

Summary
我々は、様々な成長因子の存在下での腸溶神経堤細胞の遊走電位の正確な定量を可能にex vivoで細胞遊走アッセイを提示する。
Abstract
神経堤細胞(NCC)は、背側神経管に由来し、開発脊椎動物の胚全体で広範囲に移動し、一過性および多能細胞集団である。末梢グリアおよびニューロンを提供することに加えて、NCCはメラノサイトだけでなく、頭蓋顔面骨格の大部分を生成します。 NCC遊走および分化は、神経管に沿った軸方向の原点と地域の異なる細胞外の合図への曝露との組み合わせによって制御される。細胞外のリガンドのような貢献は、腸神経系(ENS)、ローカルに(特に)を制御する神経核の複雑な相互接続されたネットワーク腸の筋肉の動きや腸の運動を形成する際に特に顕著である。尾ファッションに吻側に - - 将来の腸の全長にENSのほとんどが植民地化するために長い旅に着手するNCCの小さな初期プールに由来している。知られているいくつかのシグナル伝達経路のうち、腸NCCのコロニー形成に影響を与え、GDNF / RETシグナル伝達は、最も重要なものとして認識されている。確かに、消化管間葉によるRETリガンドGDNFの時空間的に制御された分泌は、胚性腸へと内誘引およびRET発現腸NCCの指導のために主に責任があります。ここでは、GDNFを含む様々な成長因子の存在下で腸溶性NCC移行電位の正確な定量化を可能にする蛍光標識されたNCCを有するトランスジェニックマウス系統を利用して、ex vivoでの細胞遊走アッセイを記載している。
Introduction
神経堤細胞(NCC)は、胚発生の間に多くのデリバティブを形成している脊椎動物に特有の過渡的な細胞型である。この細胞集団は、非神経外胚葉1に隣接して、神経板の境界で生じる。神経胚形成の間に、神経板の曲げ成形神経管の背側縁に沿って、NCCが配置されます。 NCCは、その後分離し、離れて神経管からの移行、上皮間葉転移を受ける。 NCCは、彼らが全体の腸管神経系(ENS)、腸の壁に埋め込まれた神経の神経節の相互接続されたネットワークを形成し、消化管を含む様々な胚の構造を、植民地化。最近2,3日のように、多くの遺伝子がこの複雑な構造の開発に携わってきました。
ENSのほとんどは( つまり、将来の後脳/脊髄の境界付近)迷走神経管からのNCC発信の小さなプールに由来している4。これらの神経前駆細胞は、マウスでは胎生(E)9.0を中心に前腸に到達してから、全体胚性腸にコロニーを形成するのに約e15.0までの消化管間葉内で尾側に移行します。結腸の神経前駆細胞の小サブセットはまた、最大盲腸4と反対方向に後方腸に侵入仙骨NCCによって提供される。迷走神経と仙骨の両方NCCは、ENSの完全な形成を確実にするために、複数の移行、増殖、生存および分化を促進する手がかりが必要になります。 GDNF(グリア細胞由来神経栄養因子)、エンドセリン-3、ニューロトロフィン3、BMPは(骨形成タンパク質)、ネトリン:この点では、動物モデル - - 特に遺伝子改変マウスは、いくつかの不可欠な細胞外リガンドの同定に尽力されてきただけでなく、ソニックとインディアンヘッジホッグ(ShhおよびIhhは)5月10日 。これらのうち、(トランスフェクション中に再配列)チロシンキナーゼ膜貫通受容体を介したシグナル伝達RET GDNFは番目として認識される胚性腸へと内魅力とNCCの指導のための電子の最も重要な経路。 GDNFは消化管間葉から分泌され、RET 11,12を表現腸NCCに直接化学誘引である時空間的に制御rosrrocaudal勾配を形成している。
