

Summary
L'attività citotossica della fenantridina PJ-34 in cellule tumorali in mitosi è stata documentata in tempo reale dai confocale vivo. PJ-34 sradicato cancro al seno umano MDA-MB-231 cellule che ospitano in più-centrosomi nella mitosi. A differenza della normale mitosi bi-focale, l'extra-centrosomi non sono stati raggruppati in due poli del fuso in presenza di PJ-34.
Abstract
Derivati Fenantrene che agiscono come potenti inibitori PARP1 impedito il raggruppamento bi-focale di centrosomi soprannumerari nelle celle multi-centrosomica tumorali umane in mitosi. Il fenantridina PJ-34 è stata la più potente molecola. Declustering di extra-centrosomi provoca insufficienza mitotico e la morte cellulare nelle cellule multi-centrosomica. Tumori umani più solidi hanno alta presenza di extra-centrosomi. L'attività di PJ-34 è stata documentata in tempo reale da imaging confocale di cancro al seno umano vivo MDA-MB-231 cellule trasfettate con vettori codificanti per fluorescente γ-tubulina, che è altamente abbondante nei cntrosome e per fluorescente istone H2b presente in i cromosomi. Cromosomi aberranti arrangiamenti e de cluster-foci γ-tubulina che rappresentano centrosomi declustered sono stati rilevati nelle transfettate cellule MDA-MB-231 dopo il trattamento con PJ-34. Un-cluster in più-centrosomi nei due poli del fuso preceduto la loro morte cellulare. Questi risultati legati for la prima volta l'attività citotossica esclusivo recentemente rilevata del PJ-34 in cellule tumorali umane con supplemento cntrosome-de-clustering in mitosi e insufficienza mitotico che porta alla morte cellulare. Secondo i risultati precedenti osservati con l'imaging confocale di cellule fisse, PJ-34 cellule tumorali debellata esclusivamente con il multi-centrosomi senza danneggiare le cellule normali subiscono mitosi con due centrosomi e mandrini bi-focale. Questa attività citotossica di PJ-34 non è stata condivisa da altri potenti inibitori PARP1, ed è stata osservata in PARP1 deficienti MEF extracentrosomes ospitano, suggerendo la sua indipendenza di PARP1 inibizione. Vivere confocale offerto uno strumento utile per l'individuazione di nuove molecole sradicare le cellule durante la mitosi.
Introduction
Fenantrene deriva inibitori PARP1, tra cui PJ-34, sono stati progettati per proteggere le cellule quiescenti di morte cellulare per apoptosi indotta dal consumo di energia PARP1 mediata riparazione del DNA in condizioni di stress (ictus o infarto del miocardio) 1. Tuttavia, di recente abbiamo scoperto che PJ-34, a concentrazione due volte più alta di quella che induce PARP1 inibizione, può causare la morte delle cellule esclusivamente in cellule tumorali umane 2,3. Più rapida è la proliferazione della cellula era, più efficiente l'eradicazione delle cellule era. L'attività citotossica di PJ-34 è stato attribuito a extra-centrosomi de-clustering in mitosi 2. Molte cellule tumorali porto umano multicentrosomes 4,5. Incubazione di cellule umane di cancro al seno MDA-MB-231, che ospitano centrosomi soprannumero, con 20 micron PJ-34 efficientemente debellate queste cellule entro 72-96 ore senza danneggiare le cellule quiescenti o alcuni proliferativi benigni ospitare due centrosomi in mitosi
Bipolare assemblea centrosoma è cruciale per la formazione del fuso bipolare in mitosi 4,5. Pertanto, le cellule con più di due centrosomi hanno sviluppato un meccanismo molecolare poco capito, raggruppando i loro centrosomi ai due poli 4-9. Fallimento di montaggio bipolare dei loro centrosomi può causare multipolare distorto mandrini e aberrante cromosomi segregazione che arresta il ciclo cellulare in G2 / M arresto, e porta alla morte cellulare attribuite al fallimento mitotico 4,5. I meccanismi molecolari alla base di olio extra-centrosomi de-cluster sono intensamente studiate <sup> 10. La comprensione di questo meccanismo di morte consentirà eradicazione esclusivo delle cellule tumorali risparmiando i tessuti sani 5,10.
