

Summary
유사 분열을 겪고있는 암 세포에있는 phenanthridine PJ-34의 세포 독성 활동은 라이브 공 촛점 이미징 실시간으로 기록되었다. PJ-34 근절 인간의 유방암 MDA-MB-231 세포의 유사 분열에 여분의 중심체를 숨겨 세포. 정상 이중 초점 유사 분열과는 달리, 여분 중심체는 PJ-34의 존재에있는 두 개의 스핀들 극에 클러스터되지 않았습니다.
Abstract
강력한 PARP1 억제제 역할을 페난 트렌 유도체 유사 분열의 멀티 centrosomal 인간의 암 세포에있는 여분 중심체의 이중 초점 클러스터링을 막았다. phenanthridine PJ-34은 가장 강력한 분자이다. 여분 중심체의 Declustering 멀티 centrosomal 세포에서 유사 분열 장애 및 세포 죽음을 발생합니다. 대부분의 고체 인간의 암은 여분 중심체의 높은 발생했습니다. PJ-34의 활동은 살아있는 인간의 유방암 중심체에 매우 풍부 형광 γ-tubulin의를 위해 인코딩 벡터에과 형광 히스톤 H2B 선물로 형질 전환 된 MDA-MB-231 세포의 공 촛점 이미징에 의해 실시간으로 기록되었다 염색체. 비정상적인 염색체 배열 및 declustered 중심체를 나타내는 데 클러스터 γ-tubulin의 병소는 PJ-34 치료 후 형질 전환 MDA-MB-231 세포에서 발견되었다. 두 개의 스핀들 극 유엔 클러스터 여분 중심체는 세포의 죽음을 앞에. 이러한 결과는 FO를 연결R 처음 여분 중심체 유사 분열에 드 클러스터링 및 세포 사망에 이르는 세포 분열 장애를 가진 인간의 암 세포에있는 PJ-34의 최근 발견 독점 세포 독성 활동. 두 개의 중심체와 이중 초점 스핀들과 분열을 겪고 정상 세포를 저해하지 않고 고정 된 세포 다 중심체와 PJ-34 전용 근절 암 세포의 공 촛점 이미징에 의해 관찰 이전의 연구 결과에 따르면. PJ-34이 세포 독성 활성이 다른 강력한 PARP1 억제제에 의해 공유되지 않은, 그리고 PARP1 억제의 독립성을 제안, PARP1 부족 MEF 숨겨 extracentrosomes에서 관찰되었다. 공 촛점 이미징을 살고 유사 분열 동안 세포를 근절 새로운 분자를 식별하기위한 유용한 도구를 제공했다.
Introduction
페난 트렌는, PJ-34 등 PARP1 억제제, 파생 스트레스 조건 (뇌졸중이나 심근 경색 1)에 따라 에너지 소비 PARP1 매개 DNA 수선에 의해 유도 된 세포 사멸 세포 사멸에서 정지 세포를 보호하도록 설계되었습니다. 그러나 최근 우리는 PJ-34, 그 유도 PARP1 억제보다 두 배 높은 농도에서 독점적으로 인간의 암 세포 2,3에서 세포 죽음을 일으킬 수 있다는 것을 발견. 세포의 더 급속한 확산이었다, 세포의보다 효과적인 근절했다. PJ-34의 세포 독성 활동은 여분 중심체 유사 분열 2 드 클러스터링에 기인 하였다. 사람의 암세포 항구는 4,5 multicentrosomes. 효율적으로 유사 분열에서 두 개의 중심체를 숨겨 정지 세포 또는 일부 양성 증식 세포를 저해하지 않고 72-96 시간 이내에 세포를 박멸 20 μM PJ-34와 여분 중심체를 항구 인간의 유방암 세포 MDA-MB-231의 배양
양극성 중심체 어셈블리는 유사 분열 4,5 양극성 스핀들 형성에 매우 중요합니다. 따라서 두 개 이상의 중심체를 가진 세포는 두 개의 기둥 4-9에서 자신의 별도의 중심체를 클러스터링, 거의 이해하지 분자 메커니즘을 개발했습니다. 자신의 중심체의 양극 어셈블리의 실패 체포 G2 / M 정지의 세포주기 및 세포 죽음에 이르게 4,5 유사 분열 장애에 기인하는 다극 왜곡 스핀들 및 탈선 염색체 분리가 발생할 수 있습니다. 여분 중심체 드 클러스터링을 기본 분자 메커니즘을 집중적으로 조사하는 <> 10을 먹다. 건강한 조직 5,10을 살려주는 동안이 죽음의 메커니즘을 이해하는 것은 암 세포의 독점적 인 박멸을 가능하게 할 것이다.
