

Summary
Цитотоксической активности фенантридин PJ-34 в раковых клетках митоз было зафиксировано в реальном времени с помощью конфокальной микроскопии живых. PJ-34 искоренили рака молочной железы человека MDA-MB-231 клетки укрывательство особо центросомам в митоза. В отличие от обычных бифокальные митоза, экстра-центросомам не были сосредоточены в двух полюсах веретена в присутствии PJ-34.
Abstract
Фенантрен производные действуют как мощные ингибиторы PARP1 помешало бифокальные кластеризации нештатных центросомам в мульти-центросомной человеческих раковых клеток в митозе. Фенантридин PJ-34 был самым мощным молекулы. Декластеризация экстра-центросомам вызывает отказ митотических и гибель клеток в многоквартирных центросомной клеток. Большинство твердых опухолей человека имеют высокую возникновения экстра-центросомам. Деятельность PJ-34 было зафиксировано в реальном времени с помощью конфокальной микроскопии живых рака молочной железы человека MDA-MB-231 клеток, трансфицированных векторов, кодирующих для люминесцентных γ-тубулина, который очень распространен в центросомам и для люминесцентных гистонов в настоящее H2b хромосом. Аномальные механизмы хромосом и де-кластерной γ-тубулина очагов представляющих declustered центросомам были обнаружены в Трансфицированные MDA-MB-231 клеток после лечения с PJ-34. Un-кластерной экстра-центросомам в два полюса шпинделя предшествовало их гибели клеток. Эти результаты связаны FOR впервые недавно обнаружены эксклюзивные цитотоксической активности PJ-34 в раковых клетках человека с очень центросомам де-кластеризации в митоза и митотического недостаточности приводит к гибели клеток. В соответствии с предыдущими выводами наблюдали конфокальной микроскопии фиксированных клетках, PJ-34 исключительно искоренить раковые клетки с несколькими центросомам не повреждая нормальные клетки митоз с двумя центросомам и бифокальные шпинделей. Это цитотоксической активности PJ-34 не разделяют другие мощные ингибиторы PARP1, и наблюдался в PARP1 дефицитный MEF укрывательстве extracentrosomes, предлагая свою независимость PARP1 торможения. Живите конфокальной микроскопии предложили полезным инструментом для выявления новых молекул искоренения клетки во время митоза.
Introduction
Фенантрен полученных PARP1 ингибиторов, в том числе PJ-34, предназначены для защиты покоящихся клеток от апоптоза индуцированных энергоемкие PARP1 опосредованной репарации ДНК в условиях стресса (инсульт или инфаркт миокарда) 1. Однако в последнее время мы обнаружили, что PJ-34, в два раза большей концентрации, чем вызывающие PARP1 торможения, может только вызвать гибель клеток в раковые клетки человека 2,3. Чем быстрее пролиферацию клетки было, тем более эффективным искоренением клетки было. Цитотоксической активности PJ-34 был приписан к дополнительным центросомам-де-кластеризации в митозе 2. Многие человеческие раковые клетки гавани multicentrosomes 4,5. Инкубация человека клеток рака молочной железы MDA-MB-231, который питает сверхштатный центросомы, с 20 мкМ PJ-34 эффективно искоренить эти клетки в течение 72-96 часов без ухудшения покоящихся клеток или некоторые доброкачественные пролиферативные клетки несущий две центросомы в митозе
Биполярное центросоме Ассамблеи имеет решающее значение для формирования биполярного веретена в митозе 4,5. Таким образом, клетки, имеющие более двух центросомы разработали едва понимать молекулярные механизмы, кластеризации их дополнительное центросомы на два полюса 4-9. Отказ биполярной сборки их центросомам может вызвать искаженное многополярного шпинделей и сегрегации хромосом аберрантными что аресты клеточного цикла в G2 / M ареста, и приводит к гибели клеток связано с недостаточностью митотического 4,5. Молекулярные механизмы, лежащие в основе экстра-де-центросомам кластеризации интенсивно исследуются <вир> 10. Понимание этого механизма смерти позволит эксклюзивные искоренение раковые клетки, щадя здоровые ткани 5,10.
Таким образом, соединения, которые активируют митотической катастрофы гибель клеток предложить новый способ селективной терапии рака, который может быть эффективным в широком диапазоне человеческого твердых cancers.Our результаты показывают, что конфокальной микроскопии может быть использован для идентификации молекул, влияющие экстра-центросомы кластеризации митоза 2,3, оказания этих соединений рака таргетинга лекарств-кандидатов.
