

Summary
אנו מתארים את גישות ההדמיה אנו משתמשים כדי לחקור את ההפצה וניידות של חלבוני ניאון transfected תושב בreticulum endoplasmic (ER) באמצעות confocal ההדמיה של תאי חיים. אנחנו גם ultrastructurally לנתח את ההשפעה של הביטוי שלהם על הארכיטקטורה של תא subcellular זה.
Abstract
השומנים והחלבונים בתאי אוקריוטים הם החליפו ברציפות בין תאי תאים, אם כי אלה לשמור על ההרכב הייחודי שלהם ופונקציות למרות תנועת interorganelle המולקולרית האינטנסיבית. הטכניקות מתוארות במאמר זה הן אמצעי רב עוצמה של לימוד חלבון וניידות שומנים בדם וסחר in vivo ו בסביבה הפיזיולוגית שלהם. התאוששות הקרינה לאחר photobleaching (FRAP) וירידה בקרינה בphotobleaching (פליפ) נמצאת בשימוש נרחב בטכניקות הדמיה לחיות תאים ללימוד סחר תאיים באמצעות מסלול Exo-endocytic,, לוקליזציה היווצרות של קומפלקסי חלבונים, והחלבונים ההמשכיות בין האברונים או subcompartments בmicrodomains שומנים בדם, אשר כולם יכולים להיות שנצפו בתנאים פיסיולוגיים ופתולוגיים. המגבלות של גישות אלו הן בעיקר כתוצאה מהשימוש בחלבוני היתוך ניאון, והחסרונות הפוטנציאליים שלהם כוללים artifactual מעל מפורשיון בתאים ואת האפשרות של הבדלים בקיפול והלוקליזציה של חלבונים מתויגים וילידים. לבסוף, כגבול של רזולוציה של מיקרוסקופיה אופטית (כ 200 ננומטר) אינו מאפשר חקירה של המבנה העדין של ER או subcompartments הספציפי שיכול לנבוע בתאים תחת לחץ (כלומר חוסר חמצן, מתן תרופה, ביטוי יתר של הטרנסממברני חלבוני תושב ER) או תחת מצבים פתולוגיים, אנו משלבים הדמיה לחיות תאים של תאי transfected תרבותיים עם ultrastructural ניתוחים הבוסס על מיקרוסקופ אלקטרונים הילוכים.
Introduction
גילוי חלבון פלואורסצנטי ירוק (GFP) וריאנטים הרפאים שלה, ופיתוח המקביל של מיקרוסקופ פלואורסצנטי של, נפתח אפיקים חדשים לחלוטין לחקירה של חלבון התנהגות בתאים. טכניקות כגון התאוששות הקרינה לאחר photobleaching (FRAP) וירידה בקרינה בphotobleaching (פליפ), שהם אפשריים בגלל היכולת הפנימית של fluorophores לכבות הקרינה שלהם תחת תאורה אינטנסיבית, מבוססות על הדמיה לחיות תאי confocal והשימוש בtransfected חלבוני היתוך ניאון 1-3. הם נמצאים בשימוש נרחב כדי להעריך לא רק את הלוקליזציה של חלבונים, אלא גם הניידות שלהם והובלת ועי, אשר יכול לחשוף רמזים חשובים הנוגעים לתפקודם 4.
התכונה הייחודית של תאי אוקריוטים היא נוכחותם של תאים תאיים שיש לי קומפוזיציות שומנים וחלבונים ספציפיות. למרות האברונים הם בידוד ע פיזיאד, הם צריכים לתקשר אחד עם השני ולשתף את המרכיבים מולקולריים על מנת לשמור על הומאוסטזיס הסלולר. מסלול ההפרשה מבטיח שהחלבונים והשומנים מסונתזים בחדר המיון להגיע ליעד הסופי הנכון שבו הם מפעילים את תפקידם. יכולים גם להיות מחוברים אברונים תאיים על ידי אתרים ליצירת קשר דינמיים המאפשרים למולקולות (שומנים) שתוחלפנה ישירות בין תאים. יתר על כן, חלבונים רבים להתאספו במתחמי heteromeric גדולים או הקשורים למיני שומנים מסוימים (רפסודות שומנים בדם / microdomains) בכדי להפוך לפונקציונלי פעיל או להיות מועבר ליעדן הסופי. כל היבטים ביולוגיים אלה משפיעים באופן משמעותי את מאפייני הקינטית של חלבונים, ולכן ניתן לחקור כראוי באמצעות הטכניקות מתוארות להלן.
