

Summary
Den cytotoksiske aktiviteten til phenanthridine PJ-34 i kreftceller som gjennomgår mitose ble dokumentert i sanntid ved konfokal levende avbildning. PJ-34 utryddet menneskelig brystkreft MDA-MB-231 celler som ekstra-centrosomes i mitose. I motsetning til normal bi-fokale mitose, ble de ekstra-centrosomes ikke gruppert i de to spindel polene i nærvær av PJ-34.
Abstract
Fenantren derivater som fungerer som potente PARP1 hemmere hindret bi-fokale clustering av overtallige centrosomes i multi-centrosomal humane kreftceller i mitose. Den phenanthridine PJ-34 var den mest potente molekylet. Declustering av ekstra-centrosomes fører mitotisk svikt og celledød i multi-centrosomal celler. Mest solide kreft hos mennesker har høy forekomst av ekstra-centrosomes. Aktiviteten av PJ-34 ble dokumentert i sanntid ved confocal avbildning av levende menneske brystkreft MDA-MB-231 celler transfektert med vektorer som koder for fluorescerende γ-tubulin, som er svært rik på de centrosomes og for fluorescerende histone H2b stede i kromosomene. Avvikende kromosomer ordninger og de-klynger γ-tubulin brennpunkter representerer declustered centrosomes ble oppdaget i transfekterte MDA-MB-231 celler etter behandling med PJ-34. Un-gruppert ekstra-centrosomes i de to spindel polene innledet sin celledød. Disse resultatene knyttet for første gang nylig oppdaget eksklusive cytotoksisk aktivitet av PJ-34 i humane kreftceller med ekstra-centrosomes de-clustering i mitose, og mitotic svikt fører til celledød. Ifølge tidligere funn observert ved confocal avbildning av faste celler, PJ-34 utelukkende utryddet kreftceller med multi-centrosomes uten å svekke normale celler gjennomgår mitose med to centrosomes og bi-fokale spindler. Dette cytotoksisk aktivitet av PJ-34 ble ikke delt av andre potente PARP1 hemmere, og ble observert i PARP1 mangelfulle MEF skjuler extracentrosomes, noe som tyder sin uavhengighet av PARP1 hemming. Leve confocal bildebehandling tilbudt et nyttig verktøy for å identifisere nye molekyler utrydde celler under mitose.
Introduction
Fenantren avledet PARP1 hemmere, inkludert PJ-34 ble utviklet for å beskytte hvilende celler fra celledød ved apoptose indusert av energikrevende PARP1 mediert DNA-reparasjon under stress forhold (slag eller hjerteinfarkt) 1. Men nylig oppdaget vi at PJ-34, på to ganger høyere konsentrasjon enn det inducing PARP1 hemming, kan utelukkende føre til celledød i humane kreftceller 2,3. Jo mer hurtig spredning av cellen var, jo mer effektiv utryddelse av cellene. Cellegiften aktiviteten til PJ-34 ble tilskrevet ekstra-centrosomes de-clustering i mitose to. Mange menneskelig kreft celler havn multicentrosomes 4,5. Inkubasjon av menneskelig brystkreft celler MDA-MB-231, som havna overtallige centrosomes, med 20 mikrometer PJ-34 effektivt utryddet disse cellene innen 72-96 timer uten å svekke hvilende celler eller noen godartede prolifererende celler som to centrosomes i mitose
Bipolar sentrosomen montering er avgjørende for bipolar spindel formasjonen i mitose 4,5. Derfor har celler med mer enn to centrosomes utviklet et knapt forstått molekylære mekanismen, clustering sine ekstra centrosomes på to poler 4-9. Unnlatelse av bipolar montering av sine centrosomes kan forårsake multipolar forvrengt spindler og avvikende kromosomer segregering at arrestasjoner celle-syklus i G2 / M arrest, og fører til celledød tilskrives mitotic svikt 4,5. De molekylære mekanismene bak ekstra-centrosomes de-clustering er intensivt undersøkt <sup> 10. Forstå dette dødsfallet mekanismen vil gjøre eksklusive utrydding av kreftceller mens sparsom sunt vev 5,10.
Således kan forbindelser som aktiverer mitotisk katastrofe celledød tilbyr en ny måte av en selektiv cancer terapi, som kan være effektive i et bredt spekter av humane solide cancers.Our resultater tyder på at konfokal avbildning kan anvendes for å identifisere molekyler som påvirker ekstra-centrosomes clustering i 2,3 mitose, rendering disse forbindelser kreft målretting narkotika kandidater.