Così, composti che attivano la morte cellulare mitotico catastrofe offrono una nuova modalità di una terapia del cancro selettivo, che può essere efficace in una vasta gamma di risultati cancers.Our solidi umani suggeriscono che l'imaging confocale può essere utilizzato per identificare molecole che influenzano extra-cntrosome clustering in mitosi 2,3, rendendo questi composti cancro mira candidati farmaci.
Abbiamo documentato l'attività citotossica del fenantridina PJ-34 tramite la scansione cellule tumorali umane fisse e vivo (con alta presenza di extra-centrosomi in mitosi) contro le cellule normali. Un passo-passo le procedure di imaging utilizzate per identificare l'attività citotossica di PJ-34 in cellule tumorali umane di seguito è incluso.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Cell Culture Preparazione
MDA-MB-231 cellule sono state acquistate da ATCC (American Type Culture Collection) e conservati in azoto liquido.
- Seed 10 6 MDA-MB-231 cellule nel diametro 92 mm piastra di Petri in 10 ml di terreno completo contenente Dulbeco Modified Eagle Medium (DMEM), 10% siero di cavallo, 1% di L-glutammina, e 1% Penstrep-amfotericina B. Consentono alle cellule di proliferare a circa 80-100% di confluenza.
- Togliere terreno di coltura dal piatto e scartare.
- Lavare lo strato cella brevemente con 0,25% (w / v) Tripsina-EDTA soluzione per eliminare ogni traccia di siero.
- Aggiungere 2,0 ml di soluzione di tripsina-EDTA per piatto e osservare le cellule al microscopio invertito fino strato di cellule viene dispersa (di solito entro 5-15 min).
- Aggiungere 18 ml di terreno di crescita completo e aspirare delicatamente le cellule dalla pipetta. Trasferire in una provetta.
- Centrifugare la sospensione cellulare a 1.200 giri al minuto.
- Risospendere il pellet cellulare in 24 ml di culture medie.
- Aggiungere 2 ml di sospensione cellulare a 35 millimetri di vetro piatti inferiori (circa il 25% di confluenza) e posto in incubatore (5% di CO 2, 37 ° C).
- Soluzioni:
- Terreno completo per la proliferazione delle cellule: DMEM con 10% FBS, 1% Antibiotici (100 unità / ml di penicillina G, 100 mg / ml di streptomicina, soluzione Pen-Strep-Ampho) e 2 mM L-glutammina.
- Soluzione di tripsina-EDTA, contenente 0,25% tripsina-EDTA.
- Piatti:
Diametro 92 mm Petri.
35 mm Diametro poli-D-lisina rivestite con fondo di vetro piatti della cultura.
2. Preparazione di cellule per Live confocale
- Seed 2 x 10 5 MDA-MB-231 cellule nel fondo di vetro piatti cultura in 2 ml di terreno completo come indicato nella sezione 1. Quando la coltura cellulare raggiunge confluenza del 60-70% (circa 3-4 x 10 5 cellule per piatto), procedere con la trasfezione.
- Trasfezione le cellule con due plasmidi ENCODzione delle proteine di fusione γ-tubulina-GFP (per la rilevazione fluorescente di centrosomi) e istone-RED (H2b-RED, per la rilevazione fluorescente dei cromosomi) utilizzando il reagente di trasfezione liposomiale Jet-PI, seguendo il protocollo di produzione. Brevemente, mescolare 2 ug di ciascun plasmide in un tubo con 100 microlitri di NaCl (150 mM). Mescolare il reagente di trasfezione (100 pl) con 100 microlitri di NaCl (150 mM) in un secondo tubo, e incubare 5 min a temperatura ambiente (RT). Poi unire le due soluzioni, miscela (con vortice lieve) e spin-down. Incubare per 30 min a RT.