따라서, 유사 분열 재앙 세포 죽음을 활성화 화합물은 공 촛점 이미징 여분 중심체 클러스터링에 영향을 미치는 분자를 식별하는 데 사용할 수있는 것이 좋습니다 인간의 고체 cancers.Our 결과의 넓은 범위에서 효율적일 수 있습니다 선택적 암 치료의 새로운 모드를 제공 유사 분열 2,3, 약물 후보를 대상으로 이들 화합물의 암을 렌더링합니다.
우리는 고정 살아있는 인간의 암 세포 (유사 분열의 여분 중심체의 높은 발생에) 대 정상 세포를 스캔하여 phenanthridine PJ-34의 세포 독성 활성을 기록했다. 단계별로 인간의 암 세포 PJ-34의 세포 독성 활동을 식별하는 데 사용되는 이미징 절차의 단계 설명은 아래에 포함되어 있습니다.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. 세포 배양 준비
MDA-MB-231 세포는 ATCC (미국 유형 문화 수집)에서 구입하여 액체 질소에 보관 하였다.
- Dulbeco 수정 이글 중간 (DMEM), 10 % 말 혈청, 1 % L-글루타민, 1 % Penstrep-Amphotericine B를 포함하는 10 ML 완전 배지 92 mm 직경 페트리 접시에 씨앗 10 6 MDA-MB-231 세포는 세포를 허용 80 ~ 100 % 정도의 합류로 증식한다.
- 요리에서 배지를 제거하고 폐기합니다.
- 0.25 % 혈청의 모든 흔적을 제거하는 (W / V) 트립신 - EDTA (에틸렌 다이아 민 테트라 초산) 솔루션을 짧게 세포 층을 씻으십시오.
- 세포 층 (보통 5 - 15 분 이내) 분산 될 때까지 거꾸로 현미경으로 세포를 접시 관찰 트립신 - EDTA (에틸렌 다이아 민 테트라 초산) 솔루션의 2.0 ML를 추가합니다.
- 18 ML 전체 성장 매체를 추가하고 부드럽게 피펫으로 세포를 대기음. 튜브에 전송할 수 있습니다.
- 1,200 rpm으로 세포 현탁액을 원심 분리기.
- CU의 24 ML에 세포 펠렛을 다시 일시 중지매체를 lture.
- 35mm 유리 바닥 요리 (25 % 합류에 대한) 및 인큐베이터의 장소 (5 % CO 2, 37 ° C)에 세포 현탁액 2 ㎖를 추가합니다.
- 솔루션 :
- 세포 증식을위한 완벽한 매체 : 10 % FBS와 DMEM, 1 % 항생제 (100 단위 / ML 페니실린 G, 100 ㎍ / ㎖ 스트렙토 마이신, 펜 연쇄상-Ampho 솔루션), 2 mM의 L-글루타민.
- 0.25 % 트립신 - EDTA (에틸렌 다이아 민 테트라 초산)를 포함하는 트립신 - EDTA (에틸렌 다이아 민 테트라 초산) 솔루션.
- 요리 :
92mm 직경 페트리 접시.
35mm 직경 폴리 - D - 라이신 코팅 유리 바닥 문화 요리.
2. 라이브 공 촛점 이미징 세포의 준비
- 시드 2 × 10 제 1 항에서 언급 한 2 ㎖ 완전 배지에서 유리 바닥 문화 요리 5 MDA-MB-231 세포. 세포 배양은 60 % ~ 70 %의 합류 (접시 당 3-4 × 10 5 세포에 대한)에 도달하면, 형질 진행합니다.
