

Summary
We presenteren een ex vivo celmigratie analyse die exacte kwantificering van enterische neurale celmigratie potentieel in aanwezigheid van verschillende groeifactoren maakt.
Abstract
Neurale cellen (NCC) is een voorbijgaande en multipotent celpopulatie die afkomstig is van de dorsale neurale buis en migreert intensief rond de ontwikkeling van gewervelde embryo. Naast het leveren van perifere glia en neuronen, NCC genereren melanocyten evenals de meeste van de craniofaciale skelet. NCC migratie en differentiatie wordt door een combinatie van de axiale oorsprong langs de neurale buis en hun blootstelling aan regionaal verschillende extracellulaire signalen. Een dergelijke bijdrage van extracellulaire liganden is vooral duidelijk tijdens de vorming van het enterische zenuwstelsel (ENS), een complex netwerk van onderling verbonden neurale ganglia die lokaal regelt (onder andere) gut spierbewegingen en darmmotiliteit. Het merendeel van de ENS is afgeleid van een kleine initiële pool van NCC die een lange reis ondernemen om te koloniseren - in een rostrale om caudale mode - de gehele lengte van de aspirant-darm. Onder verschillende signaalwegen bekend bijbeïnvloeden enterische NCC kolonisatie, wordt GDNF / RET signalering erkend als de belangrijkste. Inderdaad, spatiotemporeel gecontroleerde afscheiding van de RET ligand GDNF door de darm mesenchym is voornamelijk verantwoordelijk voor het aantrekken en begeleiden van-RET uitdrukken enterische NCC naar en binnen de embryonale darm. Hier beschrijven we een ex vivo celmigratie assay gebruikmakend van een transgene muis lijn bezit fluorescerend gemerkte NCC, die nauwkeurige kwantificering van enterische NCC migratiepotentieel in aanwezigheid van verschillende groeifactoren, waaronder GDNF maakt.
Introduction
Neurale cellen (NCC) is een voorbijgaande celtype uniek is voor gewervelde dieren dat veel derivaten maakt tijdens de ontwikkeling van het embryo. Deze celpopulatie ontstaat op de grens van de neurale plaat, naast niet-neurale ectoderm 1. Tijdens neurulatie, buigen van de neurale plaat plaatsen NCC langs de dorsale rand van de vorm neurale buis. NCC ondergaan dan een epitheliale-mesenchymale transitie, scheiden en migreren weg van de neurale buis. NCC koloniseren verschillende embryonale structuren, waaronder het spijsverteringskanaal waar zij vormen de gehele enterische zenuwstelsel (ENS), een onderling verbonden netwerk van neurale ganglia ingebed in de darmwand. Zoals onlangs beoordeeld 2,3, hebben vele genen betrokken bij de ontwikkeling van deze ingewikkelde structuur.
De meeste ENS is afgeleid van een kleine pool van NCC afkomstig van de vagale neurale buis (dwz in de toekomstige achterhersenen / ruggenmerg grens) 4.Deze neurale voorlopercellen bereikt de voordarm rond embryonale dag (e) 9.0 in muizen en vervolgens migreren caudaal in de darm mesenchym tot ongeveer e15.0 om de hele embryonale darmen koloniseren. Een kleine subset van colon neurale voorlopercellen verschaft ook sacrale NCC, die de achterste darm tot de blindedarm 4 binnendringen in de tegenovergestelde richting. Zowel de vagale en sacrale NCC vereisen meerdere migratie-, proliferatie-, survival-en-differentiatie bevorderen signalen om volledige vorming van de ENS zorgen. In dit verband diermodellen - vooral genetisch gemodificeerde muizen - zijn geweest bij de identificatie van verschillende essentiële extracellulaire liganden opgeleverd GDNF (glial cel-afgeleide neurotrofe factor), endotheline-3, neurotrofine-3, BMP (bot morfogenetische eiwitten), Netrin , evenals Sonic en Indiase Hedgehog (Shh en Ihh) 5-10. Van deze, GDNF signalering door de tyrosine kinase transmembraanreceptor RET (Rearranged tijdens transfectie) wordt erkend als ee meest kritieke pad voor het aantrekken en begeleiden van NCC naar en binnen de embryonale darm. GDNF wordt afgescheiden door de darm mesenchym en vormt een spatiotemporeel gecontroleerde rosrrocaudal gradiënt die direct chemoattractive enterische NCC, die RET 11,12 expressie.
