

Summary
Biz canlı hücre konfokal görüntüleme vasıtasıyla, endoplazmik retikulum (ER) içerisinde yerleşik transfekte floresan protein dağılımı ve hareketliliği incelemek için kullanılan görüntüleme yaklaşımı tarif eder. Biz de ultrastruktural bu subselüler bölmesinin mimarisi üzerine kendi ifade etkisini analiz.
Abstract
Bu yoğun interorganelle moleküler trafik rağmen ayırt edici bileşimin ve fonksiyonlarını muhafaza rağmen ökaryotik hücrelerde lipidler ve proteinlerin sürekli hücre bölmelerine arasında değiş tokuş edilir. Bu yazıda anlatılan teknikler in vivo ve fizyolojik ortamda protein ve lipid hareketlilik ve kaçakçılığı okuyan güçlü araçlardır. (FLIP) Photobleaching içinde (sıkı bağlamak) ve floresan kaybı photobleaching sonra floresan kurtarma yaygın exo-endositik yolu, organellere veya alt bölme arasındaki süreklilik, protein komplekslerinin oluşumu ve protein yerelleştirme yoluyla hücre içi ticaretini incelemek için canlı hücre görüntüleme teknikleri kullanılır Lipid mikro bölgeleri olarak, her biri, fizyolojik ve patolojik koşullar altında gözlemlenebilir. Bu yaklaşımların sınırlamaları nedeniyle floresan füzyon proteinlerinin kullanımı ile esas olarak ve potansiyel sakıncaları aşırı ifade suni içerirhücrelerde iyonu ve etiketli ve yerli protein katlanması ve lokalizasyon farklılıklar olasılığı. Son olarak, optik mikroskopi (yaklaşık 200 nm) çözünürlük limiti olarak stres altında hücrelerde kaynaklı olabilir ER veya belirli bir alt bölme ve ince yapısı soruşturma izin vermez (örneğin, hipoksi, ilaç uygulama, transmembran aşırı ifadesi ER ikamet proteinler) ya da patolojik koşullar altında, ultrastrüktürel ile kültür transfekte edilmiş hücrelerin canlı hücre görüntüleme kombine transmisyon elektron mikroskobu göre analiz eder.
Introduction
Yeşil floresan proteininin (GFP) ve spektral varyantları, ve floresan mikroskopi paralel gelişme keşif, hücrelerin protein davranış soruşturma için tamamen yeni yollar açtı. Çünkü yoğun bir aydınlatma altında floresan söndürmek için Flüoroforlann içsel kapasitesinin mümkündür (FLIP), ışıkla in (sıkı bağlamak) ve floresan kaybı photobleaching sonra floresan kurtarma gibi teknikler, konfokal canlı hücre görüntüleme dayalı ve kullanımı transfekte floresan füzyon proteinleri 1-3. Çıktıkları fonksiyon 4 ile ilgili önemli ipuçları ortaya çıkarabilir proteinlerin lokalizasyonu, aynı zamanda onların hareketlilik ve veziküler taşıma, sadece değerlendirmek için kullanılır.
Ökaryotik hücrelerin benzersiz özelliği spesifik lipid ve protein bileşimleri olan hücre içi bölmelerin varlığıdır. Organelleri fiziksel isolat rağmened, birbirleriyle iletişim ve hücresel homeostazı sürdürmek için moleküler bileşenleri paylaşmak gerekir. Salgı yolu ER sentezlenen proteinler ve lipidler onların işlevini uyguladıkları doğru son hedefe ulaşmasını garanti eder. Hücre içi organelleri de moleküller (lipidler) doğrudan bölmeleri arasında değiş tokuş edilmesine izin dinamik iletişim siteleri tarafından bağlanabilir. Ayrıca, birçok protein büyük heteromerik komplekslerine asamblajı ya da işlevsel olarak aktif olmaya ya da nihai hedefe taşınması amacıyla özel lipid türleri (lipid sal / mikro bölgeleri) ile bağlantılı olması. Bu biyolojik açıdan tüm büyük proteinlerin kinetik özelliklerini etkileyen ve bu nedenle, uygun bir şekilde aşağıda tarif edilen teknikler vasıtasıyla incelenebilir.
