

Summary
우리는 살려서 살짝 익히고의 컬렉션을 설명
Abstract
대부분의 생물은 공간적 제어 유전자 발현에 관련된 세포 내 목적지로의 mRNA를 집중. 최근의 연구는 사체의 대부분은 세포와 배아 랜덤하지 않은 위치로 지역화 된 것을 증명하고있다. 지역화의 mRNA를 확인하는 한 가지 방법은 화학적으로 관심 세포 구조를 정화하고 모든 관련 증명서를 확인하는 것입니다. 최근에 개발 된 높은 처리량 시퀀싱 기술을 사용하여 지금은 세포 내 구조와 관련된 모든 RNA를 식별 할 간단합니다. 성적 식별을 용이하게하면은 완전 서열 게놈을 가진 유기체로 작동 할 필요가 있습니다. 세포 내 구조물의 생화학 정화에 대한 하나의 매력적인 시스템은 개구리 Xenopus의 laevis의에서 생산되는 계란 추출물이다. 그러나, X. laevis의 현재 성적 식별을 방해 완전히 순서가 게놈을 가지고 있지 않습니다. 이 문서에서 우리는 방법을 설명관련 개구리, X.에서 계란 추출물을 생산하는 완전히 순서가 게놈을 가지고 tropicalis. 우리는 미세 소관의 중합, 정화 및 성적 증명서 격리에 대한 세부 정보를 제공합니다. 이 문서가 미세 소관 - 관련 증명서의 식별을위한 특정 방법을 설명하는 동안, 우리는 쉽게 다른 세포 내 구조물에 적용되고 지역화 된 RNA를 식별하기위한 강력한 방법을 제공 할 것이라고 믿습니다.
Introduction
유전자 발현의 시공간적 제어 모든 셀에 대한 중요하고, 1 패터닝 초기 배아의 컨트롤에 특히 중요합니다. 유전자 발현의 공간 제어 세포 또는 배아 내의 특정 목적지로의 mRNA의 적극적인 현지화를 통해 이루어집니다. 많은 매우 큰 세포 유형에서 (예를 들어, 난자, 배아, 그리고 뉴런) mRNA의 현지화 코딩 단백질의 작용 사이트로 단백질 발현을 제한하는 데 사용됩니다. 지역화 된 mRNA의 단백질 생산의 많은 라운드를 촉진 할 수 있기 때문에 그것은 개인의 단백질 분자를 지역화하는 것보다 발현을 집중하는 것이 더 효율적입니다. 그들은 더 코딩 단백질 2의 국산화를 제한하는 역할을 목적지에 도달 할 때까지 현지화의 mRNA는 일반적으로 translationally 억압된다. 배아 패턴을 제어하는 RNA 지방화의 많은 잘 문서화 된 경우뿐만 아니라, 여러 연구가 현지화의 mRNA를 문서화인코딩 된 단백질의 작용 사이트에 게시합니다. 눈에 띄는 예는 운동성 섬유 아세포의 선두 가장자리에 β-액틴 3 Arp2 / 3의 mRNA의 현지화 및 감수하고 유사 분열 스핀들 5-7 많은 유사 분열 레귤레이터의 mRNA의 현지화 (가) 있습니다.
지역화의 mRNA의 고전적인 사례의 대부분은 산모 효과 돌연변이 유전자 화면을 통해 확인되었다 나중에 지역화 된 RNA를 인코딩하기로 결정 하였다. 그러나 최근의 게놈 넓은 연구 지역화의 RNA 범위로 폭 넓은 통찰력을 제공하기 시작했다. Drosophila의 배아 원위치 하이브리드 화면에서 최근 모두의 mRNA ~ 70 % 많은 소설 대상 8 등 특정 지역화를 가지고 것을 보여 주었다. 마우스 섬유 아세포에서 pseudopodia의 정화 지역화의 mRNA 9의 다양한 그룹을 확인했다. 감수 제노에서 미세 소관의 생화학 적 정제를 사용하여 우리 그룹에서 작업고름 계란 스핀들 5,7와 copurify의 mRNA 확인 수백을 추출합니다. 우리의 작업의 mRNA가 코딩 된 단백질의 작용 사이트에 지역화 된 아이디어를 지원하는 미세 소관 지역화의 mRNA의 대부분은 유사 분열을 제어하는 기능 단백질을 인코딩 것으로 나타났다. 또한, 생화학 정화하여 세포 내 부분에서 mRNA의 농축을 감지하는 능력은 지역화의 mRNA의 식별이 방식의 힘을 강조한다.
