

Summary
De cytotoxische activiteit van de fenanthridine PJ-34 in kankercellen ondergaan mitose werd in real time gedocumenteerd door levende confocale beeldvorming. PJ-34 uitgeroeide menselijke borstkanker MDA-MB-231 cellen die extra-centrosomes in mitose. In tegenstelling tot normale bifocale mitose, werden de extra-centrosomes niet geclusterd in de twee polen spil in aanwezigheid van PJ-34.
Abstract
Fenantreen derivaten die als krachtige PARP1 remmers voorkomen de bifocale clustering van boventallige centrosomes in multi-centrosomal menselijke kankercellen in de mitose. De phenanthridine PJ-34 was de meest potente molecule. Ontclustering van extra-centrosomes veroorzaakt mitotische falen en celdood in multi-centrosomal cellen. Meest solide menselijke kankers hebben hoge voorkomen van extra-centrosomes. De activiteit van PJ-34 werd in real-time gedocumenteerd door confocale beeldvorming van levende menselijke borstkanker MDA-MB-231-cellen getransfecteerd met vectoren die coderen voor fluorescente γ-tubuline, dat veel voorkomt in de centrosomes en fluorescerende histone H2b aanwezig de chromosomen. Afwijkende chromosomen regelingen en de-geclusterde γ-tubuline foci die declustered centrosomes werden gedetecteerd in de getransfecteerde MDA-MB-231-cellen na behandeling met PJ-34. Un-geclusterde extra-centrosomes in de twee polen spindel voorafgegaan hun celdood. Deze resultaten gekoppeld for het eerst onlangs ontdekt exclusieve cytotoxische activiteit van PJ-34 in humane kankercellen met extra centrosomes de-clustering in mitose en mitotische storing die tot celdood. Volgens eerdere bevindingen waargenomen door confocale beeldvorming van vaste cellen, PJ-34 uitsluitend uitgeroeid kankercellen met multi-centrosomes zonder afbreuk normale cellen ondergaan mitose met twee centrosomes en bi-focale spindels. Deze cytotoxische activiteit van PJ-34 werd niet gedeeld door andere krachtige PARP1 remmers, en werd waargenomen in PARP1 deficiënte MEF herbergen extracentrosomes, suggereert haar onafhankelijkheid van PARP1 remming. Wonen confocale beeldvorming bood een nuttig instrument voor het identificeren van nieuwe moleculen uitroeien van cellen tijdens mitose.
Introduction
Fenantreen afgeleid PARP1 remmers, waaronder PJ-34, werden ontworpen om latente cellen van apoptotische celdood geïnduceerd door de energieverbruikende PARP1 gemedieerde DNA-reparatie onder stress-omstandigheden (beroerte of myocardinfarct) te beschermen 1. Echter, recent ontdekten we dat PJ-34, twee keer hogere concentratie dan dat inducerende PARP1 remming, kan uitsluitend leiden tot celdood in menselijke kankercellen 2,3. De snellere van de proliferatie van de cel was, hoe efficiënter de uitroeiing van de cellen was. De cytotoxische activiteit van de PJ-34 werd toegeschreven aan extra-centrosomes de-clustering in mitose 2. Veel menselijke kankercellen haven multicentrosomes 4,5. Incubatie van menselijke borstkankercellen MDA-MB-231, die boventallige centrosomen haven, met 20 uM PJ-34 efficiënt uitgeroeid deze cellen binnen 72-96 uur zonder afbreuk rustende cellen of een goedaardige delende cellen die twee centrosomes in mitose
Bipolaire centrosome montage is cruciaal voor bipolaire vorming spil in mitose 4,5. Daarom hebben cellen met meer dan twee centrosomes een nauwelijks begrepen moleculaire mechanisme ontwikkeld clustering hun extra centrosomes tweepolig 4-9. Falen van een bipolaire assemblage van hun centrosomes kan multipolaire vervormde spindels en afwijkende chromosomen segregatie dat de arrestaties de cel-cyclus in de G2 / M arrestatie, en leidt tot celdood toegeschreven aan mitotische mislukking 4,5 veroorzaken. De moleculaire mechanismen die ten grondslag liggen aan extra-centrosomes de-clustering worden intensief onderzocht <sup> 10. Inzicht in dit mechanisme dood zal exclusieve uitroeiing van kankercellen in staat stellen terwijl sparen gezonde weefsels 5,10.
