van de GUI. De cellen worden geïncubeerd bij 37 ° C en 5% CO 2.
- Kies welke celcultuur-protocol dient te worden gestart 20. Men kan kiezen uit een aanhanger celcultuur-22, het kweken en uitbreiding van de embryonale stamcellen (ES) cellen op de muis feeder cellen 23 of kweken van de opschorting cells 22.
- Selecteer het aanhangend celkweek protocol (figuur 1) en zorg ervoor adherente cellijn specifieke parameter-bestanden zijn zo aangepast dat de samenvloeiing drempel (het gebied van de omnitray plaat die cellen bevat) is vastgesteld op 70%. Stel de totale behandeling met trypsine tijd voor twee minuten.
- Instrueer de robot om nieuwe omnitray platen voor te bereiden, met een seeding cel nummer van 2 x 10 6 cellen per plaat.
- Input omnitray platen in de celkweek systeem met behulp van de geautomatiseerde hechtende celkweek protocol (figuur 1). Dit protocol omvat de volgende stappen: platen worden geïncubeerd en afgebeeld totdat ze de vooraf gedefinieerde samenvloeiing drempel. Als cellen niet de samenvloeiing drempel niet bereiken binnen vijf lezingen, worden de platen uit het systeem verwijderd. Bij het bereiken van de door de gebruiker gedefinieerde samenvloeiing drempel, worden de cellen gewassen, getrypsiniseerd en geteld. Een vooraf bepaald aantal van de cellen worden toegevoegd om nieuwe omnitray platen en of er voldoende aantallen cellen, een bepaald gevoellozeer van assay platen worden vervoerd naar het dek en een bepaald aantal cellen afgegeven in elk putje. Assay platen kunnen direct worden afgebeeld met behulp van de geïntegreerde microscoop of-uitgang van het systeem voor verdere verwerking.
- Instrueer het systeem 4 assay platen per omnitray plaat voor te bereiden, met een totaal van 5.000 cellen per well.
2. shRNA virus productie en plating in de assay platen (verplicht Tijd: 6 dagen)
- Grow bacteriële glycerol voorraden die de shRNA vectoren (Open Biosystems, TRC1), overnachting in 2 ml Luria-Bertani medium medium met 100 ug / ml van ampicilin (Sigma-Aldrich).
- Extract plasmiden volgende protocol van de fabrikant (Promega Wizard MagneSil TFX).
- Produceren virus met behulp van de RNAi Consortium high-throughput Lentivirale Production (96 wells-plaat) protocol 24. Het werken met lentivirus is relatief veilig, omdat het virus deeltjes gebruikt worden voor transductie zijn replicatie-deficiënt en split-gen verpakking strategieën worden gebruikt voor de productie. Echter, bij het werken met lentivirus, extra bioveiligheid procedures nodig zijn om het risico voor zichzelf en anderen 25 te minimaliseren. Alle experimenten moeten worden uitgevoerd in een MLII of BSL2 veiligheidsniveau laboratorium. Alle kunststoffen (pipetten, plastic borden, media) die in contact zijn geweest met lentivirus deeltjes mogen worden geïncubeerd met bleekwater voor 24 uur voorafgaand aan de beschikking.
- Bereken de multipliciteit van infectie (MOI) van de lentivirus door het bepalen van het percentage GFP-positieve cellen met behulp van de pLKO.1 GFP plasmide (Sigma-Aldrich).
- Plaat lentivirus in de assay platen, met een MOI van 3.
3. Lentivirale transductie en neuronale differentiatie van SH-SY5Y cellen (Benodigde tijd: 6 dagen)
- Cellen worden toegevoegd aan de assay platen (zie stap 1.7). Laad assay platen met shRNA lentivirus in het geautomatiseerde celcultuur.
- Na 24 uur, de media op deassay platen zullen worden veranderd in Opti-MEM die 0,5% FBS en 0,1 uM retinoinezuur aan de differentiatie te starten. Differentiatie van SH-SY5Y cellen maakt visualisatie van neuritische structuren en synchroniseert de celdeling.