대부분의 지역화 된 RNA를 자신의 최종 목적지 10 전송을 달성하기 위해, 굴지 또는 소관하거나, 세포 골격에 능동 수송을 사용합니다. 이 골격 프로세스를 요점을 되풀이 할 수있는 생체 시스템을 가지고하는 데 필요한 생화학 적 방법을 사용하여 특정 목적지로 지역화 된 RNA를의 범위와 유형의 더 나은 이해를 얻을합니다. 골격 생물학을 공부를위한 최고의 시스템 중 하나는 생산 된 계란 추출물입니다개구리 Xenopus의 laevis의. X.에서 수정란에서 laevis의 계란 추출물은 세포 골격 프로세스의 다양한 배열을 공부하는 수십 년 동안 사용되어 왔습니다 및 골격 조립 및 역학 11을 제어하는 메커니즘을 분자에 대한 우리의 이해에 많은 기여했다. 또한, X. laevis의 계란 추출물은 미세 소관과 관련 단백질 12,13의 대규모 정화 모든 의무이며, 계란 추출물 14-16의 각종 유형의 생산을위한 잘 설계 방법이 있습니다. 그러나 게놈 연구에 X의 사용에 몇 가지 단점이있다 모델 시스템으로 laevis의.
수십 년 동안 Xenopus의 laevis의 개구리는 큰 난자의 크기와 강력한 외부 개발 17 때문에 발달 및 세포 생물학의 연구를위한 강력한 시스템이었다. 또한, 많은 세포 PROCESSE 요점을 되풀이 할 수 계란 추출 시스템의 개발테스트 튜브의이 개구리에게 강력한 실험적인 모델을 만들었습니다. 그러나 Xenopus의 laevis의이 genome.In 대비의 allotetraploid 자연 둔화 된 완전한 게놈 서열의 부족에 의해 방해되었습니다 밀접한 관련이 종 Xenopus의 tropicalis는 2010 년 18 서열화 된 이배체 유전체 있습니다. 반면 X. tropicalis는 X.만큼 실험적으로 다루기 쉬운되지 않습니다 laevis의 17 시퀀스 게놈의 가용성은 게놈 넓은 분석을 수행 할 수있는 매력적인 모델 시스템을 만든다.
이 보고서에서 우리는 X.에서 감수 분열 II-, 세포 증식 억제 인자 체포 추출물 (CSF)를 확인하는 방법을 설명 tropicalis 19. 우리는 다음이 추출물에서 미세 소관과 관련 RNA를 정화하는 간단한 방법을 설명합니다. RNA를 그런 다음 최근에 개발 된 높은 처리량 시퀀싱 기술을 사용하여 시퀀싱 의무 라이브러리로 변환 할 수 있습니다. 라이브러리 회그들은 총 추출물에 비해 미세 소관 샘플에 충실 특정의 mRNA를 식별하는 개구리의 게놈에 정렬 할 수 있습니다 순서가 있습니다. 이 게놈 넓은 규모의 소관 표적 mRNA의 현지화를 감지 할 수있는 강력한 방법을 제공합니다. 지역화의 mRNA를 감지 할 수있을뿐 아니라, 높은 처리량 시퀀싱 및 시퀀스 게놈의 사용은 공공 데이터베이스 주석에 현재 존재하지 않는 새로운 사본을 발견의 가능성을 제공합니다.
Protocol
1. X. tropicalis 계란의 생성
모든 Xenopus의 tropicalis 개구리는 NASCO에서 정렬되었습니다. 우리의 개구리는 재순환 시스템은 27로 유지 수생 서식지에 보관됩니다 ° C. X의 관리를위한 급수 시스템을위한 많은 옵션이 있습니다 tropicalis가.이 개구리 종에 대한 몇 가지 좋은 일반 정보 (할 랜드와 그레인저 연구소의 웹 사이트에서 찾을 수 있습니다 http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). 우리의 개구리는 (0.4 g Ciclid 호수 소금 0.6 g 바다 소금, 물, 산도 7.0의 리터 당 0.625 g NaHCO3를) 20로 구성 tankwater에 유지됩니다. X.에 대한 높은 염분이다 ~ 1,800 μS의 전도도,이 제조법 결과 tropicalis. 그러나, 우리는 우리의 개구리 생에 성공할 것으로 나타났습니다의 환경과 난자의 질이 향상됩니다. 다른 tankwater 조리법은 일반 X 용 나열된 자원에 위 확인할 수 있습니다 tropicalis 관리.
