

Summary
Biz yüksek içerik görüntüleme ile otomatik hücre kültür birleştirerek yüksek verimli bir şekilde, çoklu hücresel süreçleri ve yapıları görselleştirmek ve ölçmek için bir metodoloji açıklanmaktadır. Bu yöntemler daha fonksiyonel ek açıklama genomların yardım yanı sıra hastalık gen ağları ve potansiyel ilaç hedefleri belirleyebilir.
Abstract
Genomların işlevsel açıklama, inşaat, moleküler ağlar ve yeni ilaç hedef tanımlama 1-4 ehemmiyetli bir mesele olarak ele alınması gereken önemli sorunlar olarak karşımıza çıkıyor. Çoklu tamamlayıcı 'omics yaklaşımları, genetik risk faktörleri ve birçok nörodejeneratif hastalıkların altında yatan patojenik mekanizmaları olarak ipuçları vermiş, ancak en bulgular hala işlevsel doğrulama 5 gerektirir . Örneğin Parkinson Hastalığı (PH) hastalığı için risk faktörü olarak tanımlanan pek çok yeni lokusların, ancak altta yatan etken varyant (ler) veya patojenik mekanizma için yeni bir genom ilişki çalışmasında, 6, 7 bilinmemektedir. Ilişkili her bölgede birçok gen içerebileceği gibi, hücre biyolojisi, geleneksel teknikleri kullanarak, hastalığı ile ilişkili fenotipleri genlerin her biri işlevsel bir değerlendirme çok uzun sürecektir.
Moleküler ağlar anlamak için de ihtiyaç vardır bu bağlantıyıneden fenotipleri genetik mutasyonlar. Bu hastalık fenotipleri bozulmuştur birden fazla etkileşimlerin sonucu olduğunu bekleniyor. Geleneksel moleküler yöntemler kullanılarak bu ağların İmar zaman alıcı olacaktır. Ayrıca, ağ bileşenleri tek tek bağımsız çalışmalar, indirgemecilik yaklaşım, tahminler, muhtemelen ağ karmaşıklığını 8 hafife . Bu, kısmen, küçümsenmesi, istenmeyen ya da toksik yan etkileri nedeniyle ilaç onayı düşük başarı oranı ile açıklanabilir. İY / HC hücresel tarama yaklaşımları kullanarak, bu yollar içinde anahtar düğümleri tanımlamak hastalığı ile ilgili yollarının bir ağ bakış açısı kazanmak, terapötik müdahale için daha uygun olan hedeflerin tanımlanmasına yol açabilir.
Yüksek verim tarama (HTS), 9-12, bu sorunları gidermek için ideal bir yöntem. ancak geleneksel yöntemlerle simpli kullanılan tek boyutlu tüm iyi hücre deneyleri,STIC karmaşık biyolojik süreçler için readouts. Onlar gibi, morfoloji özellikleri, 13, 14 aksonal ulaşım açıkları veya değişiklikler gibi nörodejeneratif hastalıklarda görülen birçok fenotipleri aynı anda ölçmek için koyamadık . Bu yaklaşım, bir hücre alt grubunda meydana gelen hücresel süreçleri ya da patojenik olayları dinamik doğasını araştırmak için kullanılan olamazdı. Bir çok-boyutlu fenotipleri geçmek için bu tür özellikleri ölçmek için yüksek içerik tarama (HCS) 4, 15-17 olarak nitelendirdi. HCS HTS göre çeşitli tedirginlikler hücresel yanıt daha ayrıntılı bir gösterimini sağlar, aynı anda çeşitli süreçlerin hücre tabanlı nicelikleme.
HCS HTS 18, 19 üzerinden pek çok avantajı vardır, ancak nöronal modelleri, yüksek verimli (HT) yüksek içerik (HC) ekran yapmak, yüksek maliyet, çevresel değişim ve insan hatası nedeniyle sorunludur. 'Phenomics ölçekte hücresel tepkileri tespit etmek içinHC görüntüleme birini kullanarak, hassasiyet ve tekrarlanabilirlik artırırken, değişim ve hatayı azaltmak için vardır.
Bu yazıda shRNA ekranlarında nöronal hücre modellerinde otomatik Hücre kültü 20 ve HC görüntüleme kullanarak doğru ve güvenilir yapmak için bir yöntem açıklanmaktadır. Biz biz mutasyona uğramış otozomal resesif parkinsonizm 21 neden belirli bir protein, DJ1, modülatörler tanımlamak için bu metodoloji nasıl kullanıldığını açıklar.
