

Summary
L'activité cytotoxique de l'phénanthridiniques PJ-34 dans les cellules cancéreuses en mitose a été documentée en temps réel par imagerie confocale en direct. 34-PJ cancer du sein humain éradiqué MDA-MB-231 cellules hébergeant des extra-centrosomes dans la mitose. Contrairement à la mitose bifocal normale, les extra-centrosomes sont regroupés dans les deux pôles du fuseau, en présence d'PJ-34.
Abstract
Dérivés de phénanthrène agissant comme inhibiteurs puissants PARP1 empêché le regroupement bi-focal de centrosomes surnuméraires dans les cellules cancéreuses humaines multi-centrosomales en mitose. Le phénanthridiniques PJ-34 est la molécule la plus puissante. Declustering d'extra-centrosomes provoque l'échec de la mitose et la mort cellulaire dans les cellules multi-centrosomales. Cancers humains les plus solides ont une forte présence des extra-centrosomes. L'activité de PJ-34 a été documentée en temps réel par imagerie confocale de cancer du sein humain vivant des cellules MDA-MB-231 transfectées avec des vecteurs codant pour fluorescent γ-tubuline, ce qui est très abondant dans les centrosomes et fluorescent histone H2B présent dans les chromosomes. Arrangements de chromosomes aberrants et de cluster-foyers γ-tubuline représentant centrosomes declustered ont été détectés dans les cellules transfectées MDA-MB-231 après le traitement avec PJ-34. Un-cluster-centrosomes supplémentaire dans les deux pôles du fuseau précédé leur mort cellulaire. Ces résultats liés for la première fois l'activité cytotoxique exclusive récemment détecté de PJ-34 dans les cellules cancéreuses humaines avec des extra-centrosomes de-regroupement dans la mitose, et l'échec de la mitose conduisant à la mort cellulaire. Selon les résultats précédents observés par imagerie confocale de cellules fixes, PJ-34 cellules cancéreuses exclusivement éradiqué avec Multi-centrosomes sans altérer les cellules normales en mitose avec deux centrosomes et les broches bi-focal. Cette activité cytotoxique de PJ-34 n'a pas été partagée par d'autres PARP1 inhibiteurs puissants, et a été observée dans PARP1 déficientes MEF extracentrosomes hébergeant, suggérant son indépendance de PARP1 inhibition. Vivez l'imagerie confocale offre un outil utile pour identifier de nouvelles molécules éradiquer les cellules en mitose.
Introduction
Phénanthrène dérivé PARP1 inhibiteurs, y compris PJ-34, ont été conçus pour protéger les cellules au repos de la mort cellulaire par apoptose induite par le consommateur d'énergie PARP1 médiation réparation de l'ADN dans des conditions de stress (accident vasculaire cérébral ou infarctus du myocarde) 1. Toutefois, récemment, nous avons découvert que PJ-34, à une concentration deux fois plus élevée que celle induisant PARP1 inhibition, peut exclusivement provoquer la mort cellulaire dans les cellules cancéreuses humaines 2,3. Le plus rapide de la prolifération de la cellule était, le plus efficace à l'éradication des cellules était. L'activité cytotoxique de PJ-34 a été attribuée à des extra-centrosomes de-regroupement dans la mitose 2. Beaucoup humain cellules cancéreuses port multicentrosomes 4,5. L'incubation des cellules cancéreuses du sein humain MDA-MB-231, qui abritent centrosomes surnuméraires, avec 20 uM PJ-34 éradiquée efficacement ces cellules au sein de 72-96 heures sans altérer les cellules au repos ou certaines cellules proliférantes bénignes abriter deux centrosomes dans la mitose
Assemblée centrosome bipolaire est cruciale pour la formation du fuseau bipolaire dans la mitose 4,5. Par conséquent, les cellules avec plus de deux centrosomes ont mis au point un mécanisme moléculaire à peine compris, le regroupement de leurs centrosomes supplémentaires à deux pôles 4-9. Le défaut de montage bipolaire de leurs centrosomes peut causer multipolaire déformée broches et aberrant chromosomes ségrégation qui arrête le cycle cellulaire en G2 / M arrestation et conduit à la mort cellulaire attribuées à l'échec de la mitose 4,5. Les mécanismes moléculaires sous-jacents à des extra-centrosomes de-clusters sont intensivement étudiées <sup> 10. La compréhension de ce mécanisme de mort permettra l'éradication exclusive des cellules cancéreuses tout en épargnant les tissus sains 5,10.