- 개구리가 누워 계란을 자극하는 세 개의 연속적인 일에 융모 성선 자극 호르몬 (hCG를) 주입된다 : 첫째, hCG의 솔루션의 두 가지 농도를 준비합니다. 1000 U / ML의 최종 농도 10 ML 살균 이온 H 2 O의 동결 건조 된 hCG의 분말을 Resuspend 10,000 U. 다음, 100 U / ML의 최종 농도 10 ML H 2 O 1,000 U / ㎖ hCG의 용액 1 ㎖를 희석. 4 ° C.에서 두 솔루션을 저장
- 1 일에 2시부터 오후 3시까지 사이 hCG를 함께 주입하여 누워 계란을위한 4-6 개구리를 준비합니다. 0.2 ML 100 U / ML hCG를 솔루션으로 배설강 주변 등의 림프 낭의 각 개구리를 주입. 이후 두 주사하는 동안 개구리가 빠른 데 누워 계란 중에 개구리 폐기물 현재의 양을 최소화하지만, 선택적이다.
- 하루에 2 같은 개구리를 주입2시에서 오후 3시까지 사이에 0.2 ML 100 U / ML hCG를 솔루션.
- 3 일에 오전 7시에서 10시까지 사이에 0.2 ML 1,000 U / ㎖ hCG의 솔루션과 같은 개구리를 주입. 신선한 tankwater와 6 쿼트 플라스틱 물통을 채우기, 25 어둠 속에서 개구리와 장소를 추가 ° C. : 알을 낳기 위해 개구리를 설정 이 주사 후, 달걀 누워 4 시간 후에 시작하고 7 시간에 완료됩니다. 개구리 25의 최소 유지되는 환경에서 계란 ° C.를 배치한다
- 솔루션을 추출하고 계란을 수집하기 직전에 준비 장비를 확인합니다.
20X MMR : 100 MM의 HEPES, pH를 7.8, 2 mM의 EDTA (에틸렌 다이아 민 테트라 초산) 산도 7.8, 2 M NaCl을, KCl을 40 밀리미터, 20 밀리미터 MgCl 2, 40 MM의 염화칼슘 2. 상온에서 오토 클레이브 및 저장. 준비를 추출하기 직전 1X의 MMR 1 L를 준비합니다.
10X XB : 100 MM의 HEPES, pH7.7, 10 MM MgCl 2, 1 mM의 염화칼슘, 1 M KCl을, 500 MM의 자당. 4 ° C.에서 압력솥 및 저장 1X XB J 1 L를 준비준비를 추출하기 전에 UST. Dejelly 솔루션 : 10 N NaOH를 가진 7.8-8.0에 탈 H 2 O와 산도의 250 ML 3 % 시스테인 솔루션을 준비합니다. 준비를 추출하기 직전에 준비합니다.
CSF-XB은 : 1X XB 200 mL를 취하여 2 ML 0.5 M EGTA 산도 7.7, 200 ㎕의 1 M MgCl 2를 추가합니다. 준비를 추출하기 직전에 준비합니다.
CSF-XB + : CSF-XB 50 mL를 취하여 LPC의 50 μl를 (10 ㎎ / ㎖ 각 펩틴, Pepstatin의 주식, 그리고 DMSO에 Chymostatin)을 추가합니다. 50 μL 사이토 D을 (DMSO에서 10 ㎎ / ㎖)를 추가합니다. 준비를 추출하기 직전에 준비합니다.
소독을 녹여 필터링 탈 H 2 O, 전자 레인지의 0.2 % 젤라틴 용액을 준비합니다. 상온에서 보관하십시오.
예약 2 베크 2 × ½ 인치 초원 심 분리기 튜브.
쿠션 각각 H 2 O의 0.5 ML로 두 개의 15 mL 유리 둥근 바닥 원심 분리기 튜브를 준비합니다튜브를 원심 분리기.
화재 광택 유리 파스퇴르 피펫을 확인합니다. 넓은 구멍을 노출하는 5 ¾ 인치 유리 피펫의 끝을 스냅, 새로운 노출 피펫 팁을 부드럽게 화염에 노출.