- Ga door incubatie van de test platen in de differentiatie media voor 5 dagen. Dit zorgt voor maximale knockdown van target-gen expressie.
- Op dag 5, voeg 50 uM H 2 O 2 de helft van de assay platen gedurende 24 uur tot translocatie van DJ1 te stimuleren om de mitochondriën.
- Op dag 6, toe te voegen Mitotracker CmxROS (Invitrogen) aan de cellen, bij een uiteindelijke concentratie van 200 nM per putje en incubeer bij 37 ° C gedurende 30 minuten.
- Men kan instrueren van het systeem om de afbeelding platen direct met de HC-imager of platen kunnen geëxporteerd worden uit het systeem voor verdere verwerking.
4. Geautomatiseerde immunokleuring van assay platen (verplicht Tijd: 2 dagen)
De beeldkwaliteit is paramount voor het uitvoeren van een gevoelige en betrouwbare HCS. Schade aan de cellulaire monolaag als gevolg van onjuiste pipetteren kan leiden tot slechte beeldkwaliteit en reproduceerbare resultaten. Om de cellaag schade te minimaliseren, werd de immunokleuring uitgevoerd met behulp van een robot station. De procedure is vergelijkbaar met een die al eerder is beschreven, maar 26 is aangepast om de doorvoer te verhogen en verbruiksgoederen het gebruik te verminderen.
- Fix cellen met 100 pi van 4% paraformaldehyde voorverwarmd tot 37 ° C. Incubeer gedurende 20 minuten bij kamertemperatuur.
- Was de cellen met 200 pi PBS gedurende 5 minuten, 3 keer.
- Incubeer de assay platen met 200 pi PBS met 0,1% Triton (PBST) voor 10 minuten.
- Was de cellen met 200 pi PBS gedurende 5 minuten, 3 keer.
- Incubeer de assay platen met 200 ul van blok buffer (PBST met 5% FBS) gedurende 1 uur bij kamertemperatuur.
- Was de cellen met 200 pi PBS gedurende 5 minuten, 3 keer.
- Incubeer met de evdoor primaire antilichamen overnacht bij 4 ° C:
- Geit DJ1 N20 (Santa Cruz, 5 ug / ml)
- Konijn β-tubuline III (Sigma-Aldrich, 1 ug / ml)
- Op de volgende dagen, wassen cellen met 200 pi PBS gedurende 5 minuten, 3 keer.
- Incubeer de assay platen met de volgende secundaire antilichamen 1 uur bij kamertemperatuur:
- AlexaFluor 488 ezel anti-geit (Invitrogen, 2 ug / ml)
- AlexaFluor 647 geit anti-konijn (Invitrogen, 2 pg / ml
- Was de cellen met 200 pi PBS gedurende 5 minuten, 3 keer.
- Incubeer de cellen met Hoechst (Invitrogen, 1 ug / ml) gedurende 10 minuten.
- Was de cellen met 200 ul PBS gedurende 5 minuten, 3 keer.
- Bewaar platen bij 4 ° C totdat ze kunnen worden afgebeeld.
5. Hoog gehalte beeldacquisitie en beeldanalyse (Duur: 5 dagen)
- Beeld een totaal van 30 velden per goed gebruik van de 20x objectief. Visualiseer DJ1 met de FITC filter set, de mitochondriën met het TRITC filter set, β-III tubuline met de Cy5 filter set en de kernen met behulp van de UV-filter set (figuur 3).
- Analyseren van de beelden met behulp van de compartimentale analyse Bioapplication (Cellomics, ThermoFisher) om de gemiddelde intensiteit van de Mitotracker signaal binnen de mitochondria te bepalen. (Figuur 4B, F).