Grubumuz ER mimarisi ve farklı alt incelemek için yaygın olarak kullanılan sıkı bağlamak ve elektron mikroskobu ile birlikte FLIPetki. ER salgı yolunun ilk istasyonu ve protein ve lipid 5 sıralama önemli bir rol oynar. Bu kimin ayrı alt alan birçok farklı işlevleri (yani protein ve lipit biyosentezi ve ticareti, protein katlanması, Ca 2 + depolama ve serbest ve ksenobiyotik metabolizma) yansıtan son derece dinamik bir organelidir. Bu uzaysal olarak, morfolojik, ve fonksiyonel olarak farklı olmasına rağmen, ancak, bu etki birbirleri ile süreklidir ve bunların nispi bolluk fizyolojik ve patolojik koşullar altında hücrelerde değiştirilebilir. Iyi bilinen ve ER genellikle mekansal ayrılmış alanlar nükleer zarf ve pürüzsüz ve pürüzlü ER olmakla birlikte, biz ve diğerleri ER yapı, çeşitli hücre tiplerinde daha ayrıntılı mimari ve üç boyutlu organizasyon ile olduğunu göstermiştir ve var Ayrıca, hipoksi, ilaç olarak stresli uyaranlara vasıtasıyla tahrik edilebilir, fizyolojik koşullar altında dokularınuygulama veya ER-yerleşik transmembran proteinlerin 2,6 arasında aşırı ekspresyonu (ve buradaki referanslar).
Ayrıca son zamanlarda insan hastalıklarının 1,7 hücre modellerinde bu tür yapıların varlığını göstermiştir. Pürüzsüz ER yığılmış sisternalarında kaynaklanan onlar da kendi mimarlık temelinde karmellae, lameller ve kristalloid ER olarak bilinmesine rağmen, onlar, 2003 6 düzenlenen düz endoplazmik retikulum (OSER) kolektif adı verildi, hangi gibi onların boyutu, değişebilir. Hücreler kuyruk demirlemiş (TA) ER-yerleşik proteinler (d EGFP-ER), trans GFP zayıf dimerize eğilimi dramatik ER organizasyon ve yapısını değiştirir sitozolik bölgeye kaynaşmış GFP ile transfekte sonra. Sıkı bağlamak ve FLIP deneyler d EGFP-ER OSERs içinde yayılmak için serbest olduğunu gösterdi ve OSER için retiküler ER hareket eder ve tersi <gerçeği/ Em> agrega çevreleyen retiküler ER sürekli olduğunu göstermektedir. Ultrastrüktürel analizi bize nano düzeyde OSER mimarlık ve örgüt ayrıntılı bir açıklama ile floresan veri ilişkilendirmek için izin verdi: OSERs her zaman düzgün ER eşleştirilmiş sisternalarında yığınlarının kadar yapılır ama böyle düzenli sinüs düzenlenmiş gibi mekansal organizasyon farklı biçimlere sahip olabilir diziler veya ağırşak, veya altıgen "kristalloid" boru şeklinde diziler. Bu yeniden düzenlemeler de fizyolojik koşullar 9 ve hipoksi 10, 11, ilaç tedavisi ve kanser 9 Aşağıdaki gibi stresler altında hücrelerde bulunan edilmiş olarak, ultrastrüktürel marker olarak önemli bir potansiyele sahip olabilir, kübik morfolojileri 8 yol açar.
GFP füzyon proteinleri kullanılarak bu ilk gösteri sonra, farmakolojik tedavilere 12, asse tepki olarak ER etki proliferasyonunu analiz etmek için kullanılan deneyler görüntülemep hücreleri 13 oligomerize etmek, ve 1,8 patojenite ile ilgili olabilir ER kaynaklı hücre içi agrega oluşumunda bir mutant, ALS TA-bağlantılı protein rolünü araştırmak için floresan protein eğilimidir. Bu, (çok sayıda nörodejeneratif hastalıkların 14 oluşur) hücre içi agrega oluşumu toksik mutant proteinleri ve çevreleyen hücre bileşenleri 15 arasındaki etkileşimleri önlemek için tasarlanan bir koruyucu mekanizma olabileceğini öne sürülmüştür.