他の機能の中でも、ENSは、腸壁内の平滑筋との相互作用を介して、消化管内での移動を規制する。ヒルシュスプルング病における腸結果の末端領域の神経の神経節が存在しない場合:影響を受けたセグメントの強壮収縮が詰まり、消化された材料の上流に蓄積し、腸や腹部の大規模な膨満につながる。ヒルシュスプルング病は、およそ1 5000での出生数を発生します。腸溶NCCの吻側 - 尾側移行パターンは、ヒルシュスプルング病の病因に主要な寄与因子であると考えられている。コロン、NCCの移行のソースから最も遠いとBの最後の部分植民地化されるowelは、ENS形成における欠陥の影響を最も受けている。腸溶NCC遊走におけるその重要な役割に応じて、GDNF / RETシグナル伝達の破壊は、ヒルシュスプルング病13の主既知の遺伝的原因である。
[5キロバイト]-GFP 14 Gata4p命名- -回遊NCCは、緑色蛍光タンパク質(GFP)で標識された良好なNCCとENSの発達を研究するために、我々は、トランスジェニックマウス系統を生成した。我々は次に、現在では、GDNFのような種々の成長因子の存在下で腸溶性NCC移行電位の正確な定量化を可能にする他のグループ11,12,15によって発表された研究から適合ex vivoでの細胞遊走アッセイを、完成。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
倫理声明
マウスを含む実験は医学研究に使用される動物の世話や操作のための動物実験ガイドラインのカナダの評議会後に実施された。動物の操作を含むプロトコルはモントリオールのケベック大学の施設内倫理委員会によって承認された(化委員会Institutionnelデ·保護DES Animaux、参照番号0512-R3-650から0513)。
1。コラーゲンゲルの調製
組織培養フードの下で、無菌方式で動作します。
- 標準的な抗生物質を含む完全な5X DMEM(イーグル改変必須培地)を準備します。のDMEM粉末3.37グラムと水20ml中のNaHCO 3の0.925グラムを溶解する。 0.22μmのフィルターを通すことにより滅菌する。無菌100Xペニシリン/ストレプトマイシン、および滅菌熱不活性化ウシ胎児血清25ミリリットルの2.5ミリリットルを追加します。 4℃で保存する
- 氷の上に、800μLをミックス完全5XのDMEM600μlのと1N NaOHを17μlのコラーゲンI溶液(0.02 N酢酸、濾過滅菌中3.77 mg / ml)を。 3ミリリットルの最終容量に滅菌水で希釈します。ミックス内の関連する成長因子が含まれる。私たちは通常、腸NCCの移行を刺激するために10 ngの/ mlでGDNFを使用しています。
- 24ウェルプレート内の単一行の各ウェルに沈着約480μL。泡を避けてください。残りの行は、細胞遊走に対する他の成長因子の効果を試験するために使用することができる。
- コラーゲンは、37℃で滅菌インキュベーター中で少なくとも1時間重合しましょ
2。動物16の解剖
- 交配を設定し、翌朝膣栓を確認してください。デイE0.5が膣栓が検出されたその日の正午であること、女性を隔離し、E12.5まで12日待ちます。
- イソフルランで妊娠マウスを麻酔して、CO 2吸入によって安楽死させる。
- 70%エタノールでマウスをスプレーします。腹部Sを持ち上げる親族や解剖ハサミで腹腔を開きます。
- 氷冷PBS(リン酸緩衝生理食塩水)を充填したガラスペトリ皿に子宮を除去する。各胚の移植部位を分離するために、個々のdeciduum腫脹との間で横方向に子宮をカット。
- 氷冷PBSで満たされた別のガラスシャーレに別々に移植部位に取り組んでいます。解剖顕微鏡下で、子宮の筋肉層を除去するために細かい鉗子を使用しています。
- 胚を明らかにするために、内臓卵黄嚢と羊膜を開きます。彼らが開発し、腸と密接に絡み合っているように、胎盤/内臓卵黄嚢に胚を結ぶ血管を切断する際には注意してください。
- 首に胚の頭を切断。
- ただ濃い赤色の肝臓の上に、胚の腹腔内に、閉じた鉗子を挿入し、腹腔内の横方向の開口部を作るために(圧力をかけて停止する)自体の鉗子を開いてみましょう。に向けて開かれた鉗子を引き下げ完全に腹部を開くには、胚の後端。
