

Summary
Vi præsenterer en ex vivo celle migration assay, der tillader præcis kvantificering af entero neurale potentiale våbenskjold celle migration i tilstedeværelsen af forskellige vækstfaktorer.
Abstract
Neuralkam celler (NCC) er en forbigående og multipotent celle befolkning, der stammer fra den dorsale neuralrøret og migrerer udførligt i hele udviklingen af hvirveldyr embryo. Ud over at give perifere glia og neuroner, NCC generere melanocytter samt det meste af skelettet kranio-ansigtsbehandling. NCC migration og differentiering styres af en kombination af deres aksiale oprindelse langs neuralrøret og deres eksponering for regionalt forskellige ekstracellulære signaler. Sådanne bidrag af ekstracellulære ligander er især tydeligt under dannelsen af det enteriske nervesystem (ENS), et kompleks indbyrdes forbundet netværk af neurale ganglier at lokalt styrer (blandt andet) gut muskelbevægelser og tarm motilitet. De fleste af ENS er afledt af en lille indledende pulje af NCC at foretage en lang rejse for at kolonisere - i en rostralt caudal fashion - hele længden af den potentielle tarmen. Blandt flere signalveje, der vides atpåvirke enteriske NCC kolonisering, er GDNF / RET signalering anerkendt som den vigtigste. Faktisk spatiotemporally styret sekretion af RET ligand GDNF af tarmen mesenkymet er hovedansvarlig for tiltrækning og vejledning af RET-udtrykker enteriske NCC til og inden for embryonale tarmen. Her beskriver vi en ex vivo-celle migration assay, der gør brug af en transgen mus linje besidder fluorescensmærket NCC, som muliggør en præcis kvantificering af enterisk NCC migration potentiale i nærvær af forskellige vækstfaktorer, herunder GDNF.
Introduction
Neuralkam celler (NCC) er en forbigående celletype unikke for hvirveldyr, der danner mange derivater under fosterudviklingen. Denne celle population opstår på grænsen til den neurale plade, der støder op til ikke-neurale ectoderm 1. Under neurulation, bøjning af de neurale plade steder NCC langs ryggens kant danner neuralrøret. NCC derefter gennemgå en epitelial-mesenkymal overgang, adskillelse og migrere væk fra neuralrøret. NCC kolonisere forskellige embryoniske strukturer, herunder fordøjelseskanalen, hvor de danner hele enteriske nervesystem (ENS), et sammenhængende netværk af neurale ganglier indlejret i tarmvæggen. Som det for nylig revideret 2,3, har mange gener er involveret i udviklingen af denne indviklede struktur.
De fleste af ENS er afledt fra en lille pulje af NCC oprindelse fra vagus neuralrøret (dvs. omkring den potentielle baghjerne / rygmarv afgrænsning) 4.Disse neurale progenitorer nå foregut omkring embryonisk dag (e) 9.0 i mus og derefter migrere kaudalt i tarmen mesenkymet til ca e15.0 at kolonisere hele embryonale tarmene. En mindre delmængde af colon neurale stamfædre er også leveret af sakral NCC, der invadere den bageste tarmen i den modsatte retning op til coecum 4.. Både vagal og sakrale NCC kræver flere migration-, proliferation-, overlevelse-og differentiering-fremmende signaler for at sikre fuldstændig dannelse af ENS. I denne henseende dyremodeller - især genetisk modificerede mus - har været medvirkende til at identificere flere væsentlige ekstracellulære ligander: GDNF (gliacelle neurotrofisk faktor), Endothelinreceptor-3, neurotrofin-3, BMP (knoglemorfogene proteiner), Netrin , såvel som Sonic og indiske Pindsvin (Shh og Ihh) 5-10. Af disse er GDNF signalering gennem tyrosinkinase transmembranreceptor RET (rearrangerede under transfektion) anerkendes som the mest kritiske vej for tiltrækning og vejledning af NCC til og inden for embryonale tarmen. GDNF udskilles af tarmen mesenkymet og danner en spatiotemporally kontrolleret rosrrocaudal forløb, der er direkte chemoattractive til enteriske NCC, som udtrykker RET 11,12.