Vi har dokumentert den cytotoksiske aktiviteten til phenanthridine PJ-34 ved å skanne faste og levende humane kreftceller (med høy forekomst av ekstra-centrosomes i mitose) versus normale celler. En trinnvis beskrivelse av imaging prosedyrer som brukes til å identifisere den cytotoksiske aktiviteten til PJ-34 i humane kreftceller er inkludert nedenfor.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
En. Cell Culture Forberedelse
MDA-MB-231-celler ble kjøpt fra ATCC (American Type Culture Collection) og lagret i flytende nitrogen.
- Seed 10 6 MDA-MB-231 celler i 92 mm diameter petriskål i 10 ml komplett medium som inneholder Dulbeco Modified Eagle Medium (DMEM), 10% hest serum, 1% L-glutamin, og 1% Penstrep-Amfotericin B. Tillat celler å spre seg til ca 80-100% samløpet.
- Fjern kultur medium fra fatet og kast.
- Vask celleskiktet kort med 0,25% (w / v) Trypsin-EDTA-oppløsning for å fjerne alle spor av serum.
- Legg 2,0 ml trypsin-EDTA-oppløsning for å antenne og observere cellene ved invertert mikroskop inntil celle laget dispergeres (vanligvis i løpet av 5 til 15 min.)
- Legg 18 ml komplett vekstmedium og forsiktig aspireres cellene med pipette. Overføring til et rør.
- Sentrifuger cellesuspensjonen ved 1.200 rpm.
- Re-suspendere cellen pellet i 24 ml cuLTURE medium.
- Tilsett 2 ml celle suspensjon til 35 mm glassbunn retter (ca 25% samløpet) og plasseres i inkubatoren (5% CO 2, 37 ° C).
- Løsninger:
- Komplett Medium for celler spredning: DMEM med 10% FBS, 1% Antibiotika (100 enheter / ml penicillin G, 100 mikrogram / ml streptomycin, Pen-streptokokk-Ampho løsning) og 2 mM L-glutamin.
- Trypsin-EDTA-løsning, som inneholdt 0,25% Trypsin-EDTA.
- Retter:
92 mm diameter petriskåler.
35 mm diameter poly-D-lysin belagt glassbunn kultur retter.
2. Utarbeidelse av celler for Live Confocal Imaging
- Seed 2 x 10 5 MDA-MB-231 celler i glassbunn kultur retter i 2 ml komplett medium som nevnt i § 1. Når cellekultur når samløpet av 60-70% (ca 3-4 x 10 5 celler per parabol), fortsette med transfeksjon.
- Transfektere celler med to plasmider ENCODing fusjon proteiner γ-tubulin-GFP (for fluorescerende påvisning av centrosomes) og Histone-RED (H2b-RED, for fluorescerende påvisning av kromosomer) med liposomale transfeksjon reagens Jet-PI, som følge av produksjon protokollen. Kort, bland 2 ug fra hver plasmid i et rør med 100 ul NaCl (150 mM). Bland transfeksjon reagens (100 ul) med 100 ul NaCl (150 mM) i et andre rør, og inkuber 5 min ved romtemperatur (RT). Deretter kombinere de to løsningene, bland (med mild vortex) og spinn-ned. Inkuber i 30 minutter ved RT.
- Under inkuberingen av den transfeksjon blandingen, vaskes cellene en gang med PBS og erstatte cellen medium med 2 ml varmt DMEM med ingen kosttilskudd (37 ° C).
- Tilsett transfeksjon blandingen til cellene i DMEM og deretter returnere cellene til inkubatoren (37 ° C, 5% CO2) i 8 timer.
- Etter 8 timers inkubasjon, erstatte DMEM med 2 ml komplett medium og inkuberes cellene i inkubator i 24hr.
- 24 timer etter transfeksjon, erstatte mediet av cellene med 2 ml fullstendig medium inneholdende 20 uM PJ-34.
- Inkuber cellene for ytterligere 18 timer (37 ° C, 5% CO2).
- Emne cellene til å leve konfokal avbildning i minst 16 timer i avbildning kammer holde cellene ved 5% CO2 og 37 ° C.
