

Summary
Den cytotoksiske aktivitet af phenanthridin PJ-34 i cancerceller undergår mitose blev dokumenteret i realtid ved levende konfokal billeddannelse. PJ-34 udryddet human brystkræft MDA-MB-231 celler, der huser ekstra-centrosomes i mitose. I modsætning til normal bi-omdrejningspunkt mitose blev de ekstra-centrosomes ikke grupperet i de to spindel poler i tilstedeværelse af PJ-34.
Abstract
Phenanthren derivater fungerer som potente PARP1 hæmmere forhindrede bi-omdrejningspunkt klyngedannelse af overtallige centrosomes i multi-centrosomal humane kræftceller i mitose. Den phenanthridin PJ-34 var den mest potente molekylet. Declustering ekstra-centrosomes forårsager mitotisk svigt og celledød i multi-centrosomal celler. Mest solide menneskelige kræftformer har høj forekomst af ekstra-centrosomes. Aktiviteten af PJ-34 blev dokumenteret i realtid ved konfokal billeddannelse af levende human brystcancer MDA-MB-231-celler transficeret med vektorer, der koder for fluorescerende γ-tubulin, som er meget rigelige i centrosomes og fluorescerende histon H2b stede i kromosomerne. Afvigende kromosomer arrangementer og de-klynger γ-tubulin foci repræsenterer declustered centrosomes blev påvist i de transficerede MDA-MB-231-celler efter behandling med PJ-34. Un-klynger ekstra-centrosomes i de to spindel poler forud deres celledød. Disse resultater er forbundet for første gang den nyligt opdaget eksklusiv cytotoksiske aktivitet af PJ-34 i humane cancerceller med ekstra centrosomes de-clustering i mitose, og mitotisk svigt fører til celledød. Ifølge tidligere resultater observeret ved konfokal imaging af faste celler, PJ-34 udelukkende udryddet kræftceller med multi-centrosomes uden at forringe normale celler undergår mitose med to centrosomes og bi-knudepunkt spindler. Denne cytotoksiske aktivitet af PJ-34 deltes ikke af andre potente PARP1 hæmmere, og blev observeret i PARP1 deficiente MEF huser extracentrosomes, tyder dens uafhængighed af PARP1 hæmning. Levende konfokal imaging tilbudt et nyttigt redskab til at identificere nye molekyler udrydde celler under mitose.
Introduction
Phenanthren afledte PARP1 hæmmere, inklusive PJ-34, blev designet til at beskytte hvilende celler fra apoptotisk celledød induceret af energikrævende PARP1 medieret DNA-reparation under stress betingelser (slagtilfælde eller myokardieinfarkt) 1. Men for nylig opdagede vi, at PJ-34, på det dobbelte højere koncentration end inducerende PARP1 hæmning kan udelukkende medføre celledød i humane cancerceller 2,3. Den hurtigere spredning af cellen var, jo mere effektiv udryddelse af cellerne var. Den cytotoksiske aktivitet PJ-34 blev tilskrevet ekstra-centrosomes de-clustering i mitose 2.. Mange human cancer celler havnen multicentrosomes 4,5. Inkubation af humane brystkræftceller MDA-MB-231, som huser overtallige centrosomes, med 20 uM PJ-34 effektivt udryddet disse celler inden 72-96 timer uden at forringe hvilende celler eller nogle godartede prolifererende celler huser to centrosomes i mitose
Bipolar centrosome samling er afgørende for bipolar spindel dannelse i mitose 4,5. Derfor har celler med mere end to centrosomes udviklet en næppe forstået molekylære mekanisme, clustering deres ekstra centrosomes på to poler 4-9. Manglende bipolar samling af deres centrosomes kan forårsage multipolær forvrænget spindler og afvigende kromosomer segregation, at anholdelser celle-cyklus G2 / M anholdelse, og fører til celledød tilskrives mitotisk fiasko 4,5. De molekylære mekanismer bag ekstra-centrosomes de-clustering intensivt undersøgte <sup> 10. Forståelse dette dødsfald mekanisme vil gøre det muligt for eksklusiv udryddelse af kræftceller, mens besparende raske væv 5,10.
