

Summary
Biz döllenmemiş toplanması tarif
Abstract
Birçok organizma mekansal ve zamansal kontrolü gen özgü hücre içi hedeflere mRNA lokalize. Son çalışmalar transcriptome çoğunluğu hücreleri ve embriyolarda bir tesadüfi olmayan konuma lokalize olduğunu göstermiştir. Lokalize mRNA belirlemek için bir yaklaşım biyokimyasal ilgi bir hücresel yapı arındırmak için ve tüm ilişkili transkript belirlemektir. Yakın zamanda geliştirilen yüksek verimli sıralama teknolojilerini kullanarak artık bir hücre içi yapı ile ilgili tüm RNA'lar tanımlamak için basittir. Transkriptlerini tanımlamayı kolaylaştırmak için, bir tam sıralı bir organizma genomu ile birlikte çalışmak için gereklidir. Hücre içi yapıların biyokimyasal arıtma için bir çekici sistemi kurbağa Xenopus laevis üretilen yumurta alıntılar vardır. Ancak, X laevis şu anda transkript kimlik engellemektedir tam sıralı genom, yoktur. Bu yazıda bir yöntem tarifilgili kurbağa, X gelen yumurta özler üretmek için Bir tam sıralı bir genomu olan tropicalis. Biz mikrotübül polimerizasyon, arıtma ve transkript izolasyonu için ayrıntıları. Bu makalede mikrotübüle bağlı transkript tanımlanması için özel bir yöntem tarif ederken, bunun kolayca diğer hücre içi yapılarına uygulanabilir ve lokalize RNA'lar belirlenmesi için güçlü bir yöntem sağlar inanıyoruz.
Introduction
Gen ifadesinin mekansal ve zamansal kontrolünün tüm hücreler için önemlidir ve 1 dizilmesine erken evredeki embriyonik bölgesinin kontrolü için özellikle önemlidir. Gen ifadesinin mekansal kontrol hücreleri ya da embriyolar içinde belirli bir noktaya mRNA'ların aktif lokalizasyonu ile elde edilir. Çok sayıda çok büyük hücre tipleri (örneğin, oositler, embriyolar ve nöronlar) mRNA lokalizasyonu kodlanmış protein eylem sözlük protein ekspresyonu sınırlamak için kullanılır. Yerelleştirilmiş bir mRNA protein üretimi birçok tur katalize çünkü bireysel protein molekülleri tespit etmek için daha bir mRNA tespit etmek için daha etkilidir. Onlar daha kodlu protein 2 lokalizasyonu neden olmaktadır hedeflerine, ulaşana kadar lokalize mRNA genellikle translationally baskı vardır. Embriyonik desenlendirme kontrol etmek için RNA lokalizasyonu çok iyi belgelenmiş durumda ek olarak, çeşitli çalışmalar lokalize mRNA belgeledikodlanmış protein eylem sitesine. Tanınmış örnekler hareketli fibroblast öncü için β-aktin 3 ve Arp2 / 3 4 mRNA lokalizasyonu ve mayoz ve mitoz iğ 5-7 birçok mitoz düzenleyiciler için mRNA lokalizasyonu içerir.
Lokalize mRNA'ların klasik örneklerin çoğu anne etkisi mutasyonlar için genetik ekranlarında belirlendi ve daha sonra lokalize RNA'lar kodlamak için belirlenmiştir. Ancak, son genom çalışmaları lokalize RNA'lar kapsamına geniş bilgi sağlar başladı. Drosophila embriyolar in situ hibridizasyon ekranında yeni bir tüm mRNA'ların ~ 70% birçok yeni destinasyonlarda 8 dahil olmak üzere belirli bir yeri, sahip olduğunu göstermiştir. Fare fibroblastlardan pseudopodia saflaştırılması lokalize mRNA 9 çeşitli bir grup belirledi. Mayoz Xeno gelen mikrotübül biyokimyasal arıtma kullanarak gruptan çalışmairin yumurta mil 5,7 ile copurify mRNA'ların tespit yüzlerce ayıklar. Çalışmalarımız mRNA'lar, kodlanmış protein eylem sitesine lokalize olduğu fikrini desteklemektedir, mikrotübül-lokalize mRNA'ların çoğunluğu mitoz kontrol işlevi proteinleri kodlayan gösterdi. Ayrıca, biyokimyasal arıtma tarafından bir hücre içi kısmını mRNA zenginleştirme tespit yeteneği lokalize mRNA belirlenmesi için bu yaklaşımın gücünü vurgulamaktadır.
En lokalize RNA'lar nihai hedef 10 ulaşım ulaşmak için, aktin veya mikrotübül ya, hücre iskeleti aktif taşıma kullanın. Bu iskelet işlemleri özetlemek olduğunu in vitro sistemde bir olması için gerekli olan biyokimyasal bir yaklaşım kullanarak belirli hedeflere lokalize RNA'lar ölçüde ve türleri daha iyi anlamak için. Sitoskeletal biyoloji eğitimi için önde gelen sistemlerinden biri üretilen yumurta özleri olankurbağa Xenopus laevis. X den döllenmemiş yumurta laevis yumurta özleri iskelet süreçleri geniş bir dizi çalışma için yıllardır kullanılmaktadır ve iskelet montaj ve dinamikleri 11 kontrol mekanizmaları ve moleküllerin anlayışımızı çok katkıda bulunmuştur. Ayrıca, X. laevis yumurta özü ve mikrotübüllerin 12,13 ilişkili proteinlerin büyük ölçekli saflaştırılması için uygun olan ve yumurta özü 14-16 çeşitli üretimi için iyi tasarlanmış bir yöntem vardır. Bununla birlikte, genomik çalışmaları için X'in kullanımı için çeşitli dezavantajları vardır bir model sistem olarak laevis.
