

Summary
Описаны подходы изображений мы используем, чтобы исследовать распределение и мобильность трансфектированных флуоресцентных белков резидентов в эндоплазматической сети (ER) с помощью конфокальной микроскопии живых клеток. Мы также ультраструктурно проанализировать влияние их экспрессии на архитектуру этого субклеточной отсека.
Abstract
Липиды и белки в клетках эукариот постоянно обмениваются клеточных отсеках, хотя они сохраняют свою особую состав и функции, несмотря на интенсивный interorganelle молекулярной трафика. Методы, описанные в этой статье, являются мощным средством изучения белков и липидов мобильность и торговлей в естественных условиях и в их физиологической среде. Восстановление флуоресценции после фотообесцвечивания (FRAP) и потери флуоресценции в фотообесцвечивания (Flip) широко используются методы проживанию ячейки изображения для изучения внутриклеточных торговлей через экзо-эндоцитотический пути, локализация образование белковых комплексов, и белка непрерывность между органелл или субкомпартментов в липидных микродоменов, все из которых могут наблюдаться при физиологических и патологических состояний. Ограничения этих подходов в основном за счет использования флуоресцентного слитых белков, и их потенциальные недостатки включают артефактом над-экспрессионов в клетках и возможность различий в складывания и локализации меченых и нативных белков. Наконец, как предел разрешения оптической микроскопии (около 200 нм) не позволяет исследование тонкой структуры ER или конкретных субкомпартментов которые могут происходят в клетках в условиях стресса (то есть гипоксия, введение лекарственного средства, по-выражение трансмембранного белки ER-резидентов) или при патологических состояниях, мы объединяем живых клеток изображений культивируемых трансфекции клеток с Ультраструктурные анализирует на основе просвечивающей электронной микроскопии.
Introduction
Открытие зеленого флуоресцентного белка (GFP) и ее спектральных вариантов, и параллельного развития флуоресцентной микроскопии, открыли совершенно новые возможности для исследования поведения белка в клетках. Такие методы, как флуоресценции восстановления после фотообесцвечивания (FRAP) и потери флуоресценции в фотообесцвечивания (Flip), которые являются возможным из-за внутренней емкости флуорофорами погасить их флуоресценции при интенсивном освещении, основаны на конфокальной микроскопии живых клеток и использование трансфецировали флуоресцентные белки слияния 1-3. Они широко используются для оценки не только локализацию белков, но и их мобильность и везикулярного транспорта, который может выявить важные подсказки, касающиеся их функции 4.
Уникальной особенностью эукариотических клеток является наличие внутриклеточных компартментах, имеющих определенные липидные и белковые композиции. Хотя органеллы физически изолированныйе изд, они должны взаимодействовать друг с другом и совместно молекулярные компоненты для поддержания клеточного гомеостаза. Секреторный путь гарантирует, что белки и липиды, синтезированные в ЭР достичь правильной конечного пункта назначения, в которых они осуществляют свои функции. Внутриклеточных органелл также может быть связано с тем, динамических контактных сайтов, которые позволяют молекулы (липиды), которые будут непосредственно обмениваются отсеков. Более того, многие белки в собранном в больших гетеромерных комплексов или связаны с конкретными видами липидных (липидного плоты / микродоменов), чтобы стать функционально активный или перевозиться к месту назначения. Все эти биологические аспекты значительно влиять на кинетические свойства белков, и поэтому может быть соответствующим образом исследованы с помощью методов, описанных ниже.
Наша группа широко используется FRAP и FLIP в сочетании с электронной микроскопии для изучения архитектуры ЭР и его различных субдомены. ER является первой станцией секреторного пути и играет ключевую роль в белков и липидов сортировки 5. Это очень динамичный органеллы которого отличается поддоменов отразить его много различных функций (то есть белков и липидов биосинтеза и торговлей людьми, белок складывающиеся, Са 2 + хранения и выпуска, и ксенобиотиков метаболизма). Однако, хотя они морфологически, пространственно и функционально различны, эти домены являются непрерывными друг с другом, и их относительное содержание может быть изменено в клетках при физиологических и патологических состояний. Самый известный и обычно пространственно разделенные домены ER являются ядерная оболочка, а гладкая и грубая ER, однако, мы и другие показали, что есть ER структуры с более сложной архитектурой и трехмерной организации в различных типах клеток и тканей в физиологических условиях, которые также могут быть вызваны с помощью стрессовые стимулы, такие как гипоксия, препаратавведения или более-выражение ER-резидентом трансмембранных белков 2,6 (и ссылки в ней).