Onder andere functies, de ENS regelt verkeer binnen het spijsverteringskanaal door de interactie met gladde spier in de darmwand. Afwezigheid van neurale ganglia in het terminale gebied van de darm leidt tot ziekte van Hirschsprung: tonische contractie van het aangetaste segment leidt tot verstopping stroomopwaarts accumulatie van verteerd materiaal en massale uitzetting van de darm en buik. Ziekte van Hirschsprung komt ongeveer een op 5000 levende geboorten. De rostro-caudale migratiepatroon van enterische NCC wordt beschouwd als de belangrijkste factor die bijdraagt aan de etiologie van de ziekte van Hirschsprung zijn. De dikke darm, het verst van de bron van migrerende NCC en laatste portie bOwel te worden gekoloniseerd, is het meest gevoelig voor defecten in ENS formatie. In overeenstemming met haar cruciale rol in enterische NCC migratie, verstoring van GDNF / RET signalering is de belangrijkste bekende genetische oorzaak van de ziekte van Hirschsprung 13.
Om beter te bestuderen NCC en ENS ontwikkeling genereerden wij een transgene muis lijn - vernoemd Gata4p [5kb]-GFP 14 - waarin trekkende NCC zijn gelabeld met Green Fluorescent Protein (GFP). We vervolgens geperfectioneerd een ex vivo cel migratie assay, aangepast van gepubliceerd werk van andere groepen 11,12,15, die nu zorgt voor een precieze kwantificering van enterische NCC migratie potentieel in de aanwezigheid van verschillende groeifactoren, zoals GDNF.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Ethiek verklaring
Experimenten met muizen werden uitgevoerd na Canadese Raad van Animal Care richtlijnen voor de zorg en de manipulatie van dieren die in het medisch onderzoek. Protocollen met betrekking tot de manipulatie van dieren werden goedgekeurd door de institutionele ethische commissie van de Universiteit van Quebec in Montreal (Comite Institutionnel de Protection des Animaux; referentienummer 0512-R3-650-0513).
1. Bereiding van collageen Gels
Werken in een steriele manier, onder een weefselkweek kap.
- Bereid compleet 5x DMEM (Eagle's gemodificeerd essentieel medium) inclusief standaard antibiotica. Los op 3,37 g DMEM poeder en 0,925 g NaHCO3 in 20 ml water. Steriliseren die door een 0,22 urn filter. Voeg 2,5 ml steriele 100x penicilline / streptomycine en 25 ml steriel warmte geïnactiveerd foetaal runderserum. Bewaren bij 4 ° C.
- Op ijs, meng 800 ulCollageen I-oplossing (3,77 mg / ml in 0,02 N azijnzuur, filter-gesteriliseerd), 600 gl 5x volledige DMEM en 17 ui 1 N NaOH. Verdun met steriel water tot een eindvolume van 3 ml. Neem relevante groeifactoren in de mix. Wij gebruiken meestal GDNF bij 10 ng / ml tot enterische NCC migratie stimuleren.
- Borg ongeveer 480 pl in elke well van een enkele rij in een 24-well plaat. Vermijd bubbels. De resterende rijen kan worden gebruikt om het effect van andere groeifactoren op celmigratie testen.
- Laat het collageen polymeriseren ten minste 1 uur in een steriele incubator bij 37 ° C.
2. Dissectie van dieren 16
- Opgezet paringen en controleer vaginale stekkers de volgende ochtend. Dag E0.5 zijnde uur op de dag van de vaginale plug wordt gevonden, te isoleren van de vrouwelijke en wacht 12 dagen tot E12.5.
- Verdoven zwangere muizen met isofluraan en euthanaseren door CO 2 inhalatie.
- Spuit de muis met 70% ethanol. Til de buik skin en open de buikholte met dissectieschaar.
- Verwijder de baarmoeder in een glazen petrischaal gevuld met ijskoude PBS (fosfaatgebufferde zoutoplossing). Snijd de baarmoeder dwarsrichting tussen afzonderlijke deciduum zwellingen elk embryo implantatie isoleren.
- Werken op elke implantatie apart in een ander glas petrischaal gevuld met ijskoude PBS. Onder een microscoop ontleden, gebruik fijne tang om de spierlaag van de baarmoeder te verwijderen.
- Open de viscerale dooierzak en amnion het embryo te onthullen. Wees voorzichtig bij het doorsnijden van de bloedvaten toetreding tot de embryo naar de placenta / viscerale dooierzak, als ze worden verweven met de ontwikkeling van darmen.