Ainsi, les composés qui activent la mort des cellules catastrophe mitotique offrent un nouveau mode de traitement du cancer sélectif, qui peut être efficace dans un large éventail de résultats de cancers.Our solides humaines suggèrent que l'imagerie confocale peut être utilisé pour identifier les molécules qui affectent extra-centrosomes regroupement en mitose 2,3, ce qui rend ces composés ciblant le cancer des candidats-médicaments.
Nous avons documenté l'activité cytotoxique de l'phénanthridiniques PJ-34 par les cellules cancéreuses humaines fixe et direct (avec une forte présence d'extra-centrosomes dans la mitose) par rapport aux cellules normales de numérisation. Une étape par étape description des procédures d'imagerie utilisées pour identifier l'activité cytotoxique de PJ-34 dans les cellules cancéreuses humaines est inclus ci-dessous.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Culture cellulaire Préparation
Cellules MDA-MB-231 ont été achetés auprès de l'ATCC (American Type Culture Collection) et stockés dans de l'azote liquide.
- Seed 10 6 cellules MDA-MB-231 cellules dans 92 mm de diamètre boîte de Pétri dans 10 ml de milieu complet contenant Dulbeco Modified Eagle Medium (DMEM), 10% de sérum de cheval, 1% de L-glutamine et 1% Penstrep-Amphotéricine B. Laisser les cellules à proliférer à environ 80-100% de confluence.
- Retirer milieu de culture de plat et le jeter.
- Laver la couche de cellules brièvement avec 0,25% (p / v) solution de trypsine-EDTA pour éliminer toute trace de sérum.
- Ajouter 2,0 ml de solution de trypsine-EDTA à plat et d'observer les cellules au microscope inversé jusqu'à ce que la couche de cellules est dispersée (généralement dans les 5 à 15 min).
- Ajouter milieu de croissance complet de 18 ml et aspirer doucement les cellules de la pipette. Transférer dans un tube.
- Centrifuger la suspension cellulaire à 1200 tours par minute.
- Re-suspendre le culot cellulaire dans 24 ml de cuLTURE moyenne.
- Ajouter 2 ml de suspension cellulaire à 35 plats à fond de verre mm (environ 25% de confluence) et placer dans un incubateur (5% de CO 2, 37 ° C).
- Solutions:
- Milieu complet pour la prolifération des cellules: DMEM avec 10% de FBS, 1% d'antibiotiques (100 unités / ml de pénicilline G, 100 pg / ml de streptomycine, la solution Pen-Strep-Ampho) et 2 mM de L-glutamine.
- Solution de trypsine-EDTA, contenant 0,25% de trypsine-EDTA.
- Plats:
92 boîtes de Pétri mm de diamètre.
35 mm de diamètre plats poly-D-lysine verre enduit culture fond.
2. Préparation des cellules pour l'imagerie confocale en direct
- Seed 2 x 10 5 cellules MDA-MB-231 cellules dans des boîtes de culture à fond de verre dans 2 ml de milieu complet tel que mentionné dans la section 1. Quand la culture cellulaire arrive à confluence de 60-70% (environ 10 5 cellules 3-4 x par boîte), procéder à la transfection.
- Transfection des cellules avec deux plasmides ENCODtion des protéines sur la fusion γ-tubuline-GFP (pour la détection fluorescente de centrosomes) et Histone-RED (H2b-RED, pour la détection par fluorescence des chromosomes) en utilisant le réactif de transfection liposomale Jet-PI, en suivant le protocole de fabrication. En bref, mélanger 2 pg de chaque plasmide dans un tube avec 100 pi de NaCl (150 mM). Mélanger le réactif de transfection (100 pi) avec 100 pi de NaCl (150 mM) dans un second tube et incuber 5 min à température ambiante (RT). Puis combiner les deux solutions, mélange (en utilisant vortex doux) et de spin-down. Incuber pendant 30 min à température ambiante.