- 두 개의 플라스미드는 엔코딩으로 세포를 형질제조 프로토콜에 따라 리포솜의 transfection 시약 제트-PI를 사용하여 융합 단백질 γ-tubulin의-GFP (중심체의 형광 검출)와 히스톤-RED (H2B-RED, 염색체의 형광 검출을) ING. 간단히, 100 ㎕의 염화나트륨 (150 ㎜)과 관의 각 플라스미드의 2 μg을 혼합한다. 두 번째 튜브에 100 ㎕의 염화나트륨 (150 밀리미터)로 형질 시약 (100 μL)를 혼합하고 실온 (RT)에서 5 분 알을 품다. 다음 두 가지 솔루션, 혼합 (약한 소용돌이를 사용하여) 결합 스핀 다운. RT에서 30 분 알을 품다.
- 형질 혼합물의 배양 기간 동안 PBS로 두 번 세포를 씻으없이 보충 따뜻한 DMEM 2 ㎖ (37 ° C)로 세포 매체를 교체하십시오.
- 부드럽게 DMEM에 세포의 transfection 혼합물을 추가하고 8 시간 동안 (37 ° C, 5 % CO 2) 배양기에 세포를 반환합니다.
- 24 시간 동안 배양 8 시간 후 2 ㎖ 완전 배지와 DMEM를 교체하고 인큐베이터에서 세포를 배양시간.
- 24 시간 형질 전환 한 후 2 ㎖ 완전 배지는 20 μM PJ-34를 포함하여 세포의 매체를 교체하십시오.
- 추가 18 시간 (37 ° C, 5 % CO 2)에 대한 세포를 품어.
- 5 % CO 2, 37 ℃에서 세포를 유지 이미징 챔버에 적어도 16 시간 동안 공 촛점 이미징을 살 수있는 대상 세포
- 병렬로, 다음과 같이 형광 현미경을 사용하여 형질 효능 36 시간 게시물 형질을 검사 :
- 시드 2 × 10 1 coverslip에 잘 당 2 ㎖ 완전한 매체를 포함하는 6 - 잘 접시에 5 MDA-MB-231 세포.
- 섹션 2.2-2.5에서 언급 한 세포를 형질.
- 36 시간 포스트 transfection에 차가운 메탄올에 부화하여 coverslip을에 장착 된 형질 전환 세포 수정 : 아세톤 (1:1) 용액, 7 분, -20 ° C.
- 고정 솔루션을 기음과 화학 후드에서 건조 장착 세포와 coverslip을 보자.
- DAPI와 골드 antifade 시약을 연장하고 일을하게 적용전자 coverslip에 6 시간 동안 어둠 속에서 건조.
- 형광 현미경 슬라이드를 검사하고 세포의 전체 인구 (DAPI에 의한 DNA의 얼룩)에서 형질 세포 (적색 및 녹색 신호)의 비율을 계산합니다. 100-200 세포가 계산 될 때 원하는 형질의 비율이 20-40 %이다.
3. 라이브 공 촛점 이미징 스캐너 설정의 기술적 인 매개 변수
- ScanMode XYZT; 구멍 [통풍] 1.00, 줌 3.5, 해상도 8 비트, 레이저 DPSS 561 nm의, 아르곤, 가시 레이저 488 nm의, 레이저 그 / 네 눈에 보이는 633 nm의, 목표 HCX PL APO CS 63X 1.40 OIL UV, 수치 조리개 1.4; 굴절률 1.52, 속도 700 Hz에서 검색합니다.
- 이미지를 3 차원 프레 젠 테이션 IMARIS 이미징 소프트웨어 7.0으로 제조 하였다.
4. 고정 된 세포의 유사 분열의 공 촛점 이미징
- 시드 2 × 10 5 MDA-MB-231 유방암 세포 (ATCC), 정상 마우스 배아 섬유 아세포 (MEF) 또는 PARP1 부족 ME2 ㎖ 완전 배지에서 6 - 잘 접시에 유리 coverslips는에 F (박사 프랑소와 Dantzer에 의해 제조 PARP - - /). 의 coverslips는 2 시간 건조하고, 6 - 잘 접시의 각 우물에 배치 멸균 DD 물로 세척하여 다음, 96 % 에탄올로 세척 하였다.
- 매체에 PJ-34 (10-30 μM)를 추가하고 필요한 기간 (일반적으로 최대 96 시간)에 대한 세포를 품어.
- PBS로 한 번 coverslips를 세척 (인산염 완충 식염수), 그리고 얼음처럼 차가운 메탄올에서 배양하여 세포를 고정 : 아세톤 (1:1) 용액, 7 분, -20 ° C.