- Voor het bepalen van de gemiddelde overlap coëfficiënt tussen DJ1 en de mitochondria, het analyseren van de beelden met behulp van de Cellomics Colocalisation bioapplication (Cellomics, ThermoFisher). Definieer regio's van belang (ROI) als volgt: ROI A - kern (Figuur 4A, E), ROI B - mitochondriën (Figuur 4 B, F). Uit te sluiten ROI A van ROI B tot analyse van alleen het cytoplasma te verzekeren. Definieer de mitochondriën als doel regio I en DJ1 als doel regio II (figuur 4C, G).
- Analyseren van de beelden met behulp van de neuronale Profiling bioapplication (Cellomics, Thermofisher) om de gemiddelde lengte van de neurieten trace van de β-III tubuline vlekken (Figuur4D, H).
- Afbeelding platen met behulp van de Opera LX geautomatiseerde confocale lezer (Perkin-Elmer). Beeld een totaal van 30 velden per goed gebruik van de 60x objectief met onderdompeling in water. Visualiseer mitochondria met de 561 nm laser en kernen met UV-excitatie.
- Analyseren van de beelden met behulp van de Spot-Edge-Ridge (SER) textuur functies algoritme. De SER-Ridge filter stuurt intensiteit in pixels vormen van nok-achtige patronen. Hoe meer gefragmenteerd de mitochondria, hoe hoger de SER-Ridge score (figuur 8).
6. Data normalisatie en analyse
- Importeer de gegevens van de beeldanalyse software in de Bioconductor CellHTS2 pakket voor de R-software-omgeving (R versie 2.11.1, Bioconductor versie 2.6).
- Logaritme basis (2) om te zetten van de gegevens voorafgaand aan de mediaan per plaat op basis van normalisatie 27, 28. Niet van toepassing de variantie aanpassing per plaat.
- Het identificeren van factoren van een fenotype, gebruik dan een twee weg ANOVA tussen de different behandeling groepen, dwz Scrambled geïnfecteerde onbehandelde cellen vs scrambled gif behandelde cellen versus target-gen ten opzichte van onbehandelde cellen target-gen behandelde cellen (figuren 5-7).
7. Representatieve resultaten
Mutaties binnen DJ1 aanleiding geven tot een vroeg begin-recessief parkinsonisme 21, maar het is onduidelijk hoe het verlies van DJ1 aanleiding geeft tot de ziekte fenotype. Het is bekend dat cellen tekort van DJ1 zijn gevoeliger voor oxidatieve stress-geïnduceerde celdood en in reactie op oxidatieve stress, DJ1 translokeert van het cytoplasma naar de mitochondriën 29, 30. Met de bouw van HC assays op deze fenotypes te controleren, kunnen we identificeren genen die reguleren of te beïnvloeden fenotypes geassocieerd met DJ1. Deze benadering kan helpen bij het ontcijferen van de trajecten waarbinnen DJ1 functies en die kunnen worden betrokken bij de ziekte pathogenese.
Voorbeeld van een epistatisch interactie met DJ1 (figuur 5): Knockdown van DJ1 in blootgestelde cellenvan toxines resulteert in een groter verlies van de levensvatbaarheid van de cellen (BAR-B: image-B) in vergelijking met cellen die geïnfecteerd zijn met gecodeerde lentivirus (BAR-A: image-A). Knockdown van target-gen A heeft een vergelijkbaar effect met die waargenomen in cellen met een DJ1 knockdown (BAR-C: image-C). Knockdown van zowel DJ1 en target-gen A resulteert in een significant grotere verlies van de levensvatbaarheid van de cel dan het verlies van beide genen alleen (BAR-D: image-D). Dit suggereert een epistatisch interactie tussen DJ1 en target-gen A.