- Durante l'incubazione della miscela di trasfezione, lavare le cellule una volta con PBS e sostituire il mezzo cella con 2 ml di DMEM caldo senza integratori (37 ° C).
- Aggiungere delicatamente la miscela di trasfezione per le cellule in DMEM e poi tornare celle verso l'incubatore (37 ° C, 5% CO 2) per 8 ore.
- Dopo 8 ore di incubazione, sostituire il DMEM con 2 ml di mezzo completo e incubare le cellule in incubatore per 24hr.
- 24 ore dopo la trasfezione, sostituire il medium delle cellule con 2 ml di terreno completo contenente 20 mM PJ-34.
- Incubare le cellule per ulteriori 18 ore (37 ° C, 5% CO 2).
- Oggetto le cellule a vivere confocale per almeno 16 ore in camera di imaging mantenere le cellule al 5% di CO 2 e 37 ° C.
- In parallelo, esaminare l'efficacia di trasfezione 36 ore dopo la trasfezione usando la microscopia a fluorescenza come segue:
- Seme 2 x 10 5 MDA-MB-231 cellule in piastra da 6 pozzetti contenente 1 coprioggetto per pozzetto in 2 ml di mezzo completo.
- Trasfezione le cellule come accennato nelle sezioni 2,2-2,5.
- 36 ore dopo la trasfezione fissare le cellule transfettate montati su un vetrino da incubazione in metanolo freddo: acetone (1:1) soluzione, 7 min, -20 ° C.
- Aspirare la soluzione di fissaggio e lasciare che il vetrino con le cellule montati a secco in una cappa chimica.
- Applicare Prolungare reagente antifade Oro con DAPI e lasciate °e vetrino ad asciugare al buio per 6 ore.
- Esaminare il vetrino al microscopio a fluorescenza e di calcolare la percentuale di cellule trasfettate (segnali rosso e verde) della popolazione totale di cellule (colorazione del DNA con DAPI). La percentuale desiderata transfezione è circa 20-40% 100-200 quando le cellule sono contate.
3. Parametri tecnici delle impostazioni dello scanner confocale in diretta
- ScanMode XYZT; Pinhole [arioso] 1,00; Zoom 3,5; Risoluzione 8 bit; laser DPSS 561 nm; argon, laser visibile a 488 nm laser He / Ne visibile 633 nm; Obiettivo HCX PL APO CS 63X 1.40 OLIO UV; Apertura numerica 1.4; Velocità di scansione 700 Hz; indice di rifrazione 1.52.
- Immagine 3-D di presentazione sono stati preparati da Imaris di imaging software 7.0.
4. Confocale di mitosi in cellule fissate
- Seed 2 x 10 5 MDA-MB-231 delle cellule di cancro al seno (ATCC), normali fibroblasti embrionali di topo (MEF), o PARP1 carente MEF (PARP - / -, preparato dal dottor Francoise Dantzer) su vetrini in 6 pozzetti in 2 ml di terreno completo. Coprioggetto sono stati lavati con etanolo al 96%, in seguito da lavaggio con acqua sterile DD, asciugata per 2 ore, e collocato in ciascun pozzetto di piatto 6-bene.
- Aggiungere PJ-34 (10-30 mM) al mezzo e incubare le cellule per il periodo necessario (di solito fino a 96 ore).
- Lavare i coprioggetti una volta con PBS (tampone fosfato), e fissare le cellule utilizzando incubazione in metanolo ghiacciata: (1:1) soluzione di acetone, 7 min, -20 ° C.
- Aspirare la soluzione di fissaggio e lasciare che i coprioggetti ad asciugare in cappa chimica (in questa fase, i vetrini possono essere conservati a -20 ° C per diverse settimane).
- Lavare i coprioggetti una volta con PBST (PBS supplementato con 0,1% di Tween-20) per permeabilize le membrane cellulari e di bloccare le cellule con 10% NDS (asino siero normale) in PBST ('soluzione bloccante') per 1 ora a RT.