הקבוצה שלנו בשימוש נרחב FRAP ולהעיף בשילוב עם מיקרוסקופ אלקטרונים כדי ללמוד את הארכיטקטורה של חדר המיון ותת השונים שלהתחומים. חדר המיון הוא התחנה הראשונה של מסלול ההפרשה וממלא תפקיד מרכזי בחלבון ושומני מיון 5. זהו אברון דינמי מאוד משנה שברורה לשקף את תפקידיה הרבים ושונים (כלומר חלבון וביוסינתזה שומנים וסחר, קיפול חלבונים, Ca 2 + אחסון ולשחרר, וחילוף חומרי xenobiotic). עם זאת, למרות שהם מורפולוגית, מרחבית, ותפקודיים שונים, תחומים אלה הם רציפים אחד עם השני, והשפע היחסי שלהם יכול להיות שונה בתאים בתנאים פיסיולוגיים ופתולוגיים. ידוע ביותר ותחומים בדרך כלל במרחב נפרד מחדר המיון הם מעטפת הגרעין, וER החלק ומחוספס, עם זאת, אנחנו ואחרים הראינו כי יש מבני ER עם ארכיטקטורה משוכללת יותר וארגון תלת ממדים בסוגי תאים שונים ו רקמות בתנאים פיסיולוגיים שיכול גם להיגרם על ידי אמצעים של גירויים מלחיצים כגון היפוקסיה, סמיםממשל, או על הביטוי של חלבוני ER-תושב הטרנסממברני 2,6 (והפניות בו).
יש לנו גם לאחרונה הפגין את נוכחותם של מבנים כאלה במודלים של תא של מחלות אנושיות 1,7. מקורם cisternae המוערמות של ER חלק, הם קבלו את השם הקולקטיבי של reticulum המאורגן החלק endoplasmic (Oser) ב2003 6, למרות שהם ידועים גם בשם karmellae, lamellae, וER גבישן על בסיס הארכיטקטורה שלהם, אשר, כמוהם גודל, יכול להשתנות. אחרי התאים transfected עם GFP התמזגו לאזור cytosolic של חלבונים (ת"א) ER-תושב (ד EGFP-ER) מעוגני זנב, הנטייה החלשה dimerizing של ה-GFP בטרנס באופן דרמטי משנה את הארגון ומבנה של חדר המיון. FRAP והניסויים FLIP הראו כי EGFP-ER ד הוא בחינם כדי לפזר בתוך OSERs, והעובדה שהוא נע מחדר מיון רשתי לOser ולהיפך </ Em> מצביע על כך שהמצרפים הם רציפים עם ER הרשתית שמסביב. ניתוח Ultrastructural אפשר לנו לתאם את נתוני הקרינה עם תיאור מפורט של אדריכלות Oser וארגון ברמת ננו: OSERs תמיד בנוי מערמות של cisternae לזווג של ER החלק אבל ייתכן שיש לי צורות שונות של ארגון המרחבי, כגון מסודר באופן קבוע סינוסי מערכים או מערבולות, או מערכים צינורי משושה "גבישן". שחלופים אלה להוביל למורפולוגיות מעוקב 8 אשר, כפי שהם כבר נמצאים בתאים בתנאים פיסיולוגיים 9 ולחצים כגון היפוקסיה 10, טיפול תרופתי 11, וסרטן 9 הבאים, ייתכן שיש פוטנציאל משמעותי כסמני ultrastructural.
לאחר הפגנה ראשונה זה באמצעות חלבוני GFP היתוך, השתמשנו ניסויי הדמיה כדי לנתח את ההתפשטות של תחומים ER בתגובה לטיפולים תרופתיים 12, assess הנטייה של חלבוני ניאון לoligomerise בתאי 13, וכדי לחקור את התפקיד של מוטציה, חלבון ת"א ALS צמוד להיווצרות של אגרגטים תאיים ממוצא ER שעשויה להיות רלוונטי ל1,8 פתוגניות שלה. היה מי שהציע כי היווצרות של אגרגטים תאיים (אשר מתרחשת במחלות ניווניות רבות 14) עשויה להיות מנגנון הגנה שנועד למנוע האינטראקציות בין חלבוני מוטציה רעילים ומרכיבי התא המקיפים 15.
להלן תיאור של שילוב של שיטות מיקרוסקופית אופטיות ואלקטרונים לחקירת מבנים בטרמינל C-שתחומים הידרופובי מוכנסים לתוך הקרום של ER, וניתוח של ההתנהגות הדינמית שלהם ואת ההשפעות של יתר הביטוי שלהם בתחום ER אדריכלות בתאים בתרבית (ראה תרשים 1 לתרשים זרימה של פרוטוקול הניסוי).