Aldus verbindingen die mitotische catastrofe celdood activeren bieden een nieuwe manier van een selectieve kankertherapie, die efficiënt kan in een breed spectrum van menselijke vaste cancers.Our resultaten suggereren dat confocale beeldvorming kan worden gebruikt om moleculen die extra-centrosomes clustering worden geïdentificeerd mitose 2,3, waardoor deze verbindingen kanker targeting kandidaat-geneesmiddelen.
We hebben de cytotoxische activiteit van de fenanthridine PJ-34 gedocumenteerd door het scannen van vaste en levende menselijke kankercellen (met een hoog voorkomen van extra-centrosomes in mitose) versus normale cellen. Een stapsgewijze beschrijving van de beeldvormende procedures voor de cytotoxische activiteit van PJ-34 in menselijke kankercellen te identificeren is hieronder opgenomen.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Cel Cultuur Voorbereiding
MDA-MB-231-cellen werden gekocht bij ATCC (American Type Culture Collection) en opgeslagen in vloeibare stikstof.
- Seed 10 6 MDA-MB-231-cellen in 92 mm Petri-schaal in 10 ml compleet medium bevattende Dulbeco Modified Eagle Medium (DMEM), 10% paardenserum, 1% L-glutamine en 1% Penstrep-Amfotericine B. Laat cellen te prolifereren tot 80-100% confluentie.
- Verwijder kweekmedium uit schotel en gooi.
- Spoel even de cellaag met 0,25% (w / v) Trypsine-EDTA-oplossing om alle sporen van serum te verwijderen.
- Voeg 2,0 ml trypsine-EDTA-oplossing voor te schotelen en te observeren cellen door omgekeerde microscoop totdat cellaag wordt verspreid (meestal binnen 5 tot 15 min).
- Voeg 18 ml compleet groeimedium Zuig de cellen door pipet. Transfer naar een buis.
- Centrifugeer de celsuspensie bij 1200 rpm.
- Resuspendeer de cel pellet in 24 ml culture medium.
- Voeg 2 ml celsuspensie tot 35 mm glazen bodem gerechten (ongeveer 25% confluentie) en plaats in de incubator (5% CO 2, 37 ° C).
- Oplossingen:
- Compleet Medium voor celproliferatie: DMEM met 10% FBS, 1% antibiotica (100 eenheden / ml penicilline G, 100 ug / ml streptomycine, Pen-Strep-Ampho oplossing) en 2 mM L-glutamine.
- Trypsine-EDTA-oplossing die 0,25% trypsine-EDTA.
- Gerechten:
92 mm diameter petrischalen.
35 mm diameter poly-D-lysine gecoate glazen bodem cultuur gerechten.
2. Voorbereiding van de Cellen voor Live confocale Imaging
- Zaad 2 x 10 5 MDA-MB-231 cellen in glazen bodem cultuur gerechten in 2 ml compleet medium zoals vermeld in paragraaf 1. Wanneer celcultuur samenvloeiing van 60-70% (ongeveer 3-4 x 10 5 cellen per schaaltje) bereikt, verder met transfectie.
- Transfecteren de cellen met twee plasmiden encoding van de fusie-eiwitten γ-tubuline-GFP (voor fluorescentie detectie van centrosomes) en histon-RED (H2b-RED, voor fluorescerende detectie van chromosomen) met behulp van de liposomaal transfectiereagens Jet-PI, na de vervaardiging protocol. Kort mix 2 ug van elk plasmide in een buis met 100 ul NaCl (150 mM). Meng de transfectie reagens (100 pl) met 100 ul NaCl (150 mM) in een tweede buis en incubeer 5 min bij kamertemperatuur (RT). Dan combineer de twee oplossingen, mix (met een milde vortex) en spin-down. Incubeer gedurende 30 minuten bij kamertemperatuur.