Мы также недавно продемонстрировали наличие таких структур в клеточных моделей заболеваний человека 1,7. Возникнув от уложенных цистерн гладкого ЭР, им дали коллективное имя организованной гладкой эндоплазматической сети (Осер) в 2003 году 6, хотя они также известны как karmellae, ламелями, и кристаллоидным ER на основе их архитектуры, которая, как и их размер, может варьироваться. После того как клетки трансфицируют GFP, слитый с цитозольной области хвост-якорь (ТП) ER-резидентом белки (г EGFP-ER), слабо димеризации тенденция GFP в транс резко изменяет организацию и структуру ЭР. FRAP и FLIP эксперименты показали, что г EGFP-ER может свободно диффундировать в OSERs, и тот факт, что она движется от ретикулярной ER к Осер и наоборот </ EM> указывает, что агрегаты непрерывного с окружающей ретикулярной ER. Ультраструктурный анализ позволил нам сопоставить данные флуоресценции с подробным описанием Осер архитектуры и организации на наноуровне: Бюро входят всегда состоят из стопок в паре цистерн гладкого ЭР, но может иметь различные формы пространственной организации, такие, как регулярно устраиваются синусоидальной массивы или оборотов, или шестиугольные "кристаллоидные" трубчатые массивы. Эти перестройки приводят к кубических морфологии 8, которые, как они были найдены в клетках в физиологических условиях 9 и следующих напряжений, таких как гипоксия 10, медикаментозного лечения 11 и рак 9, могут иметь значительный потенциал в качестве ультраструктурных маркеров.
После этого первого демонстрации с использованием гибридных белков GFP, мы использовали эксперименты изображений для анализа распространения ER доменов в ответ на фармакологического лечения 12, Ассесс тенденция флуоресцентных белков в oligomerise в клетках 13, а также расследовать роль мутанта, ALS-связанного белка ТА в формировании внутриклеточных агрегатов ER происхождения, которые могут иметь отношение к его патогенности 1,8. Было высказано предположение, что формирование внутриклеточных агрегатов (что имеет место во многих нейродегенеративных заболеваний 14) может быть защитный механизм, предназначенный для предотвращения взаимодействия между токсичных мутантных белков и окружающих компонентов клетки 15.
Далее следует описание комбинации оптических и электронных методов микроскопии для исследования конструкций, чей С-концевой гидрофобные домены вставляются в мембрану ЭР, а также анализ их динамического поведения и последствий их чрезмерной экспрессии на домене ER архитектура в культивируемых клетках (см. рисунок 1 для блок-схему экспериментального протокола).
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Плазмиды, Культура клеток, и Трансфекцию с ER флуоресцентных белков
- Плазмида, которую используют в данном исследовании, состоит из расширенной версией GFP слитый на его С-конца к хвостовой области ER изоформы цитохрома крысы B (5) (сокращенно здесь как B (5)) через линкерную последовательность. В регионе хвост содержит всю последовательность (Pro94-Asp134), которая остается мембраны связаны после трипсина расщепления родной б (5), в том числе 17-остатков TMD (трансмембранный домен), по бокам вверх и вниз по полярных последовательностей (ИБП и DPS) . Линкер состоит из Myc эпитопа с последующим [(Gly) 4 Ser] 3, и весь кДНК вставлен в сайты Hind3-Xba1 в экспрессирующий вектор млекопитающего pcDNA3. Детали конструкции этой плазмиды были описаны в предыдущей публикации, в котором он упоминается как GFP-ER 16.