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. פלסמיד, תרבית תאים, וtransfection עם חלבונים ניאון ER
- פלסמיד המשמש במחקר זה מורכב מגרסה משופרת של ה-GFP התמזגו בC-הסופית שלה לאזור הזנב של איזופורם ER של b ציטוכרום חולדה (5) (מקוצר כאן ב (5)) באמצעות רצף מקשר. אזור הזנב מכיל את כל הרצף (Pro94-Asp134) שנשאר קרום הקשורים לאחר מחשוף טריפסין של b ילידים (5), ובכלל זה TMD 17 השאריות (תחום הטרנסממברני), מוקף ברצפי קוטב מעלה או במורד (UPS ו DPS) . מקשר מורכב מepitope myc אחריו [4 Ser (גלאי)] 3, וכל cDNA מוכנס לתוך אתרי Hind3-Xba1 של pCDNA3 וקטור ביטוי של היונקים. פרטי הבנייה של פלסמיד זה תוארו בפרסום קודם שבו הוא מכונה ה-GFP-ER 16.
- לגדול COS-7 תאים בנשר בינוני השתנה Dulbecco (DMEM) בתוספת פטה 10%סרום שור l, L-גלוטמין 2 מ"מ, 1% פניצילין / סטרפטומיצין בחממה על 37 מעלות צלזיוס ו10% CO 2.
- Transfection. צלחת 3 x 10 5 תאים על coverslip זכוכית עגול בצלחת 6 היטב ו, ביום שלאחר מכן, transfect עם מערכת JetPEI כפי שתוארו על ידי היצרן. שים לב שיחס JetPEI / DNA האופטימלי נבדק על מנת לבסס את יעילות transfection המרבית בהתאם לפלסמיד והקו סלולרי בשימוש: במקרה שלנו, JetPEI: יחס ה-DNA של 2:1 מוביל ל70-80% יעילות transfection.
2. בשידור חי הקרינה סריקת מיקרוסקופיה confocal
- Live-תא הדמיה. שים את coverslip על שתאי transfected היו זורעים לתוך תא תא תרבות פלדה ל24 coverslips מ"מ מלא DMEM w / o פנול אדומה, בתוספת 10% FBS, 2 מ"מ L-גלוטמין, 1% עט / סטרפטוקוקוס, 25 HEPES מ"מ , 50 מיקרוגרם / מיליליטר cycloheximide ו1:100 OxyFluor כדי למנוע את הדגימות מphotobleaching. Microscop confocal SP5הדואר מצויד בחממה בטמפרטורה מבוקרת CO 2 (37 ° C ו 5% CO 2) משמש לניסויי הדמיה תא חי, עם GFP-ER ד שדמיין באמצעות לייזר 488 ננומטר ומסנן 525/50 פליטה לעבור להקה.
- התאוששות הקרינה לאחר photobleaching (FRAP). צייר אזור של עניין (ROI), המקביל למבנה Oser, ולהלבין אותו באמצעות 20 חזרות ושילוב של 488 ננומטר (של לייזר 30 mW ארגון 100%, המקביל ל 5.5-6 μW במדגם) ו405 ננומטר (60% לייזר דיודה 405 30 mW, המקביל ל 11.6 μW במדגם) לייזרים אשר, בניסיון שלנו, מוביל לphotobleaching יעיל והמהיר.
- רשום את ההתאוששות של הקרינה בROIs המולבנת על ידי לקיחת מסגרת אחת כל 10 שניות ל10 דקות (זמן = פיקסל 1.61 μsec / px).
- אובדן הקרינה photobleaching (פליפ). לצייר את ההחזר על ההשקעה המתאימה למבנה Oser, ואקונומיקה כמתואר לעיל. הלבנת חוזרת על עצמו כל 30 שניות,ותמונות שלאחר הלבנת-נרשמות כל 10 שניות ל30 דקות (זמן = פיקסל 1.61 μsec / px).
- FRAP וניתוח FLIP. כל התמונות נותחו באמצעות תוכנת ImageJ (http://rsbweb.nih.gov/ij/download.html). בניסויים FRAP, התאוששות הקרינה של ההחזר על ההשקעה המולבנת נמדדת לאורך זמן ומנורמל לקרינה הכוללת של תא מולבן, שתמיד בדקה כדי להיות קבוע לאורך זמן.
- לניסויי FLIP, לצייר את ההחזר על ההשקעה מחוץ לOser מולבן ומכסה את כל התא. למדוד עוצמת הקרינה שלה לאורך זמן ולנרמל את רמות הקרינה של החזר על השקעה שצוירו על תא מולבן על מנת לתקן את כל ירידה בקרינה הנגרמת על ידי ההדמיה עצמו.
- בכל הניסויים, מופחתים אות הרקע (שנקבעה באזור שמחוץ לתאים) מעוצמות הניאון של ROIs. לבסוף, תשרטט את התוצאות באמצעות Grתוכנת aphPad פריזמה.