- Tijdens de incubatie van de transfectie mengsel, was de cellen eenmaal met PBS en vervang het celmedium met 2 ml warm DMEM zonder supplementen (37 ° C).
- Voorzichtig voeg de transfectie aan de cellen in DMEM en vervolgens terug de cellen aan de incubator (37 ° C, 5% CO2) gedurende 8 uur.
- Na 8 uur incubatie vervangen DMEM met 2 ml compleet medium en incubeer de cellen in de incubator gedurende 24hr.
- 24 uur na transfectie vervangen medium van de cellen met 2 ml compleet medium dat 20 uM PJ-34.
- Incubeer de cellen gedurende nog 18 uur (37 ° C, 5% CO2).
- Behoudens de cellen confocale beeldweergave ten minste 16 uur in beeldkamer houden van de cellen in 5% CO2 en 37 ° C.
- Parallel, onderzoekt transfectie werkzaamheid 36 uur na transfectie met behulp van fluorescentie microscopie als volgt:
- Seed 2 x 10 5 MDA-MB-231 cellen in 6-wells plaat met 1 dekglaasje per putje in 2 ml compleet medium.
- Transfecteren de cellen zoals vermeld in paragrafen 2.2-2.5.
- 36 uur na transfectie stellen de getransfecteerde cellen gemonteerd op een dekglaasje door incubatie in koude methanol: aceton (1:1) oplossing, 7 min, -20 ° C.
- Zuig het fixatie-oplossing en laat het dekglaasje met de gemonteerde cellen te drogen in een chemische kap.
- Solliciteer ProLong Gold antifade reagens met DAPI en laat stee dekglaasje te drogen in het donker gedurende 6 uur.
- Onderzoek de schuif onder fluorescentiemicroscoop en bereken het percentage van de getransfecteerde cellen (rode en groene signalen) van de totale populatie van cellen (DNA kleuring door DAPI). De gewenste transfectie levert dat ongeveer 20-40% bij 100-200 cellen geteld.
3. Technische parameters van de Live confocale Imaging Scanner Instellingen
- ScanMode xyzt; Pinhole [luchtige] 1.00; Zoom 3,5; Resolutie 8 bits; Laser DPSS 561 nm; Argon, zichtbare laser 488 nm; Laser Hij / Ne zichtbaar 633 nm; Doelstelling HCX PL APO CS 63X 1,40 OIL UV; Numerieke diafragma 1.4; Scan snelheid 700 Hz; Brekingsindex 1.52.
- Afbeelding 3-D presentatie werden bereid door Imaris imaging software 7.0.
4. Confocale Imaging van mitose in vaste cellen
- Zaad 2 x 10 5 MDA-MB-231 borstkankercellen (ATCC), normale muis embryonale fibroblasten (MEF), of PARP1 gebrekkige MEF (PARP - / -, bereid door Dr Francoise Dantzer) op glazen dekglaasjes in 6-well plaat in 2 ml compleet medium. Dekglaasjes werden gewassen met 96% ethanol, gevolgd door wassen met steriel DD water, gedroogd gedurende 2 uur, en geplaatst in elk putje van de 6-well schotel.
- Voeg PJ-34 (10-30 uM) aan het medium en incubeer de cellen gedurende de vereiste periode (meestal maximaal 96 uur).
- Was de dekglaasjes eenmaal met PBS (fosfaat gebufferde zoutoplossing), en zet de cellen middels incubatie in ijs-koude methanol: aceton (1:1) oplossing, 7 min, -20 ° C.
- Zuig het fixatie-oplossing en laat de dekglaasjes te drogen in chemische hood (in dit stadium, de dekglaasjes worden op -20 ° C gedurende enkele weken bewaard).