HC görüntüleme HT yöntemleri ile çok yönlülüğünü birleştiren doğru bir bolluk fenotipleri ölçmek için mümkündür. Bu sonradan genomu anlayışımıza, hastalığın patogenezinde yer yolların yanı sıra potansiyel terapötik hedefler belirlemek ilerlemek için yararlanılabilir.
Protocol
1. Otomatik yapışık hücre kültürü süreci (Şekil 1) 20
- Hücre kültürü plakaların girişi için otomatik hücre kültür sistemi hazırlayın. Yük sarf malzemeleri (örneğin pipet uçları, hücre kültürü plakaları, tahlil tabak) grafik kullanıcı arayüzü (GUI) kullanarak sistemi. Yeterli hücre kültür ortamı var olun, fosfat tamponlu salin (PBS) ve tripsin robotik sistem.
- SH-SY5Y nöroblastom hücre hattı plaka başına 2 x 10 6 hücre, el ile tohum iki omnitray plakalar. Opti-MEM hücrelerin% 10 fetal sığır serumu (FBS) ile koruyun. GUI kullanarak hücre kültürü robot tabak koyun. Hücreler 37 ° C ve% 5 CO 2 'de inkübe olacaktır .
- Kültür protokolü hangi hücrenin 20 başlatılan gerektiğini seçin. Bir yapışık, hücre kültürü süreci 22, kültür ve fare besleyici hücreler 23 veya süspansiyon cel kültür embriyonik kök (ES) hücrelerinin genişlemesi seçebilirls 22.
- Yapışık hücre kültürü protokolü (Şekil 1) seçin ve yapışık hücre hattı spesifik parametre dosyaları izdiham eşik değeri% 70 olarak belirlenmiştir (hücreleri içeren omnitray plaka alan) olacak şekilde ayarlanır. Sağlamak. Iki dakika toplam trypsinization zaman ayarlayın.
- Plaka başına 2 x 10 6 hücre tohumlama hücre sayısı, yeni omnitray plakaları hazırlamak için robot bilgilendirin.
- Hücre kültür sistemi otomatik yapışık hücre kültürü protokolü (Şekil 1) kullanarak girdi omnitray plakalar. Bu protokol, aşağıdaki adımları içerir: plakaları inkübe ve önceden tanımlanmış izdiham eşik ulaşana kadar görüntülü. Hücreler 5 okuma içinde izdiham eşik ulaşmak yoksa, tabak sistemden kaldırılır. Kullanıcı tanımlı izdiham eşik ulaştıktan sonra, hücre, yıkanmış tripsinize ve sayılır. Hücrelerin önceden tanımlanmış bir sayıda yeni omnitray plakaları eklendi ve hücrelerin yeterli sayıda, belirli bir uyuşma varsatahlil plakaları er güverte taşınır ve belirli bir hücre sayısı her kuyuya vazgeçtik. Testi plakaları doğrudan, daha fazla işlem için sistem entegre bir mikroskop veya çıkış kullanarak görüntülü olabilir.
- Ortalama 5.000 hücrelerinin toplam 4 omnitray plaka başına tahlil plaka hazırlamak için sistem bilgilendirin.
2. tahlil tabak içine shRNA virüs üretim ve kaplama (gerekli Zaman: 6 gün)
- Ampisilin 100 mg / ml (Sigma-Aldrich) içeren Luria Bertani orta medya 2ml gecede shRNA vektörler (Açık Biyosistem, TRC1) içeren bakteriyel gliserol stokları büyütün.
- Üretici protokolü (Promega Sihirbazı MagneSil TFX) plazmid ayıklayın.
- RNAi Konsorsiyumu High-Throughput lentiviral Üretim (96 plaka) 24 protokolü kullanarak virüs üretin. Transdüksiyon için kullanılan virüs parçacıkları çoğaltma eksiktir ve bölünmüş çünkü lentivirüs ile Çalışma nispeten güvenligen ambalaj stratejileri, kendi üretimi için kullanılır. Ancak, lentivirüs ile çalışırken, ek biyogüvenlik prosedürleri kendini ve diğerleri 25 riskini en aza indirmek için gerekli. Tüm deneyler MLII veya BSL2 güvenlik düzeyi laboratuvarda yapılmalıdır. Lentivirüs parçacıkları ile temas halinde olan tüm plastik (pipetler, plastik tabaklar, medya), tasfiye etmeden önce, 24 saat çamaşır suyu ile inkübe edilmelidir.
- Plazmid pLKO.1 GFP (Sigma-Aldrich) kullanarak GFP pozitif hücrelerin yüzdesi belirleyerek lentivirüs enfeksiyon çokluğu (İçişleri Bakanlığı) hesaplayın.