Således forbindelser, som aktiverer mitotisk katastrofe celledød tilbyde en ny form for en selektiv cancerterapi, som kan være effektive i en bred vifte af humane faste cancers.Our resultater antyder, at konfokal billeddannelse kan anvendes til at identificere molekyler, der påvirker ekstra-centrosomes klyngedannelse i mitose 2,3, hvilket gør disse forbindelser kræft målretning lægemiddelkandidater.
Vi har dokumenteret den cytotoksiske aktivitet af phenanthridin PJ-34 ved at scanne faste og levende humane kræftceller (med høj forekomst af ekstra-centrosomes i mitosen) versus normale celler. En trin-for trin beskrivelse af billeddiagnostiske procedurer, der anvendes til at identificere den cytotoksiske aktivitet PJ-34 i humane cancerceller er medtaget nedenfor.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1.. Cell Culture Forberedelse
MDA-MB-231-celler blev indkøbt fra ATCC (American Type Culture Collection) og opbevaret i flydende nitrogen.
- Seed 10 6 MDA-MB-231 celler i 92 mm diameter petriskål i 10 ml komplet medium indeholdende Dulbeco Modified Eagle Medium (DMEM), 10% hesteserum, 1% L-glutamin og 1% Penstrep-Amphotericin B. Tillad celler at formere til omkring 80-100% konfluens.
- Fjern dyrkningsmedium fra fad og kassér.
- Vask cellelag kortvarigt med 0,25% (w / v) trypsin-EDTA-opløsning for at fjerne alle spor af serum.
- Tilsæt 2,0 ml trypsin-EDTA-opløsning til parabol og observere celler ved omvendt mikroskop indtil cellelag dispergeres (sædvanligvis inden for 5 til 15 min).
- Tilføj 18 ml komplet vækstmedium og forsigtigt aspireres cellerne med pipette. Overførsel til et rør.
- Centrifuger cellesuspensionen ved 1200 rpm.
- Genopslæmmes cellepelleten i 24 ml cuLTURE medium.
- Tilsæt 2 ml cellesuspension til 35 mm glasbund retter (omkring 25% sammenløb) og anbring dem i inkubator (5% CO 2, 37 ° C).
- Løsninger:
- Komplet Medium for celler spredning: DMEM med 10% FBS, 1% antibiotika (100 enheder / ml penicillin G, 100 ug / ml streptomycin, Pen-Strep-ampho opløsning) og 2 mM L-glutamin.
- Trypsin-EDTA-opløsning, indeholdende 0,25% trypsin-EDTA.
- Retter:
92 mm diameter petriskåle.
35 mm diameter poly-D-lysin-overtrukne glasbund kultur retter.
2.. Udarbejdelse af celler til Live Confocal Imaging
- Seed 2 x 10 5 MDA-MB-231 celler i glasbund kultur retter i 2 ml komplet medium som nævnt i § 1. Når cellekultur når sammenløb af 60-70% (ca. 3-4 x 10 5 celler pr skål), så fortsæt med transfektion.
- Transficere cellerne med to plasmider ENCODING fusion proteiner γ-tubulin-GFP (til fluorescerende detektion af centrosomes) og histon-RED (H2b-RED, for fluorescerende påvisning af kromosomer) ved hjælp af liposomal transfektionsreagens Jet-PI efter fremstillingen protokol. Kort fortalt, blandes 2 ug fra hvert plasmid i et rør med 100 pi NaCl (150 mM). Bland transfektionsreagens (100 ul) med 100 pi NaCl (150 mM) i et andet rør, og inkuber 5 minutter ved stuetemperatur (RT). Derefter kombinere de to løsninger, mix (ved hjælp af mild vortex) og spin-down. Inkuber i 30 minutter ved stuetemperatur.
