

Summary
細胞周期ステージの進行と動態の追跡微妙な変化は、BrdUとヨウ化プロピジウムを経由して全ゲノムDNAを染色した核酸の代謝標識の組み合わせの使用によって達成することができます。このメソッドは、それによって順番に細胞周期の進行に影響を及ぼす非特異的DNA損傷の導入を防止し、サイクリング細胞の化学物質の同期の必要がなくなります。
Abstract
細胞周期のさまざまな段階を経て開始とその後の進行の正確な制御は、細胞の増殖に極めて重要である。細胞周期の分裂は、成長と重要な細胞周期成分の再現性と規制緩和が発癌1,2の沈殿イベントに関与しているの不可欠な部分です。抗がん治療における分子のエージェントが頻繁に細胞周期の分裂3の規制と調整を担当生物学的経路を対象としています。細胞周期動態は細胞の種類に応じて変化する傾向があるが、細胞周期の4つの段階の中の細胞の分布は、マイトジェン、成長因子の発現の一貫したパターンに起因する特定の細胞株内ではなく、一貫性があります。遺伝毒性のイベントや他の細胞ストレスがinstiを可能にするために逮捕されたり、特定の細胞周期フェーズの一時的なポーズ、その結果、細胞周期の進行を一時的にブロックになることが適切な応答機構のgation。
実験的にそれらの細胞周期の進行の段階への参照を使用して、細胞集団の振る舞いを観察する能力は、細胞生物学における重要な前進である。そのような細胞分裂などの一般的な手順は、細胞周期4-6の特定の段階で細胞を単離するために使用され、分画遠心法またはフローサイトメトリーベースのソートを振り払う。これらを分画し、細胞周期相濃縮集団はその後、実験的な治療に供される。区切られた画分の収量、純度、生存率は、しばしば、これらの物理的な分離メソッドを使用して、妥協することができます。同様に、細胞集団の分離分画した細胞が選択された細胞周期の段階から進行することができる実験的な治療の開始の間の時間経過は、実装を成功させると、これらの実験の解釈に重要な課題を提起することができます。
STに他のアプローチUDY細胞周期の段階は、細胞を同期させる化学物質の使用が含まれています。各細胞周期ステージのための鍵となる代謝プロセスの化学的阻害剤による細胞の処理は、次の段階に細胞周期の進行を阻止するのに有用である。デオキシヌクレオチド、DNAのビルディングブロックの供給を制限することにより、G1 / S時点で、例えば、リボヌクレオチド還元酵素阻害剤ヒドロキシウレアを停止した細胞。他の顕著な化学物質がアフィジコリンによる治療を含む、有糸分裂紡錘体の形成を妨げるどちらもG1停止のためのポリメラーゼα阻害剤、コルヒチンとノコダゾールによる治療は、M期で細胞を停止し、最終的に、DNA鎖ターミネーター5 fluorodeoxyridineによる治療が開始するS期停止7-9これらの化学物質による治療は、特定の段階で細胞の全人口を同期させる有効な手段である。化学物質の除去と、細胞が一斉に細胞周期に復帰。テストエージェントは、次のリリースの治療細胞周期阻止化学物質から誘発される薬物反応が均一で、細胞周期ステージ特定の集団からのものであることを保証します。しかし、化学シンクロナイザの多くのため、挑戦している様々な応答経路(テストエージェント対シンクロナイザまで)の参加を離れてからかう、遺伝毒性の化合物が知られている。
ここでは、彼らの娘細胞の分裂と分離に至るまで、積極的にDNA複製の段階からその進行を介して細胞を自転車の集団を以下の代謝標識法を説明します。フローサイトメトリーの定量化と相まって、このプロトコルのいずれかを機械的または化学的に誘導され、携帯は一般的に他の10の細胞周期の同期化の方法論に関連付けられているストレスの存在下で細胞周期の進行運動を測定するために有効になります。以下のセクションでは、方法論だけでなく、生物医学研究への応用のいくつかを説明します。
Protocol
1。細胞調製

- 約60%コンフルエントの密度を達成するために細胞をプレート。細胞は、収集時に対数増殖期にある必要があります。 MCF7細胞では、これは適切なメディアで5×10 5細胞/ 10 cmのプレートに播種することによって達成される。 HT29とLS180細胞を6×10 5細胞/ 6 cmのプレートで播種されています。我々は、DMEM培地、10%FBSおよび1xペニシリン/ストレプトマイシンを補充した使用されます。で正しい陽性および陰性コントロールのための種子プレートに必ずテストサンプルに加えて。彼らは次のとおりです。