- Incubare le cellule fisse permeabilizzate con Antibodi primariaes per 2 ore a temperatura ambiente (per i mandrini e centrosomi colorazione). Gli anticorpi sono diluiti nella soluzione bloccante come seguono: tubulina anti-α (diluizione 1:250) e anti-tubulina γ (diluizione 1:200). Anticorpi primari sono applicati come segue: applicare 100 microlitri (in una goccia) di una miscela di anticorpi in soluzione per ogni coprioggetto bloccaggio sulla copertina 6 pozzetti (il coperchio è capovolto). Mettere delicatamente il vetrino sulla goccia anticorpi, le cellule seminate di fronte alla caduta. Incubare i coprioggetti rivolti gli anticorpi per 2 ore a temperatura ambiente.
- Posizionare i coprioggetti indietro nei pozzetti e lavare le cellule 3 volte con PBST. Quindi utilizzare la stessa procedura descritta in 4.6 per etichettare le cellule sui vetrini con gli anticorpi secondari fluorescenti. Incubare le cellule sui vetrini con gli anticorpi secondari per 1 ora, RT, al buio. Gli anticorpi sono diluiti in soluzione di blocco come segue: Alexa Fluor 488 (diluizione 1:1000; verde) e Alexa Fluor 568 (diluizione 1:1000;rosso).
- Montare il coprioggetto utilizzando prolungare reagente antifade Oro con DAPI (per i cromosomi colorazione) e incubare una notte a temperatura ambiente al buio per asciugare.
- Esaminare i vetrini al microscopio confocale.
5. La vitalità cellulare Misurato da produzione di ATP
Produzione di ATP è misurata da un kit di analisi ATP rilevamento luminescente.
- Seed le cellule in piastra a 96 pozzetti, circa 20.000 cellule in 800 microlitri media in ciascun pozzetto. Tre pozzi vuoti dovrebbero essere utilizzati per la determinazione dello sfondo luminescenza del mezzo.
- Preparare una serie di diluizioni standard di ATP da circa 10 micron a 100 micron e tenere in ghiaccio.
- Aggiungere 50 ml di detersivo per ogni pozzetto e agitare la piastra per 5 minuti in agitatore orbitale, 700 giri.
- Ricostituire ciascun flacone di 'sostrato liofilizzato' con 5 ml di 'tampone substrato' nel kit.
- Aggiungere 50 ml della soluzione di substrato ricostituito ai pozzetti, e agitarela piastra per 5 minuti su agitatore orbitale, 700 giri.
- Tenere la piastra al buio per 10 minuti.
- Misurare luminescenza di ogni bene da ELISA lettore di micropiastre.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 è un acqua stabile solubile fenantridina 1 (Figura 1). I nostri risultati precedenti hanno rivelato la morte cellulare e la de-raggruppati extra-centrosomi in diversi tipi di cellule tumorali a più centrosomica fissi che sono stati trattati con PJ-34. Al contrario, le cellule proliferanti normali non hanno perso di valore 2,3. Centrosomi sono stati identificati mediante doppia marcatura con anticorpi diretti contro centrine1 e γ-tubulina nelle cellule fisse extra-centrosomica 2.
Qui, l'attività citotossica di PJ-34 è stata documentata in queste cellule extra-centrosomica vivo in tempo reale utilizzando la microscopia confocale in diretta. Vivere il cancro al seno umano MDA-MB-231 cellule, che hanno una elevata incidenza (> 50%) di extra-centrosomi 4,5, sono stati esaminati per almeno 16 ore per l'imaging confocale focalizzata sulle cellule transfettate con γ-tubulina-GFP ( etichettatura fluorescente di γ-tubulina foci 2) e con istone H2b-RED (fluorescente labeling di cromosomi). Da sei a dieci cellule transfettate vivo vengono scansionate in parallelo in ogni esperimento. Doppio immunomarcatura dei fuochi γ-tubulina nelle cellule trasfettate con centrin1 era tecnicamente impossibile.