- 3 bir İçişleri Bakanlığı ile tahlil tabak içine Plaka lentivirüs.
3. Lentiviral iletimi ve SH-SY5Y hücreleri (gerekli Zaman: 6 gün) nöronal farklılaşma
- Hücreleri (1.7 adım bakınız) tahlil plakaları eklenir. Otomatik hücre kültürü sistemi shRNA lentivirüs içeren tahlil tabak yerleştirin.
- 24 saat sonra, medyatahlil plakalar% 0.5 FBS ve farklılaşma sürecine başlamak için 0.1 mcM retinoik asit içeren Opti-MEM değişmiş olacaktır. SH-SY5Y hücrelerin farklılaşması nöritik yapıların görüntülenmesini sağlar ve hücre bölünmesi eşitler.
- 5 gün boyunca farklılaşma medya test plakaları inkübasyon devam edin. Bu hedef gen ekspresyonu maksimum demonte sağlar.
- 5 gün, mitokondri DJ1 translokasyonu teşvik etmek için 24 saat boyunca test plakaların yarısı 50 mcM H 2 O 2 ekleyin.
- 6 gün, yanı başına 200 nM final konsantrasyonda hücrelere Mitotracker CmxROS (Invitrogen) ekleyin ve 37 ° inkübe ° C'de 30 dakika.
- Bir sistem doğrudan HC görüntüleyici veya plakaları daha fazla işlem için sistem ihraç edilebilir kullanarak görüntü plakaları talimat verebilirsiniz.
4. Tahlil plakaları otomatik olarak immün (gerekli Zaman: 2 gün)
Görüntü kalitesi paramounduyarlı ve güvenilir bir HCS yürütülmesi için t. Yanlış pipetleme nedeniyle hücresel tek tabaka hasar, kötü görüntü kalitesine ve irreproducible sonuçları yol açabilir. Hücre tabakası hasarı en aza indirmek için, immün bir robot istasyonu kullanılarak yapıldı. Bu prosedür, daha önce 26 olan birine benzer ama verimi arttırmak ve sarf malzeme kullanımını azaltmak için özelleştirilmiş oldu.
- 100 4% paraformaldehid 37 ul önceden ısıtılmış ° C hücreleri saptamak Oda sıcaklığında 20 dakika inkübe edin.
- 5 dakika, 3 kez PBS ile 200 ul hücreleri yıkayın.
- 10 dakika boyunca% 0,1 Triton (PBST) içeren PBS 200 ul tahlil plakaları inkübe edin.
- 5 dakika, 3 kez PBS ile 200 ul hücreleri yıkayın.
- Tahlil plakaları oda sıcaklığında 1 saat için blok tampon 200 ul (% 5 FBS ile PBST) ile inkübe edin.
- 5 dakika, 3 kez PBS ile 200 ul hücreleri yıkayın.
- Foll ile inkübe4 geceleme nedeniyle primer antikor ° C
- Keçi DJ1 N20 (Santa Cruz, 5 mg / ml)
- Tavşan β-III tubulin (Sigma-Aldrich, 1 mcg / ml)
- Ertesi gün, 5 dakika, 3 kez PBS ile 200 ul hücreleri yıkayın.
- Tahlil plakaları oda sıcaklığında aşağıdaki sekonder antikor 1 saat inkübe
- AlexaFluor 488 eşek anti-keçi (Invitrogen, 2 mcg / ml)
- AlexaFluor 647 keçi anti-tavşan (Invitrogen, 2 mcg / ml
- 5 dakika, 3 kez PBS ile 200 ul hücreleri yıkayın.
- 10 dakika boyunca; Hoechst (1 mcg / ml Invitrogen) ile inkübe hücreleri.
- 200 ul PBS ile 3 kez 5 dakika, hücreleri yıkayın.
- Plakları, 4 ° C 'ye kadar görüntülenebilir.
5. (Zaman: 5 gün) içeriği yüksek görüntü elde etme ve görüntü analizi
- Toplam 20x objektif lens ile de ortalama 30 alan bir görüntü. FI ile DJ1 görselleştirinTC filtre seti, TRITC filtre seti, Cy5, filtre seti ve UV filtre seti (Şekil 3) kullanarak çekirdekleri β-III tubulin mitokondri.
- Mitokondri içinde Mitotracker sinyali ortalama yoğunluğunu belirlemek için komparmantal Analizi Bioapplication (Cellomics, ThermoFisher) kullanarak görüntüleri analiz edin. (Şekil 4B, F).
