

Summary
Nous décrivons la collecte de non fécondé
Abstract
De nombreux organismes localiser les ARNm vers des destinations sub-cellulaires spécifiques à l'expression du gène de contrôle spatial et temporel. Des études récentes ont démontré que la majorité du transcriptome est localisé à une position non aléatoire dans les cellules et les embryons. Une approche pour identifier les ARNm localisés est de purifier biochimiquement une structure cellulaire d'intérêt et d'identifier tous les comptes rendus associés. Utilisation des technologies de séquençage à haut débit récemment développés, il est maintenant facile d'identifier tous les ARN associés à une structure subcellulaire. Pour faciliter l'identification du compte rendu, il est nécessaire de travailler avec un organisme dont le génome est entièrement séquencé. Un système attrayant pour la purification biochimique des structures sub-cellulaires sont extraits d'œufs produites à partir de la grenouille Xenopus laevis. Cependant, X. laevis ne dispose pas actuellement d'un génome entièrement séquencé, ce qui entrave l'identification de transcription. Dans cet article, nous décrivons une méthodepour produire des extraits d'œufs de grenouille connexe, X. tropicalis, qui possède un génome entièrement séquencé. Nous fournissons des détails pour la polymérisation des microtubules, la purification et l'isolement de la transcription. Bien que cet article décrit une méthode spécifique pour l'identification des transcriptions associées aux microtubules, nous croyons qu'il sera facilement appliquée à d'autres structures sous-cellulaires et fournira une méthode puissante pour l'identification des ARN localisées.
Introduction
Contrôle spatial et temporel de l'expression des gènes est important pour toutes les cellules, et est particulièrement important pour le contrôle des embryonnaire précoce crépitement 1. Contrôle spatial de l'expression du gène est obtenue par la localisation des ARNm actif vers des destinations spécifiques dans les cellules ou embryons. Dans de nombreux types de cellules très grandes (par exemple, les ovocytes, les embryons et les neurones) localisation de l'ARNm est utilisé pour restreindre l'expression de protéines pour le site d'action de la protéine codée. Depuis un ARNm localisée peut catalyser de nombreux cycles de production de protéines, il est plus efficace de localiser un ARNm que de localiser les molécules de protéines individuelles. ARNm localisés sont généralement translation réprimés jusqu'à ce qu'ils atteignent leur destination, ce qui permet de limiter davantage la localisation de la protéine codée 2. En plus des nombreux cas bien documentés de localisation d'ARN pour contrôler structuration embryonnaire, plusieurs études ont documenté les ARNm qui sont localiséespour le site d'action de la protéine codée. Les principaux exemples incluent la localisation de la β-actine 3 et Arp2 / 3 4 ARNm à la fine pointe de fibroblastes mobiles et de localisation des ARNm de nombreux régulateurs de la mitose à la méiose et fuseau mitotique 5-7.
La plupart des exemples classiques d'ARNm localisés ont été identifiés à travers les écrans génétiques pour les mutations à effet de maternelle et ont ensuite été déterminés pour encoder ARN localisées. Cependant, les récentes études du génome entier ont commencé à fournir un aperçu plus large dans le champ d'ARN localisées. Une récente dans l'écran d'hybridation in situ dans des embryons de drosophile démontré que ~ 70% de tous les ARNm ont une localisation spécifique, y compris de nombreuses nouvelles destinations 8. Purification des pseudopodes à partir de fibroblastes de souris a identifié un groupe diversifié d'ARNm localisé 9. Les travaux de notre groupe à l'aide purification biochimique des microtubules de la méiose Xenopus oeuf extrait identifiés centaines d'ARNm qui copurify avec la broche 5,7. Notre travail a montré que la majorité des ARNm microtubules localisées codent pour des protéines qui fonctionnent dans le contrôle de la mitose, soutenant l'idée que les ARNm sont localisés au site d'action de la protéine codée. En outre, la capacité de détecter l'enrichissement de l'ARNm en une fraction subcellulaire par purification biochimique met en évidence la puissance de cette approche pour l'identification des ARNm localisés.