- 고정 솔루션을 기음과 coverslips를 화학 후드에서 건조하자 (이 단계에서, coverslips를 여러 주 동안 -20 ° C에 보관 될 수 있습니다).
- 세포막을 permeabilize 및 RT에서 1 시간 PBST 10 % NDS (정상 당나귀 혈청) ( '차단 솔루션')로 세포를 차단하는 PBST (PBS 0.1 % 트윈 - 20과 보충) 한 번 coverslips를 씻으십시오.
- 기본 antibodi와 투과 고정 된 세포를 배양RT에서 2 시간 동안 ES (스핀들 및 중심체 염색 용). 반대로 α 튜 블린 (1:250 희석) 및 안티-γ 튜 블린 (1:200 희석) : 항체는 것과 같이 따른다 차단 용액에 희석됩니다. 일차 항체는 다음과 같이 적용됩니다 : 6 잘 플레이트 커버 (덮개가 거꾸로) 각 coverslip에 대한 솔루션을 차단하는 항체의 혼합물 100 μL를 (드롭) 적용됩니다. 부드럽게 항체 드롭 coverslip을 넣어 시드 셀의 드롭에 직면. 실온에서 2 시간 동안 항체를 직면 coverslips를 품어.
- 우물에 다시 coverslips를 배치하고 PBST로 세포를 3 회 세척한다. 다음 형광 차 항체의 coverslips에서 세포를 라벨링 4.6에서 설명한 동일한 절차를 사용합니다. 어둠 속에서, 1 시간, RT에 대한 보조 항체의 coverslips에서 세포를 품어. 항체는 다음과 같은 차단 솔루션에 희석되어 있고, 알렉사 플 루어 568 (1:1,000 희석 알렉사 플 루어 488 (녹색 1:1,000 희석)빨간색).
- 를 사용하여 coverslips를이 DAPI (염색체의 염색을 위해)와 금 antifade 시약을 연장하고 건조 어둠 속에서 RT에서 밤새 배양 마운트합니다.
- 공 초점 현미경으로 커버 전표를 확인합니다.
5. 세포 생존율은 ATP 생산에 의해 측정
ATP 생산은 발광 ATP 탐지 분석 장비로 측정됩니다.
- 96 - 웰 플레이트 각 well에 800 ㎕의 중간에있는 약 20,000 세포의 씨앗 세포를. 세 빈 우물 매체의 배경 발광 측정을 위해 사용되어야한다.
- 약 10 μM에서 100 μM의 ATP 표준 희석 시리즈를 준비하고 얼음에 보관하십시오.
- 각 웰에 세제의 50 μl를 추가하고 궤도 셰이커, 700 rpm으로 5 분 동안 접시를 흔들.
- 키트 '기질 버퍼'의 5 mL로 '동결 건조 된 기질'의 재구성 각각의 병.
- 우물에 재구성 기질 용액 50 μl를 추가하고 흔들어궤도 셰이커, 700 RPM에서 5 분 판.
- 10 분 동안 어둠 속에서 판을 보관하십시오.
- ELISA 마이크로 플레이트 리더 서로 잘 luminiscence을 측정합니다.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 phenanthridine 1 용해 안정적인 물 (그림 1)입니다. 우리의 이전 결과 세포의 죽음을 발표하고 PJ-34로 치료했다 고정 멀티 centrosomal 암 세포의 여러 유형의 엑스트라 중심체를 드 클러스터. 대조적으로, 일반적인 증식 세포는 2,3 손상되지 않았다. 중심체는 고정 여분 centrosomal 세포 2 centrine1 및 γ-tubulin에 대한 항체를 가진 두 배 레테르를 붙이기에 의해 확인되었다.
여기 PJ-34의 세포 독성 활동은 라이브 공 촛점 현미경을 사용하여 실시간으로이 라이브 여분 centrosomal 세포에서 기록되었다. 인간의 유방암 여분 중심체 4,5의 높은 발생 (> 50 %)이 MDA-MB-231 세포를 라이브 γ-tubulin의-GFP (와 형질 세포에 초점을 맞춘 공 촛점 이미징에 의해 적어도 16 시간 동안 스캔 된 형광 γ-tubulin의 병소 2 라벨링)와 히스톤 H2B-RED (형광등 LABE로염색체의 링). 6 ~ 10 라이브 형질 전환 세포를 각각의 실험에 병렬로 스캔 하였다. centrin1와 형질 세포에서 γ-tubulin의 병소의 두 immunolabeling의은 기술적으로 불가능했다.