On yıllardır Xenopus laevis kurbağa büyük oosit büyüklüğü ve sağlam dış gelişme 17 nedeniyle gelişim ve hücre biyolojisi, çalışma için güçlü bir sistem olmuştur. Ayrıca, birçok hücresel processe özetlemek olabilir yumurta özü sistemlerinin geliştirilmesibir test tüpünde s bu kurbağa güçlü bir deneysel model yaptı. Ancak, Xenopus laevis genome.In kontrast allotetraploid doğanın yavaşladı olan bir tam genom dizisi, eksikliği engel olmuştur, bir yakından ilişkili türler, Xenopus tropicalis 2010 18 sıralı bir diploid genom vardır. Iken X tropicalis X gibi deneysel olarak izlenebilir değildir laevis 17 bir sıralı genomunun kullanılabilirliği genom geniş analizleri yapmak için çekici bir model sistem yapar.
Bu yazıda X den mayoz II-, sitostatik faktör-tutuklandı özler (BOS) yapmak için bir yöntem tarif tropicalis 19. Biz o zaman bu özü mikrotübüllerin ve ilgili RNA'lar arındırmak için basit bir yöntem tarif. RNA'lar sonra yeni geliştirilmiş yüksek verimlilik sıralaması teknolojileri kullanarak sıralama için uygun kütüphaneler dönüştürülebilir. Kütüphaneler kezbunlar toplam özü göre mikrotübül örnek zenginleşmiş spesifik mRNA tespit etmek kurbağa genomuna hizalanabilir sıralanır. Bu bir genom ölçekte mikrotübül hedefli mRNA lokalizasyon tespit etmek için güçlü bir yöntem sağlar. Lokalize mRNA tespit edebilmek olmasının yanı sıra, yüksek verimli sıralama ve sıralı genomunun kullanımı kamu veritabanı açıklamalar şu anda mevcut olmayan yeni transkript keşfetme imkanı sunuyoruz.
Protocol
1. X tropicalis Yumurta Üretimi
Tüm Xenopus tropicalis kurbağa NASCO gelen sıralanmıştır. Bizim kurbağa devir daim su sistemi 27 tutulur bir Sucul Habitatlar yerleştirilmiştir ° C X'in bakımı için su sistemleri için birçok seçenek vardır tropicalis. Bu kurbağa türü ile ilgili bazı iyi genel bilgiler (Harland ve Grainger laboratuarlarının web sitelerinde bulunabilir http://tropicalis.berkeley.edu/home/ , http://www.faculty.virginia.edu/xtropicalis / .) Bizim kurbağa (0.4 gr Ciclid Lake tuzları, 0.6 g deniz tuzu, su, pH 7.0 litresi başına 0.625 g NaHCO3), 20 oluşan tankwater tutulur. X için bir yüksek tuzluluk olan ~ 1.800 ms bir iletkenlik, bu tarifi sonuçları tropicalis. Ancak, bizim kurbağa thi gelişirler bulduks çevre ve oosit kalitesi artırıldı. Alternatif tankwater tarifleri genel X için listelenen kaynaklara üzerinde bulunabilir tropicalis bakım.
- Kurbağalar döşeme yumurta uyarmak için üç ardışık günlerde insan koryonik gonadotropin (hCG) ile enjekte edilir: Birincisi, hCG çözümün iki konsantrasyonları hazırlamak. 1000 U / ml 'lik bir son konsantrasyon için 10 mL steril deiyonize H2O içinde liyofilize hCG tozun yeniden süspanse 10.000 U. Daha sonra, 100 U / ml 'lik son bir konsantrasyon için 9 mL H2O içinde 1.000 U / ml hCG çözeltisi 1 ml seyreltin. 4 ° C'de hem de çözümler Mağaza
- 1. günde, 2:00-15:00 arasında hCG ile enjekte edilerek döşeme yumurta için 4-6 kurbağa hazırlar. 0.2 mi, 100 U / ml hCG çözeltisi ile lağım yakın dorsal lenf kesenin her kurbağa enjekte edilir. Sonraki iki enjeksiyon sırasında kurbağa hızlı olan yumurtlama sırasında kurbağa atık mevcut miktarını en aza indirmek, ancak isteğe bağlıdır olacaktır.
- Günde 2, aynı kurbağa enjekte2:00-3:00 arasında 0,2 mi, 100 U / ml hCG çözeltisi eklenmiştir.