ARN plus localisées utilisent le transport actif sur le cytosquelette, soit actine ou microtubules, pour parvenir à un transport vers leur destination finale 10. Pour mieux comprendre l'ampleur et les types d'ARN qui sont localisées à des destinations spécifiques en utilisant une approche biochimique, il est nécessaire d'avoir un système in vitro qui peut récapituler les processus du cytosquelette. Un des systèmes de premier ordre pour l'étude de la biologie du cytosquelette est extraits d'œufs produitesà partir œufs non fécondés de la grenouille Xenopus laevis. X. extraits œufs laevis ont été utilisés depuis des décennies pour étudier un large éventail de processus cytosquelette et ont beaucoup contribué à notre compréhension des mécanismes et des molécules qui contrôlent l'assemblage du cytosquelette et de la dynamique 11. En outre, X. extraits œufs laevis se prêtent à des purifications à grande échelle des microtubules et des protéines associées 12,13 et il existe des méthodes bien conçues pour la production de divers types d'extraits d'œufs 14-16. Cependant, pour des études génomiques il ya plusieurs inconvénients à l'utilisation de X. laevis comme un système modèle.
Pendant des décennies, les grenouilles Xenopus laevis ont été un puissant système pour l'étude de la biologie du développement et de la cellule, en raison de la grande taille de l'ovocyte et du développement externe robuste 17. En outre, le développement de systèmes d'extraction oeuf qui peut récapituler de processe cellulaires dans un tube à essai a fait cette grenouille un modèle expérimental puissant. Cependant, Xenopus laevis a été entravée par l'absence d'une séquence complète du génome, qui a été ralentie par la nature allotétraploïde du contraste genome.In, une espèce voisine, Xenopus tropicalis, a un génome diploïde qui a été séquencé en 2010 18. Bien X. tropicalis n'est pas aussi docile expérimentalement que X. laevis 17 la disponibilité d'un génome séquencé rend un système de modèle attrayant pour effectuer des analyses échelle du génome.
Dans ce rapport, nous décrivons une méthode pour faire la méiose II-, cytostatiques extraits facteur arrêtées (CSF) de X. tropicalis 19. Nous décrivons ensuite une méthode simple pour purifier microtubules et des ARN associés à partir de cet extrait. Les ARN peuvent alors être transformés en bibliothèques prêtent au séquençage utilisant des technologies de séquençage à haut débit développés récemment. Une fois que les bibliothèquessont séquencées ils peuvent être alignés pour le génome de la grenouille à identifier les ARNm spécifiques qui sont enrichis dans l'échantillon par rapport à des microtubules extrait total. Cela donne une puissante méthode pour détecter la localisation des ARNm microtubules ciblée sur une échelle du génome. En plus d'être capable de détecter des ARNm localisés, l'utilisation de séquençage à haut débit et d'un génome séquencé offrent la possibilité de découvrir de nouveaux relevés de notes qui ne sont pas actuellement présents dans les annotations de bases de données publiques.
Protocol
1. Génération de X. tropicalis oeufs
Tous Xenopus tropicalis grenouilles sont classés du NASCO. Nos grenouilles sont logées dans un habitat aquatique système de recirculation d'eau maintenu à 27 ° C. Il ya beaucoup d'options pour les systèmes d'eau pour les soins de X. tropicalis. Quelques bonnes informations générales sur cette espèce de grenouille se trouve sur les sites Web des laboratoires Harland et Grainger ( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). Nos grenouilles sont maintenues en tankwater composé de (0,4 g ciclid Sels Lake, 0,6 g de sel marin, 0,625 g de NaHCO 3 par litre d'eau, pH 7,0) 20. Cette recette se traduit par une conductivité de ~ 1800 uS, qui est une forte salinité pour X. tropicalis. Cependant, nous avons constaté que nos grenouilles se développent dans this environnement et la qualité de l'ovocyte est améliorée. Recettes tankwater alternatives peuvent être trouvées ci-dessus dans les ressources indiquées pour le général X. soins tropicalis.