Dispersi γ-tubulina foci e cromosomi aberranti disposizione sono stati raramente rilevati in caso selezionati non trattati MDA-MB-231 cellule in mitosi. Raggruppamento bifocale di foci γ-tubulina, che rappresentano in più-centrosomi raggruppamento bifocale, è stata documentata nella maggior parte dei vivi non trattati MDA-MB-231 cellule (Figura 2), invece, centrosomi e disposizione aberrante di cromosomi non-cluster sono stati individuati in diretta trasfettate MDA-MB-231 cellule incubate con PJ-34 (20 mM), e mitosi in queste cellule cessati dalla morte cellulare (Figura 3). Queste cellule sono state incubate con PJ-34 per 18 - 24 ore prima della scansione e per ulteriori 16 ore durante la scansione (Figura 3). La documentazione tempo reale di cellula death durante la mitosi sostiene fortemente una correlazione positiva precedentemente definito tra il numero di cellule maligne umane con mandrini multipolare, in mitosi e la percentuale di morte cellulare in cellule incubate con PJ-34 (20 mM) 2.
PJ-34 agisce come un potente inibitore PARP1 1. Abbiamo quindi esaminato la possibilità di PARP1 inibizione causando la morte cellulare associata a insufficienza mitotico. Diversamente imaging dal vivo, imaging fissi cellule MDA-MB-231 abilitato esame di una vasta popolazione di cellule nelle colture cellulari, consenta di effettuare un'analisi statistica degli effetti di inibitori PARP1 in una varietà di linee cellulari tumorali umane. L'attività di PJ-34 è stato confrontato con l'attività di altri potenti, inibitori non-fenantrene PARP1 in normale o PARP1 cellule deficienti (cioè normale e PARP1 (- / -) fibroblasti embrionali di topo (MEF)) (Figura 4). PARP1 carenti MEF porto di multi-centrosomi in mitosi, ma non sonocellule tumorali t 11. Queste cellule sono state preparate dal dottor Francoise Dantzer, Strasburgo, Francia.
Fisso normale e PARP1 (- / -) MEF sono stati immunolabeled per α-e γ-tubulina che etichettato loro fusi e centrosomi, rispettivamente, come riportato prima 2. Alcune delle colture cellulari esaminati sono stati trattati con potenti inibitori, PJ-34 o di altri non-fenantrene PARP1, tra ABT-888 e AG01469, che inibiscono l'attività enzimatica di PARP1, e BSI-201, un composto che apparentemente attenua PARP1 vincolante per scalfito DNA 12-14. Nessuno degli inibitori testati PARP1 deteriorate MEF normale a concentrazioni PARP1 inibizione dell'attività (Figura 4). Al contrario, PJ-34 dose-dipendente causato un-raggruppamento di γ-tubulina foci, distorsione dei mandrini e la morte cellulare in PARP1 (- / -) MEF (Figure 4A e B). Questo non è stato osservato in MEF normale trattato con PJ-34 (Figure 4B) o in PARP1 (- / -) MEF trattato con non phenenthrene PARP1 inibitori ABT-888 o AG014699 (Figura 4C). Va notato che PJ-34 a concentrazioni superiori 20 uM ha compromesso MEF normale, sebbene MEF normale erano più resistenti a PJ-34 attività rispetto PARP1 (- / -) MEF.
