

Summary
Мы описываем коллекция неоплодотворенных
Abstract
Многие организмы локализации мРНК субклеточную конкретные направления в пространстве и во времени экспрессию генов контроля. Недавние исследования показали, что большинство транскриптом локализуется на неслучайный положение в клетках и эмбрионах. Один из подходов к выявлению локализованных мРНК является биохимически очищает клеточную структуру интересов и определить все связанные транскриптов. Использование недавно разработанных высокопроизводительных технологий секвенирования Теперь легко определить все связанные с РНК субклеточную структуры. Для облегчения идентификации стенограмму надо работать с организмом с полностью последовательность генома. Одно из привлекательных системы для биохимической очистки субклеточных структур яйца экстракты, полученные из лягушки Xenopus Laevis. Тем не менее, X. Laevis в настоящее время не имеют полностью последовательность генома, что затрудняет идентификацию стенограмму. В этой статье мы опишем методпроизводить яйца выдержки из связанных лягушка, X. tropicalis, что имеет полностью последовательность генома. Мы предлагаем детали для полимеризации микротрубочек, очистки и стенограммы изоляции. Хотя эта статья описывает конкретный метод идентификации микротрубочек связанных стенограммы, мы считаем, что он будет с легкостью применить для других субклеточных структур и даст мощный метод идентификации локализованных РНК.
Introduction
Пространственный и временной контроль экспрессии гена важна для всех клеток, и особенно важно для контроля раннего эмбрионального стучит 1. Пространственный контроль экспрессии гена достигается за счет активного локализации мРНК конкретных направлений внутри клеток или эмбрионов. Во многих очень больших типов клеток (например, ооциты, эмбрионы и нейроны) локализации мРНК используется для ограничения экспрессии белка в месте действия из кодированного белка. С локализованных мРНК может катализировать много раундов производства белка он является более эффективным для локализации мРНК, чем локализовать отдельные молекулы белка. Локализованные мРНК, как правило, поступательно репрессирован, пока они не достигнут места назначения, которое служит для дальнейшего ограничения локализации кодированного белка 2. В дополнение к множество хорошо документированных случаев локализация РНК для контроля эмбрионального паттерна, несколько исследований показали, что мРНК локализованык месту действия кодируемого белка. Известные примеры включают локализации β-актина 3 и Arp2 / 3 4 мРНК к передней кромке подвижных фибробластов и локализации мРНК для многих регуляторов митотический к мейотического и митотического веретена 5-7.
Многие из классических примеров локализованных мРНК были определены в ходе генетических экраны для материнского эффекта мутаций и позже были определены для кодирования локализованных РНК. Однако недавние генома исследования начали предоставлять широкий представление рамки локализованных РНК. Недавно на месте экрана гибридизации в эмбрионах Drosophila показали, что ~ 70% всех мРНК имеют определенную локализацию, в том числе многих направлений романа 8. Очистка псевдоподиями из мышиных фибробластов определены различные группы локализованных мРНК 9. Работы из нашей группы с использованием биохимических очистки микротрубочек от мейотическими Xenoгной яйцо извлекает выявили сотни мРНК, которые copurify со шпинделем 5,7. Наша работа показала, что большинство микротрубочек локализованных мРНК, кодирующих белки, которые функционируют в управление митоза, поддерживая идею, что мРНК локализованы в месте действия из кодированного белка. Кроме того, возможность обнаружения мРНК обогащения в субклеточных фракций биохимические очистки подчеркивается сила этого подхода для идентификации локализованных мРНК.
Большинство локализованных РНК использовать активные транспорта на цитоскелета, либо актина или микротрубочки, для развития транспорта к месту назначения 10. Чтобы получить более полное представление о масштабах и типах РНК, которые локализованы в конкретных направлений использования биохимического подхода необходимо иметь в пробирке системы, которая может повторять цитоскелета процессов. Одной из ведущих систем для изучения цитоскелета биологии яйцо экстракты, полученныеиз неоплодотворенных яиц из лягушки Xenopus Laevis. X. Laevis яйцо экстрактов были использованы на протяжении десятилетий для изучения широкого спектра цитоскелета процессов и внесли большой вклад в наше понимание механизмов и молекул, которые контролируют сборку цитоскелета и динамики 11. Кроме того, X. Laevis яйцо экстракты поддаются крупномасштабных очищения микротрубочек и ассоциированных белков 12,13 и есть хорошо продуманные методы для производства различных видов яйца экстракты 14-16. Тем не менее, для геномные исследования есть несколько недостатков использования X. Laevis в качестве модельной системы.