Blandt andre funktioner, ENS regulerer bevægelsen i fordøjelseskanalen gennem dens samspil med den glatte muskulatur i tarmvæggen. Fravær af neurale ganglier i terminale region af tarmen resulterer i Hirschsprungs sygdom: tonic sammentrækning af det pågældende segment fører til blokering opstrøms akkumulering af nedbrudt materiale og massiv udspilning af tarmen og maven. Hirschsprungs sygdom opstår cirka en ud af 5.000 levendefødte. Den Rostro-caudale migration mønster af enteriske NCC menes at være den vigtigste faktor i ætiologien af Hirschsprungs sygdom. Tyktarmen længst væk fra kilden til at migrere NCC og sidste portion bOwel at blive koloniseret, er mest modtagelige for defekter i ENS formation. I overensstemmelse med sin afgørende rolle i enterisk NCC migration, afbrydelse af GDNF / RET signalering er den vigtigste kendte genetiske årsag til Hirschsprungs sygdom 13..
For bedre at studere NCC og ENS udvikling, vi genererede en transgen mus linje - opkaldt Gata4p [5 kb]-GFP 14 - hvor vandrende NCC er mærket med grønt fluorescerende protein (GFP). Vi næste perfektioneret en ex vivo celle migration assay, tilpasset fra offentliggjorte arbejde med andre grupper 11,12,15, der nu tillader præcis kvantificering af entero NCC migration potentiale i tilstedeværelsen af forskellige vækstfaktorer, såsom GDNF.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Etik erklæring
Forsøg med mus blev udført efter canadisk Råd Animal Care retningslinjer for pleje og manipulation af dyr, der anvendes i medicinsk forskning. Protokoller, der involverer manipulation af dyr blev godkendt af den institutionelle etiske komité fra University of Quebec i Montreal (Comité Institutionnel de Protection des Animaux, referencenummer 0512-R3-650 til 0513).
1.. Fremstilling af kollagengeler
Arbejde i en steril måde, under en vævskultur hætte.
- Forbered komplet 5x DMEM (Eagle modificerede essentielle medium) herunder standard antibiotika. 3,37 g DMEM pulver og 0,925 g NaHCO3 i 20 ml vand opløses. Steriliseres ved passage gennem et 0,22 um filter. Tilsæt 2,5 ml sterilt 100x penicillin / streptomycin og 25 ml sterilt varmeinaktiveret føtalt bovint serum. Opbevares ved 4 ° C.
- På is, bland 800 piaf kollagen I-opløsning (3,77 mg / ml i 0,02 N eddikesyre, filtersteriliseret), 600 pi af komplet 5x DMEM og 17 pi af 1 N NaOH. Fortyndes med sterilt vand til et samlet volumen på 3 ml. Medtag relevante vækstfaktorer i blandingen. Vi bruger typisk GDNF ved 10 ng / ml for at stimulere enterisk NCC migration.
- Deposit omkring 480 ul i hver brønd i en enkelt række i en 24-brønds plade. Undgå bobler. De resterende rækker kan anvendes til at teste virkningen af andre vækstfaktorer på cellemigrering.
- Lad kollagen polymerisere mindst 1 time i en steril inkubator ved 37 ° C.
2. Dissektion af dyr 16
- Opsæt parringer og tjekke for vaginale propper næste morgen. Dag e0.5 være middag på dagen vaginal prop er fundet, isolere kvindelige og vente 12 dage indtil E12.5.
- Bedøver gravid mus med isofluran og aflive af CO 2 indånding.
- Spray mus med 70% ethanol. Løft maven spårørende og åbne maven hulrum med dissekere saks.
- Fjerne livmoderen i et glas petriskål fyldt med iskoldt PBS (phosphatbufret saltvand). Skær livmoderen på tværs mellem de enkelte deciduum hævelser at isolere de enkelte embryoimplantationen site.
- Arbejdet med hver implantationssted separat i en anden glas petriskål fyldt med iskoldt PBS. Under et dissektionsmikroskop bruge fin pincet til at fjerne muskel lag af livmoderen.
- Åbn visceral blommesækken og fosterhinden at afsløre embryoet. Vær forsigtig, når overskæring blodkarrene sammenføjning fosteret til moderkagen / visceral blommesæk, da de er forbundet med udviklingslandene tarmene.
- Sever fosterets hoved i nakken.