- Les grenouilles sont injectés avec la gonadotrophine chorionique humaine (hCG) sur trois jours consécutifs pour stimuler la ponte: Premièrement, préparer deux concentrations de solution hCG. Resuspendre 10000 U d'hCG poudre lyophilisée dans 10 ml stérile, déionisée H 2 O pour une concentration finale de 1000 U / ml. Ensuite, diluer une solution de 1,000 ml de hCG U / ml dans 9 ml de H 2 O pour une concentration finale de 100 U / ml. Stocker à la fois des solutions à 4 ° C.
- Au jour 1, préparer 4-6 grenouilles pour la ponte par injection avec l'hCG entre 2:00-15:00. Injecter chaque grenouille dans le sac lymphatique dorsal près du cloaque avec 0,2 ml solution hCG 100 U / ml. Avoir les grenouilles rapide au cours des deux injections ultérieures sera de minimiser la quantité de déchets grenouille présent lors de la ponte, mais est facultative.
- Le jour 2, injecter les mêmes grenouillesavec 0,2 ml solution hCG 100 U / ml entre 2:00-15:00.
- Au jour 3, injecter les mêmes grenouilles avec 0,2 ml solution hCG 1000 U / ml, entre 7:00-10:00. Mettre en place des grenouilles à pondre des œufs: remplir un seau en plastique de 6 pintes avec tankwater fraîche, ajouter les grenouilles et les placer dans l'obscurité à 25 ° C. Après cette injection, la ponte va commencer après 4 h et sera terminée d'ici 7 heures. Grenouilles devraient pondre dans un environnement qui est maintenu à un minimum de 25 ° C.
- Faire extraire solutions et disposer d'un équipement prêt immédiatement avant la collecte des œufs.
20X MMR: 100 mM d'HEPES, pH 7,8, 2 mM EDTA pH 7,8, 2 M de NaCl, 40 mM KCl, 20 mM MgCl2, 40 mM CaCl2. Autoclave et stocker à température ambiante. Préparer 1 L de 1X MMR juste avant d'extraire la préparation.
XB 10X: HEPES 100 mM, pH7.7, MgCl2 10 mM, CaCl2 1 mM, 1 M KCl, 500 mM de saccharose. Autoclave et conserver à 4 ° C. Préparer 1 L de 1X XB just avant d'extraire la préparation. Solution Dejelly: Préparer une solution de cystéine 250 de 3% en ml désionisée H 2 O et de pH de 7,8 à 8,0 avec du NaOH 10 N. Préparer juste avant d'extraire la préparation.
CSF-XB: prenez 200 ml de 1X XB et ajouter 2 ml 0,5 M EGTA pH 7,7 et 200 pi 1 M MgCl 2. Préparer juste avant d'extraire la préparation.
CSF-XB +: prendre 50 ml de CSF-XB et ajouter 50 ul de LPC (10 mg / ml chaque stock de Leupeptin, Pepstatin et chymostatine dans le DMSO). Ajouter 50 ul Cytochalasin D (10 mg / ml dans le DMSO). Préparer juste avant d'extraire la préparation.
Préparer une solution de gélatine de 0,2% en déminéralisée H 2 O, micro-ondes pour dissoudre et stériliser par filtration. Stocker à température ambiante.
Réserver 2 Beckman 2 x ½ tubes d'ultracentrifugation pouces.
Préparer deux tubes ml à centrifuger à fond rond de 15 verre avec 0,5 ml de H 2 O dans chaque pour amortir l'ultracentrifugation tube.
Faire polie au feu Pipettes Pasteur en verre. Enclenchez la fin de fréquence de 5 ¾ pouces pipettes en verre pour exposer une large ouverture, et de les exposer à la flamme pour lisser la nouvelle pointe de pipette exposée.
- Préparer un bécher de 500 ml en verre pour stocker des oeufs en agitant une solution de gélatine à 0,2% environ à revêtir les parois du bécher. Jeter la solution de gélatine à partir de bécher après utilisation.