На протяжении десятилетий лягушки Xenopus Laevis были мощной системой для изучения развития и клеточной биологии, из-за большого размера яйцеклетки и надежные внешние развития 17. Кроме того, развитие систем яйцо экстракта, который может повторять многие клеточные Processeы в пробирке сделал эту лягушку мощная экспериментальная модель. Тем не менее, Xenopus Laevis был затруднен отсутствием полного генома, которая была замедлена аллотетраплоидом природу контраста genome.In, тесно связанных видов, Xenopus tropicalis, имеет диплоидный геном, который был в 2010 году последовательно 18. В то время как X. tropicalis не столь послушной, как экспериментально X. 17 Laevis наличие генома последовательный делает его привлекательным модели системы для выполнения широкого анализа генома.
В этом докладе мы опишем метод, чтобы сделать мейоза II-, цитостатических факторов арестован экстрактов (CSF) из X. tropicalis 19. Затем мы опишем простой метод, чтобы очистить микротрубочек и связанные с РНК из этого экстракта. РНК затем могут быть преобразованы в библиотеки поддаются последовательности использованием недавно разработанного высоких технологий секвенирования пропускной способности. После библиотекиявляются последовательными они могут быть приведены в геноме лягушки определить конкретные мРНК, которые обогащены микротрубочек образца по сравнению с общим экстракта. Это обеспечивает мощный метод для обнаружения микротрубочки целевой локализации мРНК на геном масштабе. Помимо того, что в состоянии обнаружить локализованные мРНК, использование высокой пропускной последовательности генома и последовательное предлагаем возможность обнаружения новых транскриптов, которые не являются в настоящее время в общественном аннотаций к базам данных.
Protocol
1. Генерация X. tropicalis Яйца
Все лягушки Xenopus tropicalis отсортированы от НАСКО. Наши лягушки размещены в водную среду обитания системе циркуляции воды хранятся при 27 ° C. Есть много вариантов для систем водоснабжения для ухода за X. tropicalis. Некоторые хорошие общей информацией по этому видов лягушек можно найти на веб-сайтах Харланд энд Грейнджер Labs ( http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / ). Наши лягушки ведутся в tankwater, состоящей из (0,4 г соли Ciclid озера, 0,6 г морской соли, 0,625 г NaHCO 3 на литр воды, рН 7,0) 20. Этот рецепт приводит к проводимости ~ 1800 мкСм, которая является высокой соленостью для X. tropicalis. Тем не менее, мы обнаружили, что наши лягушки процветают в эту темуОкружающая среда и качества ооцитов улучшается. Альтернативные рецепты tankwater можно найти в выше перечисленных ресурсов для общего X. tropicalis помощи.
- Лягушки вводят хорионического гонадотропина человека (ХГЧ) три дня подряд, чтобы стимулировать яйценоскость: во-первых, подготовить два концентрации ХГЧ решение. Ресуспендируют 10000 ед лиофилизированные ХГЧ порошка в 10 мл стерильной деионизированной H 2 O для конечной концентрации 1000 ед / мл. Затем разбавленный 1 мл 1000 ЕД / мл ХГЧ решение в 9 мл H 2 O для конечной концентрации 100 U / мл. Храните оба решения при 4 ° C.
- В день 1, подготовить 4-6 лягушки Для откладки яиц путем введения с ХГЧ между 2:00-3:00 PM. Введите каждая лягушка в спинной лимфатический мешок рядом с клоакой 0,2 мл 100 ЕД / мл ХГЧ решение. Имея лягушки быстро в течение последующих двух инъекций позволит свести к минимуму количество отходов настоящее лягушки во время откладки яиц, но не является обязательным.
- В день 2, вводят лягушачьи0,2 мл 100 ед / мл ХГЧ решение между 2:00-3:00 PM.
- На 3 день вводить же лягушки с 0,2 мл 1000 ЕД / мл ХГЧ решение, между 7:00-10:00 AM. Настройка лягушки откладывают яйца: заполнить 6-кварт пластиковые ведра со свежим tankwater, добавьте лягушек и место в темноте при 25 ° С. После этой инъекции, яйцекладка начнется через 4 часа и будет завершено к 7-ми часов. Лягушки должны откладывать яйца в среде, которая поддерживается на уровне не менее 25 ° С.
