

Summary
We beschrijven beeldvormingsbenaderingen we gebruiken om de verdeling en mobiliteit van de getransfecteerde fluorescerende eiwitten in het endoplasmatisch reticulum (ER) inwoner via de confocale beeldvorming van levende cellen te onderzoeken. We hebben ook ultrastructureel analyseren het effect van hun expressie op de architectuur van deze subcellulaire compartiment.
Abstract
De lipiden en eiwitten in eukaryote cellen worden continu uitgewisseld tussen celcompartimenten, hoewel deze behouden hun karakteristieke samenstelling en functies, ondanks de intense interorganelle moleculaire verkeer. De in dit document beschreven technieken zijn krachtig middel van het bestuderen van eiwitten en lipiden mobiliteit en handel in vivo en in hun fysiologische omgeving. Fluorescentieherstel na fotobleken (FRAP) en fluorescentie verlies fotobleken (FLIP) worden veel gebruikt levende cellen beeldvormende technieken voor het bestuderen van intracellulair verkeer door de exo-endocytische route, de continuïteit tussen organellen of subcompartimenten, de vorming van eiwitcomplexen en eiwit lokalisatie in lipide microdomains, die kan worden waargenomen onder fysiologische en pathologische omstandigheden. De beperkingen van deze benaderingen vooral door het gebruik van fluorescente fusieproteïnen en hun potentiële nadelen zijn kunstmatig overexpressieionen in cellen en de mogelijkheid van verschillen in de vouwen en lokalisatie van merktekens en natieve eiwitten. Ten slotte de oplosgrens optische microscopie (ongeveer 200 nm) niet onderzoek naar de fijnstructuur van het ER of de specifieke subcompartimenten die kan ontstaan in cellen onder stress staan (bijvoorbeeld hypoxie, geneesmiddeltoediening, de over-expressie van transmembraan ER resident eiwitten) of onder pathologische omstandigheden, combineren we live-cell imaging gekweekte getransfecteerde cellen met ultrastructurele analyse op basis van transmissie elektronenmicroscopie.
Introduction
De ontdekking van groen fluorescerend eiwit (GFP) en de spectrale varianten en de parallelle ontwikkeling van fluorescentiemicroscopie, zijn volledig nieuwe wegen voor het onderzoek van proteïne gedrag in cellen geopend. Technieken zoals fluorescentie herstel na fotobleken (FRAP) en fluorescentie verlies fotobleken (FLIP), die mogelijk zijn door de intrinsieke capaciteit van fluoroforen hun fluorescentie blussen onder intense belichting, gebaseerd op confocale live cell imaging en het gebruik van getransfecteerde fluorescente fusie-eiwitten 1-3. Zij worden wijd gebruikt om niet alleen de lokalisatie van eiwitten, maar ook hun mobiliteit en vesiculair transport, die belangrijk aanwijzingen betreffende hun functie 4 kan onthullen beoordelen.
De unieke eigenschap van eukaryote cellen is de aanwezigheid van intracellulaire compartimenten die specifieke lipiden en eiwitten samenstellingen. Hoewel organellen zijn fysiek isolated., moeten ze communiceren met elkaar en delen moleculaire componenten om cellulaire homeostase. De secretieroute garandeert dat de gesynthetiseerd in het ER eiwitten en lipiden bereikt de juiste eindbestemming in waarin zij hun functie uitoefenen. Intracellulaire organellen kan ook worden aangesloten door middel van dynamische contact sites die moleculen (lipiden) rechtstreeks worden uitgewisseld tussen de compartimenten. Bovendien hebben veel eiwitten geassembleerd in grote heteromere complexen of geassocieerd met specifieke soorten lipiden (lipide rafts / microdomeinen) teneinde functioneel actief of hun eindbestemming worden vervoerd. Al deze biologische aspecten grote invloed op de kinetische eigenschappen van eiwitten, en derhalve passend worden onderzocht door middel van de hieronder beschreven technieken.