- 3. günde, AM 7:00-10:00 arasında, 0.2 ml 1000 U / ml hCG çözümü ile aynı kurbağa enjekte. Taze tankwater ile 6-litre plastik kova doldurmak, 25 karanlıkta kurbağa ve yer ekle ° C: yumurtalarını bırakmak için kurbağa kurmak Bu enjeksiyon sonra, yumurtlama 4 saat sonra başlayacak ve 7 saat ile tamamlanmış olacaktır. Kurbağalar 25 en az tutulur bir ortamda yumurta ° C yatıyordu gerekir
- Çözüm özü ve yumurta toplama hemen önce hazır donanıma sahip olun.
20X MMR: 100 mM HEPES, pH 7.8, 2 mM EDTA, pH 7.8, 2 M NaCl, 40 mM KCl, 20 mM MgCl2, 40 mM CaCl2. Oda sıcaklığında otoklav ve mağaza. Hazırlık ayıklamak hemen önce 1X MMR 1 L hazırlayın.
10X XB: 100 mM HEPES, pH 7,7, 10 mM MgCl2, 1 mM CaCl2, 1 M KCI, 500 mM sükroz. 4 ° C'de otoklav ve mağaza 1X XB J 'nin 1 L hazırlanmasıhazırlık ayıklamak önce ust. Dejelly çözeltisi: 10 N NaOH ile 7.8-8.0 'ye kadar deiyonize H2O ve pH değeri de 250 ml% 3 sistein çözeltisi hazırlayın. Hazırlık ayıklamak hemen önce hazırlayın.
CSF-XB: 1X XB 200 ml almak ve 2 ml 0.5 M EGTA pH 7.7 ve 200 ul 1 M MgCl2 ekleyin. Hazırlık ayıklamak hemen önce hazırlayın.
BOS-XB +: BOS-XB 50 ml almak ve LPC 50 ul (10 mg / ml her Leupeptin, Pepstatin stok ve DMSO içinde kimostatin) ekleyin. Sitokalasin B 50 ul (DMSO içinde 10 mg / ml) ilave edilir. Hazırlık ayıklamak hemen önce hazırlayın.
Sterilize çözülür ve filtre deiyonize H 2 O, mikrodalga% 0.2 jelatin çözeltisi hazırlayın. Oda sıcaklığında saklayın.
Rezerv 2 Beckman 2 x ½ inç Ultrasantrifüj tüpler.
Minder her H 2 O 0.5 ml ile iki 15 ml cam yuvarlak dipli santrifüj tüplerine hazırlayıntüp ultrasantrifüjdeki.
Yangın parlatılmış cam Pasteur pipetler olun. Geniş bir açılış ortaya çıkarmak için 5 ¾ inç cam pipetler sonunda koparmak, ve yeni ortaya pipet düzeltmek için alev maruz.
- Kat beher duvarlarına yaklaşık% 0.2 jelatin solüsyonu dönen tarafından yumurta depolanması için bir 500 ml'lik bir cam kabın hazırlayın. Kullanımdan sonra kaptan jelatin çözüm atın.
- 3. günde üçüncü enjeksiyondan sonra 6-7 saat döşeme için kullanılan plastik kova yumurta toplayın. İsterseniz, yavaşça kalan yumurta almak için bir kez her kurbağa sıkın. Taze tankwater ve% 0.2 jelatin çözeltisi ile kaplanmış 500 ml'lik bir cam kabın transferi ile bir kez yumurta yıkayın.
2. X ayıklayın hazırlanması tropicalis Yumurta
Ekstresi preparatı her adım, yaklaşık 25 ° C'de, oda sıcaklığında gerçekleştirilebilir Yıkama boyunca, yumurta tutmak için önemlidironlar ıslak kalır, böylece s sıvı altında kalmış. Havaya maruz kalma yumurta hücre döngüsü tutuklama kaçmak veya lyse neden olabilir.
- Yumurta ıslak tutmak için yeterli sıvı saklı tutarak mümkün olduğu kadar tankwater olarak Durusu. Yan yumurta içeren beher yatırın ve beher duvarına doğru ilerler ve ~ 300 mL 1X MMR eklemek, yumurtaların böylece fiziksel ajitasyon en aza indirilir. Süpernatant içeren enkaz kapalı sonra süzün, yumurta dinlendiriniz. X tropicalis yumurta bu adımda lifli, böylece aktive yumurta çıkarılması dejellying sonra yapılır. Üç 1X MMR yıkar toplam tekrarlayın.
- Dejelly yumurta. Dejelly çözüm mümkün ve eklemek yarısı kadar MMR olarak kapalı süzün. Yaklaşık 5 dakika boyunca sürekli olarak girdap. Jöle kat çözme birkaç dakika sonra süpernatant görünür olacaktır. Kapalı Durusu ve kalan dejelly çözüm ekleyin. Yumurta çok sıkı paketi sürekli kadar girdap devam ve bitkisel kutup (tüm orient kutupbeyaz pigment ile) çanak altına doğru. Hızla mümkün olduğu kadar çok dejelly çözüm olarak kapalı süzün. Yumurta dejellied Bir kez onlar mekanik manipülasyonlar çok duyarlıdır.