- DJ1 ve mitokondri arasındaki ortalama örtüşme katsayısı belirlemek için, Cellomics Colocalisation bioapplication (Cellomics, ThermoFisher) kullanarak görüntüleri analiz eder. Aşağıdaki gibi ilgi alanları (ROI) tanımlayın: ROI A - nükleus (Şekil 4A, E), YG B - mitokondri (Şekil 4 B, F). Sadece sitoplazma analizi sağlamak için ROI B ROI A dışlayın. Mitokondri hedef bölge olarak hedef bölge II (Şekil 4C, G) I ve DJ1 olarak tanımlayın.
- Β-III tubulin boyama (Şekil neurites ortalama uzunlukları iz Nöronal Profil bioapplication (Cellomics, Thermofisher) kullanarak görüntüleri analiz4D, H).
- Opera LX otomatik konfokal okuyucu (Perkin-Elmer) kullanarak görüntü plakalar. Suya daldırma ile 60x objektif lens kullanarak ortalama 30 alanların toplam görüntü. UV eksitasyon ile 561 nM lazer ve çekirdekleri ile mitokondri gözünüzde canlandırın.
- Spot-Kenar-Ridge (SER) doku özellikleri algoritması kullanarak görüntüleri analiz edin. SER-Ridge filtre sırtı benzeri desenler oluşturan piksel yoğunluğu iletir. Daha fazla parçalanmış mitokondri, yüksek SER-Ridge skoru (Şekil 8).
6. Veri normalleşme ve analiz
- R yazılım ortamı için BioConductor CellHTS2 paketi (R sürüm 2.11.1, BioConductor sürüm 2.6), görüntü analiz yazılımı, veri içe aktarın.
- Logaritma tabanı (2) plaka başına medyan temelli normalleşme 27, 28 öncesinde verileri dönüştürmek. Plaka başına varyansın ayarlaması uygulama yapmayınız.
- Bir fenotip değiştiriciler tanımlamak için, differe arasında iki yönlü ANOVAnt tedavi gruplarında yani virüslü tedavi edilmezse hücreleri Şifreli vs şifreli toksin muamele edilen hücreler hedef gen tedavi edilmezse hücreleri vs vs hedef gen tedavi hücreleri (Şekil 5-7).
7. Temsilcisi sonuçları
Mutasyonlar DJ1 içinde erken başlangıçlı resesif parkinsonizm 21 doğuran, ama DJ1 kaybı hastalığın fenotipi yol açmaktadır nasıl belli değildir. DJ1, eksik hücreleri oksidatif strese bağlı hücre ölümü daha duyarlı ve oksidatif stres, mitokondri 29 sitoplazma DJ1 translocates, 30 yanıt olduğu bilinmektedir. Bu fenotipleri izlemek için HC testleri inşa ederek, biz DJ1 ile ilişkili fenotipleri düzenleyen veya etkileyen genler tespit edebilir. Bu yaklaşım, deşifre DJ1 fonksiyonları ve hastalık patogenezinde içinde yer almış olabilir yollar yardımcı olabilir.
Epistatik DJ1 ile bir etkileşim (Şekil 5) Örnek: maruz kalan hücrelerde, demonte DJ1Görüntüyü hücre canlılığı büyük bir kaybı toksin sonuçları (BAR-B:-B) şifreli lentivirüs ile enfekte hücrelere kıyasla (BAR-A: image-A). Hedef gen demonte DJ1 demonte (image-C BAR-C) ile hücrelerinde gözlenen benzer bir etkiye sahiptir. DJ1 ve hedef genin sadece iki genin kaybı (image-Ge BAR-D) göre anlamlı olarak daha fazla hücre canlılığı kaybı sonuçları demonte. Bu epistatik DJ1 ve hedef gen A. arasında bir etkileşim de anlaşılacağı
Örnek hücreleri bir toksine maruz sitoplazma mitokondri DJ1 translocates DJ1 arasında daha yüksek bir örtüşme katsayısı ve mitokondri (BAR-A sayısal:: resim, bir karşı DJ1 translokasyon (Şekil 6) düzenleyen bir gen BAR-C: image C). B hedef gen susturulması olmuştur hücreler, daha az mitokondri DJ1 translocates hücreleri toksine maruz zaman. Bu hedef geni B mitokondri DJ1 taşınması yer almaktadır göstermektedir. (BAR-B: görüntü B ve BAR-D: image D)
Neurite uzunluğunda önemli bir artış vahşi tip SH-SY5Y hücreleri hedef gen C demonte: nöronal akıbet (Şekil 7) yer alan bir gen Örnek görüntü (BAR-B:-B), şifreli ifade lentivirüs ile enfekte hücreleri ile karşılaştırıldığında shRNA (BAR-A: Resim A). Bu etki, toksin (BAR-C ve D) ile inkübe hücreleri kaybolur.