Onze fractie heeft veel gebruikt FRAP en Flip combinatie met elektronenmicroscopie om de architectuur van de ER en de verschillende subsystemen bestuderendomeinen. De ER is het eerste station van de secretieroute en speelt een sleutelrol bij eiwit en lipide sorteren 5. Het is een zeer dynamische organel waarvan verschillende subdomeinen weerspiegelen de vele verschillende functies (dwz eiwitten en lipiden biosynthese en mensenhandel, het vouwen van eiwitten, Ca 2 + opslag en afgifte, en lichaamsvreemde metabolisme). Hoewel ze morfologisch, ruimtelijk en functioneel verschillende, deze domeinen continu met elkaar en hun relatieve abundantie kunnen in cellen worden gemodificeerd onder fysiologische en pathologische omstandigheden. De bekendste en meestal ruimtelijk gescheiden domeinen van de ER zijn de nucleaire envelop, en de gladde en ruwe ER; echter, hebben wij en anderen aangetoond dat er ER structuren met een meer uitgebreide architectuur en drie-dimensionale organisatie gewerkt in diverse celtypen en weefsels onder fysiologische omstandigheden die ook kan worden geïnduceerd door middel van stressvolle stimuli zoals hypoxie, drugadministratie, of de over-expressie van ER-ingezeten transmembraaneiwitten 2,6 (en referenties daarin).
We hebben onlangs aangetoond dat de aanwezigheid van dergelijke structuren celmodellen van menselijke ziekten 1,7. Afkomstig uit de gestapelde cisternae van gladde ER kregen ze de verzamelnaam georganiseerde gladde endoplasmatisch reticulum (OSER) 2003 6 weliswaar ook bekend als karmellae, lamellen en kristalloïde ER op basis van de architectuur die evenals de grootte, kan variëren. Nadat de cellen getransfecteerd met GFP gefuseerd aan de cytosolische regio-tail verankerd (TA) ER-resident eiwitten (d EGFP-ER), de zwak dimeriseren neiging van GFP in trans drastisch verandert de organisatie en structuur van de ER. FRAP en FLIP experimenten toonden aan dat d EGFP-ER vrij diffunderen in OSERs, en dat het zich van de reticulaire ER naar de OSER en vice versa </ Em> geeft aan dat de aggregaten zijn continu met de omringende reticulaire ER. Ultrastructurele analyse heeft ons toegestaan om de fluorescentie gegevens correleren met een gedetailleerde beschrijving van OSER architectuur en organisatie op nanoschaal niveau: OSERs zijn altijd opgebouwd uit stapels gepaarde cisternae van gladde ER, maar kan verschillende vormen van ruimtelijke organisatie, zoals regelmatig gerangschikte sinusvormige hebben arrays of kransen, of zeshoekige "kristalachtige" tubulaire arrays. Deze herschikkingen tot kubische morfologie 8 die, zoals ze gevonden in cellen onder fysiologische omstandigheden 9 en na stress zoals hypoxie 10, medicamenteuze behandeling 11 en kanker 9, kan aanzienlijk potentieel als ultrastructurele markers.
Na deze eerste demonstratie met GFP-fusie-eiwitten, gebruikten we imaging experimenten om de proliferatie van ER domeinen analyseren reactie op farmacologische behandelingen 12, assess de neiging van fluorescente proteïnen oligomerise van cellen 13 en de rol van een mutant ALS-TA gebonden eiwit bij de vorming van intracellulaire aggregaten van ER oorsprong die zijn pathogeniciteit 1,8 relevant zijn onderzoeken. Er is gesuggereerd dat de vorming van intracellulaire aggregaten (dat voorkomt in vele neurodegeneratieve ziekten 14) een beschermend mechanisme om de interacties tussen toxische mutant eiwitten en de omringende celbestanddelen 15 voorkomen kan worden.
Wat volgt is een beschrijving van een combinatie van optische en elektronenmicroscopie methoden voor onderzoek constructen waarvan de C-terminale hydrofobe domeinen in het membraan van het ER geplaatst, en een analyse van hun dynamisch gedrag en de effecten van hun over-expressie op ER- architectuur in gekweekte cellen (zie Figuur 1 een stroomdiagram van het experimentele protocol).
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Plasmide, Cultuur van de Cel, en transfectie met ER fluorescerende eiwitten
- De plasmide die in deze studie bestaat uit een verbeterde versie van GFP gefuseerd aan zijn C-terminus aan de staart gebied van de ER isovorm van ratten cytochroom b (5) (hier afgekort als b (5)) via een linkersequentie. De staart gebied bevat de hele reeks (Pro94-Asp134) dat membraangeassocieerde na de trypsine splitsing van inheemse b (5), met inbegrip van de 17-residu TMD (Transmembrane domein), geflankeerd door upstream en downstream polaire sequenties (UPS en DPS) blijft . De linker bevat de myc epitoop gevolgd door [(Gly) 4Ser] 3, en de volledige cDNA wordt in de Hind3-Xba1 plaatsen van de zoogdierlijke expressievector pCDNA3 ingevoegd. De details van de constructie van dit plasmide is beschreven in een eerdere publicatie waarin wordt verwezen als GFP-ER 16.