Voorbeeld van een gen reguleren DJ1 translocatie (Figuur 6): Wanneer de cellen worden blootgesteld aan een toxine, DJ1 translokeert van het cytoplasma naar de mitochondriën, die wordt gekwantificeerd door een hogere overlap coëfficiënt tussen DJ1 en de mitochondriën (BAR-A: image A versus BAR-C: image C). In cellen waar de target-gen B is het zwijgen opgelegd, minder DJ1 translokeert aan de mitochondria wanneer de cellen worden blootgesteld aan het gif. Dit suggereert dat doelwitgen B is betrokken bij het transport van DJ1 naar de mitochondriën. (BAR-B: afbeelding B en BAR-D: image D)
Voorbeeld van een gen betrokken bij neuronale uitgroei (figuur 7): Knockdown van target-gen C in wild type SH-SY5Y cellen resulteert in een significante toename van neurieten lengte (BAR-B: image-B) in vergelijking met cellen die geïnfecteerd zijn met lentivirus uiten van gecodeerde shRNA (BAR-A: image A). Dit effect verloren gaat in cellen geïncubeerd met toxine (BAR-C en D).
Voorbeeld van een gen betrokken bij de mitochondriale morfologie (figuur 8): Infectie van het wild type SH-SY5Y cellen met shRNA richten gen D leidt tot een afname in het mitochondriaal SER-Ridge segmentatie waarde (figuur 8, afbeelding C en D) in vergelijking met cellen die geïnfecteerd zijn met gecodeerde lentivirus (figuur 8, Image A en B).
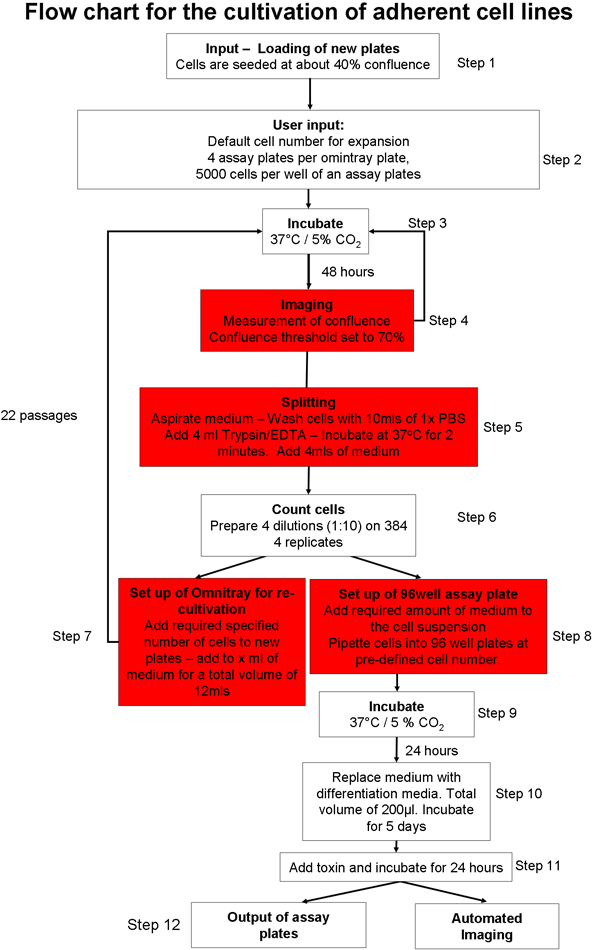
Figuur 1 Overzicht van geautomatiseerde celkweek protocol.:

Figuur 2 Schematisch overzicht van de screening, beeld analyse en statistische methoden die worden gebruikt tijdens de screening proces:. I) De cellen worden gekweekt tot ze confluent en vervolgens uitgeplaat in de assay platen met de shRNA lentivirus. Cellen worden gedifferentieerd voor 5 dagen en toxine wordt vervolgens toegevoegd aan de platen voor 24 uur. Assay platen zijn output van het systeem en immunostained. De hoeveelheid tijd die voor elk van de processen tussen haakjes aangegeven. ii) gegevens worden verkregen met behulp van een HC-imager (levensvatbaarheid van de cellen, eiwit translocatie en neuronale uitgroei) en een geautomatiseerde confocale imager (mitochondriale morfologie). De gegevens worden geëxporteerd naar CellHTS2 pakket binnen R, basis (2) log-getransformeerde en genormaliseerd. Two-way ANOVA wordt gebruikt om significante interacties tussen de verschillende variabelen te identificeren.