Мы документировали цитотоксической активности фенантридин PJ-34 при сканировании фиксированных и живых раковых клеток человека (с высокими возникновения экстра-центросомы в митоз) по сравнению с нормальными клетками. Шаг за шагом, описание процедуры отображения используются для определения цитотоксической активности PJ-34 в человеческих раковых клеток приведен ниже.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Подготовка клеточной культуры
MDA-MB-231 клетки были получены от АТСС (Американской коллекции типовых культур) и хранили в жидком азоте.
- Семенной 10 6 MDA-MB-231 клеток в 92 мм чашки Петри в 10 мл полной среды, содержащей Dulbeco изменения среду Игла (DMEM), 10% лошадиной сыворотки, 1% L-глутамина и 1% Penstrep-Amphotericine B. Разрешить клетки разрастаться до 80-100% слияния.
- Удалить из культуральной среды блюдо и отказаться.
- Промыть слой клеток вкратце с 0,25% (вес / объем) Трипсин-ЭДТА, чтобы удалить все следы сыворотки.
- Добавить 2,0 мл трипсина-EDTA решение блюдо и наблюдать клетки инвертированного микроскопа, пока слой клеток не разгоняется (обычно в пределах от 5 до 15 мин).
- Добавить 18 мл полной среды роста и осторожно аспирации клеток с помощью пипетки. Трансфер в трубке.
- Центрифуга клеточной суспензии при 1200 оборотах в минуту.
- Повторное приостановить осадок клеток в 24 мл Culture среды.
- Добавить 2 мл клеточной суспензии до 35 мм со стеклянным дном блюда (около 25% слияния) и место в инкубаторе (5% CO 2, 37 ° C).
- Решения:
- Полной среде в течение пролиферацию клеток: DMEM с 10% FBS, 1% антибиотиков (100 ед / мл пенициллина, 100 мкг / мл стрептомицина, Pen-Strep-Ampho раствор) и 2 мМ L-глутамина.
- Трипсин-ЭДТА, содержащем 0,25% трипсин-ЭДТА.
- Блюда:
Диаметром 92 мм чашки Петри.
Диаметром 35 мм поли-D-лизин покрытием блюд со стеклянным дном культуры.
2. Подготовка Клетки для живых конфокальной микроскопии
- Семенной 2 х 10 5 MDA-MB-231 клеток в культуре со стеклянным дном блюда в 2 мл полной среды, как указано в разделе 1. Когда культура клеток достигает слияния 60-70% (около 3-4 х 10 5 клеток на чашку), переходите к трансфекции.
- Трансфекции клеток с двух плазмид ENCODING слитых белков γ-тубулина-GFP (для флуоресцентной детекции центросом) и гистонов-RED (H2b-красные, для флуоресцентной детекции хромосом) с помощью липосомальной трансфекции реагента Jet-PI, после изготовления протокола. Вкратце, смешать с 2 мкг каждой плазмиды в пробирке с помощью 100 мкл NaCl (150 мМ). Смешать реагента для трансфекции (100 мкл) с помощью 100 мкл NaCl (150 мМ) в вторую трубку, и инкубируют 5 мин при комнатной температуре (RT). Затем объединить два решения, смеси (с использованием мягкого вихря) и спин-вниз. Инкубируют в течение 30 мин при комнатной температуре.
- При инкубации трансфекции смесь, промыть клетки один раз ЗФР и заменить клеточной среде с 2 мл теплой DMEM без каких-либо добавок (37 ° C).
- Аккуратно добавить трансфекции смесь клеток в DMEM, а затем вернуть клеткам в инкубатор (37 ° C, 5% СО 2) в течение 8 часов.
- Через 8 часов инкубации заменить DMEM с 2 мл полной среды и инкубируют клетки в инкубаторе в течение 24HR.
- Через 24 часа после трансфекции, замените среда клеток с 2 мл полной среды, содержащей 20 мкМ PJ-34.
- Инкубируйте клетки еще 18 ч (37 ° C, 5% СО 2).
- Тема клетки жить конфокальной микроскопии в течение не менее 16 часов в камере изображений поддержания клеток на 5% СО 2 и 37 ° C.
- Параллельно, изучить эффективность трансфекции 36 ч после трансфекции с использованием флуоресцентной микроскопии следующим образом:
- Семенной 2 х 10 5 MDA-MB-231 клеток в 6-луночный планшет, содержащий 1 покровное на лунку в 2 мл полной среды.