- Kweek COS-7 cellen in Dulbecco's Modified Eagle Medium (DMEM) gesupplementeerd met 10% fetal runderserum, 2 mM L-glutamine, 1% penicilline / streptomycine in een incubator bij 37 ° C en 10% CO2.
- Transfectie. Plaat 3 x 10 5 cellen in een ronde glazen dekglaasje op een plaat met 6 putjes en de volgende dag transfecteren het jetPEI zoals beschreven door de fabrikant. Merk op dat de optimale jetPEI / DNA verhouding werd getest om de maximale transfectie-efficiëntie afhankelijk van het plasmide en gebruikte cellijn vastgesteld: in ons geval een jetPEI: DNA verhouding van 2:1 tot 70-80% transfectie efficiëntie.
2. Levende fluorescentie confocale microscopie
- Live-cell imaging. Plaats het dekglaasje waarop de getransfecteerde cellen geënt in een stalen kweek celkamer 24 mm dekglaasjes gevuld met DMEM w / o fenolrood, aangevuld met 10% FBS, 2 mM L-glutamine, 1% pen / strep, 25 mM HEPES , 50 ug / ml cycloheximide en 1:100 OxyFluor te voorkomen dat de monsters van fotobleken. Een SP5 confocale microscoope uitgerust met een temperatuurgestuurde CO2 incubator (37 ° C en 5% CO2) wordt gebruikt voor live cell imaging experimenten met d GFP-ER gevisualiseerd met een 488 nm laser en een 525/50 band pass emissiefilter.
- Fluorescentie herstel na fotobleken (FRAP). Teken een gebied van belang (ROI), overeenkomend met een OSER structuur en ontkleuren met 20 iteraties en een combinatie van 488 nm (100% van een 30 mW Argon laser, overeenkomend met 5,5-6 μW bij het monster) en 405 nm (60% van een 30 mW 405 Diode laser, wat overeenkomt met 11,6 μW bij de steekproef) lasers die, in onze ervaring, leidt tot een efficiënte en snelle fotobleking.
- Noteer het herstel van fluorescentie in het gebleekte ROI door het nemen van een enkel frame elke 10 seconden gedurende 10 minuten (pixel tijd = 1,61 usee / px).
- Fluorescentie verlies in photobleaching (FLIP). Teken een ROI overeenkomt met een OSER structuur en bleek zoals hierboven beschreven. Het bleken wordt herhaald om de 30 sec,en post-bleken beelden worden opgenomen na elke 10 seconden gedurende 30 minuten (pixel tijd = 1,61 usee / px).
- FRAP en FLIP analyse. Alle beelden worden geanalyseerd met behulp van ImageJ software ( http://rsbweb.nih.gov/ij/download.html ). In de FRAP experimenten wordt de fluorescentie herstel van de gebleekte ROI tijd gemeten en genormaliseerd naar het totale fluorescentie van gebleekt cel, die altijd gecontroleerd tijd constant zijn.
- Voor de FLIP experimenten, trekken een ROI buiten de gebleekte OSER en die de hele cel. Meet de fluorescentie-intensiteit in de tijd en normaliseren de fluorescentie niveaus van een ROI opgesteld op ongebleekte cel om te corrigeren voor eventuele afname in fluorescentie veroorzaakt door de beeldvormende zelf.
- In alle experimenten, afgetrokken het achtergrondsignaal (bepaald in een gebied buiten de cellen) van de fluorescentie-intensiteiten van de ROI. Tot slot, de resultaten worden met behulp van GraphPad Prism software.
3. Ultrastructurele Analyse door middel van Transmissie Elektronen Microscopie
Gegeven de toxiciteit van vele reagentia dienen alle procedures uitgevoerd dragen passende lab. jas en handschoenen onder een afzuigkap.