- 코트 비커의 벽을 주위에 0.2 % 젤라틴 용액을 소용돌이로 계란을 저장하기 위해 500 ML 유리 비커를 준비합니다. 사용 후 커에서 젤라틴 용액을 버린다.
- 3 일의 세 번째 주사 후 6-7 시간을 마련하는 데 사용되는 플라스틱 통에서 계란을 수집합니다. 원하는 경우, 부드럽게 남아있는 계란을 한 번 각 개구리를 짠다. 신선한 tankwater 0.2 % 젤라틴 용액으로 코팅 된 500 mL 유리 비커로 전송 한 번 계란을 씻으십시오.
2. X.에서 추출의 준비 tropicalis 계란
추출 준비의 모든 단계는 약 25 ° C. 상온에서 수행 할 수 있습니다 세척을 통해, 그것은 계란을 유지하는 것이 중요합니다그들은 젖은 남아 있도록의 액체에서 침수. 공기에 노출되면 계란 세포주기 정지를 탈출하거나 용해가 발생할 수 있습니다.
- 계란 젖은 유지하기에 충분한 액체를 예약하는 동안 가능한 한 많은 tankwater로 가만히 따르다. 옆에 계란을 포함 비커를 기울여 비커의 벽에 천천히 ~ 300 ML 1X MMR을 추가, 계란 있도록 물리적 교반을 최소화 할 수 있습니다. 상층 액을 함유 한 부스러기 다음 가만히 따르다, 계란 정착하자. X. tropicalis 계란이 단계에서 힘줄, 그래서 활성화 된 계란의 제거는 dejellying 후에 완료됩니다. 세 1X MMR 세척의 총 반복합니다.
- Dejelly 계란. dejelly 솔루션의 가능한 추가 반만큼 MMR로 떨어져 가만히 따르다. 약 5 분 동안 지속적으로 소용돌이. 젤리 코트를 용해하는 것은 몇 분 후 상층 액에 볼 수 있습니다. 오프 가만히 따르다하고 나머지 dejelly 솔루션을 추가합니다. 계란은 매우 밀접하게 포장 때까지 계속 소용돌이에 계속해서 자신의 식물 극 (모든 방향 극백색 안료)와 접시의 바닥을 향해. 가능한 한 빨리 많은 dejelly 솔루션으로 떨어져 가만히 따르다. 계란 dejellied되면 그들은 기계적 조작에 매우 민감하다.
- 조심스럽게 계란에 XB를 추가합니다. 첫 번째 XB 세척에 용해, 푹신한, 흰색 및 pseudocleavage 계란을 제거하여 CSF의 체포를 탈출 계란을 제거합니다. 활성화 X. tropicalis 계란 상단 중앙에 정착하는 경향이, 그래서 이것들을 당겨 플라스틱 전송 피펫을 사용합니다. 또한 피부와 개구리 폐기물의 조각을 제거합니다. 부드럽게 세척 사이에 알을 소용돌이들이 비커의 바닥에 정착 할 수 있도록 ~ 300 ML 1X XB 솔루션과 계란을 세 번 총을 씻으십시오. 가능한 한 각 세척 용액의 계란 젖은 유지하면서 이전과 같이, 가만히 따르다.
- CSF-XB와 가만히 따르다 두 번 계란을 씻으십시오.
- CSF-XB 추가 + 계란에. 전자를 노출하지 않도록주의하면서, 젤라틴 처리 화재 광택 파스퇴르 피펫, CSF-XB +로 전송 계란 초 원심 분리기 튜브로 사용공기 GGS. 물 쿠션 15 mL 유리 원심 분리기 튜브 내부에서.
- 1 분 200 XG에서 임상 원심 분리기에 계란을 회전, 30 초 동안 800 XG와 스핀 속도를 높일 수 있습니다.
- 계란에서 가능한 한 많은 버퍼를 제거하기 위해 흡인기를 사용합니다. 그들은 정상에 거의 건조해야한다. 신속 HB-6 회 (또는 이에 상응하는)가 장착 Sorvall RC-6 원심 분리기 계란을 이동하고 20에서 15 분 동안 17,000 XG에 스핀 ° C.
- 1 ML의 주사기에 부착 된 18 게이지 바늘을 사용하여 색소와 지질 층 사이 노란색 세포질 레이어를 제거합니다. 구멍은 튜브의 측면 천천히 세포질 추출물 계층을 얻기 위해 주사기 배럴을 당깁니다. 색소 과립 가능한 한 피하십시오.