- Трансфекции клеток, как указано в разделах 2.2-2.5.
- 36 ч после трансфекции зафиксировать трансфицированных клеток установлен на покровное инкубацией в холодный метанол: ацетон (1:1), 7 мин, -20 ° С.
- Аспирируйте фиксирующем растворе и пусть покровное с установленными клетки для просушки в химической капотом.
- Применить продлить реагента золото Antifade с DAPI и пусть йэлектронной покровного сушить в темноте в течение 6 часов.
- Изучите скользят под флуоресцентным микроскопом и вычислить процент трансфекции клеток (красный и зеленый сигналы) от общей популяции клеток (ДНК окрашивания DAPI). Желаемый трансфекции процент составляет около 20-40% при 100-200 клетки подсчитывают.
3. Технические параметры жить конфокальной настройки сканеров
- XYZT режиме сканирования; Пинхол [воздушный] 1,00; Увеличить 3,5; разрешение 8 бит; Лазерная DPSS 561 нм; Аргон, видимый лазер 488 нм; лазерная / Ne видимой длине волны 633 нм; Цель HCX PL APO CS 63X 1,40 УФ маслом; числовая апертура 1,4; Скорость сканирования 700 Гц, показатель преломления 1,52.
- Изображение 3-D презентация были подготовлены Imaris обработки изображений 7.0.
4. Конфокальной микроскопии митоза в фиксированных клетках
- Семенной 2 х 10 5 MDA-MB-231 клеток рака молочной железы (АТСС), нормальные мышиных эмбриональных фибробластов (MEF) или PARP1 дефицитный MEF (PARP - / -, подготовленной д-ром Франсуаза Dantzer) на покровных стеклах в 6-луночный планшет в 2 мл полной среды. Покровные слои промывают 96% этанолом, после стирки путем стерильной водой DD, сушат в течение 2 ч и помещали в каждую лунку 6-луночный планшет.
- Добавить PJ-34 (10-30 мкм) в среду и инкубировать клетки в течение требуемого периода (обычно до 96 ч).
- Промыть покровные один раз ЗФР (забуференный фосфатом физиологический раствор) и зафиксировать клетки с использованием инкубации в охлажденном льдом метанол: ацетон (1:1), 7 мин, -20 ° С.
- Аспирируйте фиксирующем растворе и пусть покровные высохнуть в капот химическая (на данном этапе, покровные может храниться в -20 ° С в течение нескольких недель).
- Промыть покровные один раз PBST (PBS, дополненную 0,1% Твин-20), чтобы проницаемость клеточной мембраны и блокировать клетки с 10% NDS (нормальный осел сыворотка) в PBST ('блокирующий раствор) в течение 1 часа при комнатной температуре.
- Выдержите проницаемыми фиксированные клетки с первичными Antibodiы в течение 2 ч при комнатной температуре (для окрашивания шпинделей и центросомы). Антител разводят в блокировании решения следующим образом: анти-α тубулина (1:250 разведение) и анти-γ тубулина (разведение 1:200). Первичные антитела применяются следующим образом: применяют 100 мкл (по капле) смеси из антител в блокирующем растворе для каждого покровное на 6-луночный планшет крышки (крышки вверх дном). Осторожно положите покровное на каплю антитела, отобранные клетки, стоящие перед падением. Инкубируйте покровные сталкивается с антителами в течение 2 ч при комнатной температуре.
- Поместите покровные обратно в скважины и промыть клетки 3 раза PBST. Затем с помощью той же процедуры, описанной в 4,6 мечения клеток на покровных стеклах с флуоресцентным вторичных антител. Инкубируйте клетки на покровных стеклах с вторичными антителами в течение 1 часа, РТ, в темноте. Антитела разводили в блокирующем растворе следующим образом: Alexa Fluor 488 (разведении 1:1000; зеленый) и Alexa Fluor 568 (разведении 1:1000;красный).
- Установите покровные использованием реагента продлить Золото Antifade с DAPI (для окрашивания хромосом) и инкубировать в течение ночи при комнатной температуре в темном, чтобы высохнуть.
- Изучите покровные стекла с помощью конфокальной микроскопии.
5. Жизнеспособность клеток Измеряется продукции АТФ
АТФ измеряется с помощью люминесцентного обнаружения набора для анализа АТФ.