- Na het verwijderen van het dekglaasje van de petrischaal, bevestig de overige cellen gekweekt op de bodem van de schaal als een monolaag met gefiltreerde 2% glutaaraldehyde in 0,1 M cacodylaatbuffer, pH 7,4, gedurende 10 minuten bij kamertemperatuur.
- Schraap de cellen met behulp van een Teflon schraper en breng ze in 1,5 ml Eppendorf buizen. Pellet de cellen door middel van centrifugatie bij 9.000 g gedurende 10 minuten. Verwijder de bovenstaande vloeistof, voeg verse fixatief, en laat het een nacht bij 4 ° C.
- Was de korrels met bufferoplossing en vervolgens na vast met een oplossing van 1% osmiumtetroxide in cacodylaatbuffer gedurende 1 uur bij kamertemperatuur geroerd.
- Spoelen met MilliQ water en en bloc vlek met1% uranylacetaat met gedestilleerd water gedurende 20-60 minuten.
- Dehydrateer de monsters in toenemende ethanolreeks (70%, 80%, 90%, 100% en 100% gedurende 10 min. elk), en was tweemaal kort propyleenoxide (15 minuten elk).
- Infiltreren de monsters in een mengsel van propyleenoxide + Epon (01:01) (vanaf 2 uur tot overnacht).
- Embed in Epon epoxyhars uitgehard bij 60 ° C gedurende ten minste 24 uur.
- Sectie het handmatig geknipt hars blokken met behulp van een ultramicrotoom LEICA UC6 uitgerust met een 45 ° diamant mes om secties te verkrijgen met een dikte van 60-70 nm. Verzamel de hoofdstukken over 300 mesh koperen roosters.
- Vlekken op de secties op de grid met een verzadigde oplossing van uranylacetaat (20 min) en loodcitraat (7 min), de roosters grondig door ze onder te dompelen in bi-gedistilleerd gefilterd water, en laat ze drogen op kamertemperatuur.
- De gebrandschilderde roosters worden waargenomen met behulp van een Tecnai G2 transmissie-elektronenmicroscoop, en de beelden worden vastgelegd met behulp van een bottom-gemonteerde CCD-camera op verschillende uiteindelijke vergrotingen (doorgaans variërend van 6,000-39,000 X).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Figuur 2 toont een voorbeeld FRAP studie mobiliteit eiwit. De mobiliteit van d EGFP-ER eiwit blijkt uit de snelle fluorescentie herstel na fotobleken in gebleekt OSERs. Voor de kwantitatieve analyse, de rust en de mobiele fractie werden afgeleid uit experimenteel gemeten gegevens door het aanbrengen van de volgende mono-exponentiële vergelijking:
F (t) = F bericht + (F rec-F post) (1-e-t / τ)
waar F post is de fluorescentie-signaal na photobleaching, F rec is de maximale fluorescentie herstel waarde die wordt bereikt na het bleken, t het moment van inschrijving en τ de tijdconstante.
Let op het belang van het verwerven van beelden zonder verzadigd pixels dat de fluorescentie herstel zou kunnen veranderen en daarmee de mobiliteit eiwit analyse. Het iis ook essentieel om altijd het fluorescentiesignaal in de gebleekte ROI normaliseren de totale fluorescentie van dezelfde cel om fluorescentie-intensiteit variaties door bleken overwegen tijdens beeldacquisitie of kleine veranderingen in de focus vlak.
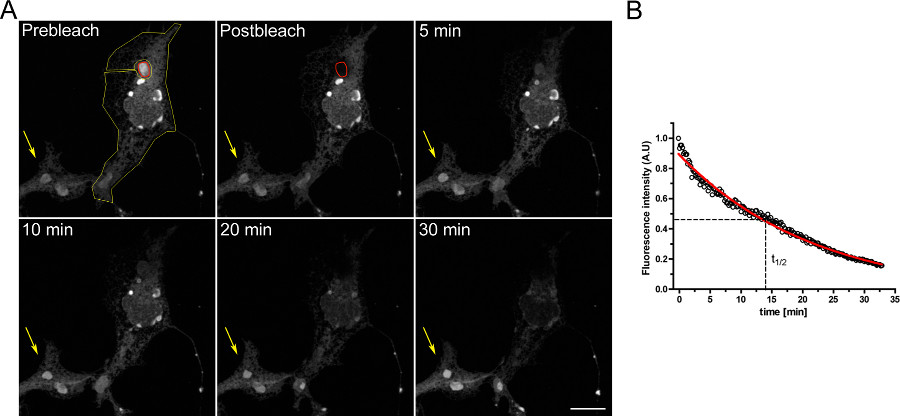
Een voorbeeld van een FLIP experiment om de continuïteit tussen intracellulaire compartimenten studie is weergegeven in figuur 3. OSERs fysiek verbonden met de rest van de ER zoals blijkt uit de geleidelijke lediging van de ER terwijl de OSER domein continu wordt gebleekt.