- Ramassez les œufs dans le seau en plastique utilisé pour la pose hr 6-7 après la troisième injection au jour 3. Si désiré, presser doucement chaque grenouille fois pour obtenir tous les œufs restants. Lavez les œufs une fois avec tankwater frais et transfert à l'bécher de 500 ml de verre revêtu d'une solution de gélatine à 0,2%.
2. Préparation de l'extrait de X. Oeufs tropicalis
Toutes les étapes de préparation d'extrait peuvent être effectués à température ambiante, environ 25 ° C. Au fil des lavages, il est important de garder à l'oeufs submergée par liquide de manière à ce qu'ils restent humides. Exposition à l'air peut causer des oeufs d'échapper à l'arrêt du cycle cellulaire ou lyse.
- Décanter autant tankwater que possible tout en réservant suffisamment de liquide pour garder les œufs humide. Inclinaison le bécher contenant les oeufs sur le côté et ajouter environ 300 ml 1X ROR lentement à la paroi du récipient, de telle sorte que l'agitation physique des oeufs est réduite au minimum. Laissez les oeufs se déposent, puis décanter le surnageant contenant les débris. X. oeufs tropicalis sont filandreuse à cette étape, de sorte que l'enlèvement des oeufs activés se fait après dejellying. Répétez l'opération pour un total de trois lavages 1X ROR.
- Dejelly les oeufs. Décanter autant MMR que possible et ajoutez la moitié de la solution dejelly. Swirl continu pendant environ 5 min. Dissolution manteaux gelée sera visible dans le surnageant après quelques minutes. Décanter et ajouter la solution dejelly restant. Continuer à tourbillon continu jusqu'à ce que les œufs emballent très bien et tout orienter avec leur pôle végétal (le pôleavec un pigment blanc) vers le fond de la boîte. Décanter rapidement off comme solution dejelly autant que possible. Une fois les oeufs sont dejellied ils sont très sensibles aux manipulations mécaniques.
- Ajouter délicatement XB aux œufs. Dans le premier lavage XB, sortez les œufs qui ont échappé CSF arrestation en supprimant, œufs blancs, et pseudocleavage gonflés lysées. Activated X. oeufs tropicalis ont tendance à s'installer dans la partie centrale supérieure, afin d'utiliser une pipette de transfert en plastique pour retirer ces out. Aussi enlever des morceaux de peau et des déchets de grenouille. Lavez les œufs un total de trois fois avec ~ solution 300 ml 1X XB, remuant doucement les oeufs entre les lavages et leur permettant de s'installer sur le fond du bécher. Comme précédemment, décanter autant de chaque solution de lavage que possible tout en gardant les oeufs humide.
- Lavez les œufs deux fois par CSF-XB et décanter.
- Ajouter CSF-XB + aux œufs. L'utilisation d'un incendie poli gélatine traitée pipette Pasteur, œufs à tubes ultra-centrifugation transfert avec CSF-XB +, en prenant soin de ne pas exposer l'eGGS dans l'air. Lieu à l'intérieur des tubes de centrifugation ml en verre 15 avec le coussin d'eau.
- Faites tourner les oeufs dans une centrifugeuse clinique à 200 g pendant 1 min, augmenter la vitesse de 800 xg et de spin pendant 30 sec.
- Utilisez un aspirateur pour éliminer autant que possible tampon à partir d'oeufs. Ils doivent être presque à sec sur le dessus. Déplacer rapidement les oeufs à un Sorvall RC-6 centrifugeuse équipée d'un rotor HB-6 (ou équivalent) et de spin 17.000 g pendant 15 min à 20 ° C.
- Retirer la couche cytoplasmique jaune entre le pigment et couches lipidiques aide d'une aiguille de calibre 18 attaché à une seringue de 1 ml. Percer la paroi du tube et tirer sur le corps de la seringue lentement pour obtenir la couche d'extrait cytoplasmique. Évitez les granules pigmentaires autant que possible.