- Under inkubation af transfektion blandingen vaskes cellerne en gang med PBS og erstatte den celle medium med 2 ml varm DMEM uden tillæg (37 ° C).
- Tilfør forsigtigt transfektion blandingen til cellerne i DMEM og derefter returnere cellerne til inkubatoren (37 ° C, 5% CO 2) i 8 timer.
- Efter 8 timers inkubering erstatte DMEM med 2 ml komplet medium og inkubere cellerne i inkubatoren i 24time.
- 24 timer efter transfektion, udskifte mediet af cellerne med 2 ml komplet medium indeholdende 20 uM PJ-34.
- Inkuber cellerne i yderligere 18 timer (37 ° C, 5% CO 2).
- Subject cellerne til at leve konfokal billedbehandling for mindst 16 timer i billedbehandling kammer holde cellerne ved 5% CO 2 og 37 ° C.
- Parallelt hermed undersøge transfektion effektivitet 36 timer efter transfektion ved hjælp af fluorescensmikroskopi som følger:
- Seed 2 x 10 5 MDA-MB-231-celler i 6-brønds plade indeholdende 1 dækglas per brønd i 2 ml komplet medium.
- Transficere celler som nævnt i § § 2,2-2,5.
- 36 timer efter transfektion fastsætte de transficerede celler monteret på et dækglas ved inkubation i kold methanol: acetone (1:1) opløsning, 7 min, -20 ° C.
- Aspirer fiksering løsningen og lad dækglasset med de monterede celler til tørre i en kemisk hætte.
- Anvend forlænge Guld antiblegemiddel reagenset med DAPI og lad the dækglas at tørre i mørke i 6 timer.
- Undersøg slide under fluorescerende mikroskop og beregne den procentdel af de transfekterede celler (røde og grønne signaler) fra den samlede population af celler (DNA-farvning af DAPI). Den ønskede transfektion omtrent 20-40%, når 100-200 celler tælles.
3.. Tekniske parametre for Levende Konfokale Imaging Scanner Settings
- ScanMode XYZT; Pinhole [luftige] 1.00, Zoom 3.5, Opløsning 8 bits, Laser DPSS 561 nm argon, synligt laser 488 nm Laser He / Ne synlige 633 nm Målsætning HCX PL APO CS 63x 1.40 OIL UV, blændetal 1.4; Scan hastighed 700 Hz Refraktionsindeks 1.52.
- Billede 3-D præsentation er udarbejdet af Imaris imaging software 7.0.
4.. Confocal Imaging i mitosen i faste Cells
- Seed 2 x 10 5 MDA-MB-231 brystkræftceller (ATCC), normal mus embryonale fibroblaster (MEF), eller PARP1 mangelfuld MEF (PARP - / -, fremstillet af Dr. Francoise Dantzer) på dækglas i 6-brønds plade i 2 ml komplet medium. Dækglas blev vasket med 96% ethanol, efterfulgt af vask med sterilt DD vand, tørret i 2 timer og anbragt i hver brønd i 6-brønds skål.
- Tilføj PJ-34 (10-30 uM) til mediet og inkuberes cellerne i den påkrævede periode (normalt op til 96 timer).
- Vask dækglassene gang med PBS (phosphatbufret saltvand), og fikseres cellerne med inkubation i iskold methanol: acetone (1:1) opløsning, 7 min, -20 ° C.
- Aspirer fiksering opløsning og lad dækglassene til tørre i kemisk hætte (på dette tidspunkt, kan dækglassene opbevares i -20 ° C i flere uger).
- Vask dækglassene gang med PBST (PBS suppleret med 0,1% Tween-20) for at permeabilisere cellemembranerne og blokere cellerne med 10% NDS (Normal Donkey Serum) i PBST (»blokerende opløsning) i 1 time ved stuetemperatur.