- Was de dekglaasjes eenmaal met PBST (PBS aangevuld met 0,1% Tween-20) aan de celmembranen doorlaatbaar en blokkeren de cellen met 10% NDS (normaal Ezel Serum) in PBST (blokkerende oplossing) gedurende 1 uur bij kamertemperatuur.
- Incubeer de gepermeabiliseerde vaste cellen met primaire antibodies gedurende 2 uur bij kamertemperatuur (voor spillen centrosomes kleuring). De antilichamen worden verdund in de blockingbuffer als volgt: anti-α tubuline (1:250 verdunning) en anti-γ tubuline (1:200 verdunning). Primaire antilichamen worden als volgt toegepast: toepassing 100 pl (een druppel) van een mengsel van de antilichamen in blokkerende oplossing per dekglaasje op 6-plaatafdekking (het deksel ondersteboven). Voorzichtig zet de dekglaasje op de antilichamen druppel, gezaaide cellen tegenover de druppel. Incubeer de dekglaasjes naar de antilichamen gedurende 2 uur bij kamertemperatuur geroerd.
- Plaats de dekglaasjes terug in de putjes en spoel de cellen 3 maal met PBST. Maak dan gebruik van dezelfde procedure in 4.6 beschreven voor het labelen van cellen op de dekglaasjes met de fluorescerende secundaire antilichamen. Incubeer de cellen op dekglaasjes met de secundaire antilichamen gedurende 1 uur, kamertemperatuur, in het donker. De antilichamen worden verdund in het blokkeren van oplossing als volgt: Alexa Fluor 488 (1:1000 verdunning, groen) en Alexa Fluor 568 (1:1000 verdunning;red).
- Monteer de dekglaasjes met ProLong Gold antifade reagens met DAPI (voor chromosomen kleuring) en incubeer overnacht bij kamertemperatuur in het donker te drogen.
- Onderzoekt de dekglaasjes door confocale microscopie.
5. Levensvatbaarheid van de cellen gemeten door ATP Production
ATP productie wordt gemeten door een luminescerende ATP detectietest kit.
- Zaad de cellen in 96-well plaat, ongeveer 20.000 cellen in 800 ul medium in elk putje. Drie blanco's moeten worden gebruikt voor de bepaling van de achtergrond luminescentie van het medium.
- Bereid ATP standaard verdunningsreeks van ongeveer 10 uM tot 100 uM en blijf op ijs.
- Voeg 50 ul van detergent aan elk putje en schud de plaat gedurende 5 min in orbitaalschudder, 700 rpm.
- Elke injectieflacon van de 'gevriesdroogde substraat' met 5 ml 'substraatbuffer' in de kit.
- Voeg 50 ul van de bereide substraatoplossing aan de putjes en schudde plaat gedurende 5 minuten op orbitaalschudder, 700 rpm.
- Houd de plaat in het donker gedurende 10 minuten.
- Meet luminescentiesignalen van elk putje met ELISA microplaatlezer.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 is een stabiele water oplosbare fenantridine 1 (figuur 1). Onze eerdere resultaten lieten celdood en de-geclusterde extra-centrosomes in verschillende soorten vaste multi-centrosomal kankercellen die behandeld werden met PJ-34. Daarentegen werden de normale delende cellen niet aangetast 2,3. Centrosomes werden geïdentificeerd door dubbele etikettering met antilichamen gericht tegen centrine1 en γ-tubuline in de vaste extra-centrosomal cellen 2.
Hier, de cytotoxische activiteit van de PJ-34 werd gedocumenteerd in deze live-extra-centrosomal cellen in real-time met behulp van live confocale microscopie. Levende menselijke borstkanker MDA-MB-231 cellen, die een hoog optreden (> 50%) van de extra-centrosomes 4,5 hebben, werden gescand gedurende ten minste 16 uur door confocale beeldvorming gericht op getransfecteerde cellen met γ-tubuline-GFP ( fluorescente labeling van γ-tubuline foci 2) en met histon H2b-RED (tl Labeling van chromosomen). Zes tot tien levende getransfecteerde cellen werden gescand in parallel in elk experiment. Dubbele immunokleuring van de γ-tubuline brandpunten in de getransfecteerde cellen met centrin1 was technisch onmogelijk.