- Dikkatlice yumurta XB ekleyin. İlk XB yıkama, parçalanmış, kabarık, beyaz ve pseudocleavage yumurta kaldırarak BOS tutuklama kaçmış yumurta çıkarın. Aktif X tropicalis yumurta üst orta yerleşmek eğilimindedir, bu nedenle bu çekin için plastik bir transfer pipet kullanın. Ayrıca deri ve kurbağa atık parçaları çıkarın. Nazikçe yıkar arasında yumurta dönen ve beher altına yerleşmek için izin, ~ 300 ml 1X XB solüsyonu ile yumurta üç kez toplam yıkayın. Mümkün olduğu kadar her bir yıkama çözeltisi yumurta ıslak tutarak daha önce olduğu gibi, süzün.
- CSF-XB ve süzün ile iki kez yumurta yıkayın.
- BOS-XB Ekle + yumurta. E maruz özen, bir jelatin ile tedavi edilen yangın cilalı Pasteur pipeti, BOS-XB + ile transferi yumurta Ultra santrifüj tüplerine kullanılmasıhavaya ggs. Su tamponu ile 15 ml bir cam santrifüj tüpleri içine yerleştirin.
- 1 dakika boyunca 200 xg'de bir klinik santrifüj yumurta Spin, 30 saniye boyunca 800 x g ve spin hızını artırır.
- Yumurta mümkün olduğu kadar tampon kaldırmak için bir aspiratör kullanın. Onlar en neredeyse kuru olmalıdır. Hızlı bir şekilde HB-6 rotor (veya eşdeğeri) ile donatılmış bir Sorvall RC-6 santrifüj yumurta taşıma ve 20 ° C'de 15 dakika 17,000 x g'de dönmeye ° C
- Bir 1 ml şırınga bağlanmış bir 18 gauge iğne kullanılarak pigment ve lipid tabakaları arasında sarı sitoplazmik katmanı kaldırın. Delinme borunun yan ve yavaş yavaş sitoplazmik özü tabakasını elde etmek için bir şırınga çekin. Pigment granülleri mümkün olduğu kadar önlemek.
- Yeni Ultrasantrifügasyon tüp sitoplazma aktarın. Özü bu adımda biraz bulutlu görünmesi normaldir. 15 ml cam santrifüj tüpüne su yastığı ile içinde yer. 20 ° C'de 10 dakika boyunca tekrar 17,000 x g'de çevirin Ext tekrarlayın18 gauge iğne ile raction.
- 1.5 ml mikrofüj tüpe sitoplazma aktarın. Özü hacmi tahmin ve özü içine Cytochalasin D ve LPC 1:1,000 sulandırmak. , 1 ml pipet ile iyice karıştırın hava kabarcıkları giriş olmadan birçok kez aşağı yukarı pipetleme ve. Sağlıklı bir kurbağa koloniden tipik bir verim özü / kurbağa yaklaşık 300-500 ul. Maksimum etkinlik korumak için, bu muhafaza edilir ve oda sıcaklığında deneysel manipülasyon (20-25 ° C) için gereklidir.
3. X arınma Taxol-stabilize Mikrotübüller tropicalis Özü
- 10 uM'lik bir son konsantrasyonda ekstresi 100-200 ul kısım için Taxol ekleyin ve 30 dakika boyunca oda sıcaklığında inkübe edin. Kontrol reaksiyonları için, mikrotübül destablilzing ilaç Nokodazol (10 uM) ile özü, eşdeğer hacimde tedavi. Analizi için işlem görmemiş ekstrenin rezervasyon 100 ul.
- Ilaç ile muamele edilmiş seyreltilir10 hacim BRB-80 (80 mM PIPES, pH 6.8, 1 mM MgCl2, 1 mM EGTA) +% 30 gliserol ile çıkarınız. BRB-80 +% 60 gliserol yastık 10 ml içeren 14 ml yuvarlak alt polipropilen borular monte edin. Geniş çaplı bir pipet, katman BRB-80 üstüne hafifçe ilaç ile muamele edilmiş ekstrakt Reaksiyon +% 60 gliserol tampon kullanarak. 20 17.000 x g'de 10 dakika santrifüj ° HB-6 rotor (veya eşdeğeri) ve tüp adaptörleri ile donatılmış bir Sorvall RC-6 santrifüj C.
- Içeren süpernatan unsedimented özü madde aspire edin ve deiyonize H 2 O. ile iki kere yıkayın arayüz Mikrotübül, mikrotübül ilişkili protein ve Taxol ile tedavi edilen örnek mikrotübül ilişkili RNA'lar içeren jel benzeri pelet rahatsız etmemek için özen, yavaş yavaş kalan yastık hacmi aspire. Nokodazol-işlemden geçirilen numune görebilir madde içermez. 1 ml TRIzol içinde pelletini tekrar ve RNA izole etmek için üreticinin talimatlarına devam edin. Untreated ekstresi (100 ml), 1 ml TRIzol doğrudan yeniden süspanse edilebilir.
- RNA-seq uygun transcriptome kütüphaneler hazırlanması için ticari kitleri şimdi var. Bu satın alınabilir http://www.illumina.com/ ve http://www.454.com/ .