- Grow COS-7 клеток в Дульбекко в модификации Дульбекко (DMEM) с добавлением 10% Фетал бычьего сывороточного, 2 мМ L-глутамина, 1% пенициллина / стрептомицина в инкубаторе при 37 ° С и 10% CO 2.
- Трансфекция. Пластина 3 х 10 5 клеток на круглом покровным стеклом в 6-луночный планшет и, на следующий день, трансфекции с системой JetPEI, как описано производителем. Следует отметить, что оптимальное соотношение JetPEI / ДНК был протестирован, чтобы установить максимальную эффективность трансфекции в зависимости от плазмиды и клеточной линии, используемой: в нашем случае JetPEI соотношение 2:01 ДНК приводит к 70-80% эффективности трансфекции.
2. Онлайн флуоресценции сканирующий конфокальный микроскопия
- Онлайн-ячейки изображения. Поместите покровное на котором трансфицированные клетки высевают в стальной камере культуре клеток в течение 24 мм покровные стекла, заполненных DMEM без фенолового красного, с добавлением 10% FBS, 2 мМ L-глутамина, 1% ручка / стрептококк, 25 мМ HEPES , 50 мкг / мл циклогексимида и 1:100 OxyFluor, чтобы предотвратить образцы из фотообесцвечивания. SP5 конфокальной Микроскопэ оснащен регулируемой температурой СО 2 инкубатора (37 ° C и 5% CO 2) используется для экспериментов изображений живых клеток, с д GFP-ER Визуализируемый помощью лазера 488 нм и фильтр с 525/50 полосовой выбросов.
- Восстановление флуоресценции после фотообесцвечивания (FRAP). Нарисуйте область интереса (ROI), что соответствует структуре Осер, и отбелить его, используя 20 итераций и комбинацию 488 нм (100% лазера 30 мВт Аргон, что соответствует 5,5-6 мкВт на образце) и 405 нм (60% от 30 мВт диод 405 лазера, что соответствует 11,6 мкВт на образце) лазеры, которые, по нашему опыту, приводит к эффективному и быстрому фотообесцвечивания.
- Запишите восстановление флуоресценции в обесцвеченных трансформирования, взяв одного кадра раз в 10 сек в течение 10 мин (время пикселей = 1,61 мкс / PX).
- Потеря флуоресценции в фотообесцвечивания (флип). Нарисовать ROI, соответствующую структуре Осер, и отбеливатель, как описано выше. Отбеливание повторяется каждые 30 сек,и пост-отбеливания изображения записываются каждые 10 сек в течение 30 мин (время пикселей = 1,61 мкс / PX).
- FRAP и анализ FLIP. Все изображения анализируются с помощью ImageJ программного обеспечения ( http://rsbweb.nih.gov/ij/download.html ). В FRAP экспериментов, восстановление флуоресценции беленой ROI измеряется во времени и нормированы на общей флуоресценции беленой клетки, которая всегда проверяется, чтобы быть постоянным во времени.
- Защелкивающейс экспериментов, нарисовать ROI за пределами отбеленной Осер и охватывающей всю ячейку. Измерить его интенсивность флуоресценции с течением времени и нормализации до уровня флуоресценции в ROI, нарисованных на небеленой клетки с целью коррекции в случае любого снижения флуоресценции, вызванной самой визуализации.
- Во всех экспериментах, вычитается фоновый сигнал (определяется в области вне клеток) из люминесцентных интенсивностей трансформирования. Наконец, участок результаты с помощью GrПрограммное обеспечение aphPad Призма.
3. Ультраструктурный Анализ методом трансмиссионной электронной микроскопии
Учитывая токсичность многих из реагентов, все процедуры должны проводиться носить соответствующую лабораторию. пальто и перчатки под вытяжкой.
- После удаления покровное от чашки Петри, исправить оставшиеся клеток, выращенных на дне чашки в виде монослоя с использованием отфильтрованного 2% глутарового альдегида в 0,1 М какодилатном буфере, рН 7,4, в течение 10 мин при комнатной температуре.