- Sever hoofd van het embryo in de nek.
- Plaats een gesloten forceps in de buikholte van een embryo, net boven de donkerrood gekleurde lever en laat de tang geopend zichzelf (langer dienen druk) een transversale opening in de buikholte maken. Trek de geopende tang naar beneden richtinghet achterste uiteinde van het embryo aan de buik volledig openen.
- Pak het bindweefsel achter de lever en trek de ingewanden van de buik, zorg dat u de darmen breken (de dikke darm wordt aan de anus verbonden).
- Snijd de dikke darm om de ingewanden te bevrijden van de rest van het embryo. De snede kan worden gemaakt op een locatie langs de colon. Reserveer de staart gedeelte voor later.
- Plagen het bindweefsel van de blindedarm, dan de rest van de darmen. Wees voorzichtig de darmen niet te verwonden tijdens het doen.
- Knip de lever en maag (de rostrale einde van de dunne darm), alsmede de mesonephros en genitale randen als sommige aanwezig zijn.
- Isoleer de dunne darm. Nogmaals, wees voorzichtig de darm niet te smoren. Vanaf nu kan de rosrrocaudal oriëntatie van het darmweefsel worden gevolgd met behulp van de scherpe kromming aanwezig is op de rostrale einde. Laat de dunne darm in PBS bij kamertemperatuur gedurende zo kort mogelijk voor embeddengoed (zie stap 3.3).
- Tenslotte registreren het aantal staart somieten voor elk embryo nauwkeurig te bepalen het stadium van de embryonale ontwikkeling.
3. Opdeling van Embryonale Darmen
- Alvorens embryo dissecties Smelt 1,5 g agarose in 100 ml PBS (fosfaat gebufferde zoutoplossing) en houdt bij 50 ° C.
- Giet gesmolten agarose in een inbedding vorm (bijv. een gesloten 2 ml microcentrifugebuis die is lengte gesneden tot ongeveer 1/4 van de buiswand te snijden). Laat de agarose afkoelen tot ongeveer 42-45 ° C, mag het alleen een beetje warm om aan te raken.
- De embryonale darm insluiten vlak voor de agarose stolt (de tijd kan geëvalueerd worden met een pincet tips verschijnt rond optreden 36-38 ° C). Houd de darm met een tang door het gevouwen rostrale einde trek langzaam via agarose langs de lengte van de matrijs. Dit helpt om de darm recht te houden terwijl de agarose sets. Laat de tZodra hier het begint te weerstaan verplaatst. Blijf op de hoogte van de rostrale-caudale oriëntatie van de darm.
- Plaats de vorm in de koelkast 2-3 minuten, zorgen voor de agarose volledig ingesteld.
- Haal de agarose uit de mal (door uit de geopende Eppendorf buis te schuiven). Met een mes, verwijder de overtollige agarose aan beide uiteinden, waardoor bezuinigingen loodrecht op de darm.
- Lijm de rostrale einde van de darm / agarose blokkeren de metalen fase van een microtoom met trillend mesje. Knip de overtollige agarose aan de zijkanten van de darm / agarose blok.
- Monteer de metalen trap op de trillende microtoom kamer. Indien nodig, stel de hoek van het podium, zodat de darm is zo verticaal mogelijk (dus loodrecht op het blad). Bedek het monster met ijskoude PBS. Breng de microtoom mes tot een paar millimeter onder de buffer oppervlak.
- Maak 200 pm vibratome dwarssneden van de staartvin meest dunne darm, zodat eagarose slice ach bevat een volledige intestinale sectie.
4. Culture of intestinale explantaten
- Zachtjes deponeren de vers gesneden darm / agarose plakjes plat op de collageen gels met een pincet, het plaatsen van een snee in het midden van elk putje.
- Incubeer 3 dagen bij 37 ° C in een vochtige 5% CO2 atmosfeer, migratie van NCC mogelijk uit het explantaat.
- Neem de darm / agarose plakjes uit de collageen-gel heel voorzichtig met een pincet. Zorg ervoor dat u de gel hieronder beschadigen.
- Vermijd de collageengel aan te raken tijdens het verdere incubatie en wasstappen om ervoor te zorgen dat de celmigratie patroon niet wordt verstoord. Bevestig met 500 ui 4% PFA (paraformaldehyde) (in PBS) per putje 1 uur bij kamertemperatuur geroerd.