3. Ultrastructural ניתוח באמצעות מיקרוסקופ אלקטרוני הילוכים
לאור רעילותם של רבים מהחומרים הכימיים, כל ההליכים צריכים להתבצע לובשים מעבדה מתאימה. מעיל וכפפות מתחת למכסת מנוע קטר.
- לאחר הסרת coverslip מצלחת פטרי, לתקן את התאים שנותרו גדלו על תחתית הצלחת כmonolayer באמצעות glutaraldehyde 2% מסוננים ב0.1 חיץ cacodylate מ ', pH 7.4, ל10 דקות בטמפרטורת חדר.
- לגרד את התאים באמצעות מגרד טפלון ולהעביר אותם לתוך 1.5 צינורות מיליליטר Eppendorf. גלולה התאים באמצעות צנטריפוגה ב9,000 גרם ל10 דקות. הסר את supernatant, להוסיף מקבע טרי, ולהשאיר לילה בשעה 4 ° C.
- לשטוף את כדורים עם החיץ, אז פוסט לתקן עם תמיסה של 1% tetroxide אוסמיום במאגר cacodylate במשך שעה 1 בטמפרטורת חדר.
- לשטוף עם מים MilliQ, וחדר כתם גוש עם1% אצטט uranyl במים מזוקקים לבין 20-60 דקות.
- מייבשים את הדגימות בסדרת אתנול (70%, 80%, 90%, 100%, ו10 דקות כל אחד 100%) הולך וגדל, ולשטוף בקצרה פעמיים בפרופילן אוקסיד (15 דקות כל אחד).
- לחדור דגימות בתערובת של תחמוצת פרופילן + קומיטה (1:1) (משעה 2 ללילה).
- להטביע בשרף אפוקסי קומיטה רפא ב60 מעלות צלזיוס במשך לפחות 24 שעות.
- סעיף גושי שרף הגזוז באופן ידני באמצעות ultramicrotome LEICA UC6 מצויד בסכין יהלום 45 ° להשיג חלקים בעובי של 60-70 ננומטר. לאסוף את החלקים ב300 רשתות נחושת רשת.
- להכתים את החלקים ברשת עם תמיסה רוויה של אצטט uranyl (20 דקות) ולהוביל ציטרט (7 דקות), לשטוף ביסודיות את הרשתות על ידי טבילתם במים מזוקקים דו מסוננים, ולאפשר להם להתייבש בטמפרטורת חדר.
- הרשתות המוכתמות הם נצפו באמצעות מיקרוסקופ אלקטרונים הילוכים Tecnai G2, והתמונות שנתפסו באמצעות בוטמצלמת CCD רכוב אום בהגדלות שונות סופיות (נעות בדרך כלל בין 6,000-39,000 X).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
איור 2 מראה מחקר FRAP דוגמא לניידות חלבון. ניידות חלבון EGFP-ER ד של מודגמת על ידי התאוששות הקרינה המהירה לאחר photobleaching בOSERs המולבן. לניתוח כמוני, חצי מהזמן והחלק נייד נגזרו מנתונים שנמדדו באופן ניסיוני על ידי התאמת משוואת monoexponential הבאה:
F (t) = F הודעה + (הודעה F rec-F) (1-e-t / τ)
שבו ההודעה F היא אות הקרינה לאחר photobleaching, rec F הוא ערך התאוששות הקרינה המרבי שיגיע אחרי הלבנה, לא עת הרישום וτ הזמן הקבוע.
נא לשים לב לחשיבות של רכישת תמונות ללא פיקסלים רוויים שיכול לשנות את התאוששות הקרינה וכתוצאה מכך, ניתוח ניידות חלבון. זה אניזה גם חיוני תמיד לנרמל את אותות הקרינה בהחזר על ההשקעה המולבנת לקרינה הכוללת של אותו התא כדי לבחון וריאציות עוצמת הקרינה בשל הלבנת במהלך רכישת תמונה או שינויים קטנים במישור הפוקוס.