- 새로운 원심 관에 세포질을 전송합니다. 추출이 단계에서 약간 흐린 나타나는 것이 정상입니다. 15 mL 유리 원심 분리기 튜브 물 방석 안쪽으로 배치합니다. 20 ℃에서 10 분 동안 다시 17,000 XG에 스핀 내선을 반복18 게이지 바늘로 raction.
- 1.5 ML의의 microfuge 튜브에 세포질을 전송합니다. 추출 양을 추정 추출물에 사이토 D 및 LPC 1:1,000 희석. , 1 ML 피펫 팁을 잘 섞어 공기 방울을 도입하지 않고 여러 번 위아래로 피펫합니다. 건강한 개구리 서식지의 전형적인 수율은 추출 / 개구리 약 300 ~ 500 μL입니다. 최대 활성을 유지하려면, 그것은 추출물을 저장하고 상온에서 실험 조작 (20-25 ° C)를 수행 할 필요가 있습니다.
3. X.에서 정화 탁솔 안정화의 미세 소관 tropicalis 추출
- 10 μM의 최종 농도에서 추출 100-200 μL 나누어지는에 탁솔 추가하고 30 분 실온에서 알을 품다. 제어 반응, 미세 소관 destablilzing 약 Nocodazole (10 μM)와 추출물의 동등한 볼륨을 처리합니다. 분석을위한 치료 추출물의 예약 100 μL.
- 약물 치료를 희석10 권 BRB-80 (80 MM의 파이프 산도 6.8, 1 mM의 MgCl 2, 1 mM의 EGTA) + 30 % 글리세롤로 추출합니다. BRB-80 + 60 % 글리세롤 쿠션의 10 ML을 포함하는 14 ML 둥근 바닥 폴리 프로필렌 튜브를 조립합니다. 넓은 구멍 피펫 팁, 레이어 BRB-80의 상단에 부드럽게 약물 치료 추출물 반응 + 60 % 글리세롤 쿠션을 사용. 20 17,000 XG에서 10 분 동안 원심 분리기 ° HB-6 회 (또는 동급) 및 튜브 어댑터가 장착 된 Sorvall RC-6 원심 분리기 C.
- 상층 액을 포함 unsedimented 추출 물질을 흡입하고, 탈 H 2 O로 두 번 인터페이스를 씻어 미세 소관, 미세 소관 - 관련 단백질과 탁솔 처리 샘플의 미세 소관 관련 RNA를을 포함하는 젤 같은 펠렛을 방해하지 돌보는, 천천히 나머지 쿠션 볼륨을 대기음. Nocodazole 처리 샘플 보이는 물질을 포함하지 않습니다. 1 ML TRIZOL에서 펠렛을 resuspend을하고 RNA를 분리 제조업체의 지시 사항을 진행합니다. Untreated 추출물 (100 μL) 1 ML의 TRIZOL에서 직접 재현 탁 할 수 있습니다.