- Au cours de l'incubation du mélange de transfection, laver les cellules une fois avec du PBS et remplacer le milieu cellulaire avec 2 ml de DMEM chaud avec aucun supplément (37 ° C).
- Incorporer délicatement le mélange de transfection des cellules dans DMEM puis retourner les cellules de l'incubateur (37 ° C, 5% CO 2) pendant 8 heures.
- Après 8 heures d'incubation, remplacer le DMEM avec 2 ml de milieu complet et incuber les cellules dans l'incubateur pendant 24hr.
- 24 heures après transfection, remplacer le milieu des cellules avec 2 ml de milieu complet contenant 20 uM PJ-34.
- Incuber les cellules pendant plus de 18 heures (37 ° C, 5% de CO 2).
- Sujet les cellules à vivre imagerie confocale pendant au moins 16 heures dans la chambre d'imagerie maintenir les cellules à 5% de CO 2 et 37 ° C.
- En parallèle, d'examiner l'efficacité de transfection 36 h après la transfection en utilisant la microscopie à fluorescence comme suit:
- Seed 2 x 10 5 cellules MDA-MB-231 cellules dans la plaque de 6 puits contenant 1 lamelle par puits dans 2 ml de milieu complet.
- Transfection des cellules comme mentionné dans les sections 2.2 à 2.5.
- 36 h après la transfection fixer les cellules transfectées montés sur une lamelle en incubation dans du méthanol froid: acétone (1:1) solution, 7 min, -20 ° C.
- Aspirer la solution de fixation et de laisser la lamelle avec les cellules montées à sec dans une hotte chimique.
- Appliquer ProLong réactif antifade d'or avec DAPI et laisser èmee lamelle de sécher dans l'obscurité pendant 6 heures.
- Examiner la lame sous microscope à fluorescence et de calculer le pourcentage de cellules transfectées (signaux rouge et vert) de la population totale de cellules (coloration de l'ADN par DAPI). Le pourcentage de transfection souhaitée est d'environ 20-40% lorsque les cellules sont comptées 100-200.
3. Paramètres techniques des confocale Paramètres du scanner d'imagerie en direct
- Scanmode XYZT; sténopé [aéré] 1,00; Zoom 3.5, résolution 8 bits; Laser DPSS 561 nm; l'argon, laser visible 488 nm; laser He / Ne visibles 633 nm; Objectif HCX PL APO CS 63X 1,40 HUILE UV; Ouverture numérique 1.4; Vitesse de numérisation 700 Hz, l'indice de réfraction de 1,52.
- Présentation de l'image 3-D ont été préparés par Imaris logiciel d'imagerie 7.0.
4. Imagerie confocale de la mitose dans les cellules fixées
- Seed 2 x 10 5 cellules MDA-MB-231 cellules mammaires cancéreuses (ATCC), les fibroblastes embryonnaires de souris normales (MEF), ou PARP1 déficient MEF (PARP - / -, préparé par le Dr Françoise Dantzer) sur des lamelles de verre en plaque de 6 puits dans 2 ml de milieu complet. Lamelles ont été lavées avec de l'éthanol à 96%, à la suite d'un lavage avec de l'eau stérile DD, séché pendant 2 heures, et placé dans chaque puits de plat à 6 puits.
- Ajouter PJ-34 (10-30 uM) au milieu et incuber les cellules pendant la période requise (habituellement jusqu'à 96 h).
- Laver les lamelles fois avec du PBS (tampon phosphate salin), et fixer les cellules à l'aide incubation dans du méthanol glacé: solution d'acétone (1:1), 7 min, -20 ° C.
- Aspirer la solution de fixation et de laisser les lamelles de sécher dans la hotte chimique (à ce stade, les lamelles peuvent être conservés dans -20 ° C pendant plusieurs semaines).