Gedispergeerd γ-tubuline foci afwijkende chromosomen arrangement werden zelden waargenomen in willekeurig geselecteerde onbehandelde MDA-MB-231 cellen in mitose. Bifocale clustering van γ-tubuline foci, ofwel extra-centrosomes bifocale clustering, is gedocumenteerd in de meeste levende behandelde MDA-MB-231-cellen (Figuur 2), daarentegen, niet-geclusterde centrosomes en afwijkende opstelling van chromosomen in levende gedetecteerd getransfecteerde MDA-MB-231-cellen geïncubeerd met PJ-34 (20 pM) en mitose in deze cellen werd beëindigd door celdood (Figuur 3). Deze cellen werden geïncubeerd met PJ-34 18 - 24 uur voor scannen en extra 16 uur tijdens het scannen (Figuur 3). De real-time documentatie van de celath tijdens mitose steunt een vooraf gedefinieerde positieve correlatie tussen het aantal menselijke kwaadaardige cellen met multipolaire spindles in mitose en het percentage celdood in cellen geïncubeerd met PJ-34 (20 pM) 2.
PJ-34 werkt als een potente remmer PARP1 1. Daarom hebben we de mogelijkheid PARP1 inhibitie onderzocht waardoor celdood geassocieerd met mitotische falen. In tegenstelling tot het, beeldvorming van vaste MDA-MB-231-cellen geactiveerd onderzoek van een grote populatie van cellen in de celkweken, waardoor statistische analyse van de effecten van remmers PARP1 in verschillende humane kankercellijnen. De activiteit van PJ-34 werd vergeleken met de activiteit van andere krachtige, niet-fenantreen PARP1 remmers in normale of PARP1 deficiënte cellen (normale en de PARP1 (- / -) muis embryonale fibroblasten (MEF)) (Figuur 4). PARP1 deficiënte MEF haven meerdere centrosomes in mitose, maar ze zijn niett tumorcellen 11. Deze cellen werden bereid door Dr Francoise Dantzer, Straatsburg, Frankrijk.
Vaste normaal en PARP1 (- / -) MEF werden immunologisch voor α en γ-tubuline hun spillen centrosomes gelabeld, respectievelijk, zoals gerapporteerd voor 2. Enkele onderzochte celkweken werden behandeld met 34 PJ-of andere krachtige, niet-fenantreen PARP1 remmers, waaronder ABT-888 en AG01469, die de enzymatische activiteit van PARP1 remmen, en BSI-201, een verbinding die schijnbaar verzwakt PARP1 bindend gejat DNA 12-14. Geen van de geteste remmers PARP1 geschaad normale MEF bij concentraties PARP1 remmende activiteit (figuur 4). Daarentegen PJ-34 dosisafhankelijk veroorzaakt un-clustering van γ-tubuline foci, verstoring van spillen celdood in PARP1 (- / -) MEF (figuren 4A en B). Dit werd niet waargenomen bij normale MEF behandeld met PJ-34 (Figure 4B) of PARP1 (- / -) MEF behandeld met niet-phenenthrene PARP1 remmers ABT-888 of AG014699 (Figuur 4C). Opgemerkt wordt dat PJ-34 bij concentraties hoger dan 20 uM heeft aantasten normale MEF, hoewel normale MEF waren resistent tegen PJ-34 activiteit dan PARP1 (- / -) MEF.