- Parallelt undersøke transfeksjon effekt 36 hr post transfeksjon med fluoriserende mikroskopi som følger:
- Seed 2 x 10 5 MDA-MB-231 celler i 6-brønn plate som inneholder en dekkglass per brønn i 2 ml komplett medium.
- Transfektere cellene som nevnt i pkt. 2.2 til 2.5.
- 36 timers post transfeksjon fikse transfekterte celler montert på et dekkglass ved inkubasjon i kald metanol: aceton (1:1)-løsning, 7 min, -20 ° C.
- Sug fiksering løsning og la dekkglass med de monterte celler til tørk i en kjemisk hette.
- Påfør forlenge Gold antifade reagens med DAPI og la the dekkglass for å tørke i mørke i 6 timer.
- Undersøke raset under fluorescerende mikroskop og beregne hvor stor andel av de transfekterte celler (røde og grønne signaler) fra den totale populasjon av celler (DNA farging av DAPI). Ønsket transfeksjon andel er ca 20-40% når 100-200 celler telles.
3. Tekniske parametere av live Confocal Imaging Skannerinnstillinger
- ScanMode XYZT; Pinhole [luftig] 1,00; Zoom 3,5; Resolution 8 bits, Laser dpss 561 nm, Argon, synlig laser 488 nm, Laser han / Ne synlige 633 nm, Objective HCX PL APO CS 63x 1,40 OIL UV; Numerisk Aperture 1.4; Skannehastiget 700 Hz; Brytningsindeks 1.52.
- Bilde 3-D presentasjon ble utarbeidet av IMARIS bildebehandlingsprogrammer 7.0.
4. Confocal Imaging av Mitosis i Faste Cells
- Seed 2 x 10 5 MDA-MB-231 brystkreft celler (ATCC), normale mus embryonale fibroblaster (MEF), eller PARP1 mangelfull MEF (PARP - / -, utarbeidet av Dr Francoise Dantzer) på glass Dekkglass i 6-brønn plate i 2 ml komplett medium. Dekkglass ble vasket med 96% etanol, etter av vask med sterilt DD vann, tørket i 2 timer, og plassert i hver brønn av seks-brønners fatet.
- Legg PJ-34 (10-30 mikrometer) til mediet og inkuberes cellene for den nødvendige periode (vanligvis opp til 96 timer).
- Vask Dekkglass en gang med PBS (fosfatbufret saltvann), og fikser cellene ved hjelp av inkubasjon på is-kald metanol: aceton (1:1)-løsning, 7 min, -20 ° C.
- Sug fiksering løsning og la Dekkglass til tørk i kjemisk hette (på dette stadiet, kan de Dekkglass holdes i -20 ° C i flere uker).
- Vask Dekkglass en gang med PBST (PBS supplert med 0,1% Tween-20) for å permeabilize cellemembraner og blokkere av cellene med 10% NDS (normal Esel Serum) i PBST ('blokkering løsning') i 1 time ved RT.
- Inkuber permeabilized faste celler med primær antibodies for 2 time ved RT (for spindler og centrosomes farging). Antistoffene blir fortynnet i den blokkerende løsning som følger: anti-α tubulin (1:250 fortynning) og anti-γ tubulin (1:200 fortynning). Primære antistoffer blir anvendt som følger: 100 pl anvende (i en dråpe) av en blanding av de blokkerende antistoffer i løsning for hver dekkglass på 6-brønners plate deksel (dekselet er opp ned). Forsiktig sette dekkglass på antistoffer drop, seeded celler mot slipp. Inkuber Dekkglass vender antistoffer i 2 timer ved romtemperatur.
- Plasser Dekkglass tilbake i brønnene og vaske cellene tre ganger med PBST. Deretter bruker du samme metode som beskrevet i 4.6 for merking celler på Dekkglass med fluorescerende sekundære antistoffer. Inkuber cellene på dekkglass med det sekundære antistoff i 1 time, RT, i mørke. Antistoffene blir fortynnet i blokkering løsningen som følger: Alexa Fluor 488 (1:1.000 fortynning, grønn) og Alexa Fluor 568 (1:1.000 fortynning;red).
- Monter Dekkglass hjelp forlenge Gold antifade reagens med DAPI (for kromosomer farging) og inkuberes over natten ved RT i mørket for å tørke.
- Undersøke dekkglass ved konfokalmikroskopi.