분산 된 γ-tubulin의 병소와 비정상적인 염색체 배열은 거의 유사 분열에서 무작위로 선택된 치료 MDA-MB-231 세포에서 발견되지 않았다. 여분 중심체에게 초점이 둘인 클러스터링을 나타내는 γ-tubulin의 병소,의 이중 초점 클러스터링 라이브 치료 MDA-MB-231 세포 (그림 2)의 대부분에 설명 된, 대조적으로, 중심체와 염색체의 비정상적인 배열을 해제 클러스터는 라이브에서 발견되었다 형질 전환 MDA-MB-231 PJ-34 (20 μM) 및 세포 사멸 (그림 3)에 의해 종료 이러한 세포의 유사 분열과 배양 세포. 이러한 세포는 18 PJ-34와 함께 배양 - 24 시간 스캔하기 전에 및 추가 16 시간 동안 (그림 3) 스캔 중에. 세포 드의 실시간 문서유사 분열 동안 ath를 강하게 유사 분열의 다극 스핀들 인간의 악성 세포의 수와 PJ-34 (20 μM) 2 배양 세포에서 세포 죽음의 비율 사이에 이전에 정의 된 양의 상관 관계를 지원합니다.
PJ-34 강력한 PARP1 억제제 1 역할을합니다. 따라서 우리는 유사 분열 실패와 관련된 세포 죽음을 일으키는 원인이 PARP1 억제의 가능성을 살펴 보았다. 라이브 영상과는 달리, 고정 MDA-MB-231 세포의 이미징함으로써 인간의 암 세포 라인의 다양한 PARP1 억제제의 효과 통계 분석을 가능하게 세포 배양에서 세포의 큰 인구의 검사를 가능하게했다. (그림 4) PJ-34의 활동은 정상에있는 다른 강력한 비 페난 트렌 PARP1 억제제 또는 PARP1 결핍 세포 (- - /) 마우스 배아 섬유 아세포 (MEF) 즉, 일반 및 PARP1 ()의 활동을 비교 하였다. PARP1 결핍 MEF 항구 유사 분열의 다중 중심체,하지만 그들은 아무 있습니다t 종양 세포 11. 이러한 세포 박사 프랑소와 Dantzer, 프랑스 스트라스부르에 의해 제조 하였다.
(- / -) 일반 고정 및 PARP1 MEF는 등의 앞에보고 각각 자신의 스핀들 및 중심체를 표시 α-및 γ-tubulin의,에 대한 immunolabeled 하였다. 검사 세포 배양 중 일부는 ABT-888 및 AG01469, PARP1의 효소 활동을 억제하고, BSI-201 등 PJ-34이나 다른 강력한 비 페난 트렌 PARP1 억제제, 외관상으로 PARP1 바인딩 감쇠 화합물로 처리 하였다 DNA 12-14 새김. 농도 억제 PARP1 활동 (그림 4)에서 정상 MEF를 손상 테스트 PARP1 억제제 없음. 반면, PJ-34은 용량 의존적으로 PARP1에 γ-tubulin의 병소, 스핀들의 왜곡과 세포 죽음의 취소 클러스터링을 발생 (- / -) MEF는 (그림 4A 및 B). 이 PJ-34으로 처리 정상 MEF에서 관찰되지 않았다 (넘겨 준다E 4B) 또는 PARP1의 (- / -) MEF는 치료가 아닌 phenenthrene PARP1 억제제 ABT-888 또는 AG014699 (그림 4C). (- / -) MEF 그것은 정상 MEF는 PARP1보다 PJ-34 활동에 저항했지만 20 μM 이하의 농도에서 PJ-34 정상 MEF를 손상 않았다는 것을 주목해야한다.