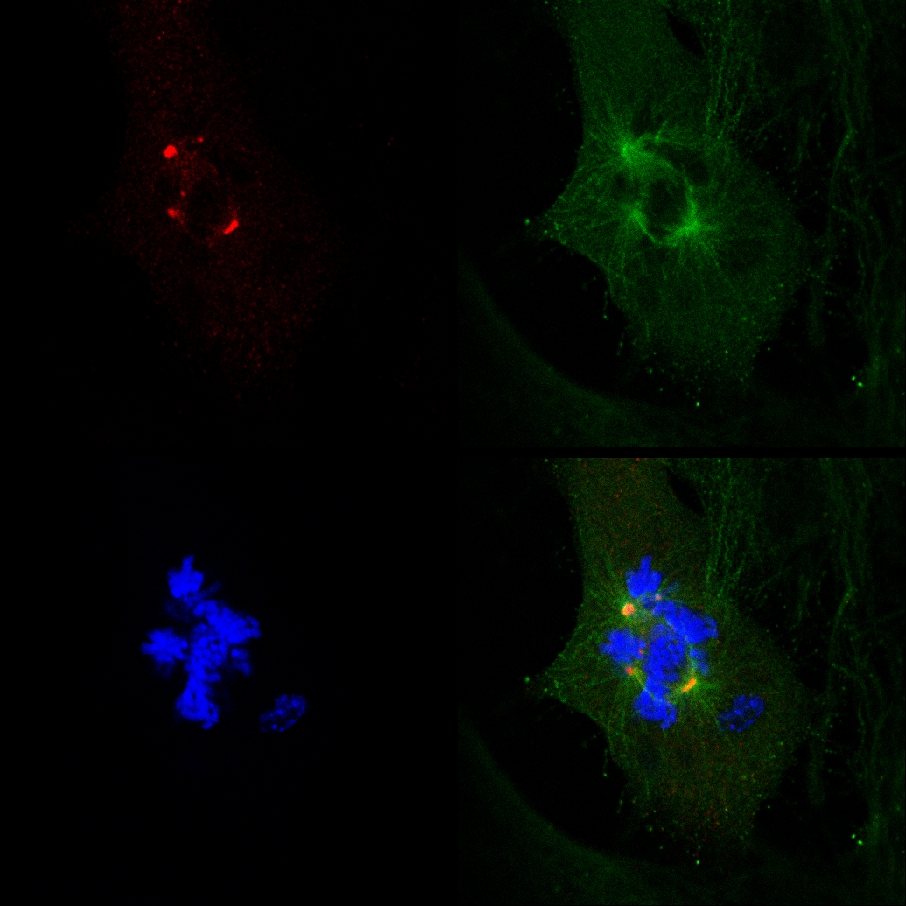
Dat PJ-34 uitgeroeid PARP1 (- / -) MEF ondanks hun PARP1 deficiëntie, en de correlatie tussen de vorming van een multi-focale spillen cel uitroeiing PARP1 (- / -) MEF geïncubeerd met PJ-34 bij concentraties hoger dan die welke aan PARP1 remming waren consistent met een causaal verband tussen extra-centrosomes de-clustering in PARP1 (- / -) en MEF PARP1 remming (figuur 4A). De cytotoxische activiteit van PJ-34 in PARP1 (- / -) MEF beter kan worden verklaard door zijn activiteit als een extra-centrosomes de-clustering middel in multi-centrosomal cellen 2 ( Aanvullende informatie "/> Figuur 3. Een live confocale beeldvorming documentatie van celdood in een willekeurig geselecteerde MDA-MB-231 cellen in mitose met niet-geclusterde γ-tubuline gelabelde extra-centrosomes. MDA-MB-231-cellen werden geïncubeerd met PJ-34 gedurende 24 uur voor het scannen en tijdens de 16 uur van levende confocale beeldvorming. PJ-34 werd aangebracht 24 uur na transfectie met vectoren die γ-tubuline-GFP (green, γ-tubuline foci centrosomes) en H2b-RED (rood, chromosomen). 
Figuur 1. De phenanthridine PJ-34: N-(6-oxo-5 ,6-dihydro-fenantridine-2-yl)-N, N-dimethyl-aceetamide. 