- Сделать извлечь решения и иметь оборудование, готовое непосредственно перед сбором яиц.
20X MMR: 100 мМ HEPES, рН 7,8, 2 мМ ЭДТА, рН 7,8, 2 М NaCl, 40 мМ KCl, 20 мМ MgCl 2, 40 мМ CaCl 2. Автоклав и хранить при комнатной температуре. Подготовить 1 л MMR 1X незадолго до извлечения подготовки.
10X XB: 100 мм HEPES, рН 7,7, 10 мМ MgCl 2, 1 мМ CaCl 2, 1 М KCl, 500 мМ сахарозы. Автоклав и хранят при температуре 4 ° С. Подготовить 1 л 1X XB JУсть до извлечения подготовки. Dejelly решение: Подготовка 250 мл 3% раствора цистеина в деионизированной Н2О и рН до 7,8-8,0 с помощью 10 N NaOH. Подготовка непосредственно перед извлечь подготовки.
CSF-ВР: взять 200 мл 1X XB и добавить 2 мл 0,5 М ЭДТА рН 7,7 и 200 мкл 1 М MgCl 2. Подготовка непосредственно перед извлечь подготовки.
CSF-XB +: возьмите 50 мл CSF-XB и добавить 50 мкл LPC (10 мг / мл каждого запас лейпептина, Пепстатин и химостатина в ДМСО). Добавить 50 мкл Цитохалазин D (10 мг / мл в ДМСО). Подготовка непосредственно перед извлечь подготовки.
Подготовьте 0,2% раствор желатина в деионизованной H 2 O, микроволновую печь, чтобы растворить и Фильтры стерилизовать. Хранить при комнатной температуре.
Резерв 2 Beckman 2 х ½ дюйма труб Ultracentrifuge.
Приготовьте два стекла 15 мл с круглым дном пробирки с 0,5 мл H 2 O в каждой, чтобы смягчитьультрацентрифужную пробирку.
Сделать пожарной полированное стекло пипетки Пастера. Привязать обрываться из 5 ¾ дюйма стеклянных пипеток подвергать широкому открытию, и не подвергайте воздействию огня, чтобы сгладить новые подвергается пипетки.
- Подготовьте 500 мл химический стакан для хранения яиц, вращая 0,2% раствор желатина вокруг, чтобы покрыть стенками сосуда. Откажитесь от раствора желатина стакан после использования.
- Сбор яиц из пластиковое ведро используется для укладки 6-7 ч после третьей инъекции в день 3. При желании, слегка сжать каждую лягушку один раз, чтобы получить оставшиеся яйца. Мытье яйца один раз свежей tankwater и передачи в 500 мл химический стакан покрытых 0,2% раствор желатина.
2. Подготовка выписки из X. Яйца tropicalis
Все стадии подготовки экстракта можно проводить при комнатной температуре примерно 25 ° С. На протяжении моет, важно сохранить яйцос погружения под жидкости, так что они остаются влажными. Воздействие воздух может вызвать яйца избежать ареста клеточного цикла или лизировать.
- Слейте столько tankwater возможно при сохранении достаточно жидкости, чтобы держать яйца влажным. Наклон стакан, содержащий яйца в сторону и добавляют ~ 300 мл 1X MMR медленно к стенке стакана, так что физическое перемешивание яйца сводится к минимуму. Пусть яйца урегулировать, то переливать супернатант, содержащий от мусора. X. tropicalis яйца волокнистой на этом этапе, так что удаление активированных яиц выполняется после dejellying. Повторите эти действия для в общей сложности три 1X моет MMR.
- Dejelly яйца. Декантируйте прочь так, MMR, как это возможно, и добавить половину раствора dejelly. Вихревой непрерывно в течение приблизительно 5 мин. Растворение желе пальто будут видны в супернатант через пару минут. Декантируйте прочь и добавить оставшийся раствор dejelly. Продолжают циркулировать непрерывно, пока яйца не упаковать очень плотно и все Востока с их вегетативный полюс (полюсс белым пигментом) в сторону дна чашки. Быстро сцеживать прочь так dejelly решение, как это возможно. Как только яйца dejellied они очень чувствительны к механическим манипуляций.