- Собрать клетки с использованием тефлоновой скребок и передавать их в 1,5 мл пробирки Эппендорфа. Гранул клеток путем центрифугирования при 9000 г в течение 10 мин. Удалить супернатант, добавить свежий фиксатор, и оставить на ночь при 4 ° С.
- Промыть гранул с буфером, а затем после исправить с помощью раствора 1% осмия в какодилатном буфере в течение 1 часа при комнатной температуре.
- Промыть MilliQ воды, и единым блоком пятно с1% уранилацетатом в дистиллированной воде для между 20-60 мин.
- Высушить образцы в увеличении этанол ряд (70%, 80%, 90%, 100% и 100% в течение 10 мин каждый), и кратко мыть дважды в окись пропилена (15 мин каждый).
- Проникнуть образцов в смеси пропиленоксида + Epon (1:1) (от 2 часов в течение ночи).
- Вставить в Epon отвержденной эпоксидной смолы при 60 ° С в течение по крайней мере 24 часов.
- Секции вручную отделанные блоки смолы с использованием ультрамикротоме LEICA UC6 оснащенный алмазного ножа 45 ° для получения разделы толщиной 60-70 нм. Соберите разделы по 300 сетки медных сетей.
- Пятно разделы по сетке с насыщенным раствором уранилацетатом (20 мин) и цитратом свинца (7 мин), тщательно промыть сетки, погружая их в би-дистиллированной фильтрованной воды, и дайте им высохнуть при комнатной температуре.
- Окрашенные сетки наблюдаются использованием Tecnai G2 просвечивающего электронного микроскопа, и образы, получены с помощью боттовскийом монтажа ПЗС-камера на различных конечных увеличениях (как правило, в пределах от 6,000-39,000 X).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Рисунок 2 показывает пример FRAP изучение подвижности белка. Подвижность г EGFP-ER белка свидетельствует быстрое восстановление флуоресценции после фотообесцвечивания в отбеленной OSERs. Для количественного анализа, половина времени и с мобильного фракция были получены из экспериментально измеренных данных путем установки следующий моноэкспоненциальной уравнение:
F (T) = F сообщению + (F REC-F сообщение) (1-е-т / τ)
где F пост сигнал флуоресценции после фотообесцвечивания, F REC является максимальным значением восстановление флуоресценции, который достигнет после отбеливания, т момент регистрации и τ постоянную времени.
Пожалуйста, обратите внимание важность получения изображений без насыщенных пикселей, которые могут изменить флуоресценции восстановления и, следовательно, анализа подвижности белка. Это яы также важно всегда нормализовать сигнал флуоресценции в обесцвеченных ROI к общему флуоресценции и той же клетке в целях рассмотрения вариантов интенсивность флуоресценции из-за обесцвечивания во время получения изображения или небольшие изменения в фокальной плоскости.
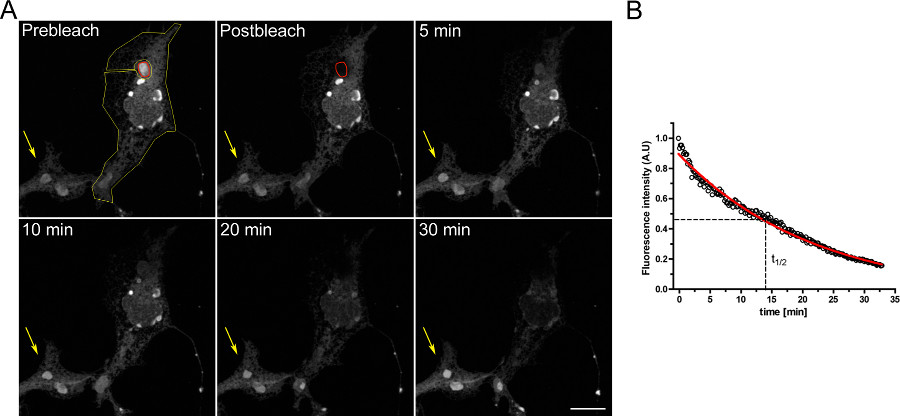
Примером FLIP эксперимент по изучению преемственность между внутриклеточными отсеками приведена на рисунке 3. Бюро входят физически соединены с остальной частью ER, как показано на прогрессивной опорожнения ER, когда область Осер непрерывно беленой.