Representative Results
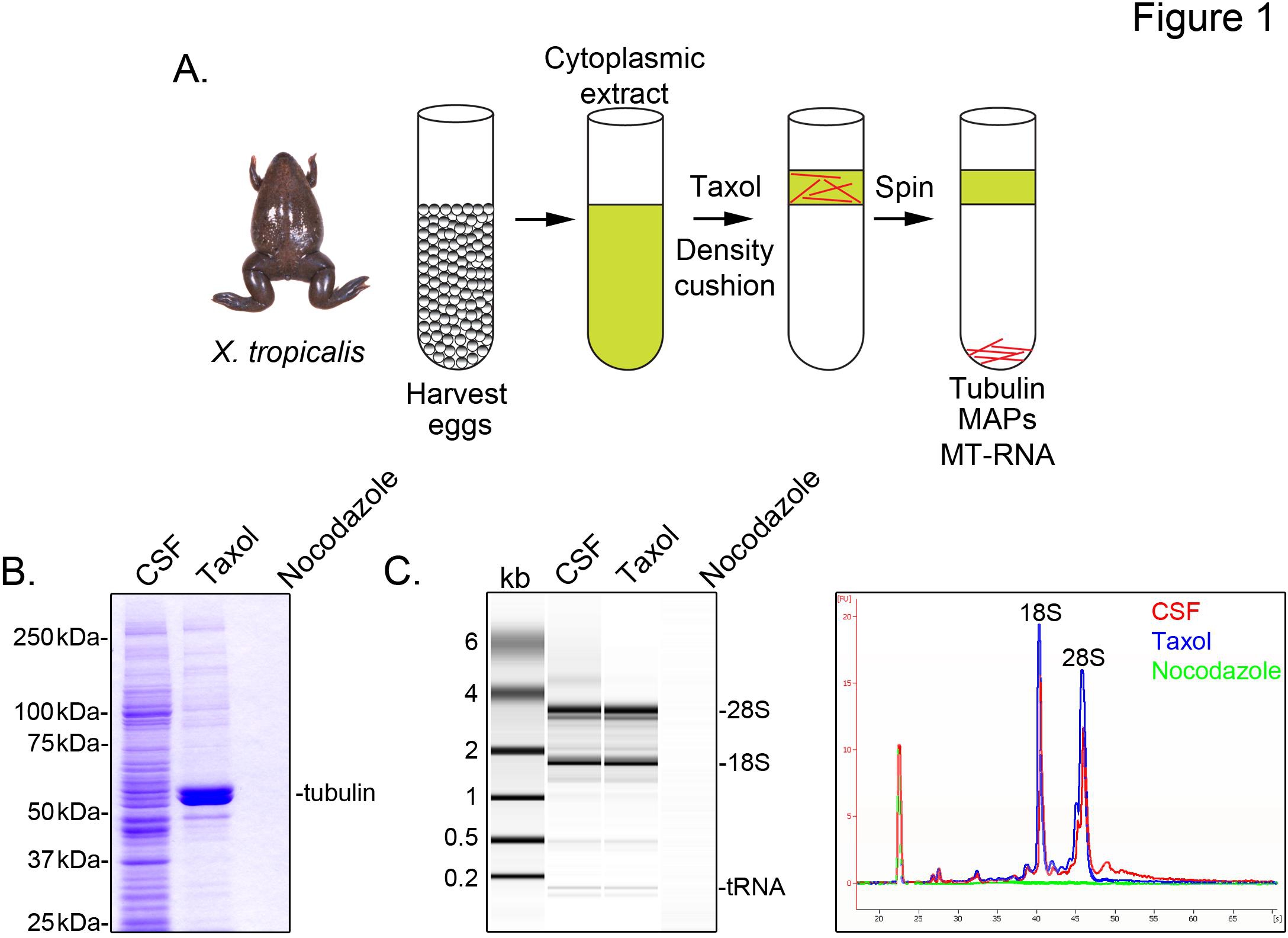
X belirlemek için mikrotübül ilişkili tropicalis transkript, biz mayoz II (BOS) metafaz tutuklandı döllenmemiş yumurta bir sitozolik özü hazırlamak. Taksol ile özü tedavisi gliserol yastık (Şekil 1A) ile çökelme ile daha da arıtılabilir stabil mikrotübül oluşumunu sağlar. Coomassie jel analizi α / β-tubulin bir taksol bağımlı bir şekilde çökeltiler ve ana protein türü temsil etmektedir, bu preparatlar (Şekil 1B) geri kazanılır olduğunu doğrular. Diğer proteinlerin düşük seviyelerini gösteren, ilaç Nokodazol depolimerizasyon mikrotübül ile tedavi hazırlıkları da taksol pelet mevcut, ama bu mikrotübül (MAP) ile özellikle ortak taksol kısmını proteinler.
Bir Agilent Bioanalyzer tüm X genel RNA kompozisyon incelemek için kullanılır tropicalis özü fraksiyonları (Şekil 1C X'te mikrotübüllerin ve iğ oluşur daha önceki bulgularla tutarlı, BOS özü ve mikrotübül içeren Taksol pelet mevcuttur laevis yumurta 5,21 ayıklayın. Jel projeksiyon bir çizgi izlemesi mRNA sinyali belirgin düşüktür ortaya mikrotübül içeren gösteren, 28S rRNA üzerindeki bölgede göç en önemlisi taksol pelet, bu X'te mikrotübüllerin ile mRNA cosediment bir alt tropicalis. Bu şekilde izole edilen RNA, ticari olarak temin edilebilen reaktifler kullanılarak, RNA-DİZ deneyler için uygundur.