Voor een goede analyse moet de verwerving van verzadigde pixels worden vermeden (zie boven), bovendien moet de verwerving parameters worden ingesteld met laser krachten zo laag mogelijk om te voorkomen fotobleken door beeldacquisitie. Om deze reden wordt sterk aanbevolen om een beeld ongebleekte cel in hetzelfde veld dat wordt gebruikt om het fluorescentiesignaal van de gebleekte c normaliserenell.
Alle experimenten worden uitgevoerd in aanwezigheid van cycloheximide, een vertaling remmer, om een verhoging van de ER fluorescentiesignaal (en dus de totale fluorescentie) voorkomen door eiwit biosynthese.
Transmissie elektronenmicroscopie aangetoond dat de fluorescente aggregaten waargenomen in gekweekte cellen getransfecteerd met EGFP-d ER vertegenwoordigen stukken soepel afgevlakt ER cisternae die zich ruimtelijk georganiseerd in goed gedefinieerde 3d geometrieën ingedeeld op basis van hun patronen: lineair of gebogen stapels (vaak verband met de nucleaire envelop, niet getoond) (Figuren 4A en B) die continue en gebieden van sinusvormige ER zijn (figuur 4A), de membranen in sommige gebieden zijn georganiseerd in roosters met een vierkant of hexagonale symmetrie (kristalloïd ER niet getoonde ). Aangrenzende cisternae worden gescheiden door een dunne laag van licht elektronen-dichte cytoplasma ongeveer 11 nm dik dat continu met het cytoplasma rond de aggregaten.

Figuur 1. Stroomdiagram van de experimentele procedure. De gekweekte cellen worden eerst getransfecteerd met jetPEI (zie Protocol) om overexpressie van het fluorescerende eiwit van belang. Na 24 uur zijn de live-getransfecteerde cellen gevisualiseerd en FRAP en FLIP experimenten worden uitgevoerd met behulp van een confocale microscoop uitgerust met een gecontroleerde temperatuur CO 2 incubator, en de opgenomen beelden worden geëxporteerd en met behulp van geschikte software (bijv. ImageJ) geanalyseerd. Voor de ultrastructurele analyse worden de getransfecteerde cellen gefixeerd, gepelleteerd en ingebed in EPON epoxyhars blokken. Ultradunne secties worden verkregen met behulp van een diamant mes, verzameld op cOpper grids, en waargenomen onder een transmissie-elektronenmicroscoop. Klik hier om een grotere afbeelding te bekijken .

Figuur 2. FRAP experiment met COS-7-cellen die transiënt getransfecteerd met EGFP d-ER. A) Twee OSER structuren (rood ROI) werden gebleekt en fluorescentie herstel werd opgetekend in de tijd. Duidelijke fluorescentie herstel kan worden gedetecteerd 1 min na-bleken, en het signaal verder toeneemt 4 min later (schaal bar 10 micrometer) B):. Kwantitatieve analyse van de FRAP experiment toont het herstel half-time en de mobiele fractie van de d EGFP ER-eiwit. Click hier om een grotere afbeelding te bekijken

Figuur 3. FLIP experiment met COS-7-cellen die transiënt getransfecteerd met d EGFP-ER. A) De continue bleken van een OSER (aangegeven door de rode ROI) veroorzaakt een geleidelijke afname in fluorescentie in de rest van de ER en andere OSER structuren binnen dezelfde cel (aangegeven door de gele ROI). De gele pijl geeft een deel van een ongebleekte cel waarin het fluorescentiesignaal is constant in de tijd. (Schaal bar 10 micrometer). B) Kwantitatieve analyse van de FLIP experiment. Klik hier om een grotere afbeelding te bekijken