- Indsæt et lukket pincet i bughulen af et foster, lige over den mørke røde farve lever og lad tangen åbne sig selv (stop anvende pres) for at gøre en tværgående åbning i maven hulrum. Træk de åbnede pincet ned modden bageste ende af embryonet at åbne maven helt.
- Grib bindevævet bag leveren og trække indvolde ud af maven, være omhyggelig med ikke at bryde tarmene (tyktarmen er knyttet til anus).
- Skær kolon at befri indvolde fra resten af embryoet. Snittet kan ske på ethvert sted langs tyktarmen. Reserve halepartiet til senere.
- Lirke bindevæv fra coecum, derefter resten af tarmene. Vær omhyggelig med ikke at såre tarmene mens du gør det.
- Skær leveren og maven (på rostralt ende af tyndtarmen), samt mesonephros og genitale kamme, hvis nogle er til stede.
- Isoler tyndtarmen. Igen, vær forsigtig med ikke at kvæle tarmen. Fra nu af kan rosrrocaudal orientering af tarmen væv kan spores ved hjælp af den skarpe krumning stede på rostral ende. Lad tyndtarmen i PBS ved stuetemperatur i så kort tid som muligt, før emsengetøj (se trin 3.3).
- Endelig registrere antallet af hale somitter for hvert foster til præcist at bestemme den fase af fosterudviklingen.
3. Sektionsinddeling af Embryonale Tarmene
- Før du fortsætter med embryo dissektioner, smelte 1,5 g agarose i 100 ml PBS (phosphatpufret saltvand) og holde ved 50 ° C.
- Hæld smeltet agarose i en indlejring støbeform (f.eks lukket 2 ml mikrocentrifugerør, der er blevet skåret i længderetningen at udskære omkring 1/4 af rørets væg). Lad agarose afkøle til ca 42-45 ° C, bør det kun være lidt varm at røre ved.
- Integrer embryonale tarmen lige før agarose størkner (dette kan vurderes med en pincet tips og vil opstå omkring 36-38 ° C). Holding tarmen med en pincet med den foldede rostralt ende trække det meget langsomt gennem agarose langs længden af formen. Dette hjælper til at holde tarmen lige mens agarose sæt. Slip tudstede så snart det begynder at modstå at blive flyttet. Hold styr på rostral-caudale orientering af tarmen.
- Sæt formen i køleskab 2-3 minutter for at sikre, at agarosen er fuldstændigt indstillet.
- Tag agarose fra formen (ved at skubbe det ud af den åbnede Eppendorf-rør). Med en kniv, tage den overskydende agarose i begge ender, hvilket gør nedskæringer vinkelret på tarmen.
- Lim rostralt endetarmen / agarose blok ned på metallet fase af en mikrotom med vibrerende kniv. Trim overskydende agarose på siderne af tarmen / agarose blok.
- Monter metal scenen på den vibrerende mikrotomet kammeret. Hvis det er nødvendigt, justere vinklen på scenen, så tarmen er så lodret som muligt (og dermed vinkelret på bladet). Dæk prøven med iskold PBS. Bring mikrotom kniv ned til nogle få millimeter under buffer overflade.
- Lav 200 um vibratome tværgående udskæringer af den bageste-mest tyndtarmen, der sikrer, at eACH agarose skive indeholder en fuld tarm afsnit.
4.. Kultur Intestinal Eksplantater
- Forsigtigt deponere nyslået tarmen / agarose skiver fladt på de kollagengeler med pincet, placere en skive mod midten af hver brønd.
- Inkuberes 3 dage ved 37 ° C i en fugtig 5% CO2 atmosfære for at muliggøre migration af NCC ud af eksplantatet.
- Tag tarmen / agarose skiver fra kollagengelen meget forsigtigt med en pincet. Pas på ikke at beskadige gelen nedenfor.
- Undgå at berøre kollagengelen direkte i de efterfølgende inkubations-og vasketrin, for at sikre, at cellemigrering mønster ikke forstyrres. Fix med 500 pi 4% PFA (paraformaldehyd) (i PBS) per brønd i 1 time ved stuetemperatur.
- Udskift fiksativ med 500 pi DAPI (4 ',6-diamidino-2-phenylindol)-opløsning (5 ug / ml i PBS) per brønd og inkuberes 10 minutter ved stuetemperatur.