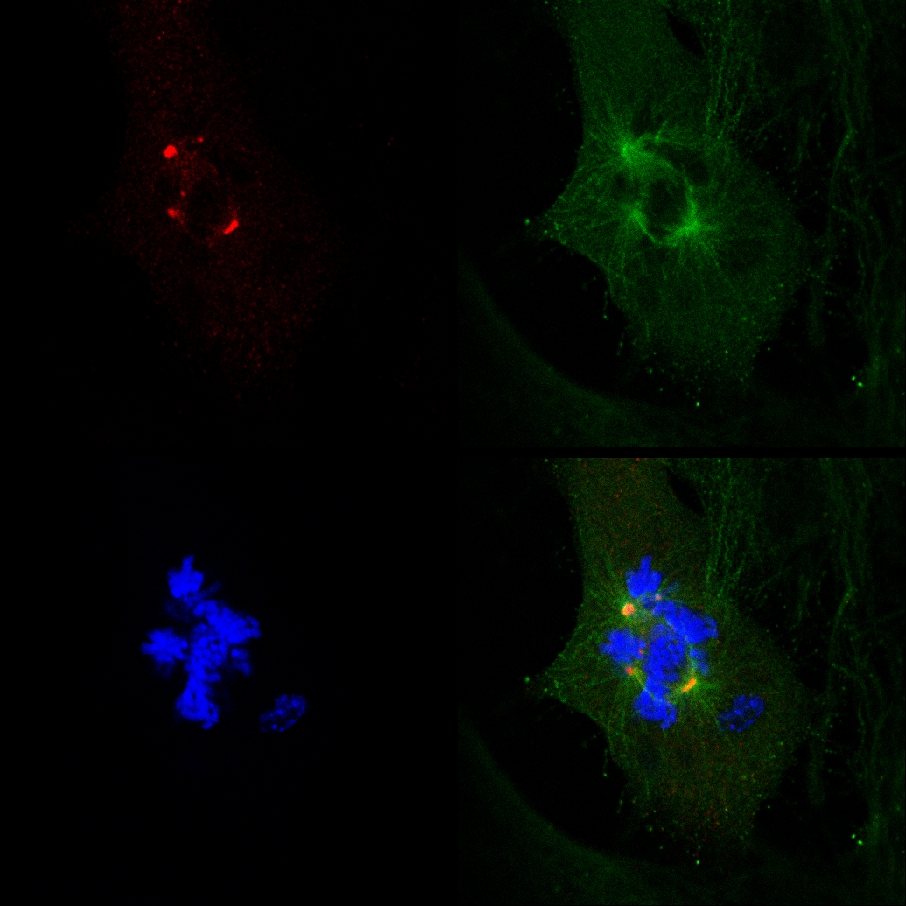
Il fatto che PJ-34 sradicato PARP1 (- / -) MEF nonostante la loro PARP1 deficienza, e la correlazione tra la formazione di mandrini multifocale ed eradicazione delle cellule in PARP1 (- / -) MEF incubate con PJ-34 a concentrazioni superiori quelli richiesti per PARP1 inibizione, non erano coerenti con un legame causale tra più centrosomi-de-clustering in PARP1 (- / -) MEF e PARP1 inibizione (Figura 4A). L'attività citotossica di PJ-34 in PARP1 (- / -) MEF potrebbe essere meglio spiegato con la sua attività come un extra-centrosomi agente di de-clustering in celle multi-centrosomica 2 ( Informazioni supplementari "/> Figura 3. Una documentazione di imaging confocale in diretta della morte cellulare in una scelta a caso MDA-MB-231 delle cellule in mitosi con un-cluster γ-tubulina etichettati in più-centrosomi. MDA-MB-231 cellule sono state incubate con PJ-34 per 24 ore prima della scansione e durante la 16 ore di confocale vivo. PJ-34 è stato applicato 24 ore dopo la trasfezione con vettori che esprimono γ-tubulina-GFP (Green, foci γ-tubulina e centrosomi) e H2b-RED (rosso, cromosomi). 
Figura 1. Il fenantridina PJ-34: N-(6-oxo-5 ,6-diidro-phenanthridin-2-il)-N, N-dimetil-acetammide. 