Figuur 4. Na fixatie en inbedding, werden cellen die hoge niveaus van d EGFP-ER waarin OSER structuren zouden worden gedetecteerd door middel van fluorescentie optische microscopie waargenomen door een transmissie-elektronenmicroscoop. A) lage vergroting van een gedeelte van het cytoplasma van een cel die een OSER bestaan uit gestapelde cisternae en golvende sinusoïdale membranen. Mitochondria (M) kan gezien worden geclusterd rond de OSER structuren, terwijl ribosomen versieren alleen de membranen van de buitenste cisternae (pijlpunten en inzet). De 11 nm dikke elektron-dichte ruimte tussen de membranen is continu met het cytoplasma (pijl en inzet) (L = lysosomen / (auto-) phagosomes) (schaal bar 1,5 micrometer; inzet 0,25 pm). B) Een OSER kan worden gevormd door lamellaire ER: dwz stapels afgeplatte ER cisternae dat continu of fragme kan zijnnted op hun verschijning in dunne secties. Blaasjes ontluikende vanaf de buitenste cisternae van de stapel af en toe kan worden waargenomen (asterisk) (PM, plasmamembraan) (Schaal bar 150 nm).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
De protocollen en beeldvormingsbenaderingen beschreven in dit document zijn gebruikt om de verdeling en mobiliteit van getransfecteerde TA fluorescente eiwitten in het ER van levende cellen resident onderzoeken. We hebben ook onderzocht het effect van overexpressie van deze eiwitten op de architecturale subcellulair compartiment via ultrastructurele analyse.
De combinatie van levende cellen confocale beeldvorming en elektronenmicroscopie staat is een zeer krachtig middel van onderzoek naar de dynamische eigenschappen van eiwitten en kan belangrijke informatie over eiwitfunctie verschaffen. De beschreven methoden zijn niet tijdrovend (meestal drie dagen van het werk), en de ontwikkeling van vele gebruiksvriendelijke software applicaties voor het verwerven en analyseren maakt fotobleking-gebaseerde live-cell imaging relatief eenvoudig.
De belangrijkste beperking van deze technieken is het gebruik van fluorescente fusieproteïnen omdatfluorescerend label opstelling kan de correcte vouwing en / of montage van het eiwit van belang. Bovendien overexpressie kan het gedrag van getransfecteerde, fluorescent gelabelde eiwitten wijzigen en derhalve niet de werkelijke eigenschappen van endogene eiwitten weerspiegelen, maar kan dit worden ondervangen door gebruik van induceerbare en stabiel getransfecteerde cellen waarin het expressieniveau kan worden precies gemoduleerd tot niveaus vergelijkbaar met die van het endogene eiwit 1,7 te verkrijgen. De neiging van kaderprogramma oligomerise is goed gedocumenteerd en kunnen gedrag (bijvoorbeeld kinetiek, ongewenste eiwit-eiwit interacties en vorming van aggregaten) van chimère eiwitten niet wezenlijk wijzigen. Het gebruik van geoptimaliseerde monomere fluorescerende eiwitten moeten derhalve 17.
Een ander kritisch aspect van dynamische weergavestudies via fluorescentie en fotobleken de tijd nodig om de fluorescentie bleken efficiënt en meet fluorescentie rehersteltools (en dus mobiliteit eiwit) bepaald, die ook afhangt van het gebied van de ROI en lokale celdikte. Als een bepaalde GFP-gelabelde eiwit een hoge diffusiecoëfficiënt kan diffusie tijdens het bleken en dus interfereren met terugwinning tijdmetingen. Om snel en efficiënt bleken te verkrijgen, is het sterk aanbevolen dat een "zoom in"-functie (indien beschikbaar) en meer dan een laser lijn worden gebruikt. Hoewel het gebruik van een snelle scan module (een resonantie scanner) kan sterk verbeteren van de snelheid van de beeldvorming tijdens de herstelfase van een experiment, in onze handen ook reduceert bleken efficiëntie. Echter kunnen alternatieve scansystemen (zoals een draaiende schijf met een speciaal photobleaching apparaat) en krachtigere lasers zowel bleken efficiënt en verkrijging snelheid.