| 私は | 陽性対照 | のみBrdUの |
| 2 | 陽性対照 | PIのみ |
| 3 | 負の制御 | たBrdU陰性、PI陰性 |
(このような、このプロトコルで説明されている一つとして、複数の時点で予備実験は、将来のコレクションを実施するために、その間の時間枠を絞り込むために使用することができることに留意する必要があります。)
- メッキ細胞を37℃でインキュベートし、5%24〜48時間のためにCO 2、それによって細胞が回復し、添付することができます。
- ブロモデオキシウリジン(BrdU)で標識細胞をパルスするには、DMEMは、wiを置き換えられます。10μMのBrdUを含む番目の新鮮な培地。細胞は、°C、5%CO 2は、DNAへのBrdUの取り込みを可能にするために37℃で1時間インキュベートされています。ネガティブコントロールとして動作するように未処理の一皿を残すようにしてください。
- パルス標識培地を除去し、細胞を1X PBSで簡単に洗浄する。
- 新鮮な増殖培地に追加され(マイナスBrdU)の、細胞を収穫するための適切なタイムポイントに達するまで37℃、5%CO 2でインキュベートし続けることが許可されています。
2。収穫と固定
- ステップ1.3から未処理の細胞は、ゼロタイムポイントと見なされます。このサンプルは、BrdUの治療直後に収集された1時間タイムポイントと一緒に収穫することができます。
- 細胞を回収するには、メディアが削除され、プレートは、1×PBSで1回リンスしています。
- 細胞は5分で燮は1500 rpmで遠心分離することによって、トリプシン処理し、培養培地中で収集され、その後、ペレット化されているernatantは破棄されます。
- 5ミリリットルの氷冷1X PBSで1から10×10 6細胞を洗浄します。 5分の上清を1500rpmで遠心分離は破棄されます。
- 細胞ペレットを氷冷PBS / 1%FBSを100μlに再懸濁されています。細胞の凝集を防止するのにPBSエイズの1%FBSを添加する。
- -20°C、細胞を固定するために70%エタノール5mlにドロップでこれらの細胞のドロップを追加します。
- 4℃で一晩で30分間またはストア氷上でインキュベートします。これは、様々の時点からのサンプルの収集を停止するときの理想的なステップです。サンプルは、それらのプロトコルの残りの部分を同時に処理できるようになり、数日間エタノールにしておくことができます。
3。 BrdUとPI染色
- 1500 rpmで5分間遠心分離により細胞をペレット化する。
- 固定液を除去するが、ボルテックスによりペレットを緩めに〜50μlを残す。
- DNAを変性させ、徐々に2NのHCl /トリトンX-100の1ミリリットルを追加するボルテックスしながら滴下した。室温で30分間サンプルをインキュベートします。
- 1500rpmで5分間サンプルを遠心分離により細胞をペレット化する。吸引し、上清を捨てる。
- 変性ステップを中和するために、0.1Mナトリウム四ホウ酸、pHが8.5の1 mlに細胞ペレットを再懸濁します。
- 1500rpmで5分間遠心します。吸引し、上清を捨てる。
- BrdU染色ミックス75μL(50μlの0.5%のトウィーン20/1%BSA / PBS +20μlのFITC標識抗BrdU + 5μlの10 mg / mlのRNaseの)に細胞ペレットを再懸濁します。
- 光から保護された45分間室温でインキュベートします。
- 細胞をペレット化する、1500 rpmで5分間サンプルを遠心します。吸引し、上清を捨てる。
- 5μg/ mlのヨウ化プロピジウムを含む1mlのPBSで細胞ペレットを再懸濁します。
4。フローサイトメトリー
- 細胞は、488 nmのレーザーとを備えたフローサイトメトリーを分析する適切なフィルタが必要です。そのようなCellQuestなどの分析ソフトウェアは、以下に概説するプロットを作成する必要があります。
- フローサイトメーターを介してサンプルを実行すると、細胞の適切な粒度分布( 図1A)を確保するために前方散乱(SSC-H)対側方散乱(FSC-H)プロットを作成します。これらのパラメータの両方が線形スケールでプロットされています。