- Transfert cytoplasme à nouveau tube d'ultracentrifugation. Il est normal que l'extrait d'apparaître légèrement nuageux à cette étape. La place à l'intérieur du tube de centrifugeuse de 15 ml en verre avec le coussin de l'eau. Tournez à nouveau 17.000 g pendant 10 min à 20 ° C. Répétez posteraction avec aiguille de calibre 18.
- Transfert cytoplasme dans un tube à centrifuger de 1,5 ml. Estimer le volume de l'extrait et diluer Cytochalasin D et LPC 1:1000 dans l'extrait. Mélangez bien avec une pointe de 1 ml pipette, pipette de haut en bas plusieurs fois sans introduction de bulles d'air. Un rendement typique d'une colonie de grenouilles en bonne santé est d'environ 300-500 ul d'extrait / grenouille. Afin de préserver l'activité maximale, il est nécessaire de stocker l'extrait et effectuer des manipulations expérimentales à température ambiante (20-25 ° C).
3. Purification Taxol stabilisées microtubules de X. tropicalis Extrait
- Ajouter Taxol à une aliquote de l'extrait de 100-200 à une concentration finale de 10 uM et incuber à température ambiante pendant 30 min. Pour les réactions de contrôle, à traiter un volume équivalent de l'extrait avec le médicament nocodazole microtubules destablilzing (10 pM). Réserve 100 ul d'extrait non traité pour analyse.
- Diluer le traité drogueextraire avec 10 volumes BRB-80 (80 mm tuyaux pH 6,8, 1 mM MgCl2, 1 mM EGTA) + 30% de glycérol. Monter 14 tubes en polypropylène à fond rond ml contenant 10 ml de BRB-80 + 60% coussin de glycérol. L'utilisation d'un large alésage pointe de pipette, la couche de réaction de l'extrait traité par médicament légèrement au-dessus de la BRB-80 + 60% coussin de glycérol. Centrifuger pendant 10 min à 17000 g à 20 ° C dans un Sorvall RC-6 centrifugeuse équipée d'un rotor HB-6 (ou équivalent) et des adaptateurs de tube.
- Aspirer le matériel extrait unsedimented contenant le surnageant et laver deux fois avec l'interface déminéralisée H 2 O. Aspirer le volume du coussin reste lentement, en prenant soin de ne pas déranger le gel culot contenant microtubules, protéines associées aux microtubules et des ARN associées aux microtubules dans l'échantillon traité au Taxol. L'échantillon Nocodazole traité ne contient pas de matière visible. Reprendre le culot dans 1 ml TRIzol et procéder aux instructions du fabricant pour isoler l'ARN. UntrExtrait eated (jusqu'à 100 pi) peut être remis en suspension directement dans 1 ml TRIzol.
- Il existe maintenant des kits disponibles dans le commerce pour la préparation de bibliothèques transcriptome appropriés pour RNA-Seq. Ceux-ci peuvent être achetés à travers http://www.illumina.com/ et http://www.454.com/ .