- 肝臓の背後にある結合組織をつかみ、(コロンは肛門に添付されている)、腸を壊さないように注意しながら、腹部から内臓を引き出します。
- 胚の他の部分から勇気を解放するためにコロンをカット。カットは結腸に沿った任意の位置で行うことができる。後でテール部分を予約。
- 盲腸から結合組織、腸の後、残りの部分を引き出す。そうしながら、腸を巻かないように注意してください。
- いくつかが存在する場合、肝臓と胃(小腸の吻側端にある)だけでなく、中腎と生殖隆起を切り取る。
- 小腸を分離します。繰り返しますが、腸をニップしないように注意してください。今後は、腸組織のrosrrocaudal向きは頭側の端に存在鋭い曲率を使用して追跡することができます。実施する前に、できるだけ短い時間、室温でPBSで小腸を残す寝具(ステップ3.3を参照)。
- 最終的に、正確に胚発生の段階を決定するために、各胚のための尾体節の数を記録します。
3。胚性腸の切片
- 胚の解剖を続行する前に100mlのPBS(リン酸緩衝生理食塩水)中に1.5グラムアガロースを融解し、50℃で維持
- 埋め込 み型内に注ぐ溶融アガロース( 例えば、aは約1/4の管壁を切除するために縦に切断された2mlのマイクロ遠心チューブを閉じた状態)。アガロースは、約42〜45℃に冷ましてみましょう、それは触るとわずかに暖かいはずです。
- ただ、アガロースが固化する前に、胚性腸に埋め込む(これは鉗子の先端を用いて評価することができ、36〜38℃付近が発生します)。折り畳んだ吻側年末までにピンセットで腸を持ち、金型の長さに沿ったアガロースを通して非常にゆっくりと引き出します。これはストレートアガロースセットしながら、腸を維持するのに役立ちます。 Tをリリースとすぐに移動しているレジストをし始めると問題。腸の吻側 - 尾側の向きを追跡する。
- アガロースが完全に設定されていることを確認するために、冷蔵庫、2〜3分で金型を置く。
- (開かれたエッペンドルフチューブの外にスライドさせて)金型からアガロースを取り出します。刃で、腸に垂直カットを作り、両端に余分なアガロースを取り出します。
- /アガロース振動刃ミクロトームの金属ステージを下にブロックし、腸の吻側端を接着します。腸/アガロースブロックの側面に余分なアガロースをトリミング。
- 振動ミクロトーム室に金属ステージをマウントします。必要に応じて腸をできるだけ垂直になるように(したがって、ブレードに垂直)ステージの角度を調整する。氷冷PBSで標本をカバーしています。バッファ面の下に数ミリまでミクロトームの刃を持参してください。
- その電子を確保し、尾最も小腸の200ミクロンビブラトーム横カットを作るACHアガローススライスは完全な腸セクションが含まれています。
4。腸管外植片の文化
- 静かに、各ウェルの中央に向かって1スライスを置く、ピンセットでコラーゲンゲルに新たに切断した腸/アガローススライス、フラットを堆積させる。
- 外植片からNCCの移動を可能にするために、湿った5%CO 2雰囲気中、37℃で3日間インキュベートする。
- ピンセットで非常に優しくコラーゲンゲルから腸/アガローススライスを取る。以下のゲルを損傷しないように注意してください。
- 細胞移動パターンが妨げられないことを確実にするために、その後のインキュベーションおよび洗浄ステップの間に直接コラーゲンゲルに触れないようにしてください。室温でよく、1時間あたりの(PBS中)を4%PFA(パラホルムアルデヒド)500μlで固定してください。
- ウェルあたり500μlのDAPI(4 '、6 - ジアミジノ-2 - フェニルインドール)溶液(PBS中の5μg/ ml)を用いて固定液を交換し、室温で10分間インキュベートする。