Figura 2. Clustering di Bi-focale di extra-centrosomi in una scelta a caso dal vivo MDA-MB-231 delle cellule in mitosi. A. Pannello superiore: centrosomi etichettato in un live MDA-MB-231 delle cellule selezionato casualmente transfettate con γ-tubulina-GFP Pannello inferiore:. Cromosomiche riorganizzazioni durante la mitosi in una scelta a caso MDA-MB-231 delle cellule trasfettate con istone H2b-RED. B. mitosi Bi-focale con cluster in più-centrosomi individuati in una setta scelto a casorato MDA-MB-231 delle cellule. Le cellule sono state trasfettate sia γ-tubulina-GFP (etichettatura γ-tubulina foci; verde) e istone H2b-RED (etichettatura cromosomi; rossa). 48 ore dopo la trasfezione, le cellule sono state esposte a un confocale vivo per 16 ore. Sei celle vengono scansionate in parallelo in ogni esperimento. Sono stati eseguiti quattro esperimenti diversi. Vedi anche informazioni supplementari. Clicca qui per ingrandire la figura . 
Figura 3. Extra-centrosomi de-cluster preceduto la morte delle cellule in vivo MDA-MB-231 cellule trattate con PJ-34. Una scelta a caso dal vivo MDA-MB-231 delle cellule in mitosi con centrosomi sparsi (1 ° fotogramma a sinistra) si è conclusa con la morte delle cellule ( 2 ° e 3 ° frame). Questo cell è stato selezionato in modo casuale in una coltura di cellule incubate per 24 ore con PJ-34 (20 micron) applicato 24 ore dopo la trasfezione con vettori che esprimono γ-tubulina-GFP (etichettatura foci γ-tubulina tra centrosomi, verde) e istone H2b-RED ( etichettatura cromosomi, rosso). La cella è stata scansionata per 16 ore per l'imaging confocale vivo. Sei celle vengono scansionate in parallelo in ogni esperimento. Sono stati eseguiti tre esperimenti diversi. Vedi anche informazioni supplementari. Clicca qui per ingrandire la figura . 
Figura 4. Una attività citotossica di PJ-34 in PARP1 (- / -) fibroblasti embrionali di topo. A. (a sinistra) La percentuale di mandrini multifocale calcolato in nnormale (linea nera) e PARP1 - / - (linea grigia) MEF, incubate per 48 ore con PJ-34 alle concentrazioni indicate. La percentuale di mandrini multipolare è stato calcolato su 20 in totale mandrini rilevato in 3 diversi esperimenti. (Destra) Ridotta sopravvivenza cellulare individuato in colture cellulari incubate per 72 ore con PJ-34 (20 micron) rispetto alla sopravvivenza delle cellule non trattate di controllo (normale linea (nero) e PARP1 - / - (linea grigia) MEF). La sopravvivenza delle cellule è stata valutata mediante la produzione di ATP le cellule '(protocollo 5). I valori medi di quattro misure per ciascuna linea cellulare in 3 diversi esperimenti sono presentati in B. Mandrini selezionato casualmente fisso normale e PARP1 -. / - MEF nella mitosi, non trattati (controllo) o incubati con PJ-34 per 48 ore al indicata concentrazioni. PJ-34 ha causato multipolare mandrini. Le cellule sono state fissate, permeabilizzate e immunolabeled per α-e γ-tubulina (etichettatura verde di fusi e etichettatura rosso di centrosomi, rispettivamente). I cromosomi sono stati etichettati con il reagente DAPI (blu). . Risultati rappresentativi di tre esperimenti diversi C. I potenti inibitori non-fenantrene PARP1 non ha influenzato centrosomi clustering in PARP1 (- / -) MEF. Mandrini di selezionati in modo casuale normale e PARP1 - / - MEF vengono presentati; MEF non trattati (controllo) o MEF trattate per 48 ore con inibitori di PARP non fenantrene, AG01469 (20 mM) o ABT888 (20 micron). I cromosomi sono etichettati con il reagente DAPI (blu). Risultati simili sono stati ottenuti in 3 diversi esperimenti. Clicca qui per ingrandire la figura .
Clicca qui per ingrandire la figura . 
Clicca qui per ingrandire la figura .