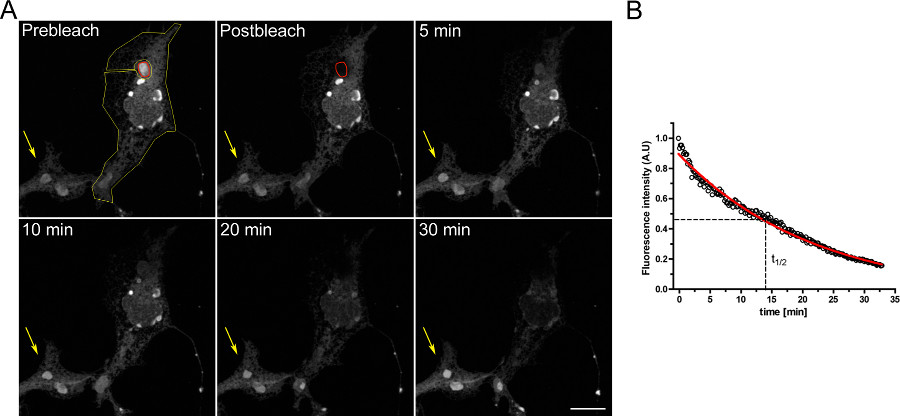
דוגמא לניסוי FLIP ללמוד את ההמשכיות בין תאים תאיים מוצגת באיור 3. OSERs מחובר באופן פיזי עם שאר חדר המיון כפי שהודגם על ידי ההתרוקנות ההדרגתית של ER כאשר תחום Oser הוא מחומצן ברציפות.
לניתוח נכון, רכישת פיקסלים רוויים יש להימנע (ראה לעיל), כמו כן, יש להגדיר את הפרמטרים לרכישה עם סמכויות לייזר נמוכות ככל האפשר על מנת למנוע photobleaching בשל רכישת תמונה. מסיבה זו מומלץ מאוד לתמונת תא מולבן באותו תחום, שישמש כדי לנרמל את אותות הקרינה של ג המולבןell.
כל הניסויים צריכים להתבצע בנוכחות cycloheximide, מעכב תרגום, על מנת למנוע כל עלייה באות הקרינה ER (וכתוצאה מכך הקרינה כוללת) בשל ביוסינתזה חלבון.
במיקרוסקופ אלקטרונים הילוכים הוכיח כי אגרגטים הניאון שנצפו בתאים בתרבית transfected עם EGFP-ER ד מייצגים כתמים חלקים ושטוח cisternae ER שמרחבית התארגן לגיאומטריות מוגדרות היטב 3D מסווג על בסיס הדפוסים שלהם: ערימות יניארי או מעוגל (לעתים קרובות הקשורים למעטפת הגרעין, לא מוצג) (איורים 4 א 'וב') שעשויה להיות רציף עם אזורים של ER סינוסי (איור 4 א); ממברנות באזורים מסוימים מאורגנות בסריגים עם סימטריה מרובעת או משושה (ER גבישן, לא מוצג ). cisternae הסמוך מופרדים על ידי שכבה דקה של אלקטרונים במקצתהציטופלסמה צפופה על 11 ננומטר העבה שהיא רציפה עם ציטופלסמה סביב המצרפים.

איור 1. תרשים זרימה של ההליך ניסיוני. התאים בתרבית הם transfected ראשון עם jetPEI (ראה פרוטוקול) על מנת יתר מבטא את חלבון היתוך הניאון של עניין. לאחר 24 שעות, תאי transfected חיים הם דמיינו וניסויים FRAP וFLIP מבוצעים באמצעות מיקרוסקופ confocal מצויד בטמפרטורת CO 2 באינקובטור מבוקר, והתמונות שתועדו מיוצאות ונותחו באמצעות תוכנה מתאימה (למשל ImageJ). לניתוח ultrastructural, תאי transfected הם קבועים, pelleted ומשובצת בגושי שרף אפוקסי EPON. סעיפי Ultrathin מתקבלים באמצעות סכין יהלום, שנאספו על גאופר רשתות, ונצפה תחת מיקרוסקופ אלקטרונים הילוכים. לחץ כאן כדי להציג דמות גדולה.