- 同時にFL3-H(染色PI)対FL2-W( 図1B)のプロットを表示します。このプロットは、細胞周期の正常な分布パターン内にある細胞の割合を分離するためにゲート(R3)を作成するために使用されます。一般に、この分布パターンは、それぞれ、細胞周期のG1およびG2の段階を表すPI染色の2Nと4N DNA含量からの細胞のつのクラスタによって特徴づけられる。これらの2つのクラスタ間に位置するセルの文字列は、細胞周期のS期に発生する進行中のDNA複製の代表です。
- プロットdisplayiを作成します。NG、y軸とx軸( 図1C)のPI(線形プロット)のFL1-Hのステップ4.3(FITC、ログプロット)からゲート細胞(R3)。このプロットは分析用のデータを収集するだけでなく、1.1で概説したさまざまなコントロールを使用して残りのパラメータを設定するために使用されます。
- メーターの上にPIのみ染色したサンプルを配置します。 x軸上の〜200℃でG1を配置するゲインを調整します。これは、染色、PIのヒストグラムプロットで視覚化することは容易になります。
- 第2の制御フローサイトメーターでのBrdUのみサンプルを実行するが含まれます。細胞の2集団(BrdU陽性対陰性のBrdU)がプロットに表示されるようにゲインを調整します。理想的には、BrdUを陰性細胞はちょうど10 -1の下に表示されるように配置されています。
- 最後のコントロールでは、BrdUを/ PI陰性サンプルです。細胞は上部または右側の象限のいずれにも表示されないことを確認するには、この負の制御を実行します。
- さまざまなプロットのパラメータが設定されていたら、フローサイトメーターは、校正およびBrdUおよびPI染色した細胞を処理するための準備ができています。正しいPI画分(4.3ステップを参照してください)のゲートである10,000個の細胞の最小値は、コレクションごとに読み取る必要があります。
- 収集したデータを分析するには、そのようなFlowJoまたはFacsDivaなどのソフトウェアが利用されています。上記の各プロットは、これらのプログラムで再現し、定量的かつ統計的な分析が行うことができます。
- 分析のエンドポイントはいくつかの連続した手順を実行します。 PI対FL2-Wプロットから正PIとしてゲート細胞とFITC対PIのプロットを作成すると、1つは、サイクリング人口を区別することができます。第2のゲートは、BrdU陽性集団を単離することにより、このプロットから作成されます。 x軸上のPIヒストグラムプロット上で、この独特の人口を表現すると、1つは、細胞周期の進行の経時変化を追跡することができます。これは時間の関数としてG1またはG2コンテンツのBrdU陽性細胞数をプロットすることにより、さらに視覚化することができます。
5。代表的な結果
通常、ヨウ化プロピジウムで染色したサイクリング細胞はそれぞれ、2Nと4N DNA含量を含有する細胞( 図2)に対応するG1とG2で明瞭なピークを持っています。 BrdUでパルスラベリングは積極的にDNA(すなわちS期)を合成している細胞の亜集団の選択的標識が可能になります。まもなくBrdUの試薬 を除去した後、すべての標識細胞は、S相( 図3)にあります。短パルスいずれかにラベル付けを制限することによって、彼らは、細胞周期の後続のフェーズを通過する数々の時間点を通して細胞のこの今の異なるサブ母集団に従うことができます。これは、G1とG2のピークの明確な欠如として、図3の1時間の時点で視覚化することができます。
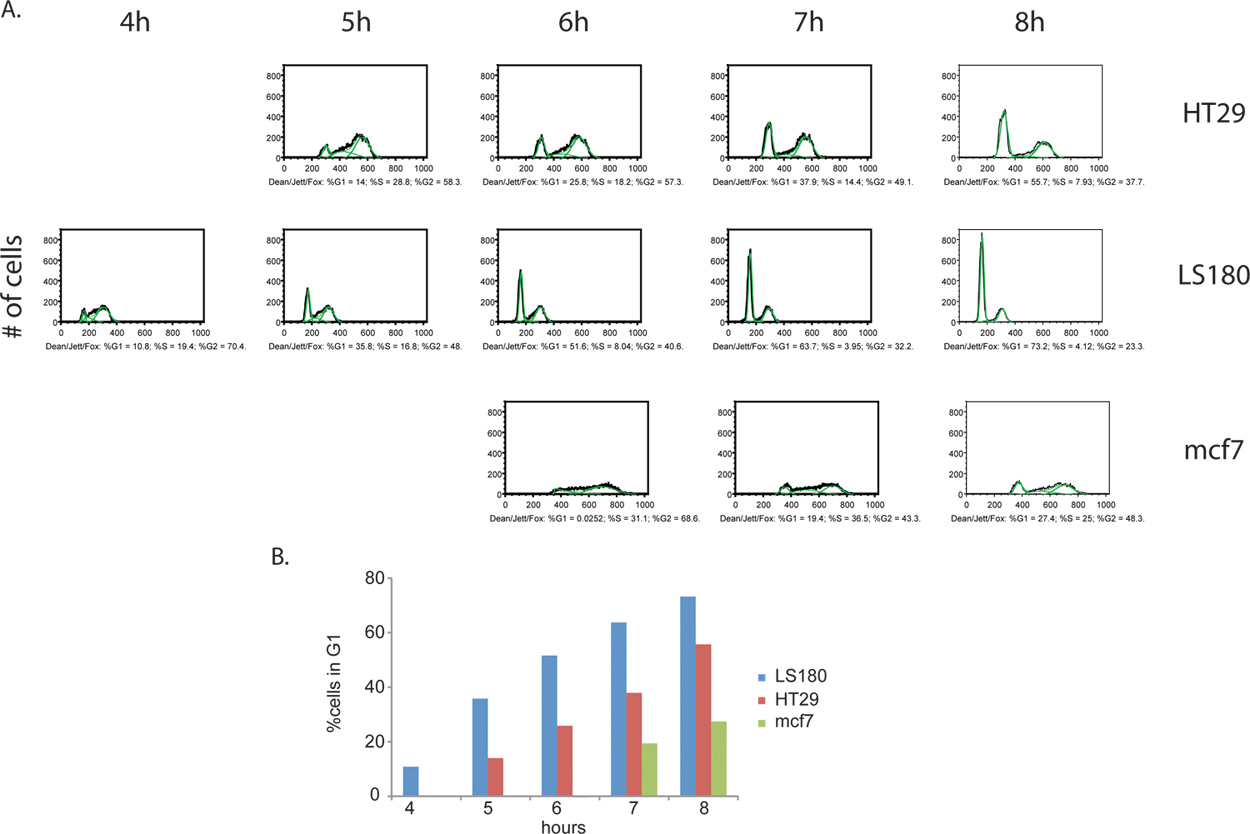
追加情報はBrdU標識ステップから派生させることができます。積極的に分裂する細胞の割合は、ESTは、測定することができますが、だけでなく、2つのサンプル間の細胞周期ステージの分布のimateも同様に決定することができます。彼らはG2にサイクル、進歩を続けているBrdUの試薬の除去後に等間隔で細胞を集めることによって、細胞を追跡することができ、最終的に最終的にG1とBrdU陽性娘細胞として浮上し、元の細胞の細胞分裂に至るまで移動するDNA含有量( 図4)。細胞周期の位相の定量化と運動解析の例を図5に記載されています。