- Inkubér permeabiliserede fikserede celler med primær antibodies i 2 timer ved stuetemperatur (for spindler og centrosomes farvning). Antistofferne fortyndes i den blokerende opløsning som følger: anti-α tubulin (1:250 fortynding) og anti-γ tubulin (1:200 fortynding). Primære antistoffer anvendes som følger: anvende 100 pi (et fald) af en blanding af antistofferne i blokerende opløsning for hvert dækglas på 6-brønds plade dæksel (dækslet er hovedet). Sæt forsigtigt dækglasset på antistoffer slip, seedede celler vender dråben. Inkuber dækglassene står antistofferne i 2 timer ved stuetemperatur.
- Placer dækglassene tilbage i brøndene og vaske cellerne 3 gange med PBST. Så brug den samme procedure som beskrevet i 4.6 for mærkning celler på dækglas med de fluorescerende sekundære antistoffer. Inkubér cellerne på dækglassene med de sekundære antistoffer i 1 time, RT, i mørke. Antistofferne fortyndes i blokerende opløsning som følger: Alexa Fluor 488 (1:1000 fortynding grøn) og Alexa Fluor 568 (1:1000 fortynding;rød).
- Monter dækglas hjælp forlænge Guld antiblegemiddel reagenset med DAPI (for kromosomer farvning) og inkuberes natten over ved stuetemperatur i mørke for at tørre.
- Undersøg dækglas ved konfokal mikroskopi.
5.. Cellelevedygtigheden Målt ved ATP Production
ATP er målt med en selvlysende ATP-assay kit.
- Frø cellerne i 96-brønds plade, omkring 20.000 celler i 800 pi medium i hver brønd. Tre tomme brønde bør anvendes til bestemmelse af baggrunden luminescens af mediet.
- Forbered ATP standardfortyndingsrækker fra cirka 10 um til 100 um og holde på is.
- Der tilsættes 50 ul af vaskemiddel til hver brønd og ryste pladen i 5 minutter i orbitalryster, 700 rpm.
- Opløs indholdet af hvert hætteglas af "frysetørret substrat" med 5 ml af 'substratbuffer "i sættet.
- Der tilsættes 50 ul af den rekonstituerede substratopløsning til brøndene, og rystpladen i 5 minutter på rysteapparat, 700 rpm.
- Hold pladen i mørke i 10 minutter.
- Måle luminiscence af hver brønd ved ELISA mikropladelæser.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 er en stabil vandopløselig phenanthridin 1 (figur 1). Vores tidligere resultater viste celledød og de-grupperet ekstra centrosomes i flere typer af faste multi-centrosomal kræftceller, der blev behandlet med PJ-34. I modsætning hertil var normale prolifererende celler ikke værdiforringede 2,3. Centrosomes blev identificeret ved at dobbeltklikke mærkning med antistoffer rettet mod centrine1 og γ-tubulin i de faste ekstra centrosomal celler 2.
Her blev den cytotoksiske aktivitet af PJ-34 dokumenteret i disse levende ekstra centrosomal celler på realtid ved hjælp af levende konfokal mikroskopi. Levende human brystcancer MDA-MB-231-celler, som har en høj forekomst (> 50%) af ekstra-centrosomes 4,5, blev scannet for mindst 16 timer ved konfokal billeddannelse fokuseret på transficerede celler med γ-tubulin-GFP ( fluorescerende mærkning af γ-tubulin foci 2) og histon H2b-RED (fluorescerende Labeling af kromosomer). Seks til ti levende transficerede celler blev scannet parallelt i hvert eksperiment. Dobbelt immunolabeling af γ-tubulin foci i de transficerede celler med centrin1 var teknisk umuligt.