5. Cell Livskraftig Målt ved ATP Produksjon
ATP-produksjon måles av en luminescerende ATP deteksjon assay kit.
- Seed cellene i 96-brønners plate, ca 20.000 celler i 800 ul medium i hver brønn. Tre blanke brønner bør brukes for bestemmelse av bakgrunnen luminescens av mediet.
- Forbered ATP standard fortynningsserier fra omtrent 10 pm til 100 uM og holde på is.
- Tilsett 50 pl av vaskemiddel til hver brønn og platen ristes i 5 min i orbital shaker, 700 opm.
- Rekonstitueres hver ampulle av den 'lyofiliserte substrat' med 5 ml av 'substrat-buffer "i settet.
- Tilsett 50 pl av den rekonstituerte løsning substrat til brønnene, og ristplaten i 5 min på orbital shaker, 700 opm.
- Hold platen i mørke i 10 min.
- Mål luminiscence av hver brønn ved ELISA mikroplateleser.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 er en stabil vannløselig phenanthridine 1 (figur 1). Våre tidligere resultater avslørt celledød og de-gruppert ekstra-centrosomes i flere typer faste multi-centrosomal kreftceller som ble behandlet med PJ-34. Derimot ble normale prolifererende celler ikke svekket 2,3. Centrosomes ble identifisert ved dobbel merking med antistoffer rettet mot centrine1 og γ-tubulin i de faste ekstra centrosomal celler to.
Her ble den cytotoksiske aktiviteten til PJ-34 dokumentert i disse live ekstra centrosomal celler på sanntid ved hjelp av levende konfokalmikroskopi. Leve menneskelig brystkreft MDA-MB-231 celler, som har en høy forekomst (> 50%) av ekstra-centrosomes 4,5, ble skannet i minst 16 timer etter confocal bildebehandling fokusert på transfekterte celler med γ-tubulin-GFP ( fluorescerende merking av γ-tubulin foci 2) og med histone H2b-RED (fluorescerende labeling av kromosomer). Seks til ti bor transfekterte celler ble skannet i parallell i hvert eksperiment. Double immunolabeling av γ-tubulin foci i transfekterte celler med centrin1 var teknisk umulig.
Spredt γ-tubulin foci og avvikende kromosomer ordningen ble sjelden påvises i tilfeldig utvalgte ubehandlede MDA-MB-231 celler i mitose. Bifokale clustering av γ-tubulin foci, som representerer ekstra-centrosomes bifokale clustering, ble dokumentert i de fleste levende ubehandlede MDA-MB-231 celler (figur 2), ble derimot un-gruppert centrosomes og avvikende arrangement av kromosomer påvist i levende transfektert MDA-MB-231-celler inkubert med PJ-34 (20 pM), og mitose i disse cellene avsluttet av celledød (figur 3). Disse celler ble inkubert med PJ-34 i 18 - 24 timer før skanning og for ytterligere 16 timer under skanning (fig. 3). Den sanntids dokumentasjon av de celleath under mitose sterkt en tidligere definert positiv korrelasjon mellom antall menneskelige ondartede celler med multi-polar spindler i mitose og prosenten av celledød i celler inkubert med PJ-34 (20 mm) 2.
PJ-34 fungerer som en potent PARP1 hemmer en. Vi undersøkte derfor muligheten for PARP1 hemming forårsaker celledød assosiert med mitotisk svikt. I motsetning til direkte avbildning, avbilding av faste MDA-MB-231-celler aktivert undersøkelse av en stor populasjon av celler i cellekultur, og dermed muliggjør statistisk analyse av virkningene av inhibitorer PARP1 i en rekke humane kreftcellelinjer. Aktiviteten av PJ-34 ble sammenlignet med aktiviteten av andre potente, ikke-fenantren PARP1 hemmere i normal eller PARP1 mangelfull celler (dvs. normal og PARP1 (- / -) mus embryonale fibroblaster (MEF)) (figur 4). PARP1 mangelfulle MEF havnen multi-centrosomes i mitose, men de er ikket kreftceller 11. Disse cellene ble utarbeidet av Dr. Francoise Dantzer, Strasbourg, Frankrike.