- Аккуратно добавьте XB к яйцам. В первой стирки XB, удалить яйца, которые избежали ареста CSF путем удаления лизированными, опухшие, белый и pseudocleavage яиц. Активированный X. tropicalis яйца обычно селятся в самом центре, так что используйте пластиковые пипетки передачи тянуть из этих. Также удалите кусочки кожи лягушки и отходов. Мойте яйца в общей сложности три раза с ~ 300 мл раствора 1X XB, аккуратно закрученных между яйцами моет и позволяя им оседать на дне стакана. Как и прежде, сцедить столько каждого промывочного раствора возможно, сохраняя при этом яйца влажным.
- Мойте яйца дважды CSF-XB и переливать.
- Добавить CSF-XB + на яйца. Используя желатин обработанные огневой полировкой пипетки Пастера перенести яйца к ультра-центрифужные пробирки с CSF-XB +, заботясь, чтобы не подвергать электроннойGGS с воздухом. Место внутри трубы 15 мл стеклянный центрифуги с водой подушку.
- Спин яйца в клинической центрифуге при 200 мкг в течение 1 мин, увеличить скорость до 800 мкг и спина в течение 30 сек.
- Используйте аспиратором удалить как можно больше буфера как можно дальше от яиц. Они должны быть почти сухой на вершине. Быстрое перемещение яйца Sorvall RC-6 Центрифуга оснащена HB-6 ротора (или эквивалент) и спина 17 000 мкг в течение 15 мин при 20 ° C.
- Удалите желтую цитоплазматической слой между пигментом и липидного слоя, используя иглу 18 калибра прикреплен к 1 мл шприца. Прокол стороне трубки и тянуть цилиндра шприца медленно получить цитоплазматической слой экстракта. Избежать пигментных гранул в максимально возможной степени.
- Передача цитоплазмы в новую пробирку ультрацентрифугирование. Это нормально для экстракта появляться слегка мутным на этот шаг. Место внутри 15 мл стеклянную пробирку центрифуги водой подушку. Спин снова 17000 мкг в течение 10 мин при 20 ° C. Повторите добraction с иглой 18 калибра.
- Передача цитоплазмы в 1,5 мл трубки микроцентрифужную. Оцените объем экстракта и разбавляют Цитохалазин D и LPC 1:1000 в экстракт. Хорошо перемешать с 1 мл пипетки, пипетки вверх и вниз много раз без введения пузырьков воздуха. Типичный выход из здоровой колонии лягушки составляет примерно 300-500 мкл экстракта / лягушки. Чтобы сохранить максимальную активность, необходимо хранить экстракта и выполнения экспериментальных манипуляций при комнатной температуре (20-25 ° С).
3. Очистка Таксол стабилизированного Микротрубочки из X. Экстракт tropicalis
- Добавить таксол до 100-200 мкл аликвоты экстракта при конечной концентрации 10 мкМ и инкубируют при комнатной температуре в течение 30 мин. Для контрольных реакций лечения эквивалентный объем экстракта микротрубочки destablilzing нокодазол наркотиков (10 мкм). Резервный 100 мкл необработанного экстракта для анализа.
- Развести наркотиков лечениеизвлечение с 10 объемами BRB-80 (80 мМ труб рН 6,8, 1 мМ MgCl 2, 1 мМ EGTA) + 30% глицерина. Сборка 14 мл с круглым дном полипропиленовые пробирки, содержащие 10 мл BRB-80 + 60% глицерина подушки. Использование широким отверстием наконечника пипетки, слой, обработанных лекарством экстракт реакцию осторожно сверху BRB-80 + 60% глицерина подушки. Центрифугируют 10 мин при 17000 х г при 20 ° С в центрифуге Sorvall RC-6 центрифуга оснащена HB-6 ротора (или эквивалент) и трубка адаптеров.
- Аспирируйте супернатант, содержащий экстракт unsedimented материала, и мыть интерфейса дважды деионизованной H 2 O. Аспирируйте оставшийся объем подушки медленно, стараясь не беспокоить гелеобразный осадок, содержащий микротрубочки, микротрубочек связанных белков и микротрубочек связанных РНК в таксолом образца. Нокодазолом обработанный образец не содержит видимых материала. Ресуспендируют гранул в 1 мл TRIzol и приступить к инструкции производителя для выделения РНК. UNTReated экстракт (до 100 мкл) может быть взвесь непосредственно в 1 мл TRIzol.