Для правильного анализа, приобретение насыщенных пикселей следует избегать (см. выше), кроме того, параметры сбора должны быть установлены с лазерными полномочий как можно ниже в избежание фотообесцвечивание за счет получения изображений. По этой причине настоятельно рекомендуется изображения неотбеленного ячейку в той же области, который будет использоваться для нормализации сигнала флуоресценции отбеленной слокоть.
Все эксперименты должны быть выполнены в присутствии циклогексимида, ингибитора перевода, с тем чтобы избежать какого-либо увеличения сигнала флуоресценции ER (и, следовательно, общее флуоресценция) из-за биосинтеза белка.
Просвечивающей электронной микроскопии показали, что люминесцентные агрегаты, наблюдаемые в культивируемых клетках, трансфицированных г EGFP-ER представляют участки спокойная, сглаженная ER цистерн, что пространственно организованной себя в четко определенных 3D геометрии, классифицированных на основе их моделей: линейные или изогнутые стеки (часто связанный с ядерной оболочки, не показан) (фиг.4А и В), которые могут быть непрерывными с регионами синусоидальной ER (фиг. 4А), а мембраны в некоторых регионах организованы в решетках с квадратной или шестиугольной симметрией (кристаллоид ER, который не показан ). Смежные цистерны разделены тонким слоем слегка электронно-плотная цитоплазма около 11 нм, что непрерывно цитоплазме, окружающей агрегатов.

Рисунок 1. Блок-схема экспериментальной методики. Культивируемые клетки сначала трансфицировали jetPEI (см. протокол), чтобы чрезмерно выразить флуоресцентный слитый белок, представляющий интерес. После 24 часов, живые трансфицированные клетки визуализируются и FRAP и FLIP эксперименты выполняются с помощью конфокальной микроскопии, оснащенный регулируемым температурным режимом СО 2 инкубатора, и снимаемые изображения экспортируются и проанализированы с помощью соответствующего программного обеспечения (например ImageJ). Для ультраструктурном анализа, трансфицированные клетки фиксируют, осаждали и заливали в эпоксидной смолы EPON блоков. Ультратонкие срезы получены с использованием алмазного ножа, собранные от сМедь сетки, и наблюдается при просвечивающего электронного микроскопа. Нажмите здесь, чтобы увеличить рисунок .

Рисунок 2. FRAP эксперимент с использованием COS-7 клеток трансфицированы с г EGFP-ER. А) Два Осер структуры (красные Rois) отбеливались, и восстановление флуоресценции регистрировали в течение долгого времени. Ясно восстановление флуоресценции можно обнаружить 1 мин после отбеливания, и сигнал дальнейшее увеличение 4 мин позже (масштабная линейка 10 мкм) Б):. Количественный анализ FRAP эксперимента, показывая восстановления полставки и мобильная фракция г EGFP -ER белка. ClИк здесь, чтобы увеличить рисунок

Рисунок 3. FLIP эксперимент с использованием COS-7 клеток трансфицированы с г EGFP-ER. А) Непрерывный отбеливание в Осер (указано красной ROI) вызывает прогрессирующее снижение флуоресценции в остальной части ER и в других структурах Осер в пределах одной ячейки (обозначается желтой ROI). Желтая стрелка указывает на часть небеленой ячейки, в которой сигнал флуоресценции является постоянным с течением времени. (Шкала бар 10 мкм). Б) Количественный анализ эксперимента FLIP. Нажмите здесь, чтобы увеличить рисунок
Рисунок 4. После фиксации и встраивания, клетки, экспрессирующие высокие уровни г EGFP-ER, в котором Осер структуры могут быть обнаружены с помощью флуоресценции оптической микроскопии наблюдалось через просвечивающего электронного микроскопа. A) Низкий увеличение из части цитоплазмы клетки, содержащей Осер, состоящую из сложенных цистерн и волнистые синусоидальные мембран. Митохондрии (М) можно увидеть сгруппированы вокруг Осер структур, в то время как рибосомы украсить только мембраны внешней цистерн (наконечники стрел и вставка). 11 нм электронно-плотность пространства между мембранами непрерывна цитоплазме (стрелка и вставка) (L = лизосом / (авто-) фагосом) (масштаб бар 1,5 мкм, вставка 0,25 мкм). Б) Осер могут образовываться на пластинчатой ER: т.е. стеки уплощенной ER цистерн, которые могут быть непрерывными или fragmented в их появления в тонких срезов. Пузырьки начинающие от крайней цистерны стека иногда можно наблюдать (звездочка) (PM, плазма мембраны) (Шкала бар 150 нм).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Протоколы и подходы визуализации, описанные в этой статье, были использованы для исследования распределения и мобильности трансфектированных TA флуоресцентных белков резидентов в ЭР живых клеток. Мы также проанализировали влияние над экспрессия этих белков на архитектуре этой субклеточной отсека с помощью ультраструктурных анализов.