- RNA-SEQ에 적합한 사체 라이브러리를 준비 시판 키트는 이제이 있습니다. 이들은을 통해 구입하실 수 있습니다 http://www.illumina.com/ 및 http://www.454.com/ .
Representative Results
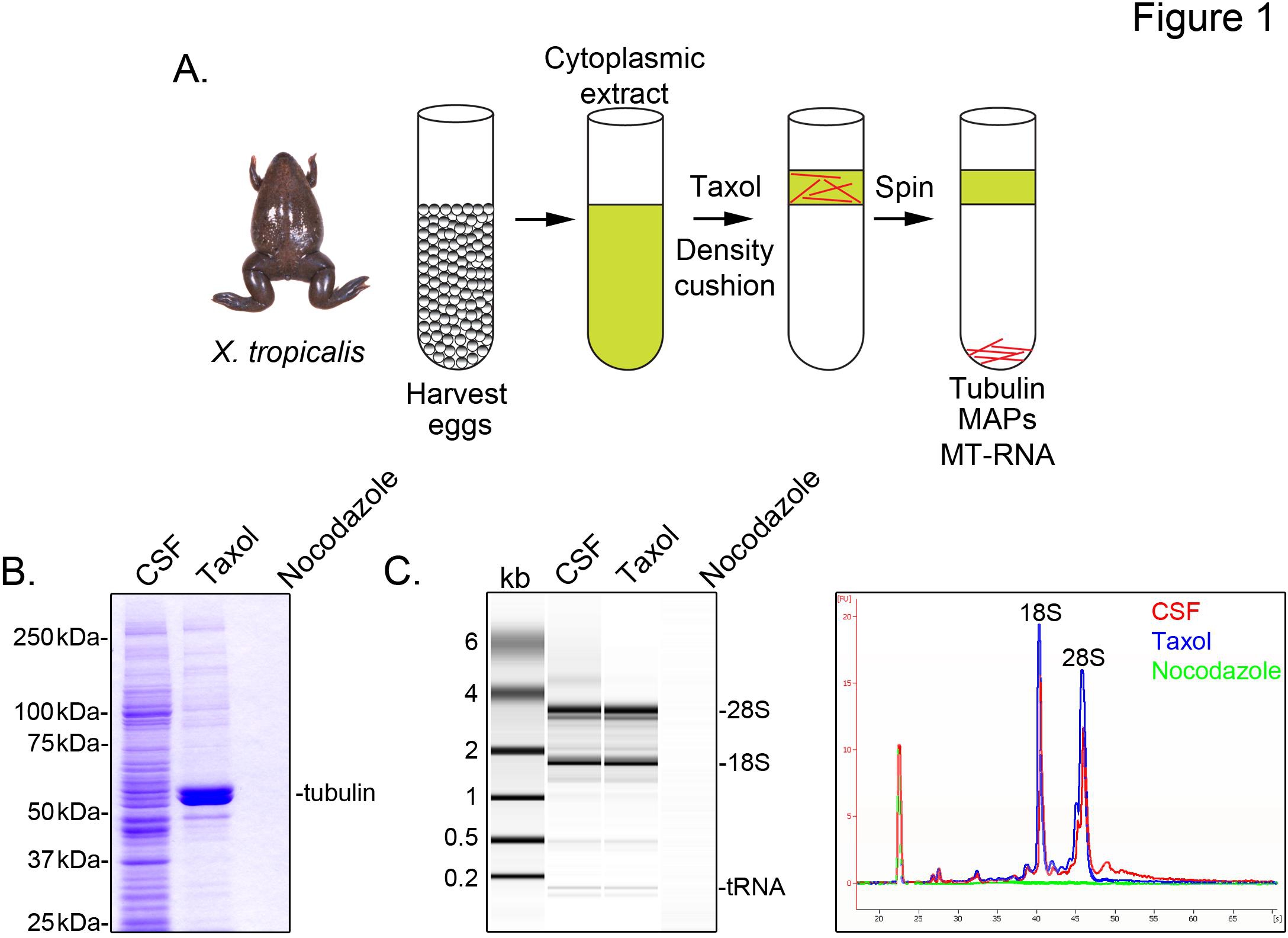
X를 식별하려면 미세 소관과 관련된 tropicalis 성적 증명서, 우리는 감수 분열 II (CSF)의 중기에서 체포 난자의 세포질 추출물을 준비합니다. 택솔 (taxol)이 추출물의 치료 글리세롤 쿠션 (그림 1A)를 통해 침전으로 정제 할 수있는 안정적인 미세 소관의 형성을 할 수 있습니다. 매시 젤 분석 α / β-튜 블린의 택솔 (taxol)에 의존하는 방식으로 퇴적물 및 주요 단백질의 종을 나타내는이 준비 (그림 1B)에서 복구하는 확인합니다. 다른 단백질의 낮은 수준을 나타내는 약물 nocodazole을 해중합 소관으로 처리 준비도 탁솔 펠렛에 존재하지만 그 소관 (MAP에)와 특별히 연관 탁솔 부분에 단백질.에게
애질런트 Bioanalyzer 모든 X에서 일반적으로 RNA 구성을 검사하는 데 사용됩니다 tropicalis 추출물 분획 (그림 1C X에서 미세 소관과 스핀들에 발생하는 이전의 연구 결과와 일치, CSF 추출물과 미세 소관을 함유 택솔 (taxol) 펠렛에 존재하는 laevis의 계란 5,21 압축을 풉니 다. 젤 투사의 라인 추적은 mRNA의 신호에 현저하게 낮은 보여 미세 소관을 함유 표시, 28S rRNA의 위의 영역 마이그레이션 특히 택솔 (taxol) 펠렛을, 그 X에서 미세 소관과의 mRNA의 cosediment의 하위 집합 tropicalis. 이러한 방식으로 절연 RNA는 시판 시약을 사용하여 RNA-SEQ 실험에 적합합니다.