Şekil 1. RNA-DİZ MT-RNA saflaştırılması. MT-RNA izole etmek için (A), saflaştırma şeması. Yumurta dişi X hasat edilir tropicalis kurbağa. Sitoplazmik bir ekstresinin hazırlanması sonra taksol mikrotübül polimerizasyon ikna etmek için ilave edilir. Mikrotübüller ve MT-RNA, gliserol yastığı sedimantasyon ile saflaştırılır. (A) 'de tarif edilen şema kullanılarak izole edilmiş protein (B), Coomassie jel analizi. Toplam CSF taksol veya Nokodazol varlığında çöktürülmüştür proteinleri ile karşılaştırıldığında ekstraktıdır. (C) RNA, Bioanalyzer jel analizi (A) 'de tarif edilen şema kullanılarak izole edilmiştir. BOS RNA izole taksol veya Nokodazol varlığında çöktürülmüştür RNA ile karşılaştırıldığında özü. Jel projeksiyon hattı izlerinin her ikisini de gösterilmiştir. Keskin, et al., (2011) izni ile yayımlanmaktadır. Büyük resim görmek için buraya tıklayın .
Discussion
Bu yazıda X BOS-tutuklandı yumurta özleri üretmek için basit bir yöntem tarif var tropicalis 19 ve mikrotübül ilişkili RNA'lar 7 incelemek için bu özü kullanın. X den BOS-tutuklandı yumurta özler üretmek için temel prosedür X için kullanılan tropicalis aynıdır birkaç önemli farklılıklar ile laevis. X ile çalışan en zor yönlerinden biri tropicalis kurbağa X karşılaştırılabilir mikrotübül çekirdeklenme veya milli montaj aktivite ile bir özü için yeterli kaliteli yumurta elde edilir laevis yumurta ayıklar. Mayoz II hücre döngüsü tutuklama, X için hormon enjeksiyonları arasındaki aralıktan kayma önlerken en uygun yumurtlama koşulları elde etmek için tropicalis X için kullanılan daha kısadır laevis ve yumurtlama başına üçüncü hCG enjeksiyonundan zamanlama da çok kısadır. X ile hCG enjeksiyonundan t zamanlama laeviso yumurtlama başında bu tampon içine bir gecede döşenecek yumurta için uygun ve etkili olduğu gibi. Ancak, hCG enjeksiyonu ve yumurta arasındaki kısa zaman X ile döşeme sık sık el kurbağalar yumurta ifade için gerekli olan tropicalis. Iki farklı kurbağalar yumurta özü yapma arasındaki bir diğer önemli fark dejellying adımdır. X ile yumurta laevis bu jöle kat yumurta beher ne kadar yakın yerleştirileceğini gözlemleyerek çözünmüş ne zaman belirlemek kolaydır o kadar büyüktür. Dejellying reaksiyon başlangıcında, yumurta daha yoğun paketi başlar. Ancak, X. tropicalis yumurta çok daha küçük ve jöle kat tek başına yoğunluğu ambalaj yumurta tarafından çözülür ne zaman belirlemek için oldukça zor olabilir. Biz, jöle kat çözündürüldü ne zaman tamamlandığını belirlemek için en güvenilir yöntem hayvan (siyah) ve bitkisel (beyaz) kutup yönü izleme olduğunu bulduk. Ne zaman tüm bitkisel kutup oribeher altına doğru ent jöle kat ekstresi ile devam etmek için yeterli kaldırıldı. Son olarak, oysa X laevis yumurta özü serin sıcaklıklarda saklanabilir (4-12 ° C) bunu X korumak için kritik olduğunu gözlemledik hazırlanması ve deneysel manipülasyonlar sırasında oda sıcaklığında tropicalis yumurta özü (20-25 ° C) biyokimyasal faaliyet korumak için. Çünkü kullanım kolaylığı farklılıklar biz X kullanmayı tercih Yumurta özü üretimi için laevis kurbağa. Ancak, ihtiyaç ya da sıralı genom, X ile bir organizma tarafından kolaylaştırdı deneyler için tropicalis mükemmel bir alternatif sistemidir.