Mitokondriyal morfolojisi (Şekil 8) yer alan bir gen Örnek: Enfeksiyon mitokondriyal SER-Ridge segmentasyonu değeri (Şekil 8, Resim C ve D) bir azalma gen-Ge sonuçlarını hedef shRNA vahşi tip SH-SY5Y hücreleri ile karşılaştırıldığında hücreleri şifreli lentivirüs (Şekil 8, Resim A ve B) ile enfekte.
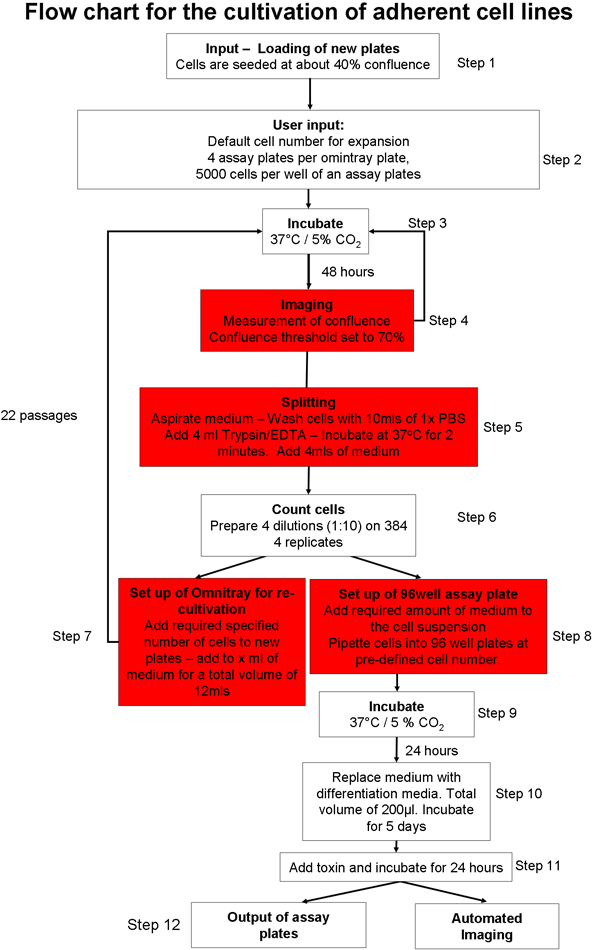
Şekil 1 otomatik hücre kültürü protokol Taslağı:

Şekil 2, tarama sürecinin şematik bakış, görüntü analizi ve tarama işlemi sırasında kullanılan istatistiksel yöntemler: shRNA lentivirüs içeren test plakaları içine belirsiz ve sonradan kaplama kadar i) Hücreler kültür. Hücreler, 5 gün boyunca farklı ve toksin sonra 24 saat boyunca plakaları eklenir. Testi plakaları sistemden çıkış ve immunohistokimyasal. Süreçlerinin her biri için alınan zamanın miktarı parantez içinde gösterilir. ii) Veri HC görüntüleyici (hücre canlılığı, protein translokasyon ve nöronal akıbet) ve otomatik bir konfokal görüntüleyici (mitokondriyal morfoloji) kullanılarak elde edilir. Veri R CellHTS2 paketi içinde ihraç edilmektedir, baz (2) değiştirdi ve normalize giriş. İki yönlü ANOVA farklı değişkenler arasındaki önemli etkileşimi tanımlamak için kullanılır.
Şekil 3 HC görüntüleme ile elde edilen hücreler kompozit görüntüler. A) Tedavi edilmeyen hücreleri, B) Hücreler H 2 O 2 ile tedavi. DJ1, yeşil, kırmızı ve mavi çekirdekleri mitokondri etiketlenmiştir. Neurite boyama vurgulu değildir.

Şekil 4 bir HCS elde edilen çok sayıda cep telefonu özellikleri Kantitasyonu . AD) Tedavi edilmemiş SH-SY5Y hücreleri. EH) H 2 O 2 tedavi SH-SY5Y hücreler. ROI A, E) Çekirdekler segmentasyonu ve tanımı; mitokondri, YG B, B, F) tanımlanması ve ölçülmesi, C, G) DJ1 belirlenmesi, Hedef kanal II; ortalama neurite uzunluğu D, G) Tanımlama ve hesaplama. Görüntünün kenarına yakın Hücreler analize dahil. Ankastre görüntüleri analizden önce görüntüleri.