사실 PJ-34 근절 PARP1보다 높은 농도 PJ-34으로 배양 MEF (- - /) 자신의 PARP1 부족하고, 다중 초점 스핀들의 형성과 PARP1의 셀 박멸의 상관 관계에도 불구하고 MEF (- - /) (- / -) MEF 및 PARP1 억제 (그림 4A) PARP1 억제에 필요한 사람들은, 추가 중심체 PARP1에 드 클러스터링 사이의 인과 결합과 일치하지 않았다. PARP1의 PJ-34의 세포 독성 (- /이 -) MEF는 더 나은 멀티 centrosomal 세포 2 여분 중심체가 드 클러스터링 에이전트 (로 그 활동에 의해 설명 될 수 기타 정보 "/> 그림 3. UN-클러스터 γ-tubulin에 표시된 여분 중심체와 유사 분열에서 무작위로 선택된 MDA-MB-231 세포에서 세포 죽음의 실시간 공 촛점 이미징 설명서를 참조하십시오. MDA-MB-231 세포를 스캔하기 전에 라이브 공 촛점 이미징 16 시간 동안 24 시간 동안 PJ-34으로 배양 하였다. PJ-34 γ-tubulin의-GFP (녹색, γ-tubulin의 병소 및 중심체)와 H2B-RED (빨강, 염색체)를 표현하는 벡터를 형질 전환 한 후 24 시간을 적용 하였다. 
그림 1. phenanthridine PJ-34 : N-(2 - 옥소-5, 6 - 디 하이드로-phenanthridin -2 - 일)-N, N-디메틸 아세트 아미드. 
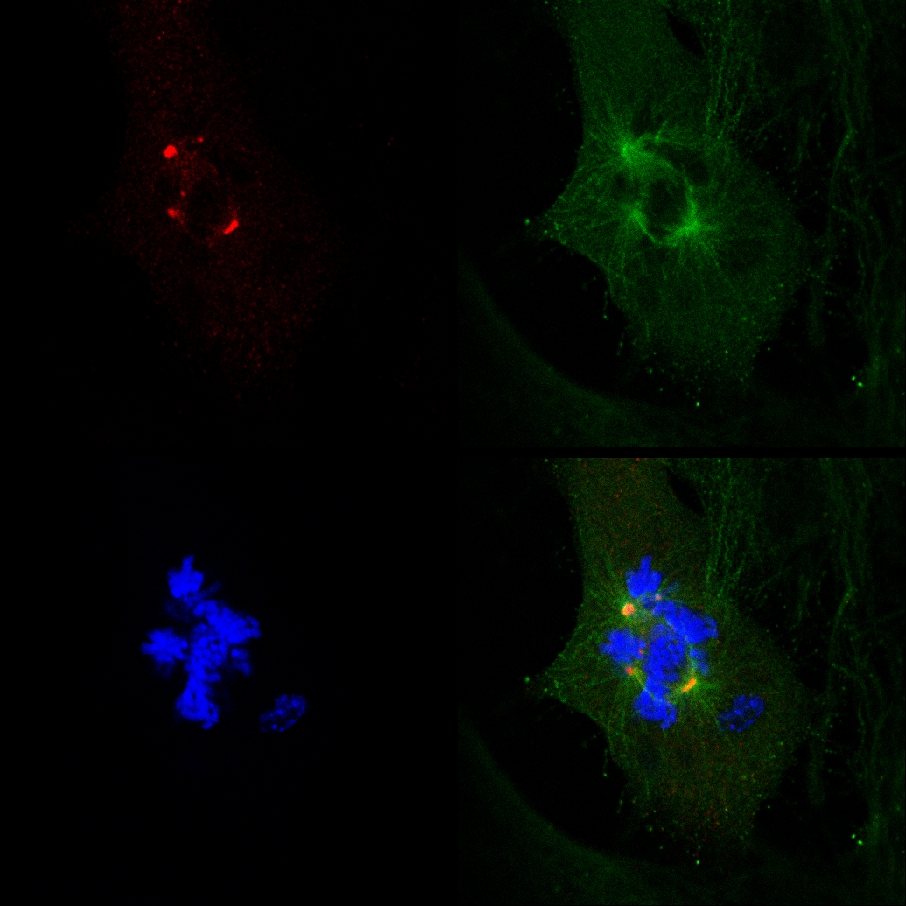
그림 2. 유사 분열에서 무작위로 선택된 라이브 MDA-MB-231 세포에있는 여분의 중심체의 이중 초점 클러스터링. A. 상단 패널 :. γ-tubulin에-GFP 낮은 패널 형질 무작위로 선택된 라이브 MDA-MB-231 세포에있는 레이블 중심체 : 히스톤 H2B-RED 형질 전환 무작위로 선택된 MDA-MB-231 세포에서 유사 분열 동안 염색체 재 배열. 무작위로 선택된 숭배에서 확인 클러스터 여분 중심체와 B. 이중 초점 유사 분열어인 MDA-MB-231 세포. 와 히스톤 H2B-RED (라벨 염색체, 빨간색) 세포는 모두 γ-tubulin의-GFP (녹색 표시 γ-tubulin의 병소)에 의해 형질 전환 하였다. 48 시간 형질 전환 후, 세포를 16 시간 동안 라이브 공 촛점 이미징에 노출되었다. 여섯 세포는 각각의 실험에 병렬로 스캔 하였다. 네 가지 실험을 수행 하였다. 보충 정보를 참조하십시오. 큰 그림을 보려면 여기를 클릭하십시오 . 