- Семенной клеток в 96-луночный планшет, приблизительно 20000 клеток в 800 мкл среды в каждую лунку. Три контрольных лунок следует использовать для определения фонового свечения среды.
- Подготовка АТФ стандартной серии разведений от приблизительно 10 мкм до 100 мкм и держать на льду.
- Добавить 50 мкл моющего средства в каждую лунку и встряхните пластине в течение 5 мин в орбитальный шейкер, 700 оборотов в минуту.
- Разведите каждый флакон 'Субстрат лиофилизированного' с 5 мл 'субстратный буфер "в наборе.
- Добавить 50 мкл восстановленного раствора субстрата в лунки и встряхиваютпластине в течение 5 мин на орбитальном шейкере, 700 оборотов в минуту.
- Храните в темном пластину в течение 10 мин.
- Измерение люминесценции каждой лунки на микропланшет-ридера ELISA.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
PJ-34 является стабильным водорастворимые фенантридин 1 (рис. 1). Наши предыдущие результаты показали гибель клеток и де-кластерной экстра-центросомам в нескольких видов основных несколькими центросомной раковые клетки, которые лечились PJ-34. В отличие от нормальных пролиферирующих клеток не нарушается 2,3. Центросомы были определены двойной маркировки с антителами, направленными против centrine1 и γ-тубулина в фиксированном экстра-центросомной клетки 2.
Здесь цитотоксической активности PJ-34 было зафиксировано в эти живые экстра-центросомной клетки в реальном времени с использованием живых конфокальной микроскопии. Живите рака молочной железы человека MDA-MB-231 клеток, которые имеют высокий возникновение (> 50%) экстра-центросомам 4,5, было проверено на наличие, по меньшей мере 16 часов по конфокальной микроскопии сосредоточены на трансфекции клеток с γ-тубулина-GFP ( флуоресцентного мечения γ-тубулина очаги 2) и с гистонов H2b-RED (флуоресцентная ЛабеЛинг хромосом). От шести до десяти живых трансфицированные клетки сканировании параллельно в каждом эксперименте. Двухместный иммунного из γ-тубулина очагов в трансфицированных клетках с centrin1 был технически невозможно.
Дисперсных γ-тубулина очагов и аберрантным расположением хромосом редко обнаружен в случайно выбранных необработанной MDA-MB-231 клеток в митоза. Бифокальный кластеризации γ-тубулина очагов, представляющих экстра-центросомам бифокальной кластеризации, было зафиксировано в большинстве живых необработанных MDA-MB-231 клеток (рис. 2), в отличие, ООН-кластерной центросомам и аберрантным расположением хромосом были обнаружены в живых трансфицированными MDA-MB-231 клетки инкубировали с PJ-34 (20 мкм), и митоза в этих клетках закончилась гибелью клеток (рис. 3). Эти клетки инкубировали с PJ-34 в течение 18 - 24 часов до начала сканирования и еще 16 ч во время сканирования (рис. 3). В режиме реального времени документации клеточного де-ате при митозе решительно поддерживает определенные ранее положительная корреляция между количеством человеческих злокачественных клеток с многополярного веретена в митозе и процент гибели клеток в клетки, инкубированные с PJ-34 (20 мкМ) 2.
PJ-34 действует как мощный ингибитор PARP1 1. Таким образом, мы рассмотрели возможность PARP1 ингибирования вызывая гибель клеток, связанных с митотической недостаточности. В отличие от живого изображения, изображение основных MDA-MB-231 клетки включен рассмотрении большой популяции клеток в культурах клеток, тем самым позволяя статистического анализа эффектов PARP1 ингибиторов в различных линий раковых клеток человека. Деятельность PJ-34 по сравнению с деятельностью других сильнодействующих, не фенантрена PARP1 ингибиторов в нормальном или PARP1 дефицитных клеток (т.е. нормальной и PARP1 (- / -) мышиных эмбриональных фибробластов (MEF)) (рис. 4). PARP1 дефицитный MEF порт мульти-центросомы в митозе, но они неТ-клетки опухоли 11. Эти клетки были получены доктором Франсуаза Dantzer, Страсбург, Франция.