Figuur 2. Bifocale clustering van extra-centrosomes in een willekeurig geselecteerde live-MDA-MB-231 cellen in mitose. A. Bovenste paneel: Labeled centrosomes in een willekeurig geselecteerde live-MDA-MB-231 cellen getransfecteerd met γ-tubuline-GFP Onderste paneel:. Chromosoom herschikkingen tijdens de mitose in een willekeurig geselecteerde MDA-MB-231 cellen getransfecteerd met histon H2b-RED. B. Bi-focale mitose met geclusterde extra-centrosomes geïdentificeerd in een willekeurig gekozen cultfigureerde MDA-MB-231 cellen. Cellen werden getransfecteerd door zowel γ-tubuline-GFP (labeling γ-tubuline foei, groen) en histon H2b-RED (labelen van chromosomen; red). 48 uur na transfectie werden cellen blootgesteld aan een live confocale beeldvorming voor 16 uur. Zes cellen werden gescand in parallel in elk experiment. Vier verschillende experimenten werden uitgevoerd. Zie ook aanvullende informatie. Klik hier om een grotere afbeelding te bekijken . 
Figuur 3. Extra-centrosomes de-clustering voorafgegaan celdood in levende MDA-MB-231 cellen behandeld met PJ-34. Een willekeurig geselecteerde live-MDA-MB-231 cellen in mitose met verspreide centrosomes (1 st kader links) eindigde door celdood ( 2 e en 3 e frames). Dit cell werd willekeurig gekozen in een celcultuur gedurende 24 uur met PJ-34 (20 uM) toegepast 24 uur na transfectie met vectoren die γ-tubuline-GFP (labeling γ-tubuline brandpunten waaronder centrosomes, groen) en histon H2b-RED ( labelen van chromosomen; red). De cel werd gescand voor 16 uur per live-confocale beeldvorming. Zes cellen werden gescand in parallel in elk experiment. Drie verschillende experimenten werden uitgevoerd. Zie ook aanvullende informatie. Klik hier om een grotere afbeelding te bekijken . 
Figuur 4. Een cytotoxische activiteit van PJ-34 in PARP1 (- / -) muis embryonale fibroblasten. A. (links) Het percentage van multi-focale spindels berekend normale (zwarte lijn) en PARP1 - / - (grijze lijn) MEF, gedurende 48 uur met PJ-34 bij de aangegeven concentraties. Het percentage multipolaire spindels werd berekend uit 20 in totaal spindels gedetecteerd in 3 verschillende experimenten. (Rechts) Verminderde overleving van cellen gedetecteerd in celkweken gedurende 72 uur met PJ-34 (20 uM) met betrekking tot de overleving van onbehandelde cellen (normale (zwarte lijn) en PARP1 - / - (grijze lijn) MEF). Celoverleving werd getest door de ATP-productie van de cellen '(protocol 5). De gemiddelde waarden van 4 metingen voor elke cellijn in 3 verschillende experimenten worden weergegeven in B. spindels willekeurig gekozen vast normaal en PARP1 -. / - MEF in mitose, onbehandelde (controle) of geïncubeerd met PJ-34 gedurende 48 uur bij de aangegeven concentraties. PJ-34 veroorzaakte multipolaire spindels. Cellen werden vastgesteld, gepermeabiliseerd en immunolabeled voor α-en γ-tubuline (groen etikettering van spindels en rode etikettering van centrosomes, respectievelijk). Chromosomen werden gelabeld met DAPI reagens (blauw). . Representatieve resultaten van 3 verschillende experimenten C. potente niet-fenantreen PARP1 remmers hadden geen invloed centrosomes clustering in PARP1 (- / -) MEF. Spindels van willekeurig geselecteerde normaal en PARP1 - / - MEF worden gepresenteerd; onbehandeld MEF (controle) of MEF behandeld voor 48 uur met niet-fenantreen PARP-remmers, AG01469 (20 uM) of ABT888 (20 uM). Chromosomen zijn gelabeld met DAPI reagens (blauw). Vergelijkbare resultaten werden verkregen in 3 verschillende experimenten. Klik hier om een grotere afbeelding te bekijken .
Klik hier om een grotere afbeelding te bekijken . 
Klik hier om een grotere afbeelding te bekijken .