그림 3. 추가 중심체는 드 클러스터링 PJ-34으로 처리 라이브 MDA-MB-231 세포에서 세포 죽음을 앞에. 세포의 죽음으로 끝 흩어져 중심체 (왼쪽에 1 차 구조 ()와 유사 분열에서 무작위로 선택된 라이브 MDA-MB-231 세포 2 차 및 3 차 프레임). 이 CE및 (히스톤 H2B-RED, LL은 무작위로 γ-tubulin의-GFP (녹색 중심체를 포함하여 라벨 γ-tubulin의 병소)를 표현하는 벡터로 형질 전환 한 후 24 시간을 적용 PJ-34 (20 μM)를 24 시간 동안 배양 세포 배양에서 선택한 라벨 염색체, 빨간색). 셀은 라이브 공 촛점 이미징에 의해 16 시간 동안 검색되었습니다. 여섯 세포는 각각의 실험에 병렬로 스캔 하였다. 세 가지 실험을 수행 하였다. 보충 정보를 참조하십시오. 큰 그림을 보려면 여기를 클릭하십시오 . 
그림 4. PARP1의 PJ-34의 세포 독성 활동 (- / -) 마우스 배아 섬유 아세포. N에서 계산 다 초점 스핀들 A. (왼쪽) 비율ormal (블랙 라인) 및 PARP1 - / - 표시된 농도에서 PJ-34로 48 시간 동안 배양 (회색 선) MEF. 다극 스핀들의 비율은 3 가지 실험에서 검출 된 총 20 스핀들에서 계산되었다. (오른쪽) 제어 치료 세포의 생존에 비해 PJ-34 (20 μM)를 72 시간 동안 배양 세포 배양에서 검출 된 세포의 생존 감소 (- / - 정상 (블랙 라인) 및 PARP1을 (회색 선) MEF). 세포 생존은 세포의 ATP 생산 (프로토콜 5)에 의해 assayed되었다. 지정된 48 시간 동안 처리되지 않은 유사 분열에서 MEF (제어) 또는 PJ-34와 배양 -. 3 가지 실험에서 각각의 세포주 4 측정의 평균 값은 무작위로 선택된 고정 정상에있는 B. 스핀들을 제시하고 PARP1됩니다 - / 농도. PJ-34 다극 스핀들가 발생했습니다. 세포 고정 투과에 대해 immunolabeled 된 α-및 γ-tubulin의 (녹색 스핀들의 라벨 및 각각 중심체의 빨간색 표시). 염색체는 DAPI 시약 (파란색)으로 표시 하였다. 3. 다른 실험의 대표적인 결과 C. 유력한 비 페난 트렌 PARP1 억제제 PARP1의 중심체 클러스터링에 영향을 미치지 않았다 (- / -) MEF합니다. 무작위로 선택된 일반의 스핀들 및 PARP1는 - / - MEF가 제공되며, 치료 MEF (제어) 또는 MEF가 아닌 페난 트렌 PARP 억제제, AG01469 (20 μM) 또는 ABT888 (20 μM)를 48 시간 동안 처리. 염색체는 DAPI 시약 (파란색)로 표시되어 있습니다. 비슷한 결과는 3 가지 실험을 하였다. 큰 그림을 보려면 여기를 클릭하십시오 .
큰 그림을 보려면 여기를 클릭하십시오 . 
큰 그림을 보려면 여기를 클릭하십시오 .