図1代表的なMCF7細胞集団の(A)前方散乱対側の散布図。代表的なMCF7細胞集団の(B)PI対幅(FL-W)をプロット。ゲーティングは最終的な分析(R3)でセルのダブレットを除外することが示されている。彼らはレーザービームとtheref通過に時間がかかるように細胞ダブレットは、単一のセルよりも大きいパルス幅を持っています。鉱石は、分析から除外することができます。代表的なMCF7細胞集団の(C)PIに対するFITC(BrdU)のプロット。ゲーティングは、FITC(BrdU)の陽性細胞(R4)を含むように示されています。

図2通常の細胞をサイクリングの総人口の代表的なヒストグラムプロット。

図3 FITC(BrdU)の陽性細胞のゲーティングした後、BrdUのパルスを除去した後1時間を収集された細胞のヒストグラムプロット。ラベルの除去後早期のタイムポイントでのBrdU陽性細胞は、DNA合成中の細胞の正常なラベルを確認し、細胞周期のS期にある細胞を用いた一貫性のあるDNAのコンテンツを表示するサンプルに対応するPIプロファイルを示しています。
 LT = "図4" />
LT = "図4" />
図4の癌細胞株、大腸癌HT29(A)及びLS180(B)、ならびに乳癌MCF7(C)との間の細胞周期進行動態の比較。細胞は、BrdUのパルスを除去した後、8時間ごとに時間を収集した。この実験では、大腸の細胞株LS180における細胞周期のG2期を通して加速、細胞周期の進行の明確なプロファイルを観察した。大腸癌細胞の2つのプロファイル間で速度を比較すると、G1ピークの出現はこのピークを欠いているHT-29細胞株に対応する時点と比較して、LS180細胞でT = 4H後のBrdUで明らかである。 2大腸の細胞株、MCF7細胞のいずれかと比較して大幅に減少率でサイクリングされています。 拡大図を表示するには、ここをクリックしてください 。
再度5 "/>
図5:BrdU標識癌細胞の(A)定量的細胞周期の位相解析。ディーン/ジェット/フォックスアルゴリズムは、HT29、LS180およびMCF7(緑色で示す)に適用した。各サンプルから得られた細胞周期の位相分布は、各フェーズの%合計BrdU陽性細胞として表現されます。各細胞株のための唯一の選択の時点では、無効な結果が得られた以前の時点のいくつかの分析として、表示されます。これらの以前の時点では、すべてのBrdU陽性細胞はS期にあり、したがって、アルゴリズムがアプリケーションに必要な個別のG1とG2のピークを、欠けている。 (B)癌細胞のヒストグラムは、細胞周期のG2 / M期を通して進行の動態を示す。 G2 / M期を通じて進行はHT29およびMCF7続いLS180細胞のために最も速くなります。 拡大図を表示するには、ここをクリックしてください 。
JPG "ALT ="図S1 "/>
図S1。フローサイトメトリーを制御します。 MCF7細胞は、細胞がPIでもBrdUの染色でもない(A)ネガティブコントロールとして表示されます。 (B)PIのみ染色した。 (C)のBrdU-FITC標識したサンプルは、。

PIに対するFITC(BrdU)の蛍光発光スペクトルの関係を示すS2。模式図を示します。蛍光チャネル1(FL1、FITC)と蛍光チャネル3(FL3、PI用)スペクトル収集ウィンドウは、対応するボックスに表示されます。 FL1とFL3検出の間には、蛍光体のスペクトルの重複はありません。 PIとFITC-BrdUのから実験データの共標識された細胞は、それぞれFL1/FL3チャネルで収集されたとき明らかに、蛍光補正は必要ありません。蛍光スペクトルは、BD Biosciences社のWebサイトから得られた。ターゲット= "_blank"> http://www.bdbiosciences.com/research/multicolor/spectrum_viewer。 拡大図を表示するには、ここをクリックしてください 。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
BrdUの取り込みをフローサイトメトリーを組み合わせることにより、我々は細胞周期動態を検討するために必要なツールを持っています。チミジンのアナログとして機能するBrdUの独特のプロパティは、サイクリング細胞のDNA含量の定量を可能にするものです。細胞周期の合成フェーズで成長娘DNA鎖へのBrdUの取り込みは1つがG2での成長期に、最終的に細胞分裂に、S期におけるDNAの複製を介して循環細胞の集団を追跡することができる内容です。 BrdUのラベルは信号がバックグラウンドレベルに減少する程度に細胞分裂によって希釈された時点で、その時点までは娘のBrdU陽性細胞は継続的に続けることができます。