Spredt γ-tubulin foci og afvigende kromosomer arrangement sjældent blev påvist i tilfældigt udvalgte ubehandlede MDA-MB-231 celler i mitose. Bifokal gruppering af γ-tubulin foci, som repræsenterer ekstra-centrosomes bifokale klyngedannelse, blev dokumenteret i de fleste levende ubehandlede MDA-MB-231 celler (figur 2) blev derimod-un klynger centrosomes og afvigende placering af kromosomer påvist i levende transficeret MDA-MB-231-celler inkuberet med PJ-34 (20 uM), og mitose i disse celler endte ved celledød (figur 3). Disse celler blev inkuberet med PJ-34 for 18-24 timer før scanning og for yderligere 16 timer under scanningen (figur 3). Den realtid dokumentation for celle death under mitosen støtter en tidligere defineret positiv korrelation mellem antallet af humane maligne celler med multipolær spindler i mitose og procentdelen af celledød i celler inkuberet med PJ-34 (20 uM) 2.
PJ-34 virker som en potent inhibitor PARP1 1.. Vi har derfor undersøgt muligheden for PARP1 hæmning fremkalder celledød forbundet med mitotisk fiasko. I modsætning til live-billedbehandling, aktiveret billeddannelse af faste MDA-MB-231-celler undersøgelse af en stor population af celler i cellekulturer hvorved statistisk analyse af virkningerne af PARP1 hæmmere i en bred vifte af menneskelige kræftceller. Aktiviteten af PJ-34 blev sammenlignet med aktiviteten af andre potente, ikke-phenanthren PARP1 hæmmere i normal eller PARP1 mangelfulde celler (dvs. normal og PARP1 (- / -) mus embryonale fibroblaster (MEF)) (Figur 4). PARP1 mangelfuld MEF havnen multi-centrosomes i mitosen, men de er ikket tumorceller 11.. Disse celler blev fremstillet af Dr. Francoise Dantzer, Strasbourg, Frankrig.
Fast normal og PARP1 (- / -) MEF blev immunolabeled for α-og γ-tubulin, der mærkede deres spindler og centrosomes henholdsvis som rapporteret før 2.. Nogle af de undersøgte cellekulturer blev behandlet med PJ-34 eller andre potente, ikke-phenanthren PARP1 inhibitorer, herunder ABT-888 og AG01469, der hæmmer den enzymatiske aktivitet af PARP1, og BSI-201, en forbindelse, der tilsyneladende dæmper PARP1 binding til hakker DNA 12-14. Ingen af de testede PARP1 inhibitorer svækkede normal MEF ved koncentrationer hæmmer PARP1 aktivitet (figur 4). I modsætning hertil dosisafhængigt PJ-34 forårsagede un-gruppering af γ-tubulin brændpunkter, fordrejning af spindler og celledød i PARP1 (- / -) MEF (figur 4A og B). Dette blev ikke observeret i normal MEF behandlet med PJ-34 (Figure 4B) eller i PARP1 (- / -) MEF behandlet med ikke-phenenthrene PARP1 inhibitorer ABT-888 eller AG014699 (fig. 4C). Det skal bemærkes, at PJ-34 i koncentrationer på over 20 uM havde forringe normal MEF, skønt normal MEF var mere resistente over for PJ-34 aktivitet end PARP1 (- / -) MEF.