- 各ウェル3Xを洗う5分間のPBS500μlで。
- 各ウェル内のコラーゲンゲル内に埋め込まれた蛍光細胞(GFPおよびDAPIチャネル)写真。
5。画像解析
私たちは、移植片培養した後に生成された画像を処理し、定量化するためにImageJの17の大規模な使用をした。
- 蛍光細胞写真と同じ倍率でマイクロスライドを画像化することから始めます。 (ストレートライン·ツールを使用して)ピクセル単位で1ミクロンの長さを測定します。
- (/設定スケールを分析、ピクセル/ミクロンの入力数)スケールを設定します。
- 各GFP蛍光写真は、8ビットグレースケール(Image/Type/8-bit)に画像形式を変更します。
- 信号(画像/調整/明るさ/コントラスト)の強さを調整します。
- 必要に応じて(;ローリングボールの半径を調整プロセス/背景を引く)、バックグラウンドノイズを差し引く。
- 細胞および細胞塊を(画像/ /しきい値を調整)、ハイライト表示し、しきい値を設定するN塊(プロセス/バイナリ/流域)を分割するために流域を適用します。
- 関心領域(ROI)を指定し、細胞数の統計情報を生成する粒子を分析(粒子を分析/分析、設定されたサイズの最小値は、残りのピクセルを除外する)。
- フェレ径(/セット測定の分析)を含むように、測定オプションを設定します。
- グループのROIと、フェレ径を決定するために、全体として細胞の広がり(ANALYZE /ツール/ ROIマネージャー/詳細/ OR、そしてROIマネージャー/追加し、ROIマネージャー/測定)の指標を測定します。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
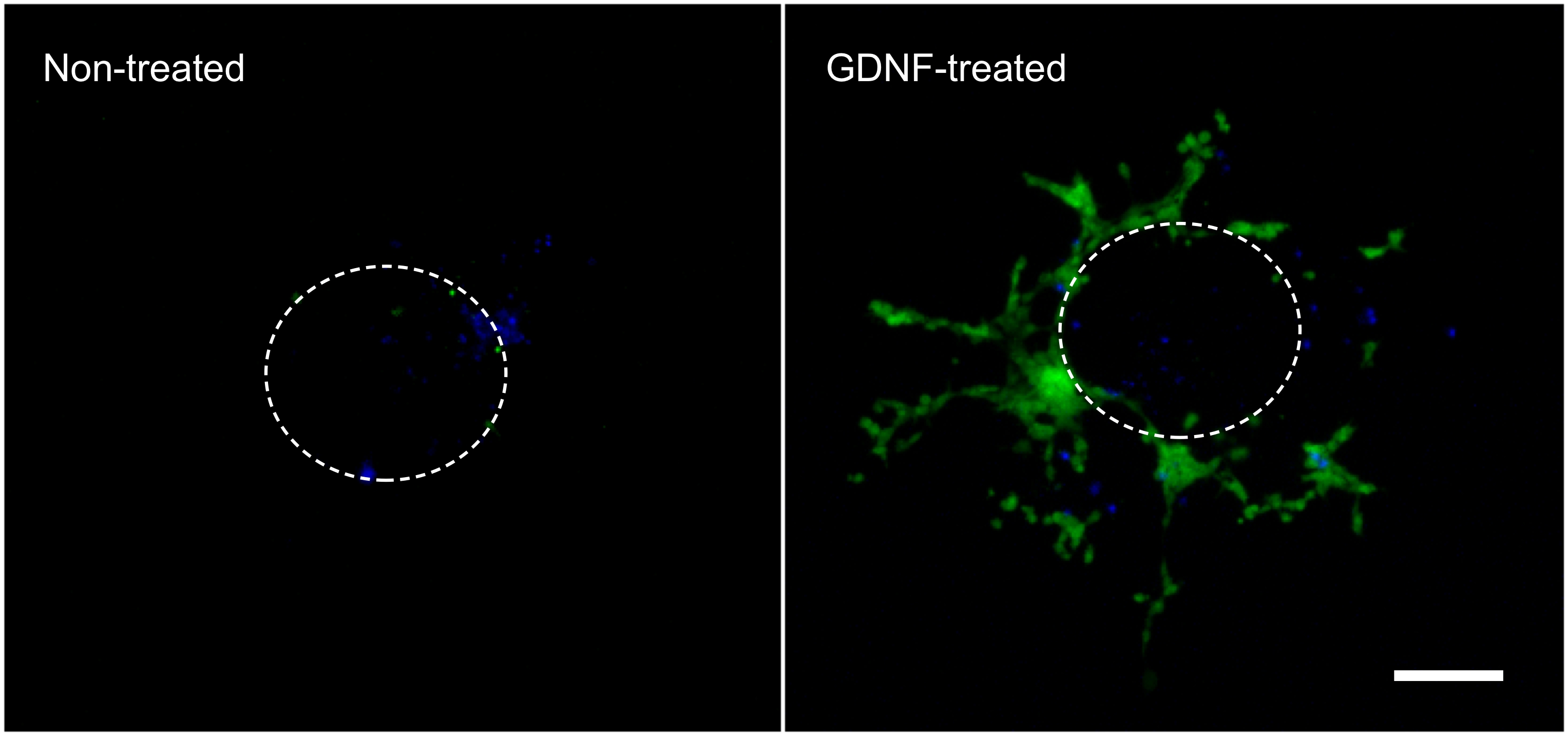
以下の結果は、ここで記載された技術( 図1)で得られるものの代表である。増殖因子の使用( すなわち、GDNF)は、腸の外植片からコラーゲンゲル( 図2)にGFPを発現する腸溶性NCCの遊走を刺激する。一部の細胞は、成長因子の非存在下で外植片から出てくるが、これらは主にGFP-標識されず、パッシブエントリーを表す。この組織はまだ頻繁に蛍光細胞によって取り込まれ、そうでなければ下にコラーゲンに横たわってセルを非表示になるように、それは、結果を記録するために、コラーゲンゲルから腸スライスを除去する必要がある。これらの結果の定量は、GDNFが存在する場合、より多くの細胞がコラーゲンゲル内に見出されることを示し、その活性な移行が( 図3)で行われる。確かに、受動的な細胞は腸管スライスの径内、WHER、すぐに外植片の下で発見された積極的にコラーゲンゲルに侵入EAS細胞が離れてそれらの起源の点から移動して、さらに広がった。

図1。外植片培養技術であって、a)200μ厚の外植片を作るために使用される尾小腸領域内の蛍光腸NCCの制服人口の概要 。スケールバー:200ミクロンB)コラーゲンを含む培養培地を24ウェルプレートに堆積し、1時間硬化させておく。アガロース包埋胚腸のビブラトーム切片をゲル(ウェル当たり1つのスライス)上に堆積され、蛍光腸溶NCC 3日間外植片から遊走させる。スライスは、その後、コラーゲンゲルに侵入した細胞を画像化する前に取り出される。s/ftp_upload/50709/50709fig1highres.jpg "ターゲット=" _blank ">拡大画像を表示するにはここをクリックしてください。

図2。腸管外植片から出てコラーゲンゲル内細胞移動。10 ng / mlのGDNFの存在下または非存在下で3日間のインキュベーションの間に腸の外植片から遊走した細胞は、固定DAPI(青)で染色し、に写真撮影したGFP標識腸NCC(緑)を示す。 70X倍率。スケールバー:100μmである。それはゲルを外された前点線は外植片のおおよそのサイズと位置を表しています。 大きな画像を見るにはここをクリックしてください。 クリックH拡大画像を表示するERE。