De meeste fluorescerende eiwitten gebruikt in FRAP en FLIP experimenten tonen een zekere mate van omkeerbare photobleaching en bliNKing die moeten worden overwogen bij het uitvoeren van kwantitatieve analyses. De schommelingen tussen fluorescerende en donkere toestanden optreden in de tweede tot minuut tijdschaal. Voor EGFP, is aangetoond dat tijdens het bleken experimenten, zou fluorescentie variaties betrekken minder dan 10% van de moleculen, dus in het huidige protocol dit fenomeen is te verwaarlozen. Als alle omstandigheden constant worden gehouden, zal dit een constante voorspanning introduceren in de resultaten. Als andere fluorescerende eiwitten worden gebruikt, waarbij de reversibele fractie significant hoger (bijvoorbeeld YFP), of op te sporen en te evalueren fotobleken reversibiliteit kan dit gebeuren door fluorescentieherstel na fotobleken in het gehele levende cel, als herstel waargenomen kan dit alleen het resultaat van fotobleken omkeerbaarheid 18 zijn.
De mogelijke toxiciteit van het licht tijdens de experimenten is een andere cruciale factor, vooral omdat fotobleking vereist sterke verlichting. Het is well bekend dat opgewekt fluoroforen vaak reageren met zuurstof vrije radicalen die kunnen invloed hebben op verschillende intracellulaire processen en zelfs cellevensvatbaarheid 19 produceren, en dus moet een evenwicht tussen efficiënte bleken en minimale fototoxiciteit stellen, bovendien moet cellevensvatbaarheid altijd gecontroleerd na live cell imaging experimenten. Gezien de korte opnameduur, hebben we het genotoxische effect van licht met korte golflengte (405 nm) niet overwegen in de in dit document beschreven voorbeeld, maar als een langere experiment nodig is, een 405 nm laser lijn moet niet worden gebruikt.
We kozen voor een correlatieve benadering transmissie elektronenmicroscopie te gebruiken vanwege de heterogeniteit van OSER architectuur en het feit dat we wilden zo veel cellen (en structuren) waarnemen mogelijk. De diversiteit van de fijne structuur van eiwitcomplexen in cellen kan een belangrijk kenmerk van verschillende ziekten en we waren geïnteresseerd in het verkrijgen van een breed scala aan samples, terwijl een correlatieve benadering maakt de observatie van minder gebeurtenissen in dezelfde periode tijd. Er moet echter correlatieve licht-elektronenmicroscopie (CLEM) de eerste keuze bij het onderzoeken gebeurtenissen in structuren die niet gemakkelijk kunnen worden geïdentificeerd (zoals minder prominent ER subdomeinen) of in een beperkt aantal cellen (zoals micro-geïnjecteerde cellen). Vermeldenswaard is dat onze experimenten werden gekenmerkt door een hoge transfectie efficiëntie (ten minste 30% van de cellen getransfecteerd), anders wordt de mogelijkheid van het waarnemen OSER structuren noncorrelatively vrij beperkt.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben niets te onthullen.
Acknowledgments
De auteurs zijn dankbaar Fondazione Filarete voor zijn hulp en steun bij de publicatie van dit artikel. We willen ook graag Centro Europeo di Nanomedicina bedanken voor het gebruik van de Tecnai G2 transmissie-elektronenmicroscoop.
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) | Invitrogen | 41966029 | |
| Dulbecco’s Modified Eagle Medium (DMEM) w/o phenol red | Invitrogen | 31053028 | |
| Fetal Bovine Serum (FBS) | Invitrogen | 10270106 | |
| Pen/Strep | Invitrogen | 15140-122 | |
| L-Glutamine 200 mM solution | Invitrogen | 25030-024 | |
| jetPEI | Polyplus Transfection | PP10110 | |
| OxyFluor | Oxyrase Inc. | OF-0005 | |
| Glutaraldehyde Grade I | Sigma Aldrich | G5882 | |
| Sodium Cacodylate Trihydrate | Sigma Aldrich | C0250 | |
| Osmium Tetroxide 4% solution | Electron Microscopy Science | 19150 | |
| Uranyl Acetate Dihydrate | Sigma Aldrich | 73943 | slightly radioactive |
| Propylene Oxide | Sigma-Aldrich | 82320 | |
| EPON embedding medium kit | Sigma-Aldrich | 45359-1EA-F | |
| Lead Citrate | Electron Microscopy Science | 17800 | |
| Bench top centrifuge | Eppendorf | 5415 D | |
| Spectral Confocal Microscope | Leica Microsystems | TCS SP5 | |
| CO2 Microscope Cage Incubation System | OkoLab | ||
| Ultramicrotome | Leica Microsystems | UC6 | |
| Diamond knife | Diatome | Ultra 45 ° | |
| Transmission Electron Microscope | FEI | Tecnai G2 | |
| GraphPad Prism Software | GraphPad Software, Inc | ||
| Steel culture cell chamber for 24 mm coverslip | Bioscience Tools | CSC-25 | |
| Electron Microscopy grids | Electron Microscopy Science | G300Cu |
References
- Fasana, E., et al. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 24, 1419-1430 (2010).