Fast normal og PARP1 (- / -) MEF ble immunolabeled for α-og γ-tubulin som er merket deres spindler og centrosomes, henholdsvis, som rapportert før to. Noen av de undersøkte celle-kulturene ble behandlet med PJ-34 eller andre potente, ikke-fenantren PARP1 inhibitorer, inkludert ABT-888 og AG01469, som inhiberer den enzymatiske aktiviteten til PARP1, og BSI-201, en forbindelse som tydeligvis demper PARP1 binding til bretter DNA 12-14. Ingen av de testede PARP1 hemmere svekket normal MEF ved konsentrasjoner hemmer PARP1 aktivitet (figur 4). I kontrast, PJ-34 doseavhengig forårsaket un-gruppering av γ-tubulin foci, forvrengning av spindlene og celledød i PARP1 (- / -) MEF (Fig. 4A og B). Dette ble ikke observert i normal MEF behandlet med PJ-34 (Figure 4B) eller i PARP1 (- / -) MEF behandles med ikke-phenenthrene PARP1 hemmere ABT-888 eller AG014699 (Figur 4C). Det bør bemerkes at PJ-34 ved konsentrasjoner over 20 pM ble svekke normal MEF, selv om normal MEF var mer resistente overfor PJ-34 aktivitet enn PARP1 (- / -) MEF.
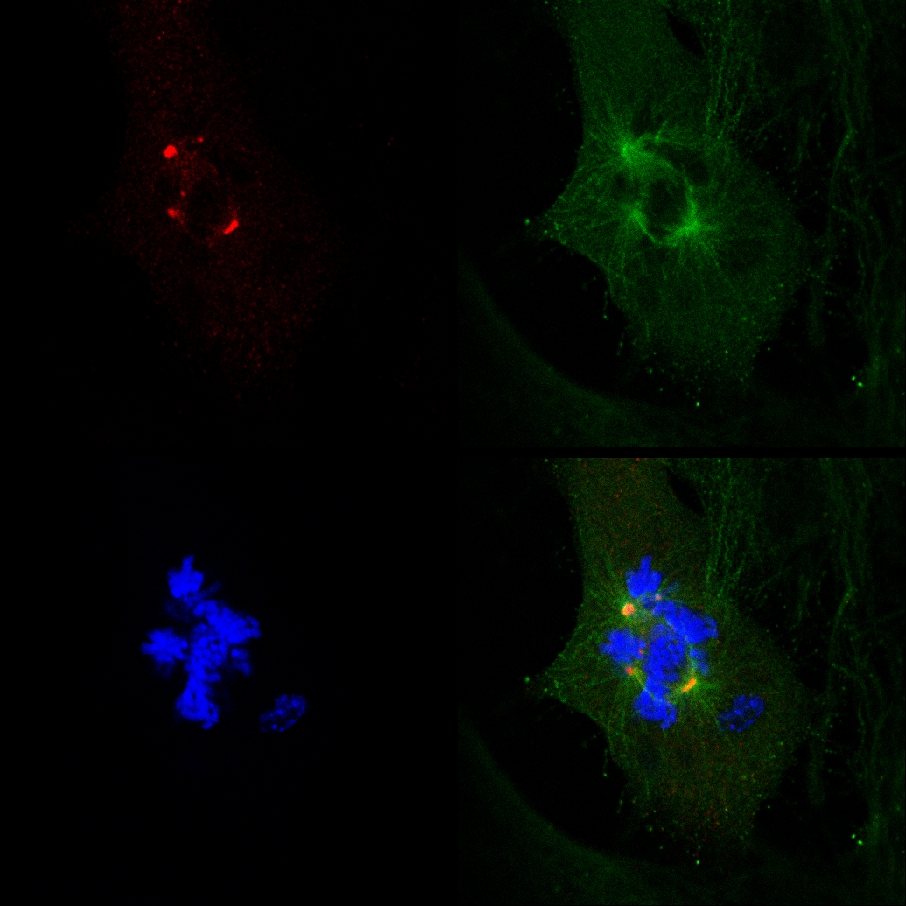
Det faktum at PJ-34 utryddet PARP1 (- / -) MEF til tross for deres PARP1 mangel, og sammenhengen mellom dannelsen av multi-focal spindler og celle utrydding i PARP1 (- / -) MEF inkubert med PJ-34 i konsentrasjoner høyere enn de som kreves for PARP1 hemming, ikke var forenlig med en kausal sammenheng mellom ekstra-centrosomes de-clustering i PARP1 (- / -) MEF og PARP1 hemming (Figur 4A). Cellegiften aktiviteten til PJ-34 i PARP1 (- / -) MEF kunne vært bedre forklart av sin virksomhet som en ekstra centrosomes de-clustering agent i multi-centrosomal celler 2 ( Tilleggsopplysninger "/> Figur 3. En live confocal bildebehandling dokumentasjon av celledød i en tilfeldig valgt MDA-MB-231 celler i mitose med un-klynger γ-tubulin merket ekstra-centrosomes. MDA-MB-231 celler ble inkubert med PJ-34 for 24 timer før du skanner og i løpet av 16 timer av levende confocal bildebehandling. PJ-34 ble brukt 24 timer etter transfeksjon med vektorer som uttrykker γ-tubulin-GFP (grønt, γ-tubulin foci og centrosomes) og H2b-RED (rød, kromosomer). 