Şekil 6 DJ1 translokasyon (resimlere yazı karşılık barlarda harf) düzenleyen bir gen belirlenmesi. Görüntüler D yoluyla mitokondri DJ1 translokasyon ölçümü için kullanılan mitokondri DJ1 (yeşil), (kırmızı) ve çekirdekler (mavi) etiketli SH-SY5Y hücrelerdir.

Şekil 7 nöronal akıbet yer alan bir gen belirlenmesi (resimlere yazı karşılık barlarda harf). A ile D arasındaki neurite uzunluk ölçümü için kullanılan β-III tubulin (yeşil) ve çekirdekler (mavi) etiketli SH-SY5Y hücreleri görüntüler.

Şekil 8 mitokondriyal morfolojisi düzenleyen bir gen belirlenmesi. Resim A ve C sırasıyla şifreli shRNA veya shRNA hedef geni D ile enfekte olan SH-SY5Y hücrelerinin kompozit görüntüler. Çekirdekleri mavi renkli iken Mitokondri kırmızı renklidir. Resim B ve D SER-Ridge kantifikasyon görselleştirme.
Discussion
Gen işlevini değiştirmek için güçlü bir genom araçlarının durumu ile kombine HT / HC hücresel tarama sistemleri, azalan maliyetler, HT / HC ekranlar akademide olağan hale gelmiştir. Bu yaklaşım zaten başarıyla araştırma, kanser 9, 31-33 ve embriyonik gelişim 34-36 ilaç hedeflerinin belirlenmesi gibi çeşitli alanlarda uygulanan ve hatta nöropsikiyatrik hastalıklarda 37,38 dahil yollar deşifre uygulama potansiyeline sahiptir. Ancak böyle bir sistemin uygulanması, genellikle en az 6 aylık bir süreç optimizasyonu ile zaman ve çaba önemli bir yatırım gerektirir. Trypsinization kez pipetleme hızları ve ekim yoğunlukları gibi tüm adımlar, hücrelerin sağlıklı olmasını sağlamak ve sürekli büyümeye ayarlanması gerekir. Bakteriyel kontaminasyonun önlenmesi, c haftalık temizlik protokolleri ile otomatik hücre kültürü karşılaştığı en zor durumlardan birikirlenme ücretsiz kültürler için gerekli olan% 70 etanol ile tüm sıvı taşıyan hatların sabit durulama ile ombination. Bu robotik geliştirmek için de gerekli olacak, böylece, yüksek çözünürlük ve -80 ° C derin dondurucular bileşik depolama için entegre edilebilir konfokal mikroskoplar gibi ek araçlar.
Gen ağları çalışma ve patojenik moleküler yolaklar ilgili genleri tanımlamak için bu yöntemi duyarlı, hızlı ve faydalı artırmak için ele alınması gereken sınırlamalar da vardır.
HT / HC ekran davranış ve güvenilir veri toplanmış olduğundan emin olmak için, çeşitli yönleriyle optimize edilmiş olması. İlk olarak, ölçüm güvenilirliği çok önemlidir ve sağlamlık ve duyarlılık testinin bağlı. Örneğin, yukarıda açıklanan deneyleri, küçük ekranlar için uygundur, ancak görüntü elde etme önce gerekli sayı işleme adımları nedeniyle geniş çaplı bir genom, uygulamak zor.Böylece, doğrudan görüntüleme için izin ve işlem adımları az sayıda nedeniyle azalmış varyasyon yol açacak muhabiri gen ifade istikrarlı hücre hatları inşa etmek olurdu. Şu anda, doğru bir tasvir ve güvenilir bir ilgi fenotipi HC tarama sürecinin büyük bir darboğaz olduğunu rakamlarla bir test tasarımı.
Birçok ekranlarında-hedef dışı etkileri sınırlı gen susturulması verimlilik ve eksik genom kapsama muzdarip her biri farklı RNAi kütüphaneler, memeli hücreleri kullanılarak yapılmaktadır. Böylece kütüphaneler, daha spesifik, güçlü ve daha iyi bir kapsama sahip olduğu yapılması gerekir. Oluşturmak için çalışmalar devam etmektedir bu tür bu tür çabalar İY / HC scre tekrarlanabilirliğini artıracak umulmaktadır tr vurur.