Сочетание живых клеток конфокальной микроскопии и электронной микроскопии представляет очень мощное средство исследования динамических свойств белков, и может предоставить важную информацию о функции белка. Описанные методы не отнимает много времени (как правило, три дня работы), а также развитие многих удобных программных приложений для сбора и анализа изображений делает фотообесцвечивания основе, получение изображений живых клеток-относительно проста.
Основным ограничением этих методов является использование флуоресцентный слитых белков, посколькуфлуоресцентной метки могут влиять на правильную укладку и / или сборку белка, представляющего интерес. Кроме того, сверхэкспрессия может изменить поведение трансфицированных, флуоресцентно меченных белков, и поэтому не может отражать действительные свойства эндогенных белков, однако это может быть преодолена использованием индуцируемых и стабильно трансфицированных клеток, в которых уровень экспрессии могут быть точно модулируется для получения уровней, сравнимых с теми, эндогенного 1,7 белка. Тенденция ФП в oligomerise широко документированы и могут существенно изменить поведение (т.е. кинетика, нежелательных белок-белковых взаимодействий и образование агрегатов) химерных белков. Поэтому использование оптимизированных мономерных белков флуоресцентных следует рассматривать 17.
Другим важным аспектом динамических исследований визуализации с использованием флуоресценции и фотообесцвечивания является время, необходимое для эффективного отбеливания флуоресценцию и измерить флуоресценции повторноновление (и таким образом подвижность белка) точно, который также зависит от площади и толщины ROI местного клеток. Если данный GFP с метками белок имеет высокий коэффициент диффузии, диффузии может произойти во время отбеливания и таким образом мешать измерения времени восстановления. Для того чтобы получить быструю и эффективную отбеливание, настоятельно рекомендуется, чтобы "увеличить" функции (при наличии) и более чем один лазерный луч можно использовать. Хотя использование быстрого модуля сканирования (т.е. резонансный сканер) может значительно повысить скорость визуализации на этапе восстановления эксперимента, в наших руках она также значительно снижает эффективность отбеливания. Тем не менее, альтернативные системы сканирования (например, вращающийся диск, снабженный специальной фотообесцвечивания устройства), а также более мощные лазеры могут улучшить как эффективность отбеливания и скорость сбора.
Большинство флуоресцентные белки, используемые в FRAP и FLIP экспериментов показывают некоторую степень обратимой фотообесцвечивания и BLInking, который необходимо учитывать при выполнении количественного анализа. Колебания между люминесцентных и темных состояний происходят во втором, чтобы минутах масштабе времени. За EGFP, Было показано, что в ходе экспериментов отбеливающих вариации флуоресценции может включать менее чем 10% молекул таким образом в настоящем протоколе это явление является незначительным. Если все условия остаются постоянными, это будет ввести постоянное смещение в результатах. Если другие флуоресцентные белки используются, в котором обратимый доля значительно выше (т.е. YFP), или для обнаружения и оценки фотообесцвечивания обратимость, это может быть сделано путем измерения флуоресценции восстановления после фотообесцвечивания во всей живой клетке, если наблюдается восстановление это может быть только результатом фотообесцвечивания обратимость 18.