Figur 1. Den phenanthridine PJ-34: N-(6-okso-5 ,6-dihydro-phenanthridin-2-yl)-N, N-dimetyl-acetamid. 
Figur 2. Bi-focal clustering av ekstra-centrosomes i en tilfeldig valgt levende MDA-MB-231 celler i mitose. A. Øvre panel: Merkede centrosomes i en tilfeldig valgt levende MDA-MB-231 celle tilført med γ-tubulin-GFP Nedre panel:. Chromosome rearrangementene under mitose i et tilfeldig valgt MDA-MB-231 celle tilført med histone H2b-RED. B. Bi-focal mitose med klynger ekstra-centrosomes identifisert i en tilfeldig valgt kultfigurert MDA-MB-231 celle. Celler ble tilført både γ-tubulin-GFP (merking γ-tubulin foci, grønn) og histone H2b-Red (merking kromosomer, rød). 48 timer etter transfeksjon, ble cellene eksponert for en levende avbildning konfokal i 16 timer. Seks celler ble skannet i parallell i hvert eksperiment. Fire forskjellige forsøk ble utført. Se også Supplementary Information. Klikk her for å se større figur . 
Figur 3. Ekstra-centrosomes de-clustering innledes celledød i live-MDA-MB-231 celler behandlet med PJ-34. Et tilfeldig valgt levende MDA-MB-231 celler i mitose med spredte centrosomes (1 st rammen på venstre) endte ved celledød ( 2. og 3. rammer). Dette cell ble tilfeldig valgt i en cellekultur ruges i 24 timer med PJ-34 (20 mm) brukes 24 timer etter transfeksjon med vektorer som uttrykker γ-tubulin-GFP (merking γ-tubulin foci inkludert centrosomes, grønn) og histone H2b-RED ( merking kromosomer, rød). Cellen ble skannet for 16 timer av live confocal bildebehandling. Seks celler ble skannet i parallell i hvert eksperiment. Tre forskjellige forsøk ble utført. Se også Supplementary Information. Klikk her for å se større figur . 
Figur 4. En cytotoksisk aktivitet av PJ-34 i PARP1 (- / -) mus embryonale fibroblaster. A. (Venstre) Andelen av multi-focal spindler beregnet normal (sort linje) og Parp1 - / - (grå linje) MEF, ruges i 48 timer med PJ-34 på de angitte konsentrasjoner. Prosentandelen av multi-polar spindler ble beregnet ut fra total spindler 20 detektert i tre forskjellige eksperimenter. (Høyre) Redusert celleoverlevelse oppdaget i cellekulturer ruges i 72 timer med PJ-34 (20 mm) i forhold til overlevelse av kontroll ubehandlede celler (normal (sort linje) og Parp1 - / - (grå linje) MEF). Celle overlevelsen ble bestemt ved cellens ATP-produksjon (protokoll 5). Gjennomsnittsverdiene av fire målinger for hver cellelinje i tre ulike eksperimenter presenteres B. Spindler i tilfeldig valgt fast normal og Parp1 -. / - MEF i mitose, ubehandlet (kontroll) eller inkubert med PJ-34 i 48 timer på den angitte konsentrasjoner. PJ-34 forårsaket multipolar spindler. Celler ble fikset, permeabilized og immunolabeled for α-og γ-tubulin (grønn merking av spindler og rød merking av centrosomes, henholdsvis). Kromosomer ble merket med DAPI reagens (blå). . Representative resultatene av tre forskjellige eksperimenter C. Potente ikke-fenantren PARP1 hemmere ikke påvirke centrosomes clustering i PARP1 (- / -) MEF. Spindler av tilfeldig valgt normal og Parp1 - / - MEF er presentert; ubehandlet MEF (kontroll) eller MEF behandlet i 48 timer med non-fenantren PARP-hemmere, AG01469 (20 mm) eller ABT888 (20 mm). Kromosomene merket med DAPI reagens (blå). Lignende resultater ble oppnådd i tre forskjellige eksperimenter. Klikk her for å se større figur .