図3。腸溶NCC移行電位の定量。3日後に腸の外植片から移動GFP発現細胞の数およびスプレッド(フェレ径)が17 ImageJソフトウェアを用いて定量した。両方の場合において、未処理およびGDNF処理条件との間の有意差は、スチューデントのt検定(; *はp <0.001)に記載がある。腸のスライス(黒い点線:260ミクロン)の平均粒径は、パッシブエントリーとアクティブ移行を区別するために含まれていました。 NT:無処理、N:処理された外植片の数は、 拡大画像を表示するには、ここをクリックしてください 。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
我々のex vivoでの外植片培養技術を正確GDNFの存在下で腸溶性NCCの移行の可能性を定量化するために使用することができる方法を示す。このような正確な定量が大幅に以前に11,12,15を説明するように、厚さ200μmのビブラトーム腸のセクションを使用しての代わりに、おおよその大きさの大部分によって容易になる。確かに、これは、私たちは再現性の高い環境でのセルの合理的な数で作業することができます。注目すべきは、外植片切片が切断される小腸の尾最領域内の蛍光腸溶NCCの均一な分布は、単一の消化管( 図1A)からの複数のセクションの分析を可能にする。また、腸NCCと軸索の両方が培養期間の終了時に、このようなアッセイ11、外植片の撤退で組織を終了することができ、私たちは渡り鳥NCCのみに集中することができますことを考える。
最も重要なステップは、しかし、Wのように、プロトコルテキストに概説された腸管外植片のelfareは、特定の注意が必要、健康的な移行NCCを得るために最も重要です。腸アガロース(3.3ステップ)に埋め込まれている場合は特に、急激な温度変化に胚組織を与えないでください。腸が組織を「料理」を避けるために、室温、できるだけ涼しい(それでも溶けた)アガロースであることを確認します。腸管外植片は、多くの場合、大型化し、アガローススライスからこぼれる、培養液で満たされたコラーゲンゲル上で繁栄する必要があります。外植片が不健康または悪化表示された場合は、インキュベーション中に死ぬ傾向があり、解剖中にそれを維持を支援する( 例えば HEPES緩衝M2またはDMEM、10%FBSを添加)を室温で培養培地を含むPBSを交換してみてください。
我々のアプローチの主要な制限は、NCCの移行に蛍光標識を付与するマウス株の利用可能性に依存することである。このようなリソースの非存在下では、mに対する抗体NCCをigrating( 例えば 、抗Retのまたは抗SOX10)は、コラーゲンゲルに侵入し細胞を標識するために使用できます。また、消化管の微小環境ははるかに複雑簡単なコラーゲンゲルよりもあることを考えると、このin vitroアッセイで得られた結果は、完全に生体内で腸NCCの動作を反映しない場合があります。生細胞イメージングを含む追加的な実験が、この挙動を評価することをお勧めします。これは、化学誘引物質としてのその役割に加えて、GDNFは、腸溶NCC 4マイグレーションの増殖を促進することが知られていることも注目すべきである。 GDNFの存在下で腸NCCの移行の可能性の我々の測定値は、おそらくこのように腸のNCCの定着につながるの生体内のメカニズムに似た真の細胞遊走および細胞増殖のミックスです。これらの二つのプロセスの間に明確な区別が所望される場合、培養培地における細胞周期遮断薬( 例えば、AZD 5438 18)の添加は、細胞migratioに分析を制限することができN。