Потенциальная токсичность света во время экспериментов является еще одним важным фактором, особенно потому, что фотообесцвечивание требует сильного освещения. Это вэйл известно, что возбужденные флуорофоров имеют тенденцию реагировать с кислородом производить свободные радикалы, которые могут повлиять на различные внутриклеточные процессы и даже клеток жизнеспособность 19, и поэтому необходимо установить баланс между эффективной отбелки и минимальной фототоксичности, кроме того, жизнеспособность клеток всегда должны быть проверены после экспериментов с изображениями живой клетки. Учитывая короткий время записи, мы не рассматривали генотоксическое действие короткой длиной волны света (405 нм) в примере, описанном в этой статье, но если больше эксперимент необходим, 405 нм лазерный луч не должен использоваться.
Мы приняли решение не использовать коррелятивную подход к просвечивающей электронной микроскопии из-за гетерогенной природы Осер архитектуры и тот факт, что мы хотели, чтобы наблюдать, как многие клетки (и структур), насколько это возможно. Разнообразие тонкой структуры белковых агрегатов в клетках может быть одной из ключевых особенностей различных заболеваний, и мы были заинтересованы в получении широкий спектр саmples, в то время как корреляционная подход позволяет наблюдать меньшим количеством событий в течение того же периода времени. Тем не менее, коррелятивный светло-электронной микроскопии (CLEM) должен быть первым выбором при исследовании событий в структурах, которые не могут быть легко идентифицированы (например, менее известных ER субдоменов) или в ограниченном количестве клеток (например, микро-инъекции клеток). Стоит отметить, что наши эксперименты характеризовались высокой степенью эффективности трансфекции (по крайней мере 30% клеток трансфицируют), иначе возможность наблюдения Осер структуры noncorrelatively весьма ограничен.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Авторы не имеют ничего раскрывать.
Acknowledgments
Авторы благодарны Fondazione Филарете за его помощь и поддержку в публикации этой статьи. Мы также хотели бы поблагодарить Centro Europeo ди Nanomedicina для использования Tecnai G2 просвечивающего электронного микроскопа.
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) | Invitrogen | 41966029 | |
| Dulbecco’s Modified Eagle Medium (DMEM) w/o phenol red | Invitrogen | 31053028 | |
| Fetal Bovine Serum (FBS) | Invitrogen | 10270106 | |
| Pen/Strep | Invitrogen | 15140-122 | |
| L-Glutamine 200 mM solution | Invitrogen | 25030-024 | |
| jetPEI | Polyplus Transfection | PP10110 | |
| OxyFluor | Oxyrase Inc. | OF-0005 | |
| Glutaraldehyde Grade I | Sigma Aldrich | G5882 | |
| Sodium Cacodylate Trihydrate | Sigma Aldrich | C0250 | |
| Osmium Tetroxide 4% solution | Electron Microscopy Science | 19150 | |
| Uranyl Acetate Dihydrate | Sigma Aldrich | 73943 | slightly radioactive |
| Propylene Oxide | Sigma-Aldrich | 82320 | |
| EPON embedding medium kit | Sigma-Aldrich | 45359-1EA-F | |
| Lead Citrate | Electron Microscopy Science | 17800 | |
| Bench top centrifuge | Eppendorf | 5415 D | |
| Spectral Confocal Microscope | Leica Microsystems | TCS SP5 | |
| CO2 Microscope Cage Incubation System | OkoLab | ||
| Ultramicrotome | Leica Microsystems | UC6 | |
| Diamond knife | Diatome | Ultra 45 ° | |
| Transmission Electron Microscope | FEI | Tecnai G2 | |
| GraphPad Prism Software | GraphPad Software, Inc | ||
| Steel culture cell chamber for 24 mm coverslip | Bioscience Tools | CSC-25 | |
| Electron Microscopy grids | Electron Microscopy Science | G300Cu |
References
- Fasana, E., et al. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 24, 1419-1430 (2010).