Bu raporda açıklanan olduğunu yöntemi mikrotübül polimerizasyon ikna etmek için bir mikrotübül-dengeleyici ajan olarak taksol kullanır. Taksol mikrotübül saflaştırılmış büyük ölçekli izolasyon kolaylaştıran sağlam bir mikrotübül stabilize edici, çünkü bu yöntem seçtik. Gibi bir yöntem incibiz tarif de olası alternatif mikrotübül polimerizasyon yöntemleri kullanılarak mikrotübül ilişkili protein ve RNA karşılaştırarak geliştirilebilir. Alternatifler GTP bağlı polimerizasyon (klasik bir teknik), 22 veya kromatin odaklı mili montaj 23 neden olduğu mikrotübül taklit etmek için bir mikrotübül polimerizer olarak Ran-GTP kullanarak kullanarak polimerizasyon içerebilir. Son olarak, mikrotübül polimerizasyon en yakın mitoz (centrosome, kromatin ve kinetochore aracılı) sırasında çekirdeğini olan mikrotübül türlerine taklit olacağını ikna etmek için sperm çekirdekleri saflaştırılmış kullanın. Mikrotübül çekirdeklenme bu alternatif kaynaklara dezavantajları çekirdekleştirici ajanlar taksol kadar hazır değildir ve onlar çekirdekli yok ya da verimli Taksol olarak mikrotübül stabilize. Bu nedenle, bu yöntemlerin her büyük ölçekli saflaştırılması için kullanmak daha zor olacaktır. Mikrotübül Nükleatörler birden fazla farklı türde karşılaştırma avantajıbu protein ve / veya mikrotübül çekirdeklenme her yolu özgü RNA'lar belirlemek mümkün olabilir olmasıdır.
Burada anlattığımız yöntem, amfibi sitoplazmik özleri yararlanır. Ancak, bu yaklaşım diğer organizmalardan özü sisteminin kullanımı için uzun olabilir. Mitotik özleri sadakatle mikrotübül montaj birçok açıdan özetlemek bu senkronize insan doku kültürü hücreleri 24 tanımlanmıştır. Başarıyla HeLa hücreleri 5'ten mikrotübüle bağlı RNA'lar tanımlamak için bu ekstreler kullandık. Mikrotübül ilişkili RNA'lar incelenmiş değil de benzer mikrotübül arıtma planları, birçok farklı organizmalar 25,26 için tanımlanmıştır. Burada açıklanan yaklaşım, mikrotübül çekirdekleştirici yeteneğine sahip bir konsantre sitoplazmik özü üretebilen herhangi bir organizma ile kullanılabilir.
Son olarak, her ne kadar bu biz de yaklaşımBurada yazıcı mikrotübül ve ilişkili protein ve RNA'lar arıtma tartışır, bu yaklaşım diğer hücre içi yapılara genel olabilir. En lokalize mRNA biyokimyasal yöntemlerle tespit edilmemiş olsa da DNA ve RNA dizi teknolojilerindeki son gelişmeler bu yaklaşımın yerel RNA'lar tanımlamak için cazip bir yöntem yapmak. Bu yaklaşımda ilgi herhangi bir hücre içi veya alt embriyo yapısı izole veya saf olabilir. Daha sonra ilgili protein ve RNA bir genom geniş ölçekte tespit edilebilir. RNA'lar sonra, toplam hücre ya da zenginleştirilmiş lokalize RNA'lar tespit etmek embriyo RNA içeriği ile karşılaştırılabilir. Bu yaklaşım, bütün olarak (27, Xenopus birinci lokalize RNA'lar tespit yaklaşım benzer hayvansal ve bitkisel ayırma), yumurta, aktin ilişkili RNA'lar ER-ilişkili RNA'lar mitokondri ilişkili RNA'lar ile ya da herhangi bir olabilir hücre içi yapısı için kullanılabilir bozulmamış ilişkili RNA'lar ile daha da saflaştırılabilir. Göremikrotübül ilişkili RNA üzerinde çalışmalarımızı bu belirli bir yerde yeni bir protein bu işlevi keşfetmek için mükemmel bir yöntem olacağını tahmin. Ayrıca, tüm yerelleştirilmiş RNA'lar yerini ve kapsamı belirlenmesi hücreleri ve embriyoların gen ekspresyon kontrol etmek için mRNA lokalizasyon kullanarak nasıl içine fikir verecektir.
Disclosures
Çıkar çatışması ilan etti.
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| Xenopus tropicalis | NASCO | LM00823MX | |
| human Chorionic Gonadotropin | Sigma-Aldrich | CG10 | |
| HEPES | Sigma-Aldrich | H4034 | |
| EDTA | Sigma-Aldrich | E5134 | |
| NaCl | Sigma-Aldrich | S3014 | |
| KCl | Sigma-Aldrich | P9541 | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| CaCl2 | Sigma-Aldrich | C8106 | |
| sucrose | Sigma-Aldrich | S0389 | |
| NaOH | Sigma-Aldrich | S5881 | |
| EGTA | Sigma-Aldrich | E3889 | |
| Leupeptin | Sigma-Aldrich | L9783 | |
| Pepstatin | Sigma-Aldrich | P5318 | |
| Chymostatin | Sigma-Aldrich | C7268 | |
| Cytochalasin D | Sigma-Aldrich | C8273 | |
| Gelatin, porcine skin | Sigma-Aldrich | G1890 | |
| PIPES | Sigma-Aldrich | P6757 | |
| Taxol | Sigma-Aldrich | T7191 | |
| Nocodazole | Sigma-Aldrich | M1404 | |
| Trizol | Invitrogen | 15596-026 | |
| L-Cysteine, free base | USB Corporation | 14030 | |
| Cichlid Lake Salt | Seachem | 47894 | |
| Marine salt | Seachem | SC7111 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| EQUIPMENT | |||
| 1 ml syringes | BD Biosciences | 309659 | |
| 18 gauge needles | BD Biosciences | 305195 | |
| 30 gauge needles | BD Biosciences | 305106 | |
| Rubbermaid Plastic bucket | Amazon | 6306 | |
| Beckman Polyallomer 2 x ½ inch Ultracentrifuge tubes | Beckman | 326819 | |
| 15 ml round-bottomed glass centrifuge tubes | Fisher Scientific | 45500-15 | |
| Rubber adapter sleeves for 15 ml tubes | Kimble-Chase | 45550-15 | |
| 5 ¾ inch glass Pasteur pipettes | Fisher Scientific | 13-678-20A | |
| 14 ml polypropylene round-bottom tube | BD Biosciences | 352059 | |
| Sorvall HB-6 rotor | Thermo Scientific | 11860 | |
| Sorvall RC-6 centrifuge | Thermo Scientific | 46910 | |
References
- Martin, K. C., Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell. 136, 719-730 (2009).