- Есть в настоящее время коммерчески доступных наборов для приготовления транскриптома библиотек, пригодных для РНК-след. Они могут быть приобретены через http://www.illumina.com/ и http://www.454.com/ .
Representative Results
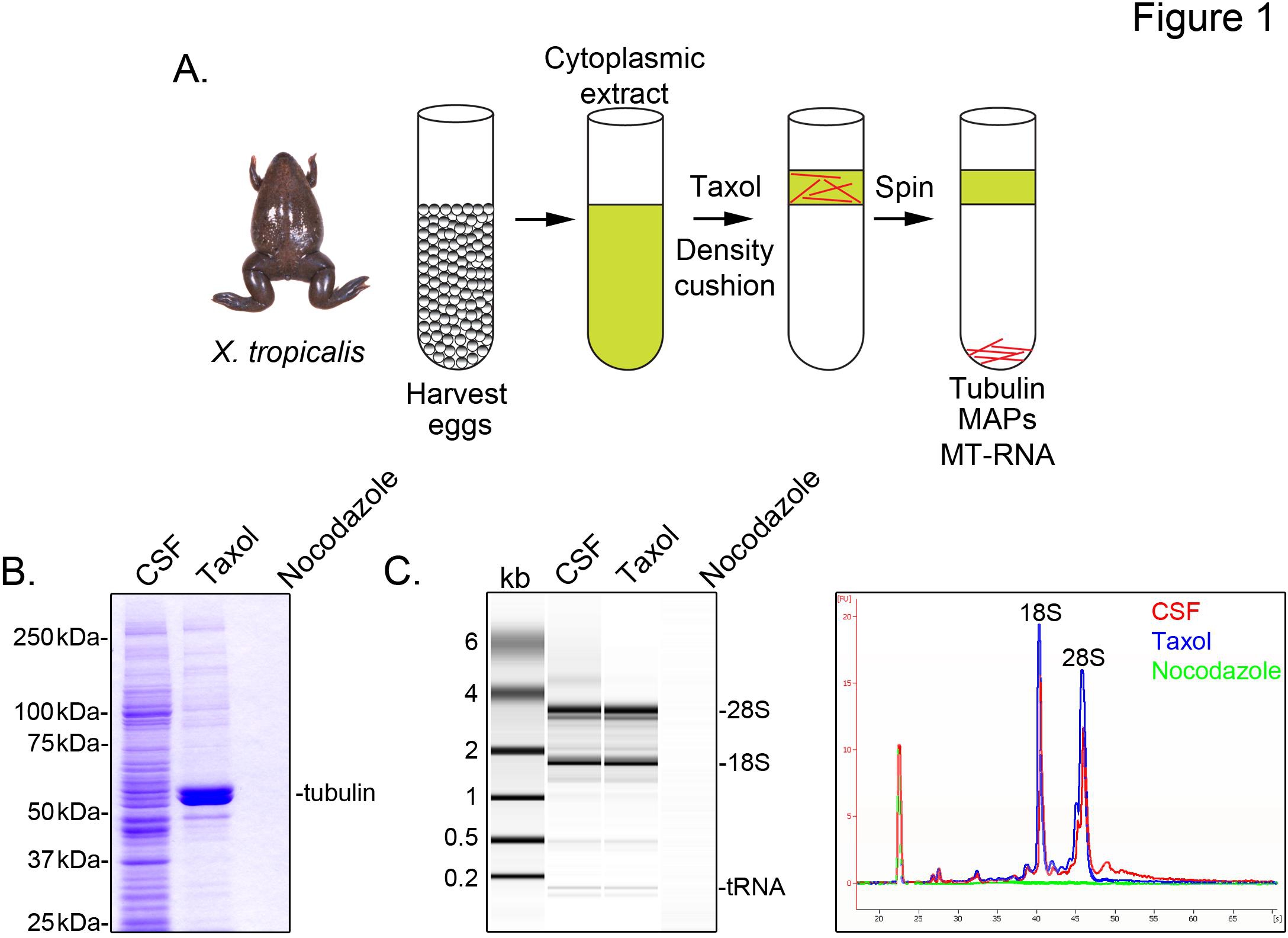
Чтобы определить X. tropicalis транскриптов, связанные с микротрубочки, мы готовим цитозольные выписку из неоплодотворенных яиц арестован в метафазе мейоза II (CSF). Лечение этого экстракта с таксолом позволяет формировать стабильный микротрубочки, которые можно выдел ли путем осаждени с помощью глицерина подушка (Фиг.1А). Coomassie геля анализ подтверждает, что α / β-тубулина отложений в таксол-зависимым образом, и представляет основные виды белка восстановлены в этих препаратах (рис. 1В). Более низкие уровни других белков, которые также присутствуют в таксол гранул, но не в препаратах, обработанных с деполимеризации микротрубочек препарата нокодазолом, указывая, что белков в таксол фракции конкретно ассоциированный с микротрубочками (MAP),.