그림 1. RNA-SEQ를위한 MT-RNA의 정제. MT-RNA를 분리하는 (A) 정화 방식. 계란은 여성의 X.에서 수확 TROpicalis 개구리. 세포질 추출물의 준비를 한 후, 택솔 (taxol)은 미세 소관의 중합을 유도하기 위해 추가됩니다. 미세 소관과 MT-RNA는 글리세롤 쿠션을 통해 침전에 의해 정화된다. (A)에 설명 된 방식을 사용하여 격리 된 단백질 (B) 매시 젤 분석. 총 CSF는 탁솔 또는 nocodazole의 면전에서 침전 단백질에 비해 압축을 풉니 다. (C) RNA의 Bioanalyzer 젤 분석 (A)에 설명 된 방식을 사용하여 격리. CSF에서 RNA 절연 탁솔 또는 nocodazole의 면전에서 침전 RNA에 비해 압축을 풉니 다. 젤 프로젝션 라인 트레이스가 모두 표시됩니다. 샤프, 등., (2011)의 허가 재판은. 큰 이미지를 보려면 여기를 클릭하십시오 .
Discussion
이 보고서에서 우리는 X.에서 CSF-체포 계란 추출물을 생산하는 간단한 방법을 설명했습니다 tropicalis 19은 미세 소관 관련 RNA를에게 7을 연구하는이 추출물을 사용합니다. X.에서 CSF-체포 계란 추출물 생산을위한 기본 절차 X. 위해 사용되는 tropicalis는 동일합니다 몇 가지 주요 차이점 laevis의. X. 작업에 가장 어려운 부분 중 하나 tropicalis 개구리는 X.에 비해 미세 소관 핵 또는 스핀들 어셈블리 활동과 추출을 할 수있을 정도로 충분한 고품질의 계란을 획득합니다 laevis의 계란 추출합니다. 감수 분열 II 세포주기 정지, X 용 호르몬 주사 사이의 간격에서 미끄러짐을 방지하면서 최적의 달걀 누워 조건을 달성하기 위해 tropicalis는 X.에 사용되는보다 짧은 laevis의, 그리고 달걀 누워의 시작 제 hCG를 주입의 시간도 훨씬 단축됩니다. X가 hCG를 주사에서 t에 타이밍을 laevis의그는 달걀 부설의 시작은 버퍼에 하룻밤 배치 할 계란 편리하고 효율적 그런 것입니다. 그러나, hCG를 주사와 계란 사이의 짧은 시간의 X. 누워 그것은 자주 수동 개구리의 알을 표현하는 것이 필요하다 tropicalis. 두 개의 서로 다른 개구리의 난자 추출물을 만들기의 또 다른 중요한 차이점은 dejellying 단계입니다. X가 계란 laevis의는 젤리 코트 계란 비커에 간격이 얼마나 밀접하게 관찰하여 녹일 때를 결정하기 쉬운 너무 큽니다. dejellying 반응이 시작될 때, 계란을 더 조밀하게 포장하기 시작합니다. 그러나 X. tropicalis 계란 훨씬 작하고 젤리 코트 혼자 밀도를 포장 달걀에 의해 용해 될 때를 결정하는 것은 매우 어려울 수 있습니다. 우리는 젤리 코트 해산 시점을 확인하는 가장 신뢰할 수있는 방법은 동물 (검정)과 식물 (흰색) 극의 방향을 모니터링하는 것으로 나타났습니다. 시 식물 극 오라비커의 아래쪽으로 ENT는 젤리 코트는 추출을 진행하기에 충분한 제거되었습니다. 마지막으로, 반면 X. laevis의 계란 추출물은 차가운 온도에서 저장할 수 있습니다 (4-12 ° C) 우리는 X를 유지하는 것이 중요합니다 것을 관찰 준비와 실험 조작 동안 실온에서 tropicalis 달걀 추출물 (20-25 ° C)의 생화학 적 활성을 유지한다. 때문에 사용의 용이성의 차이를 우리는 X를 사용하는 것을 선호 계란 추출물의 생산을위한 laevis의 개구리. 그러나 요구하거나 시퀀스 게놈, X.과 유기체에 의해 촉진되는 실험 tropicalis는 훌륭한 대안 시스템입니다.