Figuur 3. Samengestelde beelden van cellen verkregen door HC beeldvorming. A) onbehandelde cellen, B) cellen behandeld met H 2 O 2. DJ1 is gelabeld in het groen, de mitochondria in het rood en de kernen in het blauw. Neurieten kleuring is niet gemarkeerd.

Figuur 4. Kwantificering van verschillende cellulaire functies verkregen uit een HCS. AD) Onbehandeld SH-SY5Y cellen. EH) H 2 O 2 behandeld SH-SY5Y cellen. A, E) Kernen segmentatie en de definitie van ROI A, B, F) Identificatie en kwantificering van de mitochondria, ROI B, C, G) Identificatie van DJ1, Target kanaal II, D, G) Identificatie en berekening van de gemiddelde lengte van neurieten. Cellen dicht bij de rand van het beeld zijn uitgesloten van de analyse. Inzet beelden zijn beelden voorafgaand aan de analyse.

Figuur 6. Identificatie van een gen reguleren DJ1 translocatie (letters op de balken komen overeen met de belettering op de beelden). Afbeeldingen A tot D zijn SH-SY5Y cellen gelabeld voor DJ1 (groen), mitochondria (rood) en kernen (blauw) die werden gebruikt voor de kwantificering van DJ1 translocatie naar de mitochondriën.

Figuur 7. Identificatie van een gen betrokken bij neuronale uitgroei (letters op de balken komen overeen met de belettering op de beelden). Afbeeldingen A tot D zijn SH-SY5Y cellen gelabeld voor β-tubuline III (groen) en kernen (blauw), die werden gebruikt voor de kwantificering van neurieten lengte.

Figuur 8. Identificatie van een gen dat betrokken is bij het reguleren van mitochondriale morfologie. Afbeelding A en C zijn samengestelde beelden van de SH-SY5Y cellen die geïnfecteerd zijn met scrambled shRNA of een shRNA targeting-gen D respectievelijk. Mitochondriën zijn gekleurd in het rood terwijl de kernen zijn gekleurd in blauw. Afbeelding B en D zijn visualisaties van de SER-Ridge kwantificering.
Discussion
Met de dalende kosten van de HT / HC-cellulaire screening systemen, in combinatie met de beschikbaarheid van krachtige genoom-brede instrumenten om genfunctie te wijzigen, zijn HT / HC schermen steeds gemeengoed in de academische wereld. De aanpak is al met succes toegepast op diverse gebieden van onderzoek, zoals de identificatie van targets voor geneesmiddelen bij kanker 9, 31-33 en 34-36 embryonale ontwikkeling en heeft zelfs mogelijkheden voor toepassing in het ontcijferen van de wegen die betrokken zijn bij neuropsychiatrische stoornissen 37,38. Maar de uitvoering van een dergelijk systeem vergt een aanzienlijke investering van tijd en inspanning met procesoptimalisatie vaak het nemen van een minimum van 6 maanden. Alle stappen, zoals behandeling met trypsine keer, pipetteren snelheden en seeding dichtheden moeten worden aangepast, om ervoor te zorgen dat cellen gezond zijn en consistent te groeien. Preventie van bacteriële besmetting is een van de moeilijkste uitdagingen voor geautomatiseerde cel-cultuur met wekelijkse schoonmaak protocollen in combination met een constante spoelen van alle vloeibare peilingslijnen met 70% ethanol noodzakelijk voor de verontreiniging gratis culturen. Het zal ook nodig zijn om de robotica te verbeteren, zodat aanvullende instrumenten, zoals de confocale microscopen voor een hogere resolutie en -80 ° C diepvriezers voor samengestelde opslag kan worden geïntegreerd.