この技術は、種々の他の細胞外のリガンドならびに特定のシグナル伝達経路の阻害剤、およびその中の任意の組み合わせをテストするために拡張することができる。他の組織はまた、潜在的に多くの胚の構造におけるNCCの移動の研究を可能に解剖し、切片化することができます。新規かつ/またはNCC開発の可能性欠陥の特徴付けされていない変異マウス系統と組み合わせることで、私たちの技術は急速に特異的なシグナル伝達イベントに応じて移行挙動の不備をスクリーニングするために適用することができます。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者らは、開示することは何もありません。
Acknowledgments
我々は、画像処理と解析アドバイスデニスFlipoに感謝し、デビッド·W·シルバーサイズが実験室でGata4p [5キロバイト]-GFPマウスラインが生成されました。ピロンの実験室での研究はCIHR、NSERC、FRQSとFRQNTによって資金を供給される。
Materials
| Name | Company | Catalog Number | Comments |
| DMEM powder | Wisent | 219-010-XK | |
| NaHCO3 | Bioshop | SOB999 | Biotechnology grade |
| Steriflip vacuum filtration system (0.22 micron) | EMD Millipore | SCGP00525 | |
| Penicilin-Streptomycin solution, 100x | Wisent | 450-201-EL | |
| Fetal bovine serum | Wisent | 095-150 | High quality grade |
| Collagen I | BD biosciences | 354236 | |
| NaOH | Bioshop | SHY700 | Diluted from 10 N stock then sterile-filtered |
| GDNF | Cedarlane | CLCYT305 | |
| Falcon 24-well Plate | BD biosciences | 353047 | |
| Dissecting scissors | Fisher Scientific | 089515 | |
| Glass Petri dish | VWR | 89000-306 | |
| PBS | Sigma | P5493 | Cell culture grade |
| Dissecting microscope | Leica | M125 | |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | |
| Agarose | Bioshop | AGA001 | Biotechnology grade |
| Surgical blade | Feather | 21 | |
| All Purpose Instant Krazy Glue Pen | Krazy Glue | KG824 | |
| HM 650V Vibrating-Blade Microtome | Thermo Scientific | 920110 | |
| Paraformaldehyde | Sigma-Aldrich | P6148 | |
| DAPI | Sigma-Aldrich | D9564 | |
Table of specific reagents and equipment. |
|||
References
- Bronner, M. E., Le Douarin, N. M. Development and evolution of the neural crest: An overview. Dev. Biol. 366 (1), 2-9 (2012).