- Laver les lamelles fois avec PBST (PBS additionné de 0,1% de Tween-20) pour perméabiliser les membranes cellulaires et bloquer les cellules avec 10% NDS (Normal Sérum d'Ane) en PBS («solution de blocage») pendant 1 heure à température ambiante.
- Incuber les cellules fixes perméabilisées avec Antibodi primairees pendant 2 heures à température ambiante (pour les broches et centrosomes coloration). Les anticorps sont dilués dans la solution de blocage comme suit: anti-tubuline α (1:250 dilution) et anti-tubuline γ (dilution 1:200). Les anticorps primaires sont appliqués comme suit: appliquer 100 pl (dans une goutte) d'un mélange d'anticorps dans la solution de blocage pour chaque lamelle couvre-objet sur le couvercle de la plaque à 6 puits (le couvercle est à l'envers). Mettre doucement la lamelle sur la goutte d'anticorps, les cellules ensemencées face à la baisse. Incuber les lamelles auxquels sont confrontés les anticorps pendant 2 h à température ambiante.
- Placez les lamelles de retour dans les puits et laver les cellules 3 fois avec PBST. Ensuite, utilisez la même procédure décrite à l'article 4.6 pour le marquage de cellules sur les lamelles avec les anticorps secondaires fluorescents. Incuber les cellules sur les lamelles avec les anticorps secondaires pour 1 h, RT, dans l'obscurité. Les anticorps sont dilués dans la solution de blocage comme suit: Alexa Fluor 488 (1:1000 dilution; vert) et Alexa Fluor 568 (1:1000 dilution;rouge).
- Monter les lamelles à l'aide de prolonger réactif antifade or avec DAPI (pour la coloration des chromosomes) et incuber une nuit à température ambiante dans l'obscurité à sécher.
- Examiner les lamelles par microscopie confocale.
5. viabilité cellulaire mesurée par la production d'ATP
La production d'ATP est mesurée par un kit de dosage de détection d'ATP luminescent.
- Seed les cellules en plaque de 96 puits, environ 20.000 cellules dans 800 pi de milieu dans chaque puits. Trois puits vierges devraient être utilisés pour la détermination de la luminescence de fond du milieu.
- Préparer ATP série de dilution standard d'environ 10 et 100 micromètres et garder sur la glace.
- Ajouter 50 l de détergent dans chaque puits et agiter la plaque pendant 5 min dans un agitateur orbital, à 700 rpm.
- Reconstituer chaque flacon du «substrat lyophilisé» avec 5 ml de «tampon de substrat» dans le kit.
- Ajouter 50 ul de la solution de substrat reconstitué dans les puits, et secouezla plaque pendant 5 min sur agitateur orbital, à 700 rpm.
- Conservez la plaque dans l'obscurité pendant 10 min.
- Mesurer luminescence de chaque puits par ELISA lecteur de microplaques.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 est un stable soluble dans l'eau phénanthridiniques 1 (figure 1). Nos résultats précédents ont révélé la mort cellulaire et de-regroupés extra-centrosomes dans plusieurs types de cellules cancéreuses multi-centrosomales fixes qui ont été traités avec PJ-34. En revanche, les cellules proliférantes normales n'ont pas été altérées 2,3. Centrosomes ont été identifiés par un double marquage avec des anticorps dirigés contre centrine1 et γ-tubuline dans les cellules extra-centrosomales fixes 2.
Ici, l'activité cytotoxique de PJ-34 a été documentée dans ces cellules extra-centrosomales direct à temps réel en utilisant la microscopie confocale en direct. Vivre le cancer du sein humain des cellules MDA-MB-231, qui ont une fréquence élevée (> 50%) des extra-centrosomes 4,5, ont été numérisées pour au moins 16 heures par imagerie confocale porté sur les cellules transfectées avec γ-tubuline-GFP ( marquage fluorescent de γ-tubuline foyers 2) et histone H2b-RED (fluorescent Labeling de chromosomes). Six à dix cellules transfectées vivants ont été analysés en parallèle dans chaque expérience. Double immunomarquage des foyers γ-tubuline dans les cellules transfectées avec centrin1 était techniquement impossible.