- Vask hver brønd 3xmed 500 pi PBS i 5 min.
- Fotografere de fluorescerende celler (GFP og DAPI kanaler) indlejret i kollagengelen indenfor hver brønd.
5.. Image Analysis
Vi gjorde udstrakt brug af ImageJ 17 at behandle og kvantificere de billeder genereret efter eksplantation kultur.
- Start med billeddannelse en mikrometer dias med samme forstørrelse som de fluorescerende celle fotografier. Måle længden af en mikron i pixels (ved hjælp af den rette linje værktøj).
- Indstil skalaen (Analyze / Set Scale; input antal pixels / micron).
- For hver GFP-fluorescens fotografi, ændre billedet format til 8-bit gråtoner (Image/Type/8-bit).
- Juster intensiteten af signalet (Image / Juster / Lysstyrke / Kontrast).
- Fratræk baggrundsstøjen om nødvendigt (Process / Træk Baggrund, justere rullende kugle radius).
- Sæt en grænse for at fremhæve celler og celleklumper (Billede / Juster / Threshold), denn anvende et vandskel at opdele klumper (Process / Binary / Watershed).
- Analyser partikler til at angive områder af interesse (ROI) og generere celle nummer statistik (Analyze / Analyser Partikler, minimum størrelse at udelukke de resterende pixel).
- Måleapparaturet muligheder for at medtage Féret diameter (analyse / sæt målinger).
- Gruppe ROIs og måle som helhed til at bestemme Féret diameter, en angivelse af celle-spredning (Analyze / Værktøj / ROI manager / Mere / ELLER, så ROI manager / Tilføj, og ROI manager / Measure).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
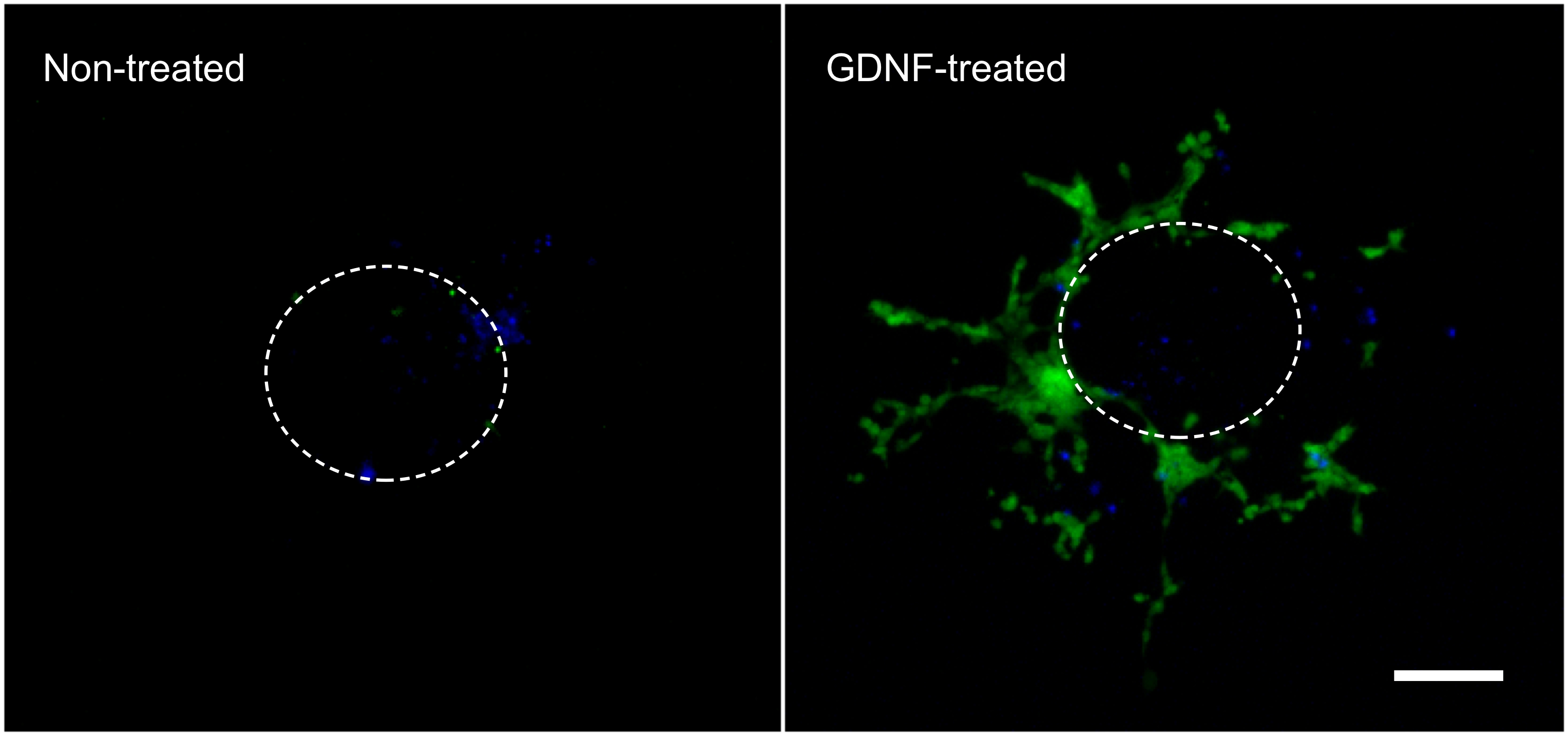
De følgende resultater er repræsentative for hvad der kan opnås med den teknik, der er beskrevet her (figur 1). Anvendelsen af vækstfaktorer (dvs. GDNF) stimulerer migreringen af GFP-udtrykkende enterisk NCC ud af tarmens eksplantatet og ind i kollagengelen (figur 2). Selvom nogle celler kommer ud af eksplantatet i fravær af vækstfaktorer, er