- Vervang het fixeermiddel met 500 ui DAPI (4 ',6-diamidino-2-fenylindool) oplossing (5 ug / ml in PBS) per putje en incubeer 10 minuten bij kamertemperatuur.
- Was elk putje 3xmet 500 ui PBS gedurende 5 minuten.
- Fotografeer de fluorescerende cellen (GFP en DAPI kanalen) ingebed in de collageen-gel in elk putje.
5. Beeldanalyse
We maakten uitgebreid gebruik van ImageJ 17 om de beelden gegenereerd nadat explantaatkweek verwerken en te kwantificeren.
- Begin met het afbeelden van een micrometer op dezelfde vergroting als de fluorescerende cel foto's. Meet de lengte van een micron in pixels (volgens de lineaire tool).
- Zet de schaal (Analyseren / Set Schaal; ingang aantal pixels / micron).
- Voor elk GFP-fluorescentie foto, wijzigt het beeld formaat naar 8-bits grijsschaal (Image/Type/8-bit).
- Pas de intensiteit van het signaal (Afbeelding / Aanpassen / Helderheid / Contrast).
- Trek het achtergrondgeluid indien nodig (Proces / Aftrekken Achtergrond; pas de rollende bal radius).
- Stel een drempel naar de cellen en celklonten (Afbeelding / Aanpassen / Threshold), markeert den toepassing een keerpunt voor de klonten (Proces / Binary / Watershed) verdelen.
- Analyseer deeltjes regio van belang (ROI) op te geven en het genereren van het aantal cellen statistieken (Analyze / Analyseer Deeltjes; vastgestelde grootte minimum om de resterende pixels uit te sluiten).
- Stel meting opties om Feret de diameter (Analyseren / Set-metingen) omvatten.
- Groep ROI en meet als geheel om de diameter van de Feret bepalen, een indicatie van de cel spread (Analyseren / Tools / ROI Manager / Meer / OR, dan ROI Manager / toevoegen, en ROI Manager / Measure).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
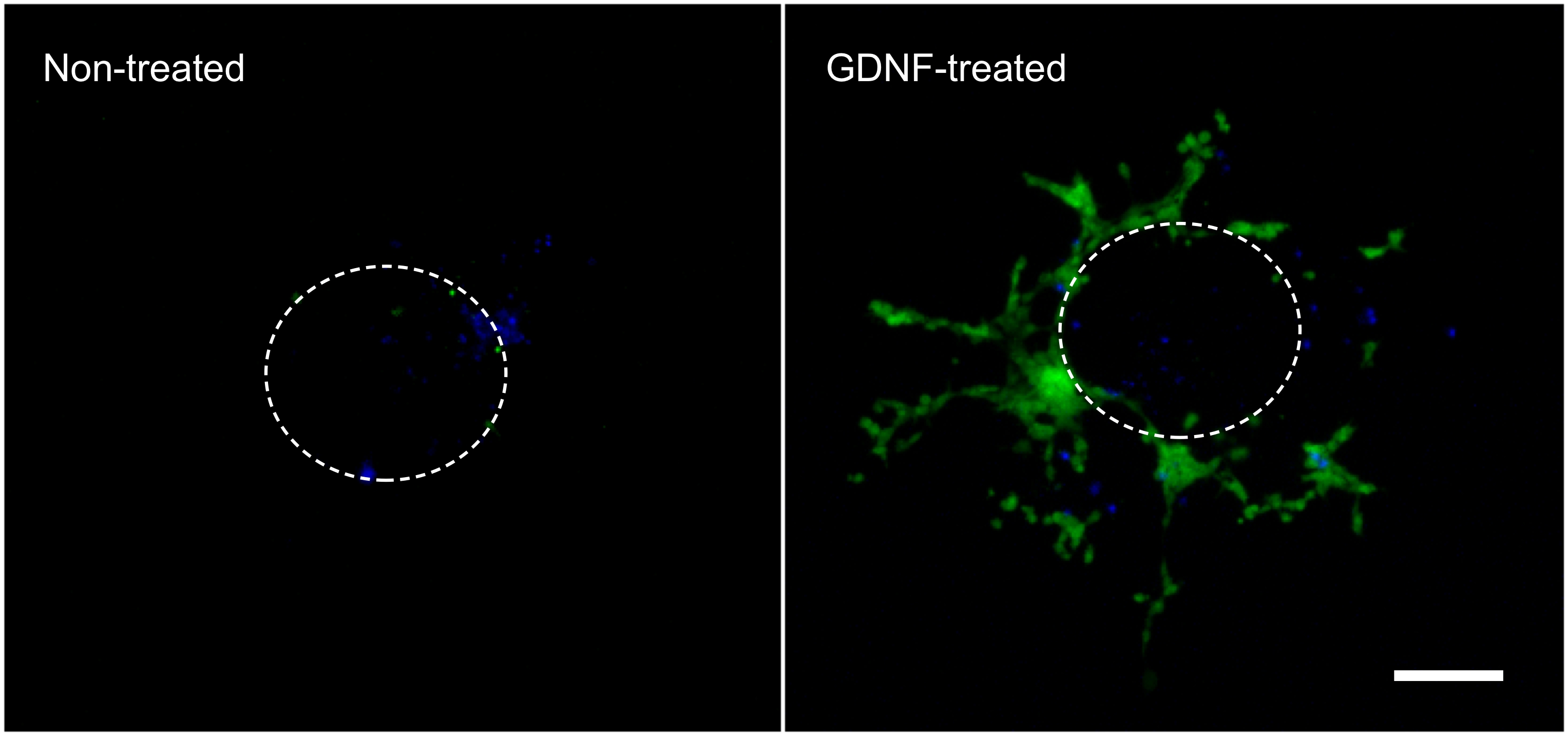
De volgende resultaten zijn representatief van wat kan worden bereikt met de hier beschreven (Figuur 1) techniek. Het gebruik van groeifactoren (bijv. GDNF) stimuleert de migratie van GFP-expressie enterische NCC uit de intestinale explantaat en in de collageengel (figuur 2). Hoewel sommige cellen uit het explantaat komen in de afwezigheid van groeifactoren, die meestal niet-GFP gemerkte en vormen passieve ingang. Het is noodzakelijk om de intestinale segment uit de collageengel te verwijderen om de resultaten te nemen, aangezien dit weefsel nog sterk bevolkt door fluorescerende cellen en anders zouden de