全DNA含有量を決定するためにヨウ化プロピジウムを用いて、S→G2 / M→G1から細胞周期の進行をフォローすることができます。細胞の循環率を追跡する能力は、多くのアプリケーションで特に重要なのfoと便利です。新規化合物11の細胞周期の段階の感度を決定するために細胞周期特異的なDNA損傷を誘発する薬や薬剤開発プロセスの間を含むrの前臨床試験。代謝標識法を用いての最も明白な利点は、細胞の化学物質の同期の除去です。細胞周期停止を誘導する最も化学的処理自体が遺伝毒性であるので、これらの化学物質の存在下での新規化合物の細胞周期の感度の影響を解剖すると、可能でないかもしれません。
我々は、PI共染色、このBrdUの代謝標識法の汎用性を実証した。このプロトコルを使用して、我々は正常細胞周期のG2 / M期を通じて細胞周期の進行の増加は酵素発現10を強制しているように形質転換されたテロメラーゼ陰性ヒト細胞で観察することができることを実証した。このメソッドは、サイクルthroのように細胞の動態を測定する際に強力ですが複数のステージぐふっ、一つの制限は、遷移時のタイミング情報の損失だけでなく、G2およびM期の範囲内である。細胞がMにG2を進めるように、我々は単にDNAの内容に基づいて、二つの相だけでなく、区別することはできませんが、またサブ-G1集団。この差別化が要求されるアプリケーションでは、式の細胞周期12,13の所望の位相に固有であるサイクリンまたは他の細胞タンパク質に対する抗体との共染色のBrdU標識細胞の潜在的可能性があります。追加の最適化はサイクリン抗体はBrdUの標識と免疫染色のプロトコルと互換性があることを保証するために必要となります。理想的には、十分に細胞周期のさまざまな段階を区別するために、いくつかのサイクリン抗体は単一のサンプルを同時に共存ラベルに必要になることがあります。当研究室では、細胞を区別するためにサイクリンBとリンヒストン3抗体とBrdUのパルス標識細胞のポストラベリングに取り組んでいます細胞周期のG2対M相インチPIとFITC-BrdUの染色に加えてサイクリン抗体との共標識は、実質的にハードウェアとデータ分析(蛍光補正の問題)の両方の観点から、フローサイトメトリーの合併症を増加させます。フローサイトメーターの新世代の到来とともに、能力と機能の進歩は、これらの技術的な懸念を緩和する必要があります。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
我々は、開示することは何もありません。
Acknowledgments
我々はFACS分析の支援についてはUBCで医学研究センターのアンディ·ジョンソンに感謝します。ウォン研究所のがん研究の資金は、カナダの癌協会総合研究所(オペレーティング·グラント#019250)でと薬学、UBCの学部の研究再投資資金から提供されています。 JMYWは、カナダリサーチチェアと保健研究キャリア開発プログラムのマイケル·スミス財団によってサポートされています。
Materials
| Name | Company | Catalog Number | Comments |
| bromodeoxyuridine | BD Biosciences | 55089 | |
| propidium iodide | Sigma-Aldrich | 287075 | 1mg/ml stock |
| FITC anti-BrdU | BD Biosciences | 347583 | |
| sodium tetraborate | Fisher Scientific | S80172 | 0.1M, pH 8.5 |
| FACS Caliber | BD Biosciences |
References
- Musgrove, E. A., Caldon, C. E., Barraclough, J., Stone, A., Sutherland, R. L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer. 11, 558-572 (2011).