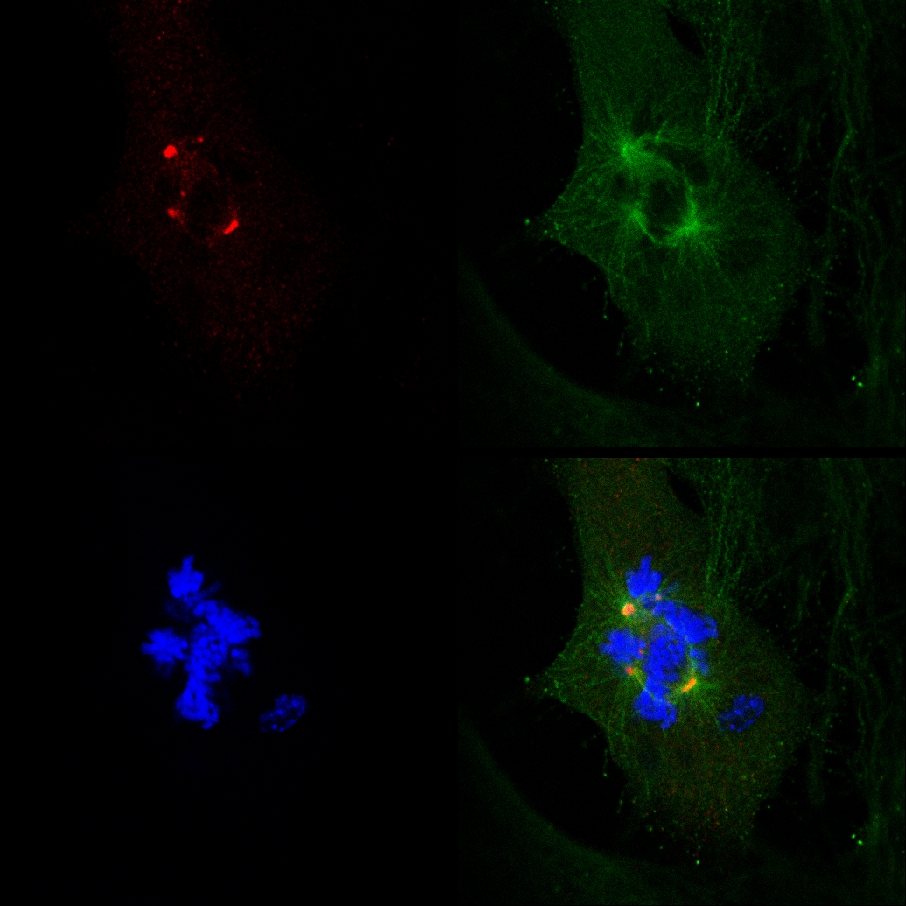
Dispersés γ-tubuline foyers et l'arrangement des chromosomes aberrants ont été rarement détectés dans les cellules non traitées choisis au hasard MDA-MB-231 en mitose. Regroupement bifocale de foyers γ-tubuline, représentant des extra-centrosomes regroupement bifocale, a été documentée dans la majorité des cellules vivantes non traités MDA-MB-231 (figure 2), en revanche, centrosomes et la disposition aberrante des chromosomes non-cluster ont été détectés en direct transfectées MDA-MB-231 cellules incubées avec PJ-34 (20 M), et la mitose dans ces cellules terminés par la mort cellulaire (Figure 3). Ces cellules ont été incubées avec PJ-34 pour les 18 - 24 heures avant la numérisation et pour supplémentaire de 16 heures lors de la numérisation (Figure 3). Les documents en temps réel de la cellule death lors de la mitose appuient fermement une corrélation positive définie précédemment entre le nombre de cellules malignes humaines avec des broches multipolaire dans la mitose et la proportion de la mort cellulaire dans des cellules incubées avec PJ-34 (20 pM) 2.