איור 2. ניסוי FRAP באמצעות COS-7 תאים transiently transfected עם EGFP-ER ד. א) שני מבני Oser (ROIs האדומה) היו מחומצן, והתאוששות הקרינה נרשמה לאורך זמן. התאוששות הקרינה ברורה יכולה להתגלות 1 דקות לאחר הלבנת, ואת אות עליות נוספות 4 דקות לאחר מכן (סרגל קנה מידה 10 מיקרומטר) B):. ניתוח כמותי של ניסוי FRAP מראה התאוששות בחצי משרה והחלק הנייד של EGFP ד -ER חלבון. Clאיכס כאן כדי להציג דמות גדולה

איור 3. ניסוי FLIP באמצעות COS-7 תאים transiently transfected עם EGFP-ER ד. א) הלבנת המתמשכת של Oser (מסומן על ידי ROI האדום) גורמת לירידה הדרגתית בקרינה בשאר חדר המיון ובמבני Oser אחרים בתוך אותו התא (מצויינים על ידי ROI הצהוב). החץ הצהוב מצביע על חלק של תא מולבן שבאות הקרינה היא קבועה לאורך זמן. ניתוח כמותי של ניסוי FLIP (בר סולם 10 מיקרומטר). ב '). לחץ כאן כדי להציג דמות גדולה
איור 4. לאחר קיבוע והטבעה, תאי מבטאי רמות גבוהות של EGFP-ER ד שבו ניתן היה לזהות מבני Oser באמצעות מיקרוסקופיה אופטית הקרינה נצפו באמצעות מיקרוסקופ אלקטרונים הילוכים. א) להציג בהגדלה נמוכה של חלק מציטופלסמה של תא המכיל Oser המורכב cisternae מוערם והקרומים סינוסי גליים. מיטוכונדריה (ז) ניתן לראות התקבצו סביב מבני Oser, בעוד שהריבוזומים לקשט רק את הקרומים של cisternae החיצוני ביותר (ראשי החץ והבלעה). חלל 11 ננומטר העבה האלקטרונים צפופים בין הקרומים הוא רציף עם ציטופלסמה (חץ והבלעה) (L = phagosomes lysosomes / (auto-)) (סרגל קנה מידה 1.5 מיקרומטר; הבלעה 0.25 מיקרומטר). B) יכול להיווצר Oser על ידי ER שבשבת: ערימות כלומר של cisternae ER השטוח שיכול להיות מתמשך או fragmented במראה שלהם בסעיפים דקים. מדי פעם ניתן להבחין בשלפוחית ניצנים מcisternae החיצונית של המחסנית (הכוכבית) (PM, קרום פלזמה) (בר סולם 150 ננומטר).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
הפרוטוקולים וגישות הדמיה מתוארות במאמר זה נעשו שימוש כדי לחקור את ההפצה וניידות של חלבוני ניאון ת"א transfected תושב בחדר המיון של תאי חיים. יש לנו גם ניתחתי את ההשפעה של ביטוי יתר של חלבונים אלה על הארכיטקטורה של תא subcellular זאת באמצעות ניתוחי ultrastructural.
השילוב של הדמיה confocal לחיות תאים במיקרוסקופ אלקטרונים מייצג הוא אמצעי חזק מאוד של חוקר את התכונות דינמיות של חלבונים, ועשוי לספק מידע חשוב הנוגעים לתפקוד חלבון. השיטות שתוארו הן לא זמן רב (בדרך כלל שלושה ימים של עבודה), ופיתוח יישומי תוכנה ידידותי למשתמש רבים לרכישת תמונה וניתוח הופך מבוסס photobleaching, הדמיה לחיות תאים פשוטים יחסית.
המגבלה העיקרית של טכניקות אלה היא השימוש בחלבוני היתוך הניאון כיתג ניאון יכול להשפיע על הקיפול ו / או הרכבה הנכון של החלבון של עניין. בנוסף, על הביטוי יכול לשנות את התנהגותם של חלבוני transfected, מתויגים פלורסנטי, ולכן עשוי שלא לשקף את התכונות אמיתיות של חלבונים אנדוגניים, אך זה ניתן להתגבר על ידי שימוש בתאים מושרה וtransfected ביציבות שברמת הביטוי יכולה להיות בדיוק מווסת להשיג רמות דומות לאלו של 1,7 חלבון אנדוגני. הנטייה של FPS לoligomerise תועדה באופן נרחב ומשמעותית יכולה לשנות את ההתנהגות (כלומר קינטיקה, אינטראקציות בין חלבונים לא רצויות והיווצרות של אגרגטים) של חלבוני chimeric. השימוש בחלבוני ניאון monomeric מותאמים צריכים להיחשב לכן 17.
עוד היבט קריטי של מחקרי הדמיה דינמיים באמצעות הקרינה וphotobleaching זה הזמן הדרוש כדי להלבין את הקרינה ביעילות ולמדוד הקרינה מחדשcovery (ולכן ניידות חלבון) דווקא, שגם תלוי באזור של ההחזר על ההשקעה ועובי תא מקומי. אם חלבון GFP-tagged ניתנו בעל מקדם גבוה דיפוזיה, דיפוזיה יכולה להתרחש במהלך הלבנת ובכך להפריע למדידות זמן החלמה. על מנת לקבל הלבנת מהירה ויעילה, מומלץ מאוד ש" זום "פונקציה (אם קיימים) וקו לייזר אחד או יותר לשמש. למרות שהשימוש במודול סריקה מהיר (כלומר סורק תהודה) יכול לשפר באופן משמעותי את המהירות של הדמיה בשלב ההתאוששות של ניסוי, בידיים שלנו זה גם מצמצם באופן משמעותי את יעילות הלבנת. עם זאת, מערכות חלופיות סריקה (כגון דיסק מסתובב מצוידים במכשיר photobleaching ייעודי), ולייזרים חזקים יותר יכולים לשפר את יעילות ומהירות הלבנת רכישה.