- Molchadsky, A., Rivlin, N., Brosh, R., Rotter, V., Sarig, R. p53 is balancing development, differentiation and de-differentiation to assure cancer prevention. Carcinogenesis. 31, 1501-1508 (2010).
- Dickson, M. A., Schwartz, G. K. Development of cell-cycle inhibitors for cancer therapy. Curr. Oncol. 16, 36-43 (2009).
- Banfalvi, G. Cell cycle synchronization of animal cells and nuclei by centrifugal elutriation. Nat. Protoc. 3, 663-673 (2008).
- van Opstal, A., Boonstra, J. Inhibitors of phosphatidylinositol 3-kinase activity prevent cell cycle progression and induce apoptosis at the M/G1 transition in CHO cells. Cell Mol. Life Sci. 63, 220-228 (2006).
- Gasnereau, I. Flow cytometry to sort mammalian cells in cytokinesis. Cytometry. A. 71, 1-7 (2007).
- Harper, J. V. Synchronization of cell populations in G1/S and G2/M phases of the cell cycle. Methods Mol. Biol. 296, 157-166 (2005).
- Pedrali-Noy, G. Synchronization of HeLa cell cultures by inhibition of DNA polymerase alpha with aphidicolin. Nucleic Acids Res. 8, 377-387 (1980).
- Merrill, G. F. Cell synchronization. Methods Cell Biol. 57, 229-249 (1998).
- Fleisig, H. B., Wong, J. M. Telomerase promotes efficient cell cycle kinetics and confers growth advantage to telomerase-negative transformed human cells. Oncogene. , (2011).
- Cai, D., Byth, K. F., Shapiro, G. I. AZ703, an imidazo[1,2-a]pyridine inhibitor of cyclin-dependent kinases 1 and 2, induces E2F-1-dependent apoptosis enhanced by depletion of cyclin-dependent kinase 9. Cancer Res. 66, 435-444 (2006).
- Pozarowski, P., Darzynkiewicz, Z. Analysis of cell cycle by flow cytometry. Methods Mol. Biol. 281, 301-311 (2004).
- Sherwood, S. W., Rush, D. F., Kung, A. L., Schimke, R. T. Cyclin B1 expression in HeLa S3 cells studied by flow cytometry. Exp. Cell Res. 211, 275-281 (1994).

{kind=link}
{kind=link}