Исправлена нормальной и PARP1 (- / -) MEF были immunolabeled для α-и γ-тубулина, что пометил свой центросомам шпинделей и, соответственно, как сообщалось ранее 2. Некоторые из рассматриваемых культур клеток обрабатывали PJ-34 или других сильнодействующих, не фенантрен PARP1 ингибиторы, включая АВТ-888 и AG01469, которые ингибируют ферментативную активность PARP1 и BSI-201, соединение, которое по-видимому ослабляет PARP1 связывания порезал ДНК 12-14. Ни один из проверенных PARP1 ингибиторы нарушением нормального MEF в концентрациях, ингибирующих PARP1 деятельности (рис. 4). В противоположность этому, PJ-34 в зависимости от дозы вызвало ООН-кластеризации γ-тубулина очагов, искажение шпинделей и гибель клеток в PARP1 (- / -) MEF (4A и B). Это не наблюдалось в нормальных MEF получавших PJ-34 (Figurэлектронной 4В) или в PARP1 (- / -) MEF получавших не-phenenthrene PARP1 ингибиторы ABT-888 или AG014699 (фиг.4С). Следует отметить, что PJ-34 в концентрации более 20 мкМ же ухудшать нормальной MEF, хотя нормального MEF были более стойкими к PJ-34 активностью, чем PARP1 (- / -) MEF.
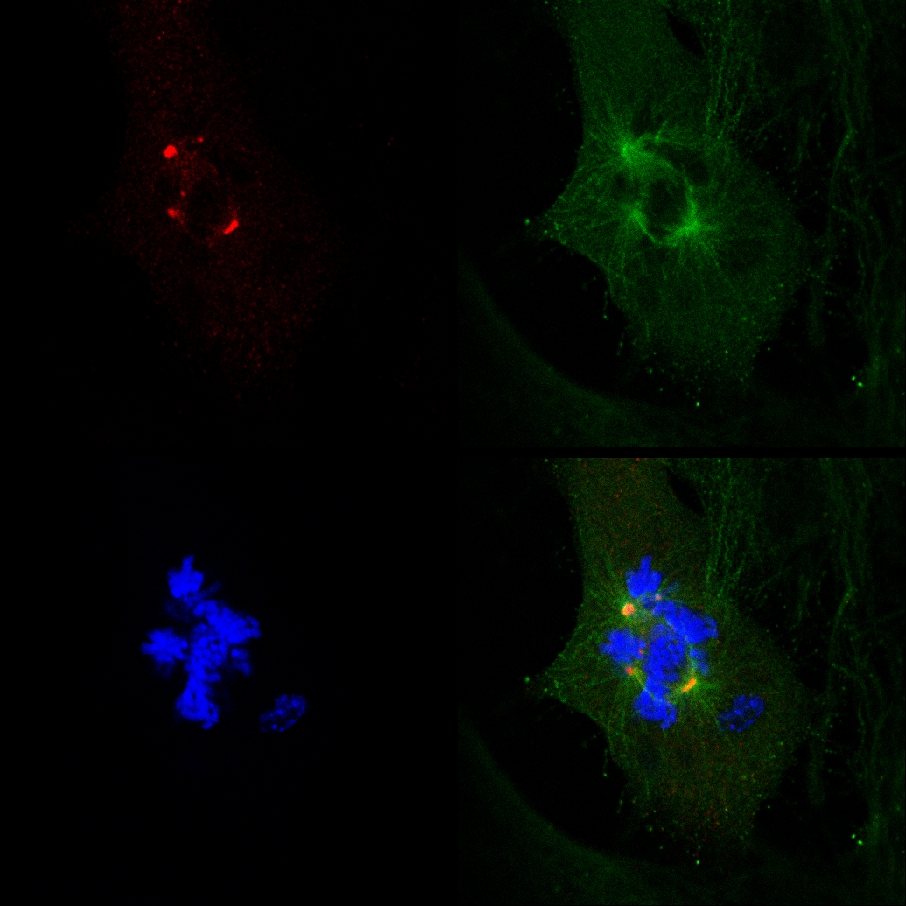
Тот факт, что PJ-34 искоренили PARP1 (- / -) MEF, несмотря на их дефицит PARP1, и корреляция между образованием многофокусной шпинделей и искоренения в клетке PARP1 (- / -) MEF инкубировали с PJ-34 в концентрациях, превышающих те, которые требуются для PARP1 торможение, не соответствовали причинной связи между дополнительными центросомам-де-кластеризации в PARP1 (- / -) MEF и PARP1 торможения (рис. 4а). Цитотоксической активности PJ-34 в PARP1 (- / -) MEF может быть лучше объяснить свою деятельность в качестве дополнительной центросомам-де-кластеризации агента в многоквартирных центросомной клетки 2 ( Дополнительная информация "/> Рисунок 3. Жить конфокальной документации визуализации клеточной гибели в случайно выбранные MDA-MB-231 клетки в митозе с ООН-кластерной γ-тубулина помечены экстра-центросомам. MDA-MB-231 клетки инкубировали с PJ-34 в течение 24 ч перед сканированием и в течение 16 ч живых конфокальной микроскопии. PJ-34 был применен 24 часа после трансфекции векторов, экспрессирующих γ-тубулина-GFP (зеленый, γ-тубулина очагов и центросомам) и H2b-RED (красный, хромосом). 