Klikk her for å se større figur . 
Klikk her for å se større figur .
Figur 2B. Supplerende figur. Et tilfeldig valgt MDA-MB-231 celle i anaphase med bi-fokale gruppert γ-tubulin merket ekstra-centrosomes ble skannet for 16 timer av live confocal bildebehandling, 48 timer etter å ha blitt tilført med vektorer som uttrykker γ-tubulin-GFP og h2b-RED (merking γ-tubulin foci og centrosomes i de faste celler (grønn) og H2b histone merking av kromosomene (rød), henholdsvis).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Leve confocal bildebehandling gitt en real-time dokumentasjon av den cytotoksiske effekten av PJ-34 i levende multi-centrosomal celler under mitose (figur 3 og utfyllende informasjon). Dette var den første live dokumentasjon tilskrive cytotoksisitet av PJ-34 i humane kreftceller til ekstra centrosomes de-clustering og celledød, noe som tyder induksjon av Mitotisk Catastrophe celledød ved PJ-34 5-9. I kontrast, ble bi-fokale clustering av super-numerario centrosomes observert i levende ubehandlede MDA-MB-231 celler gjennomgår normal mitose med bi-focali gruppert ekstra centrosomes (Figur 2, utfyllende informasjon).
Ifølge disse resultatene, leve confocal avbildning av transfekterte celler kan være nyttig for å oppdage translokasjon av proteiner involvert i ekstra-centrosomes clustering i transfekterte celler under mitose 5,9,15,16. Identifisering av proteiner som påvirkes av PJ-34 i multi-centrosOmal celler kan gi noen ledetråder for å forstå døden mekanismer aktiveres av ekstra-centrosomes de-clustering.
Fordelen med levende confocal bildebehandling i å gi informasjon i sanntid under mitose i levende celler, er også bundet til flere begrensninger. Antall celler per skannet eksperiment er begrenset. Derfor sjanser til å oppdage celler i mitose er lave, og flere gjentatte forsøk er nødvendig for en real-time dokumentasjon av mitose i de skannede celler. I tillegg er suksess svært avhengig av høy effektivitet av transfeksjon av celler med vektorer som uttrykker merkede proteiner. Dermed, til tross for å være pålitelig, er live confocal bildebehandling tidkrevende og krever svært erfarne hardt arbeidende.
Til sammenligning immunocytokjemi og konfokal avbildning av faste celler muliggjøre undersøkelse av et stort antall celler pr forsøket, som er nødvendig for en pålitelig statistisk analyse. Vi brukte denne metoden for å sammenligneeffektene av PJ-34 i mitose av normale godartede celler til dens virkninger i kreftcellene gjennomgår mitose med overtallige centrosomes 2,3. På samme måte ble denne metoden for å sammenligne effektene av PJ-34 som hos ikke-fenantren PARP1 inhibitorer i PARP (- / -) MEF husing ekstra-centrosomes (celler med høy forekomst av multi-centrosomes 11) (figur 4).
Oppsummert Våre resultater indikerer fordelen av å kombinere de mest verdifulle sanntids informasjon gitt av konfokal avbildning av levende celler i mitose (Figurene 2 og 3) med cytochemistry og konfokal analyse av faste celler. En kombinasjon av disse metoder kan være nyttige for å identifisere den cytotoksiske aktivitet av små molekyler som, i likhet PJ-34, mål-spesifikke mekanismer viktig for celleoverlevelse. Den unike avhengighet av mange menneskelige kreftceller på ekstra-centrosomes bi-polar clustering for deres spredning og survival gjengir PJ-34 en mulig kandidat for kreftterapi.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer at de har ingen konkurrerende finansielle interesser.
Acknowledgments
Finansiering kilder til denne forskningen: et felles fond av Tel Aviv University teknologioverføring selskapet, Ramot og Sheba-Medical Center (M. CA og SI.), ICRF - Israelsk Cancer Research Foundation (M. CA.) Og Israel Science Foundation ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}