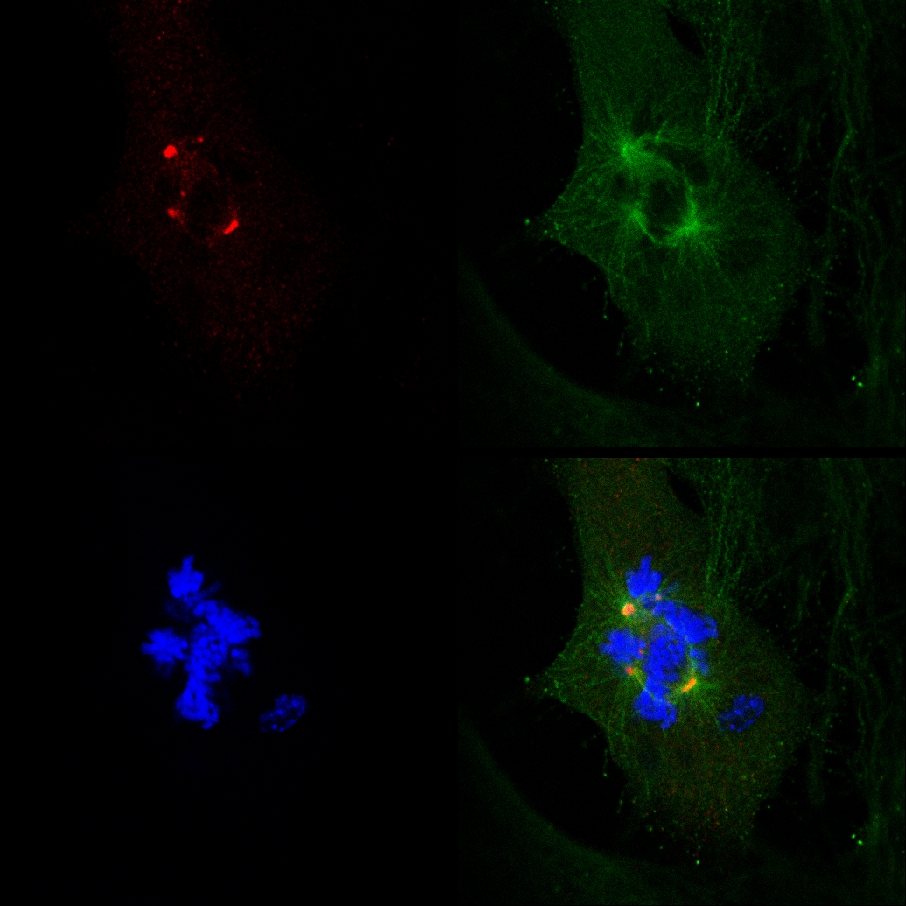
Det faktum, at PJ-34 udryddet PARP1 (- / -) MEF på trods af deres PARP1 mangel, samt sammenhængen mellem dannelsen af multifokal spindler og celle udryddelse i PARP1 (- / -) MEF inkuberet med PJ-34 ved koncentrationer højere end dem, der kræves for PARP1 hæmning, ikke var i overensstemmelse med en kausal sammenhæng mellem ekstra-centrosomes de-klyngedannelse i PARP1 (- / -) MEF og PARP1 hæmning (Figur 4A). Den cytotoksiske aktivitet PJ-34 i PARP1 (- / -) MEF bedre kunne forklares ved dens aktivitet som en ekstra centrosomes de-clustering agent i multi-centrosomal celler 2 ( Supplerende information "/> Figur 3. En live konfokal imaging dokumentation af celledød i en tilfældigt udvalgt MDA-MB-231 celle i mitosen med un-klynger γ-tubulin mærkede ekstra-centrosomes. MDA-MB-231-celler blev inkuberet med PJ-34 i 24 timer før scanning og under 16 timer af levende konfokal billeddannelse. PJ-34 blev anvendt 24 timer efter transfektion med vektorer, der udtrykker γ-tubulin-GFP (grønt, γ-tubulin foci og centrosomes) og H2b-RED (rød, kromosomer). 
Figur 1. Den phenanthridin PJ-34: N-(6-oxo-5 ,6-dihydro-phenanthridin-2-yl)-N, N-dimethyl-acetamid. 