Agilent Bioanalyzer используется, чтобы исследовать общую РНК состав во всех X. tropicalis экстракта фракций (рис. 1в X. Laevis яйцо извлечения 5,21. Линии следов геля проекция показывает мРНК сигнала, заметно ниже на микротрубочки содержащих таксол гранул, в первую очередь в области миграции выше 28S рРНК, указывая, что подмножество cosediment мРНК с микротрубочек в X. tropicalis. РНК, выделенных таким образом подходит для РНК-последующие эксперименты с использованием коммерчески доступных реагентов.

Рисунок 1. Очистка MT-РНК РНК-сл. (А) очистки схема изолировать MT-РНК. Яйца собирают из женщин X. троpicalis лягушек. После подготовки цитоплазматический экстракт, таксол добавляется к индукции полимеризации микротрубочек. Микротрубочки и МТ-РНК выдел ли путем осаждени с помощью глицерина подушке. (B) кумасси гель анализа белков, выделенных с использованием схемы, описанные в (А). Всего CSF экстракта по сравнению с белками осаждают в присутствии таксола или нокодазолом. (C) Bioanalyzer анализ в геле РНК выделяли с использованием схемы, описанной в (А). РНК, изолированной от CSF извлечения по сравнению с РНК осаждали в присутствии таксола или нокодазолом. Оба выступа гель и линией следы показаны. Печатается с разрешения от Sharp и соавт., (2011). Кликните здесь, чтобы посмотреть увеличенное изображение .
Discussion
В данном отчете мы описали простой способ получения CSF-арестованных экстрактах яиц из X. tropicalis 19 и использовать этот экстракт для изучения микротрубочек связанных РНК 7. Основная процедура получения CSF-арестованных яйцо выдержки из X. tropicalis такой же, как используемый для X. Laevis с несколько ключевых отличий. Одним из наиболее сложных аспектов работы с X. tropicalis лягушек получения достаточно высокого качества яиц, чтобы сделать выписку с зарождения микротрубочек или деятельности сборки веретена сопоставимы с X. Laevis яйца извлекается. Для достижения оптимального яйценоскость условиях, предотвращая проскальзывание мейоза II клеточного цикла, интервал между инъекциями гормонов для X. tropicalis короче, чем для X. Хепориз, и время от третьего ХГЧ инъекции до начала откладки яиц также намного короче. С X. Laevis сроки от ХГЧ инъекции Tон начала откладки яиц такова, что она является удобной и эффективной для яиц, которые будут установлены на ночь в буфере. Тем не менее, из-за более короткого времени между инъекции чХГ и откладки яиц с X. tropicalis часто приходится вручную выразить яйца от лягушек. Еще одно существенное отличие между созданием яйца выписку из двух разных лягушек dejellying шаг. С X. Laevis яйца настолько велики, что легко определить, когда слой желе растворится, наблюдая, как тесно яйца расположены в стакан. Как dejellying начинается реакция, яйца начинают упаковать плотнее. Однако X. tropicalis яйца намного меньше и это может быть довольно трудно определить, когда слой желе распущена яйцо плотность упаковки в одиночку. Мы обнаружили, что наиболее надежным методом для определения, когда слой желе растворится является контроль ориентации животных (черный) и растительные (белый) полюсов. Когда все растительные полюсов Oriных в нижней части стакана слой желе был удален достаточно, чтобы приступить к экстракта. Наконец, в то время как X. Laevis яйцо экстракт можно хранить при низкой температуре (4-12 ° C), мы обнаружили, что очень важно для поддержания X. tropicalis яйцо экстракта при комнатной температуре (20-25 ° С) в период подготовки и экспериментальных манипуляций сохранить биохимической активности. Из-за различий в простоте использования мы предпочитаем использовать X. Хепориз лягушки для производства яичного экстракта. Тем не менее, для экспериментов, которые требуют или облегчаются организм с геномом, последовательность, X. tropicalis является отличной альтернативной системы.