PJ-34 agit comme un inhibiteur puissant PARP1 1. Nous avons donc examiné la possibilité de PARP1 inhibition provoquant la mort cellulaire associée à l'insuffisance de la mitose. Contrairement à l'imagerie en direct, l'imagerie des cellules MDA-MB-231 fixes a permis l'examen d'une grande population de cellules dans les cultures cellulaires, permettant ainsi une analyse statistique des effets des inhibiteurs PARP1 dans une variété de lignées de cellules cancéreuses humaines. L'activité de PJ-34 a été comparée à l'activité d'autres puissants, non-phénanthrène PARP1 inhibiteurs dans des conditions normales ou PARP1 cellules déficientes (c. normal et PARP1 (- / -) fibroblastes embryonnaires de souris (MEF)) (figure 4). PARP1 déficientes MEF port multi-centrosomes dans la mitose, mais ils ne sontles cellules tumorales t 11. Ces cellules ont été préparées par le Dr Françoise Dantzer, Strasbourg, France.
Fixe normal et PARP1 (- / -) MEF ont été immunomarquées pour α-et γ-tubuline qui étiqueté leurs broches et centrosomes, respectivement, tel que rapporté avant 2. Certains des cultures cellulaires examinés ont été traités avec de puissants, non-phénanthrène PARP1 inhibiteurs PJ-34 ou autre, y compris ABT-888 et AG01469, qui inhibent l'activité enzymatique de PARP1 et BSI-201, un composé qui atténue apparemment PARP1 liaison à entaillé ADN 12-14. Aucun des PARP1 inhibiteurs testés avec facultés affaiblies MEF normale à des concentrations inhibant l'activité PARP1 (Figure 4). En revanche, PJ-34 dépendante de la dose a provoqué un-regroupement des γ-tubuline foyers, la distorsion de broches et de la mort cellulaire dans PARP1 (- / -) MEF (figures 4A et B). Cela n'a pas été observé dans MEF normale traités par PJ-34 (Figure 4B) ou dans PARP1 (- / -) MEF traité de non-phenenthrene PARP1 inhibiteurs ABT-888 ou AG014699 (figure 4C). Il convient de noter que PJ-34 à des concentrations dépassant 20 uM ne altèrent MEF normale, bien que MEF normale étaient plus résistants à l'activité PJ-34 que PARP1 (- / -) MEF.
Le fait que PJ-34 éradiqué PARP1 (- / -) MEF malgré leur PARP1 carence, et la corrélation entre la formation de broches multi-focale et l'éradication de la cellule dans PARP1 (- / -) MEF incubées avec PJ-34 à des concentrations supérieures celles requises pour PARP1 inhibition, n'étaient pas compatibles avec un lien de causalité entre des extra-centrosomes de-clustering dans PARP1 (- / -) du MEF et PARP1 inhibition (figure 4A). L'activité cytotoxique de PJ-34 PARP1 (- / -) MEF pourrait être mieux expliqué par son activité comme un extra-centrosomes l'agent de-clustering dans les cellules multi-centrosomales 2 ( Renseignements supplémentaires "/> Figure 3. Une documentation confocal d'imagerie en direct de la mort cellulaire dans une MDA-MB-231 cellules choisies au hasard dans la mitose avec ONU-cluster γ-tubuline marqués des extra-centrosomes. Cellules MDA-MB-231 ont été incubées avec PJ-34 pendant 24 heures avant de numériser et pendant les 16 heures de l'imagerie confocale en direct. PJ-34 a été appliqué 24 heures après transfection avec des vecteurs exprimant γ-tubuline-GFP (Green, foyers γ-tubuline et centrosomes) et H2b-RED (rouge, chromosomes). 
Figure 1. Le phénanthridine PJ-34: N-(6-oxo-5 ,6-dihydro-phenanthridin-2-yl)-N, N-diméthyl-acétamide. 