Onlar genetik göreli kolaylıkla manipüle ve çok sayıda etmek için kültürü, çünkü birçok büyük ölçekli hücre tabanlı ekranlar bir sınırlama nöroblastom hücrelerinde yapılır. Ancak, in vivo fonksiyonu, ex-vivo hücre kültürü modelleri tanımlanan 'hit' alaka beyin sinaptik bağlantıların fonksiyonu için son derece entegre bir birim olarak yoğun ve karmaşık bir ağ oluşturan son derece uzmanlaşmış hücre tipleri oluşur, özellikle de sorgulanabilir. Sonuç olarak, yukarıda açıklanan tarama yaklaşımı kullanılarak belirlenen ek teknikler kullanılarak ikincil ekranları ve daha fizyolojik ilgili modelleri 39 valide Vurduğu yaygındır. HCS sırasında tanımlanan çeviri hit artırmak için, birincil hücre ve farklılaşmış kök hücreler ya da ko-kültür sistemleri gibi daha temsili ve sofistike modelleri, İY / HC yaklaşımlar için geliştirilen ve adapte olması gerekir.
otomatik hücre kültür ve HC görüntüleme tek bir kombinasyonu ile ntent "> nöron fonksiyonu nasıl içine hızla yeni anlayışlar kazanmak ve hangi yollar hastalık gelişiminde önemli 'diğer omics' yaklaşımlar elde edilen bilgiler ile HCS / HTS verileri birleştiren belirlemek, sonra beyin hastalıkları bir sistem biyolojisi genel bakış oluşturmak için, böylece tedavi gelişimi kolaylaştırmak mümkün.Disclosures
Biz ifşa etmek başka bir şey var
Acknowledgments
Hamilton programcılar ve uzman teknik yardım için sürekli destek ve Eva Blaas için teşekkür ederiz. T5-207: SJ Ti-Pharma tarafından desteklenen; Bu çalışma iki NWO Yatırım hibe (911-07-031 ve 40-00506-98-10011), Prinses Beatrix Viyana Wetenschapsprijs 2009 ve Nörobilim Kampüsü Amsterdam tarafından desteklenmiştir.
Materials
| Name | Company | Catalog Number | Comments |
| AI.CELLHOST | Hamilton Co | http://www.hamiltonrobotics.com/en-uk/applications/cellomics/ | |
| OPTI-MEM | Invitrogen | 31985-054 | |
| RETINOIC ACID | Sigma-Aldrich | R2625 | |
| OMNITRAY PLATES | Nalge Nunc international | 465219 | |
| 96 WELL CULTURE PLATES | Greiner Bio-One | 655086 | |
| DJ1 N20 ANTIBODY | Santa Cruz Biotechnology, Inc. | SC27004 | |
| BETA-III TUBULIN ANTIBODY | Sigma-Aldrich | T3952 | |
| MITOTRACKER CMXROS | Invitrogen | M-7512 | |
| H–CHST-33342 | Invitrogen | H1399 | |
| HYDROGEN PEROXIDE | Sigma-Aldrich | 216763-100ML | |
| TRYPSIN | Invitrogen | 25050014 | |
| DULBECCO’S PHOSPHATE BUFFERED SALINE | Invitrogen | 14190086 | |
| PROMEGA WIZARD MAGNESIL TFX | Promega Corp. | A2380 | |
| SHRNA CLONES | Open Biosystems | http://www.openbiosystems.com/RNAi/shRNALibraries/ TRCLibraryDetails/ | |
| CELLOMICS BIOAPPLICATIONS | Thermo Fisher Scientific, Inc. | http://www.thermo.com/hcs |
References
- Geschwind, D. H., Konopka, G. Neuroscience in the era of functional genomics and systems biology. Nature. 461, 908-915 (2009).
- Ge, H., Walhout, A. J., Vidal, M. Integrating 'omic' information: a bridge between genomics and systems biology. Trends. Genet. 19, 551-560 (2003).
- Zhu, H., Snyder, M. 'Omic' approaches for unraveling signaling networks. Curr. Opin. Cell. Biol. 14, 173-179 (2002).
- Jain, S., Heutink, P. From single genes to gene networks: high-throughput-high-content screening for neurological disease. Neuron. 68, 207-217 (2010).
- Manolio, T. A. Finding the missing heritability of complex diseases. Nature. 461, 747-753 (2009).
- Nalls, M. A. Imputation of sequence variants for identification of genetic risks for Parkinson's disease: a meta-analysis of genome-wide association studies. Lancet. 377, 641-649 (2011).
- Simon-Sanchez, J. Genome-wide association study confirms extant PD risk loci among the Dutch. Eur. J. Hum. Genet. 19 (6), 655-661 (2011).