Figur 2. Bi-omdrejningspunkt gruppering af ekstra-centrosomes i en tilfældigt udvalgt levende MDA-MB-231 celle i mitose. A. Overpanel: Mærkede centrosomes i en tilfældigt udvalgt levende MDA-MB-231 celler transficeret med γ-tubulin-GFP Underpanel:. Kromosom re-arrangementer under mitosen i en tilfældigt udvalgt MDA-MB-231 celler transficeret med histon H2b-RED. B. Bi-omdrejningspunkt mitose med klynger ekstra-centrosomes identificeret i en tilfældigt udvalgt kultgureres MDA-MB-231 celle. Celler blev transficeret af både γ-tubulin-GFP (mærkning γ-tubulin foci, grøn) og histon H2b-RED (mærkning kromosomer, rød). 48 timer efter transfektion blev cellerne udsat for en levende konfokal billeddannelse til 16 timer. Seks celler blev scannet parallelt i hvert forsøg. Fire forskellige eksperimenter blev udført. Se også supplerende oplysninger. Klik her for at se større figur . 
Figur 3. Ekstra centrosomes de-klyngedannelse forud celledød i levende MDA-MB-231 celler behandlet med PJ-34. Et tilfældigt udvalgt levende MDA-MB-231 celle i mitosen med spredte centrosomes (1 st ramme på venstre) sluttede af celledød ( 2. og 3. rammer). Denne cell blev tilfældigt udvalgt i en cellekultur inkuberet i 24 timer med PJ-34 (20 uM) anvendt 24 timer efter transfektion med vektorer, der udtrykker γ-tubulin-GFP (mærkning γ-tubulin foci herunder centrosomes, grøn) og histon H2b-RED ( mærkning kromosomer, rød). Cellen blev scannet i 16 timer ved levende konfokal billedbehandling. Seks celler blev scannet parallelt i hvert forsøg. Tre forskellige eksperimenter blev udført. Se også supplerende oplysninger. Klik her for at se større figur . 
Figur 4.. En cytotoksisk aktivitet PJ-34 i PARP1 (- / -) mus embryonale fibroblaster. A. (Venstre) Procentdelen af multifokal spindler beregnet i normal (sort linje), og Parp1 - / - (grå linje) MEF, inkuberes i 48 timer med PJ-34 ved de angivne koncentrationer. Procentdelen af multi-polar spindler blev beregnet ud af 20 i alt spindler påvist i 3 forskellige eksperimenter. (Højre) Reduceret celleoverlevelse påvist i cellekulturer inkuberet i 72 timer med PJ-34 (20 pM) i forhold til overlevelsen af kontrol ubehandlede celler (normal (sort linie) og Parp1 - / - (grå linie) MEF). Celleoverlevelse blev analyseret ved cellernes ATP-produktion (protokol 5). Middelværdierne af 4 målinger for hver cellelinie i 3 forskellige eksperimenter præsenteres B. Spindler i tilfældigt udvalgte fast normal og Parp1 -. / - MEF i mitose, ubehandlet (kontrol) eller inkuberet med PJ-34 i 48 timer ved den angivne koncentrationer. PJ-34 forårsagede multipolær spindler. Cellerne blev fastsat, permeabiliseret og immunolabeled for α-og γ-tubulin (grøn mærkning af spindler og rød mærkning af centrosomes, henholdsvis). Kromosomer blev mærket med DAPI reagens (blå). . Repræsentative resultater af 3 forskellige eksperimenter C. Potente non-phenanthren PARP1 hæmmere ikke påvirkede centrosomes klyngedannelse i PARP1 (- / -) MEF. Spindler af tilfældigt udvalgte normal og Parp1 - / - MEF præsenteres, ubehandlet MEF (kontrol) eller MEF behandlet i 48 timer med non-phenanthren PARP-inhibitorer, AG01469 (20 uM), eller ABT888 (20 uM). Kromosomer er mærket med DAPI reagens (blå). Lignende resultater blev opnået i 3 forskellige eksperimenter. Klik her for at se større figur .
Klik her for at se større figur . 
Klik her for at se større figur .