Representative Results
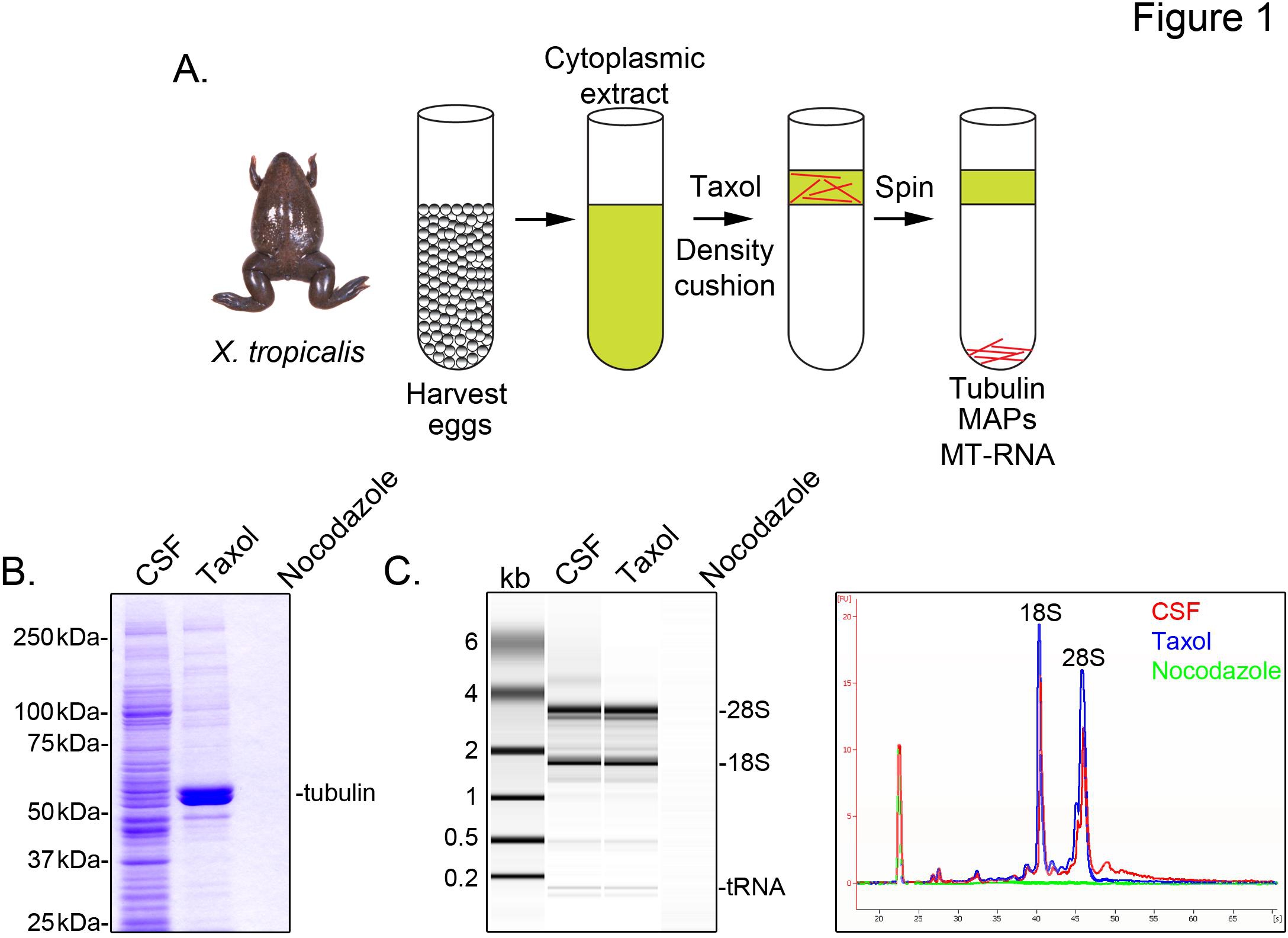
Pour identifier X. transcriptions tropicalis associés aux microtubules, nous préparons un extrait cytosolique à partir d'oeufs non fécondés arrêtés en métaphase de la méiose II (CSF). Le traitement de cet extrait avec du taxol permet la formation de microtubules stables qui peuvent être purifiés par sédimentation à travers un coussin de glycérol (figure 1A). Analyse gel Coomassie confirme que les sédiments α / β-tubuline de manière taxol-dépendante, et représente l'espèce de protéines majeures récupérés dans ces préparations (figure 1B). Les niveaux inférieurs d'autres protéines sont également présentes dans le taxol granulés, mais pas dans les préparations traitées avec le médicament dépolymérisation des microtubules nocodazole, indiquant que les protéines de la fraction taxol spécifiquement associée aux microtubules (MAP).
Un Bioanalyzer est utilisé pour examiner la composition générale ARN dans tous X. tropicalis fractions d'extraits (figure 1C X. extraire laevis oeufs 5,21. Une trace d'axe de la saillie de gel révèle le signal d'ARNm est nettement plus faible dans la microtubules contenant du taxol granulés, et plus particulièrement dans la région de migration au-dessus de l'ARNr 28S, ce qui indique un sous-ensemble des ARNm cosediment avec les microtubules dans X. tropicalis. ARN isolé de cette manière est approprié pour les expériences RNA-Seq utilisant des réactifs disponibles dans le commerce.