disse for det meste ikke GFP-mærkede og repræsenterer passiv indgang. Det er nødvendigt at fjerne den intestinale skive fra kollagengelen for at optage resultater, da dette væv stadig er stærkt befolket af fluorescerende celler og ellers ville skjule celler liggende i collagen nedenunder. Kvantificering af disse resultater viser, at mange flere celler findes inden kollagengelen når GDNF er til stede, og at aktiv vandring finder sted (figur 3). Faktisk er passive celler findes umiddelbart under eksplantaterne inden diameteren af en intestinal skive Wher EAS celler, der aktivt invaderer kollagengelen bevæge sig væk fra deres udgangspunkt og sprede sig yderligere.

Figur 1. Oversigt over eksplantatet dyrkningsteknik. A) Ensartet population af fluorescerende enterisk NCC i caudale tyndtarmen region anvendes til at fremstille 200 μ tykke eksplantater. Målestok:. 200 um B) Dyrkningsmediet indeholdende kollagen er deponeret i plader med 24 brønde og efterladt til at hærde i 1 time. Vibratome skiver af agarose-indlejret embryonale tarme aflejres på gelerne (en skive per brønd), og fluorescerende enteriske NCC får lov til at migrere ud af eksplantaterne i 3 dage. Skiverne bliver derefter taget ud, før billeddannelse de celler, der invaderede de kollagengeler.s/ftp_upload/50709/50709fig1highres.jpg "target =" _blank "> Klik her for at se større billede.

Figur 2. Cellemigrering af tarmens eksplantatet og inden kollagengelen. Celler, migrerede ud af en intestinal eksplantat i løbet af 3 dages inkubering i fravær eller nærvær af 10 ng / ml GDNF blev fikseret, farvet med DAPI (blå) og fotograferet til viser GFP-mærkede enteriske NCC (grøn). 70X forstørrelse. Skala bar: 100 um. Den stiplede linje repræsenterer den omtrentlige størrelse og placering af eksplantatet, før det blev taget af gelen. Klik her for at se større figur. Klik på here for at se større billede.