- Van Regenmortel, M. H. Reductionism and complexity in molecular biology. Scientists now have the tools to unravel biological and overcome the limitations of reductionism. EMBO. Rep. 5, 1016-1020 (2004).
- Aherne, G. W., McDonald, E., Workman, P. Finding the needle in the haystack: why high-throughput screening is good for your health. Breast. Cancer. Res. 4, 148-154 (2002).
- An, W. F., Tolliday, N. J. Introduction: cell-based assays for high-throughput screening. Methods. Mol. Biol. 486, 1-12 (2009).
- Mayr, L. M., Bojanic, D. Novel trends in high-throughput screening. Curr. Opin. Pharmacol. 9, 580-588 (2009).
- Hertzberg, R. P., Pope, A. J. High-throughput screening: new technology for the 21st century. Curr. Opin. Chem. Biol. 4, 445-451 (2000).
- Conrad, C., Gerlich, D. W. Automated microscopy for high-content RNAi screening. J. Cell. Biol. 188, 453-461 (2010).
- Thomas, N. High-content screening: a decade of evolution. J. Biomol. Screen. 15, 1-9 (2010).
- Arrasate, M., Finkbeiner, S. Automated microscope system for determining factors that predict neuronal fate. Proc. Natl. Acad. Sci. U. S. A. 102, 3840-3845 (2005).
- Dragunow, M. High-content analysis in neuroscience. Nat. Rev. Neurosci. 9, 779-788 (2008).
- Varma, H., Lo, D. C., Stockwell, B. R. High throughput screening for neurodegeneration and complex disease phenotypes. Comb. Chem. High. Throughput. Screen. 11, 238-248 (2008).
- Durr, O. Robust hit identification by quality assurance and multivariate data analysis of a high-content, cell-based assay. J. Biomol. Screen. 12, 1042-1049 (2007).
- Miller, J. Quantitative relationships between huntingtin levels, polyglutamine length, inclusion body formation, and neuronal death provide novel insight into huntington's disease molecular pathogenesis. J. Neurosci. 30, 10541-10550 (2010).
- Jain, S., Sondervan, D., Rizzu, P., Bochdanovits, Z., Caminada, D., Heutink, P. The Complete Automation of Cell Culture. Journal of Biomolecular Screening. 16 (8), (2011).
- Bonifati, V. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science. 299, 256-259 (2003).
- Ricardo, R., Phelan, K. Trypsinizing and Subculturing Mammalian Cells. J. Vis. Exp. (16), e755-e755 (2008).
- Kent, L. Culture and Maintenance of Human Embryonic Stem Cells. J. Vis. Exp. (34), e1427-e1427 (2009).
- Moffat, J. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 124, 1283-1298 (2006).
- Volksgezondheid, M. V. Integrale versie van de Regeling genetisch gemodificeerde organismen en het Besluit genetische gemodificeerde. Organismen. , (2004).
- Anderl, J. L., Redpath, S., Ball, A. J. A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J. Vis. Exp. (27), e1173-e1173 (2009).
- Wiles, A. M., Ravi, D., Bhavani, S., Bishop, A. J. An analysis of normalization methods for Drosophila RNAi genomic screens and development of a robust validation scheme. J. Biomol. Screen. 13, 777-784 (2008).
- Canet-Aviles, R. M. The Parkinson's disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. U. S. A. 101, 9103-9108 (2004).
- Blackinton, J. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 284, 6476-6485 (2009).
- Gupta, P. B. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138, 645-659 (2009).
- Luo, J. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 137, 835-848 (2009).
- Mouchet, E. H., Simpson, P. B. High-content assays in oncology drug discovery: opportunities and challenges. IDrugs. 11, 422-427 (2008).
- Vogt, A. Automated image-based phenotypic analysis in zebrafish embryos. Dev. Dyn. 238, 656-663 (2009).
- Pardo-Martin, C. High-throughput in vivo vertebrate screening. Nat. Methods. 7, 634-636 (2010).
- Vogt, A., Codore, H., Day, B. W., Hukriede, N. A., Tsang, M. Development of automated imaging and analysis for zebrafish chemical screens. J. Vis. Exp. (40), e1900-e1900 (2010).
- Ross, P. J., Ellis, J. Modeling complex neuropsychiatric disease with induced pluripotent stem cells. F1000. Biol. Rep. 2, 84-84 (2010).
- Ebert, A. D., Svendsen, C. N. Human stem cells and drug screening: opportunities and challenges. Nat. Rev. Drug. Discov. 9, 367-372 (2010).
- An, W. F., Tolliday, N. Cell-based assays for high-throughput screening. Mol. Biotechnol. 45, 180-186 (2010).