- Bergeron, K. F., Silversides, D. W., Pilon, N. The developmental genetics of Hirschsprung's disease. Clin. Genet. 83 (1), 15-22 (2013).
- Obermayr, F., Hotta, R., Enomoto, H., Young, H. M. Development and developmental disorders of the enteric nervous system. Nat. Rev. Gastroenterol. Hepatol. 10 (1), 43-57 (2012).
- Sasselli, V., Pachnis, V., Bursn, A. J. The enteric nervous system. Dev. Biol. 366 (1), 64-73 (2012).
- Sanchez, M. P., Silos-Santiago, I., et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 382 (6586), 70-73 (1996).
- Baynash, A. G., Hosoda, K., et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell. 79 (7), 1277-1285 (1994).
- Chalazonitis, A., Pham, T. D., et al. Neurotrophin-3 is required for the survival-differentiation of subsets of developing enteric neurons. J. Neurosci. 21 (15), 5620-5636 (2001).
- Goldstein, A. M., Brewer, K. C., Doyle, A. M., Nagy, N., Roberts, D. J. BMP signaling is necessary for neural crest cell migration and ganglion formation in the enteric nervous system. Mech. Dev. 122 (6), 821-833 (2005).
- Jiang, Y., Liu, M. T., Gershon, M. D. Netrins and DCC in the guidance of migrating neural crest-derived cells in the developing bowel and pancreas. Dev. Biol. 258 (2), 364-384 (2003).
- Ramalho-Santos, M., Melton, D. A., McMahon, A. P. Hedgehog signals regulate multiple aspects of gastrointestinal development. Development. 127 (12), 2763-2772 (2000).
- Natarajan, D., Marcos-Gutierrez, C., Pachnis, V., de Graaf, E. Requirement of signaling by receptor tyrosine kinase RET for the directed migration of enteric nervous system progenitor cells during mammalian embryogenesis. Development. 129 (22), 5151-5160 (2002).
- Young, H. M., Hearn, C. J., et al. GDNF Is a chemoattractant for enteric neural cells. Dev. Biol. 229 (2), 503-516 (2001).
- Amiel, J., Sproat-Emison, E., et al. Hirschsprung disease, associated syndromes and genetics: a review. J. Med. Genet. 45 (1), 1-14 (2008).
- Pilon, N., Raiwet, D., Viger, R. S., Silversides, D. W. Novel pre- and post-gastrulation expression of Gata4 within cells of the inner cell mass and migratory neural crest cells. Dev. Dyn. 237 (4), 1133-1143 (2008).
- Nagy, N., Goldstein, A. M. Endothelin-3 regulates neural crest cell proliferation and differentiation in the hindgut enteric nervous system. Dev. Biol. 293 (1), 203-217 (2006).
- Nagy, A., Gertsenstein, M., Vintersen, K., Behringer, R. Manipulating the mouse embryo: a laboratory manual. , 3rd ed, CSH Press. Cold Spring Harbor. 209-250 (2003).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671-675 (2012).
- Byth, K. F., Thomas, A., et al. AZD5438, a potent oral inhibitor of cyclin-dependent kinases 1, 2, and 9, leads to pharmacodynamic changes and potent antitumor effects in human tumor xenografts. Mol. Cancer Ther. 8 (7), 1856-1866 (2009).

{kind=link}