חלבוני הניאון ביותר בשימוש בניסויי FRAP וFLIP להראות מידה מסוימת של photobleaching וBLI הפיכיםnking שיש לקחת בחשבון בעת ביצוע ניתוחים כמותיים. התנודות בין מצבי ניאון וכהים להתרחש בשנייה לעת בהיקף דקות. לEGFP, הוכח, כי במהלך ניסויים הלבנה, וריאציות הקרינה עלולות להיות כרוכים בפחות מ -10% מהמולקולות, ובכך בפרוטוקול הנוכחי התופעה הזאת היא זניחה. אם כל התנאים נשמרים קבועים, זה יהיה להציג את הטיה קבועה בתוצאות. אם חלבוני ניאון אחרים נמצאים בשימוש, שבשבריר הפיך הוא גבוה באופן משמעותי (כלומר YFP), או כדי לזהות ולהעריך את הפיכות photobleaching, זה יכול להיעשות על ידי מדידת התאוששות הקרינה לאחר photobleaching בכל התא החי, ואם ההתאוששות הוא ציין יכול זה רק להיות התוצאה של photobleaching הפיכות 18.
פוטנציאל הרעילות של אור במהלך הניסויים היא עוד גורם קריטי, בעיקר משום שphotobleaching דורש תאורה חזקה. זה well ידוע כי fluorophores הנרגש נוטה להגיב עם חמצן כדי לייצר רדיקלים חופשיים שיכול להשפיע על תהליכים תאיים שונים ואפילו תא כדאיות 19, ולכן יש צורך להקים איזון בין הלבנת היעילה וphototoxicity המינימלי; יתר על כן, כדאיות תא תמיד צריכה להיבדק לאחר ניסויי הדמיה לחיות תאים. בהתחשב בזמן הקלטה הקצר, אנחנו לא לקחנו בחשבון את השפעת genotoxic של אור באורך גל קצר (405 ננומטר) בדוגמא שתוארה במאמר זה אבל, אם יש צורך בניסוי ארוך יותר, לא אמור לשמש קו לייזר 405 ננומטר.
אנחנו בחרנו שלא להשתמש בגישה גומל למיקרוסקופי אלקטרונים הילוכים בגלל האופי הטרוגני של אדריכלות Oser ואת העובדה שאנחנו רוצים להתבונן תאים רבים (ומבנים) ככל האפשר. הגיוון של המבנה העדין של אגרגטים חלבון בתאים עשוי להיות תכונה מרכזית של מחלות שונות והיינו מעוניינים בקבלת מגוון רחב של samples, ואילו גישה גומל מאפשרת התצפית של פחות אירועים במהלך אותה תקופת הזמן. עם זאת, במיקרוסקופ אור האלקטרון גומל (קלם) צריכה להיות הבחירה הראשונה, כאשר חוקרים אירועים במבנים שלא ניתן לזהות בקלות (כגון תחומי משנה ER פחות בולטים) או במספר מצומצם של תאים (כגון תאים מוזרקי מיקרו). ראוי לציין שהניסויים שלנו התאפיינו ברמה גבוהה של יעילות transfection (לפחות 30% מהתאים היו transfected), אחרת האפשרות של התבוננות מבני Oser noncorrelatively היא די מוגבלת.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
יש המחברים אין לחשוף.
Acknowledgments
המחברים מודים לFondazione Filarete לעזרתה ותמיכה בפרסום של מאמר זה. כמו כן, אנו רוצים להודות לCentro di Europeo Nanomedicina לשימוש במיקרוסקופ האלקטרונים הילוכים Tecnai G2.
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) | Invitrogen | 41966029 | |
| Dulbecco’s Modified Eagle Medium (DMEM) w/o phenol red | Invitrogen | 31053028 | |
| Fetal Bovine Serum (FBS) | Invitrogen | 10270106 | |
| Pen/Strep | Invitrogen | 15140-122 | |
| L-Glutamine 200 mM solution | Invitrogen | 25030-024 | |
| jetPEI | Polyplus Transfection | PP10110 | |
| OxyFluor | Oxyrase Inc. | OF-0005 | |
| Glutaraldehyde Grade I | Sigma Aldrich | G5882 | |
| Sodium Cacodylate Trihydrate | Sigma Aldrich | C0250 | |
| Osmium Tetroxide 4% solution | Electron Microscopy Science | 19150 | |
| Uranyl Acetate Dihydrate | Sigma Aldrich | 73943 | slightly radioactive |
| Propylene Oxide | Sigma-Aldrich | 82320 | |
| EPON embedding medium kit | Sigma-Aldrich | 45359-1EA-F | |
| Lead Citrate | Electron Microscopy Science | 17800 | |
| Bench top centrifuge | Eppendorf | 5415 D | |
| Spectral Confocal Microscope | Leica Microsystems | TCS SP5 | |
| CO2 Microscope Cage Incubation System | OkoLab | ||
| Ultramicrotome | Leica Microsystems | UC6 | |
| Diamond knife | Diatome | Ultra 45 ° | |
| Transmission Electron Microscope | FEI | Tecnai G2 | |
| GraphPad Prism Software | GraphPad Software, Inc | ||
| Steel culture cell chamber for 24 mm coverslip | Bioscience Tools | CSC-25 | |
| Electron Microscopy grids | Electron Microscopy Science | G300Cu |
References
- Fasana, E., et al. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 24, 1419-1430 (2010).