우리는이 보고서에서 설명한하는 방법은 미세 소관의 중합을 유도하는 미세 소관 - 안정화 에이전트로 taxol을 사용합니다. 탁솔은 미세 소관 정화의 대규모 분리를 용이하게하는 강력한 미세 소관 - 안정화 에이전트이기 때문에 우리는이 방법을 선택했다. 메소드 회우리가 기술에서 가능성이 다른 미세 소관의 중합 방법을 사용하여 미세 소관과 관련된 단백질과 RNA를 비교하여 개선 할 수 있습니다. 대안 GTP-유도 중합 (고전 기술), 22 또는 염색질 기반의 스핀들 어셈블리 (23)에 의해 유도 된 미세 소관을 모방하는 미세 소관 polymerizer으로 실행-GTP를 사용하여 사용하여 중합을 포함 할 수 있습니다. 마지막으로, 미세 소관의 중합 가까운 유사 분열 (중심체, 염색질 및 동원체 중재) 동안 유핵되는 미세 소관의 형태를 모방 될 것이다 유도하는 정자의 핵을 정화 중 하나를 사용하십시오. 미세 소관 핵 이러한 대안 소스에 대한 단점은 핵 에이전트가 탁솔처럼 쉽게 사용할 수 없다는이며, 그들은 핵을 다할 것이다 또는 효율적으로 택솔 (taxol) 등의 미세 소관을 안정화. 따라서 이러한 각각의 방법은 대규모 않는 정화를 위해 사용하는 것이 더 어려울 것입니다. 미세 소관 nucleators의 여러 다른 유형을 비교의 장점그것은 단백질 및 / 또는 미세 소관 핵의 각 경로에 특정 RNA를 식별 할 수 있다는 것입니다.
우리가 여기에서 기술 한 방법은 양서류의 세포질 추출물을 활용합니다. 그러나이 방법은 다른 생물에서 추출 시스템의 사용을 확장 할 수 있습니다. 유사 분열 추출물 충실 소관 어셈블리의 여러 측면을 요점을 되풀이하는 동기화 된 인체 조직 배양 세포 24에서 설명하고있다. 우리는 성공적으로 헬라 세포 5에서 미세 소관 관련 RNA를 식별하려면 다음 추출물을 사용했다. 미세 소관 관련 RNA를가 조사되지 않았지만 비슷한 소관 정화 방식은 여러 가지 생물 25,26에 대해 설명하고있다. 여기에 설명 된 접근 방식은 미세 소관을 핵 능력 집중 세포질 추출물을 생산할 수있는 유기체로 사용할 수 있습니다.
마지막으로, 비록 그 우리 드 방식여기에 서기는 미세 소관과 관련 단백질과 RNA를의 정화에 대해 설명합니다,이 방법은 다른 세포 내 구조에 일반화 될 수있다. 대부분의 지역화의 mRNA는 생화학 적 방법을 사용하여 식별되지 않은 반면 DNA와 RNA 시퀀싱 기술의 최근 발전이 방법에게 지역화 된 RNA를 식별 할 수있는 매력적인 방법을 확인하십시오. 이 방법에 대한 관심의 세포 내 또는 하위 배아 구조는 절연 또는 정제 할 수있다. 그런 다음, 연관된 단백질과 RNA를이 게놈 넓은 규모로 식별 할 수 있습니다. RNA를 그런 다음 전체 셀 또는 농축 지역화 된 RNA를 식별하는 배아의 RNA의 내용을 비교할 수 있습니다. 이 방법은 전체 (Xenopus의 27 번째 지역화 된 RNA를을 식별 방식과 유사한 동물과 식물의 분리), 계란, 말라 연관된 RNA를, ER-관련 RNA를, 미토콘드리아 관련 RNA를 함께 또는 할 수있는 어떤 세포 내 구조에 사용될 수 있습니다 그대로 연결된 RNA를 함께 정화. 에 따라미세 소관 관련 RNA에 대한 우리의 작업은 우리가이 지정된 위치에서 새로운 단백질이 기능을 발견 할 수있는 좋은 방법이 될 것이라고 예측하고있다. 또한, 모든 지역화 된 RNA를의 위치와 범위의 식별 세포와 배아 유전자 발현을 제어하는 mRNA의 현지화를 사용하는 방법에 대한 통찰력을 제공 할 것입니다.
Disclosures
관심 없음 충돌 선언하지 않습니다.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D. Functional analysis of the microtubule-interacting transcriptome. Mol. Biol Cell. 22, 4312-4323 (2011).
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al. Xenopus meiotic microtubule-associated interactome. PLoS One. 5, e9248 (2010).
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}