Figur 2B. Supplerende Figur. En tilfældigt udvalgt MDA-MB-231 celle i anaphase med bi-knudepunkt grupperet γ-tubulin mærket ekstra-centrosomes blev scannet i 16 timer ved levende konfokal billedbehandling, 48 timer efter at være blevet transficeret med vektorer, der udtrykker γ-tubulin-GFP og H2b-RED (mærkning γ-tubulin foci og centrosomes i de faste celler (grøn) og H2b histon mærkning af kromosomerne (rød), henholdsvis).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Levende konfokal imaging forudsat en real-time dokumentation af den cytotoksiske effekt af PJ-34 i levende multi-centrosomal celler under mitose (figur 3, og supplerende oplysninger). Dette var den første levende dokumentation tilskrive cytotoksicitet PJ-34 i humane kræftceller til ekstra centrosomes de-clustering og celledød, hvilket tyder på induktion af Mitotisk Catastrophe celledød ved PJ-34 5-9. I modsætning hertil var bi-omdrejningspunkt gruppering af super-numerary centrosomes observeret i levende ubehandlede MDA-MB-231-celler undergår normal mitose med bi-focali klynger ekstra centrosomes (figur 2, supplerende oplysninger).
Ifølge disse resultater, konfokal billedbehandling af transfekterede celler kan være nyttig til påvisning translokation af proteiner involveret i ekstra-centrosomes klyngedannelse i transficerede celler under mitose 5,9,15,16 leve. Identifikation af proteiner ramt af PJ-34 i multi-centrosOmal celler kan give nogle fingerpeg for at forstå de dødsfald mekanismer aktiveres af ekstra-centrosomes de-clustering.
Fordelen af levende konfokal billeddannelse i levere real-time information under mitosen i levende celler, er også bundet til flere begrænsninger. Antallet af celler scannet pr eksperiment er begrænset. Derfor chancer for at opdage celler i mitose er lave, og flere gentagne forsøg er nødvendige for en real-time dokumentation af mitose i de scannede celler. Desuden er succes stærkt afhængig høj effektivitet af transfektion af celler med vektorer, der udtrykker mærkede proteiner. På trods af at være pålidelige, er live konfokal imaging tidskrævende og kræver meget erfarne hårde arbejdstagere.
I sammenligning, immunocytokemi og konfokal billeddannelse af fikserede celler muliggøre undersøgelse af et stort antal celler pr eksperiment, som er nødvendig for en pålidelig statistisk analyse. Vi brugte denne metode til at sammenligneeffekter af PJ-34 i mitosen af normale godartede celler til sine virkninger i kræftceller undergår mitose med overtallige centrosomes 2,3. Tilsvarende blev denne metode anvendes til at sammenligne virkningerne af PJ-34 til at ikke-phenanthren PARP1 hæmmere i PARP (- / -) MEF huser ekstra-centrosomes (celler med høj forekomst af multi-centrosomes 11) (figur 4).
Sammenfattende indikerer vores resultater den fordel at kombinere de mest værdifulde realtidsinformation fra konfokal billeddannelse af levende celler i mitose (figur 2 og 3) med cytokemi og konfokal analyse af fikserede celler. En kombination af disse metoder kan være nyttige til at identificere den cytotoksiske aktivitet af små molekyler, der ligesom PJ-34, målrette specifikke mekanismer afgørende for celleoverlevelse. Den unikke afhængighed af mange menneskelige kræftceller på ekstra centrosomes bi-polar klyngedannelse for deres spredning og efterlevendeal gør PJ-34 en mulig kandidat til kræftbehandling.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer, at de ikke har nogen konkurrerende finansielle interesser.
Acknowledgments
Finansieringskilder af denne forskning: en fælles fond af Tel Avivs Universitets teknologioverførsel selskab, Ramot og Sheba-Medical Center (M. CA og SI.) ICRF - Israelsk Cancer Research Foundation (M. CA.), Og Israel Science Foundation ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}