Aşağıda olan C-terminal hidrofobik alanları, ER'nin zarı içine sokulur ve dinamik davranış bir analiz ve ER alanı üzerindeki aşırı ifade yapıları etkileri araştırmak için, optik ve elektron mikroskopi yöntemlerin bir kombinasyonu bir açıklamasıdır kültürlenmiş hücrelerde mimarisi (deney protokolünün bir akış şeması için Şekil 1'e bakınız).
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1.. ER Floresan Proteinleri ile plazmid, Hücre Kültürü ve Transfection
- Bu çalışmada kullanılan fare plasmid sitokrom b'nin ER izoformunun kuyruk bölgeye C-terminalinde kaynaşmış GFP geliştirilmiş bir versiyonu olan (5) bir bağ dizisi yoluyla (b olarak (5) burada kısaltılmıştır). Kuyruk bölgesi (5), üst ve alt polar diziler tarafından eteklenen 17-artık TMB (transmembran domain), (UPS ve DPS) de dahil olmak üzere, doğal B tripsin bölünmesinden sonra ilişkili membran kalan bütün dizisi (Pro94-Asp134) içerir . Bağlayıcı [(Gly) 4, Ser] 3 ve ardından myc epitopu oluşur ve tüm cDNA, memeli ekspresyon vektörü pCDNA3 ve Hind3-Xba1 sitelerine sokulur. Bu plazmidin yapımı ayrıntılar GFP-ER 16 olarak adlandırılır edildiği önceki bir yayında tarif edilmiştir.
- ,% 10 beyaz peynir ile takviye edilmiş Dulbecco Modified Eagle Medium (DMEM) içinde COS-7 hücrelerinin büyütünl sığır serumu, 2 mM L-glutamin, 37 ° C'de bir inkübatör içinde% 1 penisilin / streptomisin ve% 10 CO2.
- Transfeksiyon. Üretici tarafından tarif edildiği gibi plaka 3 x 10 5, 6 oyuklu bir plaka içerisinde yuvarlak cam lamel üzerinde hücreleri ve, bir sonraki gün, JetPEI sistemi ile transfekte. Optimal JetPEI / DNA oranı, kullanılan plazmid ve hücre serisine bağlı olarak en fazla transfeksiyon verimi kurmak amacıyla test edilmiştir Not: bizim durumda, bir JetPEI: 2:1 DNA oranı,% 70-80 transfeksiyon verimliliği sağlanır.
2. Canlı Floresan Tarama Eşodaklı Mikroskopi
- Canlı hücre görüntüleme. Transfekte hücreler, w /% 10 FBS, 2 mM L-glutamin,% 1 pen / strep, 25 mM HEPES ile desteklenmiş fenol kırmızısı, o DMEM dolu 24 mm kapak için bir çelik hücre kültür odası içine ekildi üzerinde lamel koyun , ışıkla ağartma gelen örnekleri önlemek için 50 ug / ml sikloheksimidin ve 1:100 OxyFluor. Bir SP5 konfokal mikroskopd GFP-ER 488 nm lazer ve bir 525/50 bant geçiş emisyon filtresi kullanılarak görselleştirilmiştir olması ile ısı kontrollü bir CO2 inkübatör (37 ° C ve% 5 CO2) e sahip, canlı hücre görüntüleme deneyleri için kullanılır.
- (Sıkı bağlamak) photobleaching sonra floresan kurtarma. Bir OSER yapısına karşılık gelen, (ROI) bir bölge çizin ve (örnek olarak 5,5-6 μW tekabül eden, 30 mW argon lazer% 100) 20 tekrar ve 488 nm bir kombinasyonu kullanılarak ve 405 nm çamaşır suyu bizim deneyim, verimli ve hızlı ağartmanın yol açar, (numune 11.6 μW tekabül eden 30 mW Diyot 405 lazerin% 60) lazerler.
- 10 dakika (piksel süresi = 1.61 mikro saniye olduğu / px) için tek bir kare her 10 sn alarak ağartılmış ROI'lardaki floresan kurtarma kaydedin.
- (FLIP) Photobleaching floresan kaybı. A, bir OSER yapısına karşılık gelen YG ve yukarıda tarif edildiği gibi ağartıcı çizin. Ağartma her 30 saniye tekrar edilir,ve post-beyazlatma görüntüleri 30 dakika (piksel süresi = 1.61 mikro saniye olduğu / px) boyunca her 10 sn kaydedilir.