Figur 3. Kvantificering af enteriske NCC migration potentiale. Antallet og spredning (Féret diameter) af GFP-udtrykkende celler migrerer ud af intestinale explanter efter 3 dage blev kvantificeret ved hjælp ImageJ software 17. I begge tilfælde er der en betydelig forskel mellem ubehandlede og GDNF-behandlede betingelser ifølge en Students t-test (p <0,001, *). Den gennemsnitlige diameter af intestinale skiver (punkteret Sort Serie: 260 mM) blev inkluderet for at skelne mellem passiv indrejse og aktiv migration. NT: ikke-behandlede, n:. Antal eksplantater forarbejdede Klik her for at se større billede .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Vi viser, hvordan vores ex vivo eksplantation kultur teknik kan bruges til præcist at kvantificere enterisk NCC migration potentiale i nærværelse af GDNF. En sådan præcis kvantificering er blevet betydeligt nemmere ved hjælp af 200 um tykke vibratome gut sektioner i stedet for store stykker af omtrentlige størrelse, som tidligere beskrevet 11,12,15. Faktisk er det muligt for os at arbejde med et rimeligt antal af celler i en meget reproducerbar indstilling. Det skal bemærkes, at en ensartet fordeling af fluorescerende enterisk NCC i caudale længst region af tyndtarmen, hvorfra eksplantation skiver skæres tillader også analyse af flere sektioner fra en enkelt tarm (figur 1A). I betragtning af, at både enteriske NCC og axoner kan afslutte vævet i sådanne analyser 11, tilbagetrækning af eksplantaterne ved udgangen af kultur periode lader os fokusere udelukkende på vandrende NCC.
De fleste kritiske trin er skitseret i protokollen tekst, men da welfare af tarmens eksplantat er altafgørende for at opnå en sund migrerer NCC, bør særlig forsigtighed skal træffes. Undgå at udsætte embryonale væv for pludselige temperaturændringer, især når tarmen er indlejret i agarose (trin 3.3). Sørg for, at tarmen er ved stuetemperatur og agarose så køligt som muligt (stadig smeltet) for at undgå "madlavning" vævet. Den intestinale explant skal trives på dyrkningsmediet fyldt kollagen gel, ofte vokser i størrelse og breder ud Agarosens skive. Hvis eksplantatet synes usunde eller værre, har tendens til at omkomme under inkubation, prøv at erstatte PBS med dyrkningsmedier ved stuetemperatur (f.eks HEPES-buffered M2 eller DMEM suppleret med 10% FBS) at hjælpe med at opretholde det under dissektion.
En væsentlig begrænsning for vores tilgang er, at det beror på tilgængeligheden af en mus linje opnår en fluorescerende mærke til at migrere NCC. I mangel af en sådan ressource, et antistof mod migrating NCC (f.eks anti-Ret eller anti-Sox10) kan anvendes til at mærke celler, der invaderede kollagengelen. I betragtning af, at tarmen mikromiljø er langt mere kompleks end et simpelt kollagengel resultater opnået med dette in vitro assay måske ikke fuldstændigt afspejle adfærd enteriske NCC in vivo. Yderligere forsøg med live-cell imaging anbefales at vurdere denne adfærd. Det er også bemærkelsesværdigt, at ud over sin rolle som en chemoattractant er GDNF kendt for at fremme udbredelsen af migrerende enteriske NCC 4.. Vores mål for enterisk NCC migration potentiale i nærværelse af GDNF er derfor sandsynligvis en blanding af sand cellemigration og celleproliferation, beslægtet med in vivo-mekanismer, der fører til NCC kolonisering af tarmene. Hvis der ønskes en klar sondring mellem disse to processer, kan tilføjelsen af en cellecyklus blokker (f.eks AZD 5438 18) i kulturen medier begrænse analysen til celle migration.
Denne teknik kan udvides til at afprøve forskellige andre ekstracellulære ligander samt hæmmere af specifikke signalveje, og enhver kombination deri. Andre væv kan også potentielt dissekeret og snit tillader undersøgelse af NCC migration i mange embryoniske strukturer. Kombineret med nye og / eller ukarakteriserede mutant musestammer med mulige defekter i NCC udvikling kan vores teknik anvendes til hurtigt screene for mangler i migration adfærd som svar på specifikke signalering begivenheder.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ikke noget at afsløre.
Acknowledgments
Vi takker Denis Flipo til billedbehandling og analyse rådgivning, og David W. Silversides i hvis laboratorium Gata4p [5 kb]-GFP mus linje blev genereret. Forskning i Pilon laboratorium er finansieret af CIHR, NSERC, FRQS og FRQNT.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J. The enteric nervous system. Dev. Biol. 366 (1), 64-73 (2012).
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}