Figuur 2B. Supplemental Figuur. Een willekeurig geselecteerde MDA-MB-231 cellen in anafase met bifocale geclusterde γ-gelabeld tubuline extra-centrosomes werd gescand op 16 uur via live confocale beeldvorming, 48 uur na getransfecteerd met vectoren die γ-tubuline-GFP en H2B-RED (labeling γ-tubuline foci centrosomes in de gefixeerde cellen (groen) en H2b histon etikettering van de chromosomen (rood), respectievelijk).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Wonen confocale beeldvorming voorzien van een real-time documentatie van het cytotoxische effect van PJ-34 in levende multi-centrosomal cellen tijdens mitose (Figuur 3 en aanvullende informatie). Dit was de eerste levende documentatie toeschrijven de cytotoxiciteit van PJ-34 in menselijke kankercellen extra centrosomes de-clustering en celdood, suggereert inductie van mitotische catastrofe celdood door PJ-34 5-9. Daarentegen werd bifocale clustering van super-numerary centrosomes waargenomen in levende onbehandelde MDA-MB-231 cellen die normale mitose met bi-focali geclusterde extra centrosomes (figuur 2, aanvullende informatie).
Volgens deze resultaten, live confocale beeldvorming van getransfecteerde cellen kunnen nuttig zijn voor het detecteren van translocatie van eiwitten betrokken in extra-centrosomes clustering in getransfecteerde cellen tijdens mitose 5,9,15,16 zijn. Identificatie van eiwitten die door PJ-34 in multi-centrosomal cellen kan enige aanwijzingen geven voor het begrijpen van de dood mechanismen geactiveerd door extra-centrosomes de-clustering.
Het voordeel van levende confocale beeldvorming in real-time informatie tijdens mitose in levende cellen, is ook gebonden aan verschillende beperkingen. Het aantal cellen per gescande experiment beperkt. Daarom kansen om cellen te detecteren in mitose zijn laag, en een aantal herhalende experimenten nodig zijn voor een real-time documentatie van mitose in de gescande cellen. Bovendien succes is sterk afhankelijk hoge efficiëntie van transfectie van cellen met vectoren die gemerkte eiwitten. Dus, ondanks dat betrouwbaar, leef confocale beeldvorming is tijdrovend en vraagt zeer ervaren harde werkers.
In vergelijking, immunocytochemie en confocale beeldvorming van gefixeerde cellen mogelijk onderzoek van een groot aantal cellen per experiment, die nodig is voor een betrouwbare statistische analyse. We gebruikten deze methode het vergelijkeneffecten van PJ-34 in mitose van normale goedaardige cellen om de gevolgen ervan in kankercellen ondergaan mitose met boventallige centrosomes 2,3. Evenzo werd de methode voor het vergelijken van de effecten van PJ-34 tot die van niet-fenantreen PARP1 PARP remmers (- / -) MEF harboring extra centrosomes (cellen met hoog aantal meervoudige centrosomes 11) (Figuur 4).
Samengevat, onze resultaten tonen het voordeel van het combineren van de meest waardevolle real-time informatie van confocale beeldvorming van levende cellen in mitose (figuren 2 en 3) met cytochemie en confocale analyse van gefixeerde cellen. Een combinatie van deze methoden kunnen nuttig om de cytotoxische activiteit van kleine moleculen die, zoals PJ-34, gericht specifieke mechanismen cruciaal voor celoverleving zijn. De unieke afhankelijkheid van vele menselijke kankercellen op extra-centrosomes bi-polaire clustering voor hun proliferatie en survival maakt PJ-34 een mogelijke kandidaat voor kankertherapie.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs verklaren dat zij geen concurrerende financiële belangen.
Acknowledgments
Financieringsbronnen van dit onderzoek: een gezamenlijk fonds van de Universiteit van Tel Aviv overdracht van technologie bedrijf, RAMOT en het Sheba Medical Center (M. CA en SI.), ICRF - Israëlische Cancer Research Foundation (M. CA.) En Israel Science Foundation ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}