Рисунок 1. Фенантридин PJ-34: N-(6-оксо-5 ,6-дигидро-фенантридин-2-ил)-N, N-диметилацетамида. 
Рисунок 2. Би-координационного кластеризации экстра-центросомам в случайно выбранных живой MDA-MB-231 клетки в митозе. А. Верхняя панель: Маркированный центросомам в случайно выбранных живой MDA-MB-231 ячейки трансфецированы γ-тубулина-GFP Нижняя панель:. Хромосома повторного договоренностей во время митоза в случайно выбранные MDA-MB-231 ячейки трансфецированы гистонов H2b-RED. B. бифокальные митоза с кластерными экстра-центросомам определены в случайно выбранные культаUred MDA-MB-231 клетки. Клетки трансфицировали обеими γ-тубулина-GFP (маркировка γ-тубулина очагов; зеленый) и гистонов H2b-RED (маркировка хромосом; красный). 48 ч после трансфекции клетки подвергали воздействию живого конфокальной микроскопии в течение 16 часов. Шесть клетки сканировании параллельно в каждом эксперименте. Четыре различных экспериментов. См. также дополнительную информацию. Нажмите здесь, чтобы увеличить рисунок . 
Рисунок 3. Дополнительные центросомам-де-кластеризации предшествовала гибель клеток в живом MDA-MB-231 клеток, обработанных PJ-34. Случайно выбранных живой MDA-MB-231 клетки в митозе с разбросанными центросомам (1-й кадр слева) закончилась гибелью клеток ( 2-й и 3-й кадры). Это CELL было случайно выбранных в культуре клеток инкубировали в течение 24 часов с PJ-34 (20 мкМ) использовано 24 часа после трансфекции векторов, экспрессирующих γ-тубулина-GFP (маркировка γ-тубулина очагов в том числе центросомы; зеленый) и гистонов H2b-RED ( маркировки хромосом; красный). Ячейка была отсканирована в течение 16 часов живой конфокальной микроскопии. Шесть клетки сканировании параллельно в каждом эксперименте. Три различных экспериментов. См. также дополнительную информацию. Нажмите здесь, чтобы увеличить рисунок . 
Рисунок 4. Цитотоксическую активность PJ-34 в PARP1 (- / -) мышиных эмбриональных фибробластов. А. (левый) процент многофокусной шпинделей рассчитывается в NНормальные (черная линия) и PArP1 - / - (серая линия) MEF, выдерживают в течение 48 часов с PJ-34 в указанных концентрациях. Процент многополярного шпинделей рассчитывалась из 20 шпинделей Всего обнаружено в 3 различных экспериментов. (Правый) Снижение выживаемости клеток обнаружены в культурах клеток инкубировали в течение 72 ч с PJ-34 (20 мкм) по отношению к выживаемости контрольных необработанных клеток (нормальные (черная линия) и PArP1 - / - (серая линия) MEF). Выживаемость клеток анализировали с помощью клеток АТФ (протокол 5). Средние значения 4 измерения для каждой клеточной линии в 3 различных экспериментов представлены B. шпиндели в случайно выбранных закреплен нормальных и PARP1 -. / - MEF в митозе, необработанных (контрольных) или инкубировали с PJ-34 в течение 48 ч при указанной концентрациях. PJ-34 вызвало многополярного шпинделей. Клетки фиксировали, проницаемыми и immunolabeled для α-и γ-тубулина (зеленая маркировка шпинделей и красная маркировка центросомам, соответственно). Хромосомы были помечены реагент DAPI (синий). . Представитель результаты из 3 различных экспериментах C. Мощные Номера для фенантрена PARP1 ингибиторы не влияет центросомам кластеризации в PARP1 (- / -) MEF. Шпиндели случайно выбранных нормальных и PARP1 - / - MEF представлены; MEF необработанных (контрольных) или MEF обрабатывали в течение 48 ч с не-фенантрен ингибиторы PARP, AG01469 (20 мкМ) или ABT888 (20 мкМ). Хромосомы помечены реагент DAPI (синий). Аналогичные результаты были получены в 3 различных экспериментов. Нажмите здесь, чтобы увеличить рисунок .