cellen die in het collageen eronder te verbergen. Kwantificering van deze resultaten blijkt dat veel meer cellen worden aangetroffen in de collageengel wanneer GDNF aanwezig is, en dat actieve migratie plaatsvindt (figuur 3). Inderdaad, passieve cellen vond onmiddellijk onder de explantaten, in de diameter van een intestinale segment, Wher eas cellen die actief binnenvallen het collageen gel te verplaatsen uit de buurt van hun punt van oorsprong en verder verspreid.

Figuur 1. Overzicht van de explantaatkweek techniek. A) Uniform populatie van fluorescerende enterische NCC binnen de caudale dunne darm regio gebruikt om 200 μ dikke explantaten maken. Schaalbalk. 200 urn B) Het kweekmedium dat collageen gestort in 24-well platen en liet uitharden gedurende 1 uur. Vibratome plakjes agarose ingebedde embryonale darm worden afgezet op de gels (een stukje per putje), en fluorescerende enterische NCC mogen migreren uit de explantaten gedurende 3 dagen. Vervolgens worden de schijfjes af genomen voordat de beeldvorming van de cellen die het collageen gels binnengevallen.s/ftp_upload/50709/50709fig1highres.jpg "target =" _blank "> Klik hier voor grotere afbeelding.

Figuur 2. Celmigratie van de intestinale explantaat binnen de collageengel. Cellen die gedurende de 3 dagen incubatie in de afwezigheid of aanwezigheid van 10 ng / ml GDNF uit een intestinale explantaat gemigreerd werden vastgesteld, gekleurd met DAPI (blauw) en gefotografeerd te tonen GFP-gelabelde enterische NCC (groen). 70X vergroting. Schaal bar: 100 micrometer. De stippellijn geeft de geschatte grootte en de locatie van de explantatie voordat het werd genomen uit de gel. Klik hier om een grotere afbeelding te bekijken. Klik heer voor grotere afbeelding.

Figuur 3. Kwantificering van enterische NCC migratie potentieel. Het aantal en de spreiding (Feret de diameter) van GFP-expressie cellen migreren van intestinale explants na 3 dagen werd gekwantificeerd met behulp van ImageJ software 17. In beide gevallen is er een significant verschil tussen onbehandelde en behandelde GDNF-omstandigheden volgens een Student's t-test (p <0,001; *). De gemiddelde diameter van intestinale segmenten (zwarte stippellijn: 260 um) werd toegevoegd om onderscheid te maken tussen passieve en actieve ingang migratie. Nt: niet-behandelde, n:. Aantal verwerkte explantaten Klik hier voor grotere afbeelding .