Figura 2B. Figura supplementare. Una scelta a caso MDA-MB-231 delle cellule in anafase con bi-focale cluster γ-tubulina etichettati extra-centrosomi stati scansionati per 16 ore per l'imaging confocale in diretta, 48 ore dopo essere state trasfettate con vettori che esprimono γ-tubulina-GFP e H2b-RED (etichettatura foci γ-tubulina e centrosomi nelle cellule fisse (verde) e l'etichettatura istone H2b dei cromosomi (rosso), rispettivamente).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
In diretta confocale ha fornito una documentazione in tempo reale l'effetto citotossico di PJ-34 in cellule multi-centrosomica dal vivo durante la mitosi (figura 3 e le informazioni supplementari). Questa è stata la prima documentazione vivo attribuendo la citotossicità di PJ-34 in cellule tumorali umane di centrosomi de-clustering e di morte cellulare, suggerendo l'induzione di morte cellulare mitotica Catastrophe da PJ-34 5-9. Al contrario, il clustering bi-focale di centrosomi super-numerari è stata osservata in vivo non trattati MDA-MB-231 cellule in fase di mitosi normale con bi-focali cluster centrosomi (Figura 2, le informazioni supplementari).
In base a questi risultati, vivere l'imaging confocale di cellule transfettate potrebbe essere utile per individuare la traslocazione di proteine coinvolte in extra-centrosomi clustering in cellule trasfettate durante la mitosi 5,9,15,16. Identificazione delle proteine interessate da PJ-34 in multi-Centroscellule Omal possono fornire alcuni indizi per la comprensione dei meccanismi di morte attivati da più centrosomi-de-clustering.
Il vantaggio di confocale vivo nel fornire informazioni in tempo reale durante la mitosi nelle cellule vive, è legata anche a molte limitazioni. Il numero di cellule analizzati al esperimento è limitato. Pertanto possibilità di rilevare le cellule in mitosi sono bassi, e diversi esperimenti ripetuti sono necessari per una documentazione tempo reale della mitosi nelle cellule digitalizzati. Inoltre, il successo dipende molto elevata efficienza di trasfezione di cellule con vettori esprimenti proteine marcate. Così, pur essendo affidabile, confocale vivo richiede tempo e richiede grandi lavoratori di grande esperienza.
In confronto, immunocitochimica e imaging confocale di cellule fissate consentire l'esame di un gran numero di cellule per esperimento, che è richiesto per un'analisi statistica affidabile. Abbiamo usato questo metodo per confrontare laeffetti di PJ-34 in mitosi delle normali cellule benigne ai suoi effetti sulle cellule tumorali in mitosi con centrosomi soprannumerari 2,3. Analogamente, questo metodo è stato utilizzato per confrontare gli effetti di PJ-34 a quella di inibitori non-fenantrene PARP1 in PARP (- / -) MEF ospitare supplementare-cntrosome (cellule con alta incidenza di multi-cntrosome 11) (Figura 4).
In sintesi, i nostri risultati indicano il vantaggio di combinare il più prezioso tempo reale informazioni fornite da imaging confocale di cellule vive in mitosi (figure 2 e 3) con citochimica ed analisi confocale di cellule fissate. Una combinazione di questi metodi può essere utile per identificare l'attività citotossica di piccole molecole che, come PJ-34, Obiettivo meccanismi specifici cruciale per la sopravvivenza cellulare. La dipendenza unica di molte cellule tumorali umane in extra-centrosomi clustering di bi-polare per la loro proliferazione e survivabilityAl rende PJ-34 un possibile candidato per la terapia del cancro.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Gli autori dichiarano di non avere interessi finanziari in competizione.
Acknowledgments
Le fonti di finanziamento di questa ricerca: un fondo comune di società dell'Università di Tel Aviv, il trasferimento tecnologico, RAMOT e-Sheba Medical Center (M. CA e SI.), ICRF - israeliana Cancer Research Foundation (M. CA.) E Israel Science Foundation ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}