Нажмите здесь, чтобы увеличить рисунок . 
Нажмите здесь, чтобы увеличить рисунок .
Рисунок 2B. Справочная рис. Случайно выбранных MDA-MB-231 клетки в анафазе с би-координационного кластерных γ-тубулина помечены экстра-центросомам был отсканирован в течение 16 часов живой конфокальной микроскопии, 48 часов после того, как трансфецированы векторов, экспрессирующих γ-тубулина-GFP и H2b-RED (маркировка γ-тубулина центросомам очагов и в фиксированных клетках (зеленый) и гистонов H2b маркировки хромосом (красный), соответственно).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Живой конфокальной микроскопии при условии реального времени документацию цитотоксический эффект PJ-34 в живом мульти-центросомной клеток при митозе (фиг. 3 и дополнительную информацию). Это было первое живое документации приписывая цитотоксичность PJ-34 в человеческих раковых клеток к дополнительным центросомам де-кластеризации и гибели клеток, предполагая, индукция отмирания клетки катастрофу PJ-34 5-9. В противоположность этому, бифокальные кластеризации супер-нумерарий центросомам наблюдался в живых необработанных MDA-MB-231 клеток, подвергающихся нормальной митоза с би-focali кластерных дополнительные центросомам (рис. 2, дополнительная информация).
Согласно этим результатам, жить конфокальной микроскопии Трансфицированные клетки могут быть полезны для выявления транслокации белков, участвующих в дополнительное центросомам кластеризации в Трансфицированные клетки во время митоза 5,9,15,16. Идентификация белков, пострадавших от PJ-34 в нескольких CentrosOMAL клетки могут обеспечить некоторые подсказки для понимания смерти механизмы активируются дополнительные центросомам-де-кластеризации.
Преимущество живых конфокальной микроскопии в предоставлении информации в реальном времени во время митоза в живых клетках, обязан также ряд ограничений. Число клеток отсканированы в эксперименте ограничена. Поэтому возможности для обнаружения клеток в митоз низки, и несколько повторяющихся экспериментов, необходимых для реального времени документации митоза в сканированном клеток. Кроме того, успех в значительной степени зависит высокую эффективность трансфекции клеток векторами, экспрессирующими меченых белков. Таким образом, несмотря на то, надежная, живая конфокальной микроскопии занимает много времени и требует опытных работяг.
Для сравнения, иммуноцитохимия и конфокальной микроскопии фиксированные клетки включить рассмотрение большого количества клеток на эксперимент, который необходим для надежного статистического анализа. Этот метод был использован для сравненияэффекты PJ-34 в митозе нормальных доброкачественные клетки к его последствиям в раковых клетках митоз с внештатным центросомам 2,3. Кроме того, этот метод был использован для сравнения эффектов PJ-34, что и не-фенантрена PARP1 ингибиторов PARP (- / -) MEF укрывательство особо центросомам (клетки с высоким появление нескольких центросомам 11) (рис. 4).
Таким образом, наши результаты показывают преимущество объединения наиболее ценным в реальном времени информацию, предоставленную конфокальной микроскопии живых клеток в митозе (фиг. 2 и 3) с цитохимия и конфокальной анализа фиксированные клетки. Сочетание этих методов может быть полезным для определения цитотоксической активности малые молекулы, которые, как PJ-34, ориентированы на конкретные механизмы решающее значение для выживания клеток. Уникальная зависимость многих раковых клетках человека на дополнительной-центросомам биполярной кластеризации для их распространения и Survivдр. оказывает PJ-34 возможных кандидатов для терапии рака.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторы заявляют, что у них нет конкурирующих финансовых интересов.
Acknowledgments
Источники финансирования этого исследования: совместный фонд технологий Тель-Авивского университета трансфертной компании, RAMOT и Шеба-медицинский центр (М. СА и СИ.), ICRF - израильские железы исследовательский фонд (М. CA.) И Израиль научного фонда ( СИ).