그림 2B. 보충 그림. 이중 초점 클러스터 γ-tubulin의 여분 중심체가 γ-tubulin에-GFP를 발현 벡터로 형질 전환 된 후 라이브 공 촛점 이미징, 48 시간으로 16 시간 동안 스캔 한 표시와 함께 anaphase에서 무작위로 선택된 MDA-MB-231 세포 H2B-RED (라벨 γ-tubulin의 병소와 중심체 고정 세포 (녹색)와 염색체의 H2B 히스톤 라벨 (빨강), 각각).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
공 촛점 이미징 유사 분열 동안 라이브 멀티 centrosomal 세포 (그림 3 및 보충 정보)의 PJ-34의 세포 독성 효과를 실시간으로 문서를 제공 살고 있습니다. 이 PJ-34 5-9으로 유사 분열 재해 세포 사멸의 유도를 제안, 여분의 중심체 드 클러스터링 및 세포 죽음에 인간의 암 세포에있는 PJ-34의 세포 독성을 돌리는 최초의 살아있는 문서이다. 대조적으로, 최고 numerary 중심체의 이중 초점 클러스터링 양방향 focali 클러스터 추가 중심체 (그림 2, 보충 정보)와 일반 유사 분열을 겪고 라이브 치료 MDA-MB-231 세포에서 관찰되었다.
이러한 결과에 따르면, 형질 세포의 공 촛점 이미징 유사 분열 5,9,15,16 동안 형질 전환 세포의 여분 중심체 클러스터링에 연루 단백질의 전위를 감지하는 데 유용 할 수 있습니다 살고 있습니다. 멀티 centros에 PJ-34에 의해 영향을 단백질의 식별OMAL 세포는 여분 중심체 드 클러스터링에 의해 활성화 죽음의 메커니즘을 이해하기위한 몇 가지 단서를 제공 할 수 있습니다.
살아있는 세포의 유사 분열 동안 실시간으로 정보를 제공하는 라이브 공 촛점 이미징의 장점은 몇 가지 제한 사항에 바인딩됩니다. 실험 당 스캔 세포의 수는 제한되어 있습니다. 따라서 유사 분열에서 세포를 감지하는 기회가 부족하고, 여러 가지 반복 실험 스캔 세포의 유사 분열의 실시간 문서가 필요합니다. 또한, 성공 표시된 단백질을 발현 벡터를 세포의 형질 전환의 효율에 크게 의존합니다. 따라서, 신뢰성에도 불구하고, 라이브 공 촛점 이미징 시간이 소요됩니다 및 고도로 숙련 된 일꾼을 요구한다.
고정 된 세포의 비교, 면역 세포 및 공 촛점 이미징 신뢰할 수있는 통계 분석에 필요한 실험 당 세포의 큰 숫자의 검사 수 있습니다. 우리는을 비교하기 위해이 방법을 사용여분 중심체 2,3과 분열을 겪고있는 암 세포에 미치는 영향에 대한 일반적인 양성 세포의 유사 분열의 PJ-34의 효과. 마찬가지로,이 방법은 PARP의 비 페난 트렌 PARP1 억제제의에 PJ-34의 효과를 비교하기 위해 사용되었다 (- / -) MEF는 여분 중심체 (멀티 중심체 11 높은 발생과 세포) (그림 4) 숨겨.
요약하면, 우리의 결과는 cytochemistry 고정 된 세포의 공 촛점 분석과 유사 분열의 살아있는 세포 (그림 2와 3)의 공 촛점 이미징에서 제공하는 가장 중요한 정보를 실시간으로 결합의 이점을 나타냅니다. 이러한 방법의 조합 PJ-34와 같은 세포의 생존에 대한 구체적인 메커니즘은 매우 중요 대상, 작은 분자의 세포 독성 활동을 식별하는 데 유용 할 수 있습니다. 자신의 증식과 surviv 중 하나에 대한 여분 중심체 양극 클러스터링에 많은 인간의 암 세포의 독특한 의존알은 PJ-34 암 치료에 대한 가능한 후보를 렌더링합니다.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
저자는 그들이 더 경쟁 재정적 이익이 없다는 것을 선언합니다.
Acknowledgments
이 연구의 자금 출처 : 텔 아비브 대학의 기술 이전 회사 라모트와 시바 - 메디컬 센터 (M. CA 및 SI.) ICRF의 공동 기금 - 이스라엘 암 연구 재단 (M. CA.)와 이스라엘 과학 재단 ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}