Figure 1. Purification de MT-ARN de l'ARN-seq. (A) schéma de purification pour isoler MT-ARN. Les œufs sont récoltés à partir féminin X. trogrenouilles picalis. Après une préparation d'un extrait cytoplasmique, le taxol est ajouté pour induire la polymérisation des microtubules. Les microtubules et MT-ARN sont purifiés par sédimentation à travers un coussin de glycérol. (B) Analyse en gel au bleu de Coomassie des protéines isolées à l'aide du système décrit en (A). Total des CSF extrait par rapport aux protéines sédimentées en présence de taxol ou nocodazole. (C) analyse Bioanalyzer gel de l'ARN isolé en utilisant le schéma décrit dans (A). ARN isolé à partir de CSF comparé à l'ARN extrait sédimenté en présence de taxol ou le nocodazole. Tant la projection de gel et les traces de ligne sont affichés. Reproduit avec la permission de Sharp et al., (2011). Cliquez ici pour agrandir l'image .
Discussion
Dans ce rapport, nous avons décrit une méthode simple pour produire des extraits d'œufs CSF arrêtés de X. tropicalis 19 et utiliser cet extrait d'étudier ARN associées aux microtubules 7. La procédure de base pour la production d'extraits d'œufs CSF arrêtés de X. tropicalis est le même que celui utilisé pour X. laevis avec quelques différences importantes. Un des aspects les plus difficiles à travailler avec X. tropicalis grenouilles est l'obtention d'œufs suffisamment de qualité pour faire un extrait de la nucléation des microtubules ou de l'activité d'assemblage de broche comparable à X. extrait laevis œuf. Pour obtenir des conditions de ponte optimales tout en empêchant le glissement de l'arrêt du cycle cellulaire II méiose, l'intervalle entre les injections d'hormones pour X. tropicalis est plus courte que celle qui est utilisée pour X. laevis, et le moment à partir de la troisième injection de hCG pour le début de la ponte des oeufs est aussi beaucoup plus court. Avec X. laevis le moment de l'injection d'hCG à til début de la ponte est telle qu'elle est pratique et efficace pour les œufs prévues pendant la nuit dans la mémoire tampon. Cependant, en raison du temps plus court entre l'injection d'hCG et la ponte avec X. tropicalis il est souvent nécessaire d'exprimer manuellement les oeufs de grenouilles. Une autre différence importante entre le fait extrait d'œufs de deux grenouilles différents est l'étape dejellying. Avec X. laevis les œufs sont si grands qu'il est facile de déterminer quand la couche gelée s'est dissous en observant de près les oeufs sont espacées dans le bécher. Comme la réaction dejellying commence, les œufs commencent à emballer plus dense. Cependant, X. oeufs tropicalis sont beaucoup plus petits et il peut être assez difficile de déterminer quand la couche gelée est dissous par œuf compacité seul. Nous avons constaté que la méthode la plus fiable pour déterminer quand la couche gelée a dissous est de contrôler l'orientation de l'animal (noir) et végétales pôles (blanc). Quand tout le végétal pôles orient vers le fond du bêcher la couche gelée a été éliminée suffisant pour procéder à l'extraction. Enfin, alors que X. Extrait œuf laevis peut être stocké à des températures fraîches (4-12 ° C), nous avons observé qu'il est essentiel de maintenir X. Extrait d'oeuf tropicalis à la température ambiante (20-25 ° C) pendant la préparation et manipulations expérimentales de préserver l'activité biochimique. En raison des différences dans la facilité d'utilisation, nous préférons utiliser X. laevis grenouilles destinées à la production d'un extrait d'œufs. Cependant, pour des expériences qui nécessitent ou sont facilitées par un organisme dont le génome est séquencé, X. tropicalis est un excellent système alternatif.