Метод, который мы описали в этом докладе используется таксол как микротрубочки стабилизирующего агента, чтобы вызвать полимеризацию микротрубочек. Мы выбрали этот метод, поскольку таксол является надежной микротрубочки стабилизирующего агента, который облегчает крупномасштабного выделения очищенной микротрубочек. Метод йна мы описали, вероятно, может быть улучшена путем сравнения белков и РНК, связанный с микротрубочками альтернативными методами полимеризации микротрубочек. Альтернативы могут включать полимеризации с использованием ГТФ-индуцированной полимеризации (классический метод), 22 или с помощью Ran-GTP как микротрубочки полимеризатора имитировать микротрубочек индуцированных хроматина управляемой сборки веретена 23. Наконец, использование очищенной спермы ядер, чтобы вызвать полимеризацию микротрубочек будет имитировать ближайшим к типам микротрубочек, которые зарождаются в митозе (центросомы, хроматин, и кинетохор опосредованного). Недостатки этих альтернативных источников нуклеации микротрубочек в том, что зародыши кристаллизации не так легко доступны как таксол, и они не пузырьковом или стабилизации микротрубочек так же эффективно, как таксол. Таким образом, каждый из этих методов будет более трудно использовать для крупномасштабного очисток. Преимущество сравнении нескольких различных типов микротрубочек нуклеаторамиявляется то, что он может быть возможно идентифицировать белки и / или РНК, которые являются специфическими для каждого пути нуклеации микротрубочек.
Метод, который мы описали здесь имеет преимущество цитоплазматических экстрактов амфибий. Однако этот подход может быть распространен на использование экстракта системы от других организмов. Митотическая экстракты были описаны из синхронных человеческой клетки культур тканей 24, которые добросовестно повторять многие аспекты сборки микротрубочек. Мы успешно использовали эти экстракты определить микротрубочек связанных РНК из клеток HeLa 5. Подобные схемы очистки микротрубочек были описаны для различных организмов, 25,26, хотя ассоциированный с микротрубочками РНК, которые не были рассмотрены. Описанный здесь подход может быть использован с любой организм, который можно получить концентрированный экстракт цитоплазматические способны зародышей микротрубочек.
Наконец, хотя подход, что мы де-писец здесь обсуждаются очистки микротрубочек и ассоциированных белков и РНК, этот подход может быть обобщен на других субклеточных структур. Хотя большинство локализованных мРНК не были идентифицированы с помощью биохимических методов последние достижения в ДНК и РНК технологии секвенирования делают этот подход привлекательным методом для выявления локализованных РНК. При таком подходе любой субклеточных или суб-эмбрион структуры процентных могут быть выделены или очищены. Тогда ассоциированные белки и РНК могут быть идентифицированы на геном масштабе. РНК затем можно сравнить с содержанием РНК от общей клетки или эмбрион идентифицировать обогащенный локализованных РНК. Такой подход может быть использован с целых яиц (животного и растительного разделения, аналогичный подход, который идентифицировали первую РНК, локализованных в Xenopus 27), актин связанного РНК, ER-связанной РНК, митохондрий, связанных РНК или любой субклеточные структуры, которая может быть очищен с соответствующими РНК нетронутыми. На основенаша работа по микротрубочек связанных РНК мы прогнозируем, что это будет отличный способ открыть для себя новые белки, которые функционируют на данном месте. Кроме того, определение локализации и распространенности всех локализованных РНК обеспечит понимание, как клетки и эмбрионы использовать локализации мРНК для контроля экспрессии генов.
Disclosures
Нет конфликта интересов объявлены.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D. Functional analysis of the microtubule-interacting transcriptome. Mol. Biol Cell. 22, 4312-4323 (2011).
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al. Xenopus meiotic microtubule-associated interactome. PLoS One. 5, e9248 (2010).
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}