- Besse, F., Ephrussi, A. Translational control of localized mRNAs: restricting protein synthesis in space and time. Nat. Rev. Mol. Cell Biol. 9, 971-980 (2008).
- Lawrence, J. B., Singer, R. H. Intracellular localization of messenger RNAs for cytoskeletal proteins. Cell. 45, 407-415 (1986).
- Mingle, L. A., et al. Localization of all seven messenger RNAs for the actin-polymerization nucleator Arp2/3 complex in the protrusions of fibroblasts. J. Cell Sci. 118, 2425-2433 (2005).
- Blower, M. D., Feric, E., Weis, K., Heald, R. Genome-wide analysis demonstrates conserved localization of messenger RNAs to mitotic microtubules. J. Cell Biol. 179, 1365-1373 (2007).
- Eliscovich, C., Peset, I., Vernos, I., Mendez, R. Spindle-localized CPE-mediated translation controls meiotic chromosome segregation. Nat. Cell Biol. 10, 858-865 (2008).
- Sharp, J. A., Plant, J. J., Ohsumi, T. K., Borowsky, M., Blower, M. D. Functional analysis of the microtubule-interacting transcriptome. Mol. Biol Cell. 22, 4312-4323 (2011).
- Lecuyer, E., et al. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell. 131, 174-187 (2007).
- Mili, S., Moissoglu, K., Macara, I. G. Genome-wide screen reveals APC-associated RNAs enriched in cell protrusions. Nature. 453, 115-119 (2008).
- Holt, C. E., Bullock, S. L. Subcellular mRNA localization in animal cells and why it matters. Science. 326, 1212-1216 (2009).
- Desai, A., Murray, A., Mitchison, T. J., Walczak, C. E. The use of Xenopus egg extracts to study mitotic spindle assembly and function in vitro. Methods Cell Biol. 61, 385-412 (1999).
- Gache, V., Waridel, P., Luche, S., Shevchenko, A., Popov, A. V. Purification and mass spectrometry identification of microtubule-binding proteins from Xenopus egg extracts. Methods Mol. Med. 137, 29-43 (2007).
- Gache, V., et al. Xenopus meiotic microtubule-associated interactome. PLoS One. 5, e9248 (2010).
- Cross, M. K., Powers, M. Obtaining eggs from Xenopus laevis females. J. Vis. Exp. (18), e890 (2008).
- Cross, M. K., Powers, M. Preparation and fractionation of Xenopus laevis egg extracts. J. Vis. Exp. (18), e891 (2008).
- Cross, M., Powers, M. In vitro nuclear assembly using fractionated Xenopus egg extracts. J. Vis. Exp. (19), e908 (2008).
- Harland, R. M., Grainger, R. M. Xenopus research: metamorphosed by genetics and genomics. Trends Genet. 27, 507-515 (2011).
- Hellsten, U. The genome of the Western clawed frog Xenopus tropicalis. Science. 328, 633-636 (2010).
- Brown, K. S. Xenopus tropicalis egg extracts provide insight into scaling of the mitotic spindle. J. Cell Biol. 176, 765-770 (2007).
- Godfrey, E. W., Sanders, G. E. Effect of water hardness on oocyte quality and embryo development in the African clawed frog (Xenopus laevis). Comp. Med. 54, 170-175 (2004).
- Groisman, I., et al. CPEB, maskin, and cyclin B1 mRNA at the mitotic apparatus: implications for local translational control of cell division. Cell. 103, 435-447 (2000).
- Budde, P. P., Desai, A., Heald, R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 38, 29-34 (2006).
- Kalab, P., Pu, R. T., Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 9, 481-484 (1999).
- Gaglio, T., Saredi, A., Compton, D. A. NuMA is required for the organization of microtubules into aster-like mitotic arrays. J. Cell Biol. 131, 693-708 (1995).
- Hughes, J. R., et al. A microtubule interactome: complexes with roles in cell cycle and mitosis. PLoS Biol. 6, e98 (2008).
- Suprenant, K. A., Tempero, L. B., Hammer, L. E. Association of ribosomes with in vitro assembled microtubules. Cell Motil. Cytoskeleton. 14, 401-415 (1002).
- Rebagliati, M. R., Weeks, D. L., Harvey, R. P., Melton, D. A. Identification and cloning of localized maternal RNAs from Xenopus eggs. Cell. 42, 769-777 (1985).

{kind=link}