Figure 2. Regroupement bi-focal de l'extra-centrosomes dans un live MDA-MB-231 cellules choisis au hasard dans la mitose. A. Panneau supérieur: centrosomes marqué dans un live MDA-MB-231 cellules sélectionnées au hasard transfectées avec γ-tubuline-GFP Panneau inférieur:. Réarrangements chromosomiques lors de la mitose dans une MDA-MB-231 cellules sélectionnées au hasard transfectées avec histone H2b-RED. B. mitose Bi-focal avec cluster supplémentaire centrosomes-identifiés dans une secte choisis au hasardrée MDA-MB-231 cellules. Les cellules ont été transfectées par deux γ-tubuline-GFP (étiquetage γ-tubuline foyers, vert) et histone H2b-rouge (marquage de chromosomes; rouge). 48 heures après transfection, les cellules ont été exposées à une imagerie confocale en direct pendant 16 heures. Six cellules ont été analysés en parallèle dans chaque expérience. Quatre expériences différentes ont été réalisées. Voir aussi Renseignements supplémentaires. Cliquez ici pour agrandir la figure . 
Figure 3. Extra-centrosomes de-clustering précédé la mort cellulaire en direct MDA-MB-231 cellules traitées avec PJ-34. A choisi au hasard en direct MDA-MB-231 cellules en mitose avec centrosomes dispersés (1 er cadre de gauche) terminés par la mort cellulaire ( 2 ème et 3 ème cadres). Ce CEll a été sélectionné au hasard dans une culture de cellules incubées pendant 24 heures avec PJ-34 (20 M) appliquée 24 heures après transfection avec des vecteurs exprimant γ-tubuline-GFP (étiquetage foyers γ-tubuline y compris centrosomes, vert) et histone H2b-RED ( chromosomes étiquetage; rouge). La cellule a été scannée pendant 16 heures par imagerie confocale en direct. Six cellules ont été analysés en parallèle dans chaque expérience. Trois expériences différentes ont été réalisées. Voir aussi Renseignements supplémentaires. Cliquez ici pour agrandir la figure . 
Figure 4. Une activité cytotoxique de PJ-34 dans PARP1 (- / -) des fibroblastes embryonnaires de souris. A. (gauche), le pourcentage de broches multi-focale calculée en nOrmal (ligne noire) et PARP1 - / - (ligne grise) MEF, incubées pendant 48 heures avec PJ-34 aux concentrations indiquées. Le pourcentage de broches multi-polaire a été calculé sur un total de 20 broches détectée dans 3 expériences différentes. (À droite) réduit la survie des cellules détectée dans des cultures cellulaires incubées pendant 72 heures avec PJ-34 (20 M) par rapport à la survie des cellules non traitées de contrôle (normal ligne noire () et PARP1 - / - (ligne grise) MEF). La survie des cellules a été analysée par l'ATP production des cellules (protocole 5). Les valeurs moyennes de 4 mesures pour chaque lignée cellulaire dans 3 expériences différentes sont présentées B. Broches en choisis au hasard fixé normal et PARP1 -. / - MEF dans la mitose, non traité (témoin) ou incubées avec PJ-34 pendant 48 heures au indiqué concentrations. PJ-34 a provoqué multipolaire broches. Les cellules ont été fixées, perméabilisées et immunomarquées pour α-et γ-tubuline (étiquetage vert de broches et étiquette rouge de centrosomes, respectivement). Les chromosomes ont été marquées avec réactif DAPI (bleu). . Résultats représentatifs de 3 expériences différentes C. PARP1 inhibiteurs non-phénanthrène puissants n'ont pas affecté centrosomes regroupement dans PARP1 (- / -) MEF. Broches de hasard choisi normale et PARP1 - / - MEF sont présentés; MEF non traité (témoin) ou MEF traitées pendant 48 heures avec des inhibiteurs de PARP non-phénanthrène, AG01469 (20 M) ou ABT888 (20 M). Les chromosomes sont étiquetés avec le réactif DAPI (bleu). Des résultats similaires ont été obtenus dans 3 expériences différentes. Cliquez ici pour agrandir la figure .
Cliquez ici pour agrandir la figure . 
Cliquez ici pour agrandir la figure .