La méthode que nous avons décrite dans le présent rapport utilise taxol comme agent de stabilisation de microtubule pour induire la polymérisation des microtubules. Nous avons choisi cette méthode, car le taxol est un agent de stabilisation de microtubule robuste qui facilite l'isolement à grande échelle des microtubules purifiée. Procédé èmeà nous décrivions pourrait probablement être améliorée en comparaison des protéines et des ARN associés aux microtubules en utilisant des méthodes de polymérisation des microtubules alternatives. Alternatives pourraient inclure polymérisation utilisant GTP induite par polymérisation (une technique classique), 22 ou utilisant Ran-GTP comme une polymérisation des microtubules pour imiter les microtubules induite par l'assemblage du fuseau chromatine parcouru 23. Enfin, l'utilisation de noyaux purifiés de sperme pour induire la polymérisation des microtubules serait le plus proche synoptique des types de microtubules qui sont nucléées lors de la mitose (centrosome, chromatine, et kinetochore médiée). Inconvénients de ces sources alternatives de nucléation des microtubules sont que les agents de nucléation ne sont pas aussi facilement disponibles que le taxol et ils ne nucléation ou stabilisent les microtubules de manière aussi efficace que le taxol. Par conséquent, chacune de ces méthodes serait plus difficile à utiliser pour les purifications à grande échelle. L'avantage de comparer plusieurs types de nucléation des microtubulesest qu'il pourrait être possible d'identifier des protéines et / ou d'ARN qui sont spécifiques à chaque voie de la nucléation des microtubules.
La méthode que nous avons décrit ici tire parti des extraits cytoplasmiques d'amphibiens. Cependant, cette approche pourrait être étendue à l'utilisation du système d'extraction à partir d'autres organismes. extraits mitotiques ont été décrits à partir de cellules de culture de tissus humains synchronisés 24 que reproduire fidèlement de nombreux aspects de l'assemblage des microtubules. Nous avons utilisé avec succès ces extraits d'identifier les ARN associées aux microtubules des cellules HeLa 5. Systèmes de purification des microtubules similaires ont été décrits pour différents organismes 25,26, bien que les microtubules ARN associés n'ont pas été examinées. L'approche décrite ici peut être utilisé avec n'importe quel organisme qui peut produire un extrait cytoplasmique concentré capable de nucléation des microtubules.
Enfin, bien que l'approche que nous avons describe ici traite de la purification des microtubules et des protéines et des ARN associés, cette approche pourrait être généralisée à d'autres structures sous-cellulaires. Alors que la plupart des ARNm localisés n'ont pas été identifiés en utilisant des méthodes biochimiques les avancées récentes dans l'ADN et technologies de séquençage ARN font de cette approche une méthode intéressante pour identifier les ARN localisées. Dans cette approche, toute structure subcellulaire ou sous-embryon d'intérêt peut être isolé ou purifié. Ensuite, les protéines et les ARN associés peuvent être identifiés sur une grande échelle du génome. ARN peuvent alors être comparées à la teneur en ARN de la cellule totale ou embryon d'identifier les ARN localisées enrichis. Cette approche pourrait être utilisée avec les oeufs entiers (d'origine animale et la séparation végétale, semblable à l'approche qui a identifié les premiers ARN localisées dans Xenopus 27), l'actine ARN associés, des ARN ER-associés, des ARN mitochondries associées, ou à toute structure subcellulaire qui peut être purifiée avec des ARN associés intactes. Basé surnotre travail sur l'ARN associée aux microtubules nous prévoyons que ce serait un excellent moyen de découvrir de nouvelles protéines qui fonctionnent à un endroit donné. En outre, l'identification de l'emplacement et l'étendue de tous les ARN localisées permettra de mieux comprendre comment les cellules et les embryons utiliser la localisation des ARNm de contrôler l'expression de gènes.
Disclosures
Aucun conflit d'intérêt déclaré.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D. Functional analysis of the microtubule-interacting transcriptome. Mol. Biol Cell. 22, 4312-4323 (2011).
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al. Xenopus meiotic microtubule-associated interactome. PLoS One. 5, e9248 (2010).
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}