Subscription Required. Please recommend JoVE to your librarian.
Discussion
We tonen hoe onze ex vivo explantaatkweek techniek kan worden gebruikt om enterische NCC migratiepotentieel nauwkeurig te kwantificeren in aanwezigheid van GDNF. Dergelijke precieze kwantificering wordt sterk vergemakkelijkt door 200 urn dikke darm vibratome gedeelten plaats van grote stukken benaderende afmetingen, zoals eerder beschreven 11,12,15. Inderdaad, deze kunnen we met een redelijk aantal cellen in een zeer reproduceerbare instelling. Opmerkelijk, de uniforme verdeling van fluorescerende enterische NCC in de meest caudale deel van de dunne darm waar de explantaat plakjes gesneden maakt ook de analyse van meerdere secties van een darm (figuur 1A). Bovendien, gezien het feit dat zowel enterale NCC en axonen het weefsel kan verlaten in dergelijke assays 11, intrekking van de explantaten aan het einde van de kweekperiode laat ons uitsluitend te richten op trekkende NCC.
Meest kritische stappen in het protocol beschreven tekst, maar zoals de welfare van de intestinale explant is van cruciaal belang voor het verkrijgen van gezonde migreren NCC, bijzondere aandacht moet worden genomen. Vermijd onderwerpen van embryonale weefsels aan plotselinge temperatuurswisselingen, vooral wanneer de darm wordt ingebed in agarose (stap 3.3). Controleer de darm bij kamertemperatuur en agarose zo koel mogelijk (nog steeds gesmolten) te vermijden "koken" van het weefsel. De intestinale explant moet gedijen op de cultuur-medium gevuld collageen gel, vaak in omvang toeneemt en morsen uit de agarose slice. Als het explantaat wordt ongezonde of erger, meestal verloren tijdens incubatie plaats dan PBS met kweekmedium bij kamertemperatuur (bijv. HEPES-gebufferde M2 of DMEM aangevuld met 10% FBS) te helpen ondersteunen tijdens dissectie.
Een belangrijke beperking van onze aanpak is dat het is gebaseerd op de beschikbaarheid van een muis lijn verlenen van een fluorescent label te migreren NCC. Bij het ontbreken van een dergelijk instrument, een antilichaam tegen migrating NCC (bijv. anti-Ret of anti-SOX10) kunnen worden gebruikt om cellen die de collageengel binnengedrongen label. Aangezien bovendien de darm micro-omgeving is veel complexer dan een simpele collageengel, verkregen met deze in vitro test niet volledig weergeeft het gedrag van enterische NCC in vivo. Aanvullende experimenten met live cell imaging wordt aangeraden om dit gedrag te beoordelen. Het is ook opmerkelijk dat naast zijn rol als een chemoattractant, GDNF bekend om proliferatie van migrerende enterische NCC 4 promoten. De mate van enterische NCC migratiepotentieel in aanwezigheid van GDNF is derhalve waarschijnlijk een mix van ware celmigratie en celproliferatie, verwant aan de in vivo mechanismen die leiden tot NCC kolonisatie van de darmen. Als een duidelijk onderscheid tussen deze twee processen is gewenst, kan de toevoeging van een celcyclus blokker (bijv. AZD 5438 18) in het kweekmedium de analyse beperken cel MIGRATIEn.
Deze techniek kan worden uitgebreid tot verschillende andere extracellulaire liganden en remmers van specifieke signaalwegen, en combinaties daar testen. Andere weefsels kunnen ook potentieel worden ontleed en deelbaar, waardoor de studie van NCC migratie in vele embryonale structuren. Gecombineerd met nieuwe en / of ongekarakteriseerde mutante muizenstammen eventuele gebreken van NCC ontwikkeling kan onze techniek worden toegepast om snel te screenen op tekortkomingen in migratiegedrag als reactie op specifieke signalering gebeurtenissen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben niets te onthullen.
Acknowledgments
Wij danken Denis Flipo voor beeldverwerking en analyse-advies, en David W. Silversides in wiens laboratorium het Gata4p [5kb]-GFP muis lijn werd gegenereerd. Onderzoek in het laboratorium Pilon wordt gefinancierd door CIHR, NSERC, FRQS en FRQNT.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J. The enteric nervous system. Dev. Biol. 366 (1), 64-73 (2012).
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}