Figure 2B. Figure supplémentaire. A choisi au hasard MDA-MB-231 cellules en anaphase avec bi-focal cluster γ-tubuline marqués extra-centrosomes a été balayé pendant 16 heures par imagerie confocale en direct, 48 heures après avoir été transfectées avec des vecteurs exprimant γ-tubuline-GFP et H2B-RED (étiquetage foyers γ-tubuline et centrosomes dans les cellules fixes (vert) et l'étiquetage des histones H2b des chromosomes (rouge), respectivement).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Vivez l'imagerie confocale a fourni une documentation en temps réel de l'effet cytotoxique de PJ-34 dans les cellules multi-centrosomales en direct pendant la mitose (Figure 3 et les informations supplémentaires). Ce fut la première documentation en direct attribuant la cytotoxicité de PJ-34 dans les cellules humaines de cancer de centrosomes supplémentaires de-regroupement et la mort cellulaire, suggérant l'induction de la mort cellulaire de la catastrophe mitotique par PJ-34 5-9. En revanche, le regroupement bi-focal de centrosomes surnuméraires a été observée dans les cellules vivantes non traités MDA-MB-231 en mitose normale avec bi-Focali centrosomes supplémentaires en cluster (Figure 2, des informations supplémentaires).
Selon ces résultats, vivent imagerie confocale de cellules transfectées pourrait être utile pour la détection de la translocation de protéines impliquées dans extra-centrosomes regroupement dans les cellules transfectées pendant la mitose 5,9,15,16. L'identification des protéines affectées par PJ-34 en multi-CentrosOmal cellules peuvent fournir quelques indices pour comprendre les mécanismes de la mort activés par des extra-centrosomes de-clustering.
L'avantage de l'imagerie confocale en direct en fournissant des informations en temps réel au cours de la mitose dans les cellules vivantes, est également lié à plusieurs limitations. Le nombre de cellules numérisées par l'expérience est limitée. Par conséquent chances de détecter des cellules en mitose sont faibles, et plusieurs expériences répétitives sont nécessaires pour une documentation en temps réel de la mitose dans les cellules numérisées. En outre, le succès dépend fortement de haute efficacité de la transfection de cellules avec des vecteurs exprimant des protéines marquées. Ainsi, en dépit d'être fiable, l'imagerie confocale en direct prend du temps et exige des travailleurs acharnés très expérimentés.
En comparaison, immunocytochimie et imagerie confocale de cellules fixes permettre l'examen d'un grand nombre de cellules par expérience, qui est nécessaire pour une analyse statistique fiable. Nous avons utilisé cette méthode pour comparer l'effets de PJ-34 en mitose des cellules bénignes normales à ses effets dans les cellules cancéreuses en mitose avec centrosomes surnuméraires 2,3. De même, cette méthode a été utilisée pour comparer les effets de la PJ-34 à celle de PARP1 inhibiteurs non-phénanthrène dans PARP (- / -) MEF abriter des extra-centrosomes (cellules à forte présence de multi-centrosomes 11) (figure 4).
En résumé, nos résultats montrent l'avantage de combiner l'information en temps réel la plus précieuse fournie par l'imagerie confocale de cellules vivantes en mitose (figures 2 et 3) avec cytochimie et l'analyse confocale de cellules fixes. Une combinaison de ces méthodes pourrait être utile pour identifier l'activité cytotoxique de petites molécules qui, comme PJ-34, ciblant des mécanismes spécifiques cruciale pour la survie de la cellule. La dépendance unique de nombreuses cellules cancéreuses humaines sur le regroupement bi-polaire extra-centrosomes pour leur prolifération et survivantal rend PJ-34 un candidat possible pour le traitement du cancer.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Les auteurs déclarent qu'ils n'ont aucun intérêt financier concurrents.
Acknowledgments
Les sources de financement de cette recherche: un fonds commun de la société de l'Université de Tel Aviv de transfert technologique, Ramot et le Centre Sheba-Medical (M. CA et SI)., ICRF - Fondation de la recherche sur le cancer israélien (M. CA.) Et la Fondation israélienne des sciences ( SI).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}