Er zijn ook beperkingen die moeten worden gericht aan de gevoelige, de snelheid en het nut van deze methode gebruiken om gen-netwerken bestuderen en identificeren van genen betrokken bij pathogene moleculaire pathways te verbeteren.
Voor het uitvoeren van een HT / HC-scherm en ervoor zorgen dat betrouwbare gegevens worden verzameld, een aantal aspecten rekening te worden geoptimaliseerd. De eerste, de betrouwbaarheid van de meting staat voorop en is afhankelijk van de robuustheid en de gevoeligheid van de test. Bijvoorbeeld, de testen hierboven beschreven zijn geschikt voor kleinere schermen, maar zijn moeilijk te implementeren op een genoom-wijde schaal, door het aantal verwerking stappen die nodig zijn voordat de foto acquisitie.Zo zou een te stabiele cellijnen de uiting van de reporter-gen, die het mogelijk maken voor directe beeldvorming en leiden tot verminderde variatie te wijten aan het verminderde aantal processtappen te construeren. Op dit moment is het ontwerpen van een test die nauwkeurig en betrouwbaar toont kwantificeert een fenotype van belang is een belangrijk knelpunt in de HC screening proces.
Veel schermen zijn uitgevoerd in cellen van zoogdieren met behulp van verschillende RNAi bibliotheken, die allemaal lijden aan off-target effecten beperkt gene silencing efficiëntie en onvolledige genoom dekking. Zo bibliotheken moeten worden gemaakt die meer specifieke, krachtige en hebben een betere dekking. Er wordt gewerkt aan maken dergelijke Het is te hopen deze inspanningen zal de reproduceerbaarheid van de HT / HC scre te verbeteren en hits.
Een beperking van de vele grote schaal cell-based schermen zijn dat zij worden uitgevoerd in neuroblastoom cellen omdat ze kunnen genetisch worden gemanipuleerd en gekweekt om grote aantallen met relatief gemak. Echter, de relevantie van 'hits' die in ex vivo celcultuur modellen in vivo functie is twijfelachtig, vooral omdat de hersenen bestaan uit zeer gespecialiseerde celtypen die een dicht en ingewikkeld netwerk van synaptische verbindingen om te functioneren als een sterk geïntegreerde eenheid vormen. Als gevolg daarvan, is het gebruikelijk dat raakt geïdentificeerd met behulp van de screening methode zoals hierboven beschreven, worden gevalideerd in het tweede scherm het gebruik van extra technieken en in meer fysiologisch relevante modellen 39. Ter verbetering van de vertaling van hits die tijdens HCS, meer representatieve en geavanceerde modellen, zoals primaire cellen en gedifferentieerde stamcellen of co-cultuur systemen moeten worden ontwikkeld en aangepast voor HT / HC benaderingen.
ntent "> Met een combinatie van geautomatiseerde cel kweken en HC beeldvorming kan men snel nieuwe inzichten te krijgen in hoe neuronen functioneren en bepalen welke wegen belangrijk zijn voor ziekte-ontwikkeling. combinatie van HCS / HTS gegevens met informatie uit andere 'omics' nadert, zal het dan mogelijk zijn om een systeembiologische overzicht van hersenziekten te bouwen, waardoor het gemakkelijker therapeutische ontwikkeling.Disclosures
We hebben niets te onthullen
Acknowledgments
Wij danken de Hamilton programmeurs en specialisten voor de voortzetting van steun en Eva Blaas voor technische bijstand. Dit werk werd ondersteund door twee NWO-Investeringssubsidies (911-07-031 en 40-00506-98-10011), Het Prinses Beatrix Fonds Wetenschapsprijs 2009 en de Neuroscience Campus Amsterdam, SJ wordt ondersteund door Ti-Pharma: T5-207.
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 9, 580-588 (2009).
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M. High-content analysis in neuroscience. Nat. Rev. Neurosci. 9, 779-788 (2008).
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C. High-throughput in vivo vertebrate screening. Nat. Methods. 7, 634-636 (2010).
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