- Borgese, N., Francolini, M., Snapp, E. Endoplasmic reticulum architecture: structures in flux. Curr. Opin. Cell Biol. 18, 358-364 (2006).
- Ronchi, P., Colombo, S., Francolini, M., Borgese, N. Transmembrane domain-dependent partitioning of membrane proteins within the endoplasmic reticulum. J. Cell Biol. 181, 105-118 (2008).
- Lippincott-Schwartz, J., Snapp, E., Kenworthy, A. Studying protein dynamics in living cells. Nat. Rev. Mol. Cell Biol. 2, 444-456 (2001).
- Lee, M. C., Miller, E. A., Goldberg, J., Orci, L., Schekman, R. Bi-directional protein transport between the ER and. 20, 87-123 (2004).
- Snapp, E. L., et al. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 163, 257-269 (2003).
- Papiani, G., et al. Restructured endoplasmic reticulum generated by mutant amyotrophic lateral sclerosis-linked VAPB is cleared by the proteasome. J. Cell Sci. 125, 3601-3611 (2012).
- Almsherqi, Z. A., Kohlwein, S. D., Deng, Y. Cubic membranes: a legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 173, 839-844 (2006).
- Federovitch, C. M., Ron, D., Hampton, R. Y. The dynamic ER: experimental approaches and current questions. Curr. Opin. Cell Biol. 17, 409-414 (2005).
- Takei, K., Mignery, G. A., Mugnaini, E., Sudhof, T. C., De Camilli, P. Inositol 1,4,5-trisphosphate receptor causes formation of ER cisternal stacks in transfected fibroblasts and in cerebellar Purkinje cells. Neuron. 12, 327-342 (1994).
- Feldman, D., Swarm, R. L., Becker, J. Ultrastructural study of rat liver and liver neoplasms after long-term treatment with phenobarbital. Cancer Res. 41, 2151-2162 (1981).
- Sprocati, T., Ronchi, P., Raimondi, A., Francolini, M., Borgese, N. Dynamic and reversible restructuring of the endoplasmic reticulum induced by PDMP in cultured cells. J. Cell Sci. 119, 3249-3260 (2006).
- Costantini, L. M., Fossati, M., Francolini, M., Snapp, E. L. Assessing the tendency of fluorescent proteins to oligomerize under physiologic conditions. Traffic. 13, 643-649 (2012).
- Pyszniak, A. M., Welder, C. A., Takei, F. Cell surface distribution of high-avidity LFA-1 detected by soluble ICAM-1-coated microspheres. J. Immunol. 152, 5241-5249 (1994).
- Taylor, J. P., Hardy, J., Fischbeck, K. H. Toxic proteins in neurodegenerative disease. Science. 296, 1991-1995 (2002).
- Winklhofer, K. F., Tatzelt, J., Haass, C. The two faces of protein misfolding: gain- and loss-of-function in neurodegenerative diseases. EMBO J. 27, 336-349 (2008).
- Borgese, N., Gazzoni, I., Barberi, M., Colombo, S., Pedrazzini, E. Targeting of a tail-anchored protein to endoplasmic reticulum and mitochondrial outer membrane by independent but competing pathways. Mol. Biol. Cell. 12, 2482-2496 (2001).
- Chudakov, D. M., Matz, M. V., Lukyanov, S., Lukyanov, K. A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol Rev. 90, 1103-1163 (2010).
- Bancaud, A., Huet, S., Rabut, G., Ellenberg, J. Fluorescence perturbation techniques to study mobility and molecular dynamics of proteins in live cells FRAP, photoactivation, photoconversion, and FLIP. Cold Spring Harb. Protoc. 2010, (2010).
- Michida, T., et al. Role of endothelin 1 in hemorrhagic shock-induced gastric mucosal injury in rats. Gastroenterology. 106, 988-993 (1994).

{kind=link}