- Borgese, N., Francolini, M., Snapp, E. Endoplasmic reticulum architecture: structures in flux. Curr. Opin. Cell Biol. 18, 358-364 (2006).
- Ronchi, P., Colombo, S., Francolini, M., Borgese, N. Transmembrane domain-dependent partitioning of membrane proteins within the endoplasmic reticulum. J. Cell Biol. 181, 105-118 (2008).
- Lippincott-Schwartz, J., Snapp, E., Kenworthy, A. Studying protein dynamics in living cells. Nat. Rev. Mol. Cell Biol. 2, 444-456 (2001).
- Lee, M. C., Miller, E. A., Goldberg, J., Orci, L., Schekman, R. Bi-directional protein transport between the ER and. 20, 87-123 (2004).
- Snapp, E. L., et al. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 163, 257-269 (2003).
- Papiani, G., et al. Restructured endoplasmic reticulum generated by mutant amyotrophic lateral sclerosis-linked VAPB is cleared by the proteasome. J. Cell Sci. 125, 3601-3611 (2012).
- Almsherqi, Z. A., Kohlwein, S. D., Deng, Y. Cubic membranes: a legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 173, 839-844 (2006).
- Federovitch, C. M., Ron, D., Hampton, R. Y. The dynamic ER: experimental approaches and current questions. Curr. Opin. Cell Biol. 17, 409-414 (2005).
- Takei, K., Mignery, G. A., Mugnaini, E., Sudhof, T. C., De Camilli, P. Inositol 1,4,5-trisphosphate receptor causes formation of ER cisternal stacks in transfected fibroblasts and in cerebellar Purkinje cells. Neuron. 12, 327-342 (1994).
- Feldman, D., Swarm, R. L., Becker, J. Ultrastructural study of rat liver and liver neoplasms after long-term treatment with phenobarbital. Cancer Res. 41, 2151-2162 (1981).
- Sprocati, T., Ronchi, P., Raimondi, A., Francolini, M., Borgese, N. Dynamic and reversible restructuring of the endoplasmic reticulum induced by PDMP in cultured cells. J. Cell Sci. 119, 3249-3260 (2006).
- Costantini, L. M., Fossati, M., Francolini, M., Snapp, E. L. Assessing the tendency of fluorescent proteins to oligomerize under physiologic conditions. Traffic. 13, 643-649 (2012).
- Pyszniak, A. M., Welder, C. A., Takei, F. Cell surface distribution of high-avidity LFA-1 detected by soluble ICAM-1-coated microspheres. J. Immunol. 152, 5241-5249 (1994).
- Taylor, J. P., Hardy, J., Fischbeck, K. H. Toxic proteins in neurodegenerative disease. Science. 296, 1991-1995 (2002).
- Winklhofer, K. F., Tatzelt, J., Haass, C. The two faces of protein misfolding: gain- and loss-of-function in neurodegenerative diseases. EMBO J. 27, 336-349 (2008).
- Borgese, N., Gazzoni, I., Barberi, M., Colombo, S., Pedrazzini, E. Targeting of a tail-anchored protein to endoplasmic reticulum and mitochondrial outer membrane by independent but competing pathways. Mol. Biol. Cell. 12, 2482-2496 (2001).
- Chudakov, D. M., Matz, M. V., Lukyanov, S., Lukyanov, K. A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol Rev. 90, 1103-1163 (2010).
- Bancaud, A., Huet, S., Rabut, G., Ellenberg, J. Fluorescence perturbation techniques to study mobility and molecular dynamics of proteins in live cells FRAP, photoactivation, photoconversion, and FLIP. Cold Spring Harb. Protoc. 2010, (2010).
- Michida, T., et al. Role of endothelin 1 in hemorrhagic shock-induced gastric mucosal injury in rats. Gastroenterology. 106, 988-993 (1994).

{kind=link}