- Borgese, N., Francolini, M., Snapp, E. Endoplasmic reticulum architecture: structures in flux. Curr. Opin. Cell Biol. 18, 358-364 (2006).
- Ronchi, P., Colombo, S., Francolini, M., Borgese, N. Transmembrane domain-dependent partitioning of membrane proteins within the endoplasmic reticulum. J. Cell Biol. 181, 105-118 (2008).
- Lippincott-Schwartz, J., Snapp, E., Kenworthy, A. Studying protein dynamics in living cells. Nat. Rev. Mol. Cell Biol. 2, 444-456 (2001).
- Lee, M. C., Miller, E. A., Goldberg, J., Orci, L., Schekman, R. Bi-directional protein transport between the ER and. 20, 87-123 (2004).
- Snapp, E. L., et al. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 163, 257-269 (2003).
- Papiani, G., et al. Restructured endoplasmic reticulum generated by mutant amyotrophic lateral sclerosis-linked VAPB is cleared by the proteasome. J. Cell Sci. 125, 3601-3611 (2012).
- Almsherqi, Z. A., Kohlwein, S. D., Deng, Y. Cubic membranes: a legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 173, 839-844 (2006).
- Federovitch, C. M., Ron, D., Hampton, R. Y. The dynamic ER: experimental approaches and current questions. Curr. Opin. Cell Biol. 17, 409-414 (2005).
- Takei, K., Mignery, G. A., Mugnaini, E., Sudhof, T. C., De Camilli, P. Inositol 1,4,5-trisphosphate receptor causes formation of ER cisternal stacks in transfected fibroblasts and in cerebellar Purkinje cells. Neuron. 12, 327-342 (1994).
- Feldman, D., Swarm, R. L., Becker, J. Ultrastructural study of rat liver and liver neoplasms after long-term treatment with phenobarbital. Cancer Res. 41, 2151-2162 (1981).
- Sprocati, T., Ronchi, P., Raimondi, A., Francolini, M., Borgese, N. Dynamic and reversible restructuring of the endoplasmic reticulum induced by PDMP in cultured cells. J. Cell Sci. 119, 3249-3260 (2006).
- Costantini, L. M., Fossati, M., Francolini, M., Snapp, E. L. Assessing the tendency of fluorescent proteins to oligomerize under physiologic conditions. Traffic. 13, 643-649 (2012).
- Pyszniak, A. M., Welder, C. A., Takei, F. Cell surface distribution of high-avidity LFA-1 detected by soluble ICAM-1-coated microspheres. J. Immunol. 152, 5241-5249 (1994).
- Taylor, J. P., Hardy, J., Fischbeck, K. H. Toxic proteins in neurodegenerative disease. Science. 296, 1991-1995 (2002).
- Winklhofer, K. F., Tatzelt, J., Haass, C. The two faces of protein misfolding: gain- and loss-of-function in neurodegenerative diseases. EMBO J. 27, 336-349 (2008).
- Borgese, N., Gazzoni, I., Barberi, M., Colombo, S., Pedrazzini, E. Targeting of a tail-anchored protein to endoplasmic reticulum and mitochondrial outer membrane by independent but competing pathways. Mol. Biol. Cell. 12, 2482-2496 (2001).
- Chudakov, D. M., Matz, M. V., Lukyanov, S., Lukyanov, K. A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol Rev. 90, 1103-1163 (2010).
- Bancaud, A., Huet, S., Rabut, G., Ellenberg, J. Fluorescence perturbation techniques to study mobility and molecular dynamics of proteins in live cells FRAP, photoactivation, photoconversion, and FLIP. Cold Spring Harb. Protoc. 2010, (2010).
- Michida, T., et al. Role of endothelin 1 in hemorrhagic shock-induced gastric mucosal injury in rats. Gastroenterology. 106, 988-993 (1994).

{kind=link}