Materials
| Name | Company | Catalog Number | Comments |
| REAGENTS | |||
| DMEM | Invitrogen (GIBCO) | 41965 | |
| FBS (Fetal bovine Serum) | Invitrogen (GIBCO) | 12657 | |
| Pen-Strep-Ampho solution | Biological Industries, Israel | 03-033-1B | |
| L-glutamine | Invitrogen (GIBCO) | 25030-024 | |
| 0.25% Tripsin-EDTA | Invitrogen (GIBCO) | 25200 | |
| 92 mm Petri dishes | Nunc,Thermo scientific | 150350 | |
| 35 mm poly-D-lysine coated glass bottom culture dishes | MatTek Corporation, USA | P35GC-0-14-C | |
| Luminescent ATP detection assay kit | Abcam | ab113849 | |
| NDS (Normal Donkey Serum) | Jackson ImmunoResearch | 017-000-121 | |
| Anti α-tubulin antibody | Sigma | T9026 | 1:250 dilution (IF) |
| Anti γ-tubulin antibody | Sigma | T5192 | 1:200 dilution (IF) |
| Alexa Fluor 488 Goat Anti-Mouse IgG | Invitrogen | A-11017 | 1:1,000 dilution (IF) |
| Alexa Fluor 568 Donkey Anti-Rabbit IgG | Invitrogen | A-10042 | 1:1,000 dilution (IF) |
| ProLong Gold antifade reagent with DAPI (mounting) | Invitrogen | P36935 | |
| JetPEI (liposomal transfection reagent ) | Polyplus | 101-10 | |
| EQUIPMENT | |||
| Confocal microscope | Leica (Mannheim, Germany) | TCS SP5II | |
References
- Jagtap, P., et al. Novel phananthridine inhibitors of poly(adenosine 5'-diphosphate-ribose) synthetase: Potent cytoprotective and antishock agents. Crit. Care Med. 30, 1071-1082 (2002).
- Castiel, A., et al. A small molecule exclusively eradicates human cancer cells: Extra-centrosomes de-clustering agent. BMC Cancer. 11 (1), 412 (2011).
- Inbar-Rozensal, D., et al. A selective eradication of human nonhereditary breast cancer cells by phenanthridine -derived polyADP-ribose polymerase inhibitors. Breast Cancer Res. 11 (6), R78 (2009).
- Gergely, F., Basto, R. Multiple centrosomes: together they stand, divided they fall. Genes Dev. 22, 2291-2296 (2008).
- Godinho, S. A., Kwon, M., Pellman, D. Centrosomes and cancer: how cancer cells divide with too many centrosomes. Canc. Met. Rev. 28, 85-98 (2009).
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2, 688-698 (2001).
- Walczak, C. E., Heald, R. Mechanisms of mitotic spindle assembly and function. International Rev. of Cytology. 265, 111-158 (2008).
- Ogden, A., Rida, P. C. G., Aneja, R. Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy. Cell Death Differ. 19, 1255-1267 (2012).
- Kramer, A., Anderhub, S., Maier, B. Mechanisms and Consequences of centrosomes clustering in cancer cells. The Centrosome: Cell and Molecular mechanisms of functions and disfunctions in disease. Schatten, E. , Humana Press Springer. 255-277 (2012).
- Galimberti, F., et al. Anaphase Catastrophe Is a Target for Cancer Therapy. Clin. Cancer Res. 17, 1218-1222 (2011).
- Kanai, M., et al. Haploinsufficiency of poly(ADP-ribose) polymerase-1-mediated poly(ADP-ribosyl)ation for centrosome duplication. Biochem. Biophys. Res. Commun. 359, 426-430 (2007).
- Gartner, E. M., Burger, A. M., Lorusso, P. M. Poly(ADP-ribose) polymerase inhibitors: a novel drug class with a promising future. Cancer J. 16, 83-90 (2010).
- Wahlberg, E., et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nature Biotechnology. 30, 283-288 (2012).
- Rouleau, M., Patel, A., Hendze, M. J., Kaufmann, S. H., Poirier, G. G. PARP inhibition: PARP1 and beyond. Nature Rev. Cancer. 10, 293-301 (2010).
- Leber, B., et al. Proteins Required for Centrosome Clustering in Cancer Cells. Sci. Transl. Med. 2 (33), 33-38 (2010).
- Kwon, M., et al. Mechanisms to suppress multipolar division in cancer cells with extra centrosomes. Gene Dev. 22, 2189-2203 (2008).

{kind=link}
{kind=link}