- Sıkı bağlamak ve FLIP analizi. Görüntülerin tüm ImageJ yazılımı (kullanarak analiz edilir http://rsbweb.nih.gov/ij/download.html ). FRAP deneylerde, ağartılmış ROI floresan iyileşme süresi ve her zaman içinde sabit olduğu kontrol edilir ağartılmış hücrenin toplam floresan, normalize boyunca ölçülür.
- FLIP deneyler için, ağartılmış OSER ve bütün bir hücresinin kaplayan bir dış ROI çizin. Zaman içinde floresan yoğunluğu ölçülür ve görüntüleme kendisinin neden floresan herhangi bir azalma için düzeltme amacıyla bir beyazlatılmamış hücre üzerinde çekilen bir ROI floresan seviyeleri için normalize.
- Deneylerin tamamında, İB'nin floresan şiddetleri dan (hücrelerin dışındaki bir alanda belirlenen) bir arka plan sinyali çıkarılır. Son olarak, Gr kullanarak sonuçları arsaaphPad Prism yazılımı.
3. Transmisyon Elektron Mikroskobu Yoluyla Ultrastrüktürel Analizi
Reaktiflerin birçok toksisite göz önüne alındığında, tüm prosedürleri, uygun bir laboratuarı giyen yürütülmelidir. davlumbaz altında ceket ve eldiven.
- Petri kabı lamel ayrılmasından sonra, oda sıcaklığında 10 dakika boyunca 0.1 M kakodilat tampon maddesi, pH 7.4 içinde, süzüldü% 2 glutaraldehid ile bir tek tabaka olarak çanak alt, üzerinde yetiştirilen kalan hücreleri gidermek.
- Bir Teflon kazıyıcı kullanarak hücreleri kazıyın ve 1.5 ml Eppendorf tüpleri içine aktarın. 10 dakika boyunca 9000 g'de santrifüj vasıtasıyla Pelet hücreleri. Süpernatantı taze fiksatif ekleyin ve 4 ° C'de gece bekletin
- Tamponu ile yıkayın peletleri, daha sonra oda sıcaklığında 1 saat boyunca kakodilat tampon içinde% 1 ozmiyum tetroksit çözeltisi ile post-sabitleyin.
- MilliQ su ile durulayın, ve ile blok leke tr20-60 arasında dakika için damıtılmış su içinde% 1 uranil asetat.
- Etanol serisi (% 70,% 80,% 90,% 100 ve 10 dakika her biri için% 100) artan örnekleri dihidrat ve (15 dk) propilen oksit içinde kısa süreyle iki kez yıkayın.
- (2 saat ilâ gece boyunca) propilen oksit + EPON (1:1) oluşan bir karışım içerisinde örnekleri sızmak.
- En az 24 saat boyunca 60 ° C'de tedavi Epon epoksi reçine gömme.
- Bölüm 60-70 nm kalınlığında bölümleri elde etmek için bir 45 ° bir elmas bıçak ile donatılmış bir ultramikrotom LEICA UC6 kullanarak el ile kesilmiş reçine bloklar. 300 örgü bakır ızgaralar üzerinde bölümleri toplayın.
- Uranil asetat (20 dakika) içinde doymuş bir çözeltisi ile ızgara üzerinde bölümleri leke ve sitrat (7 dakika) yol, iyice bi damıtılmış süzüldü, su içine daldırılarak ızgaraları yıkama ve bunları oda sıcaklığında kurumaya bırakın.
- Boyanan ızgaraları Tecnai G2 transmisyon elektron mikroskobu kullanılarak gözlenmiştir, ve görüntüler bir Bott kullanılarak yakalanır(genelde 6,000-39,000 X kadar) farklı nihai büyütme om monte CCD kamera.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Şekil 2, protein hareketlilik örnek FRAP çalışmayı gösterir. D EGFP-ER proteininin hareketlilik beyazlatılmış OSERs olarak ışıkla ağartma sonra hızlı bir floresan kurtarma ile gösterilir. Nicel analiz için, yarım saat ve cep kısmı, aşağıdaki denklem monoexponential uydurularak deneysel ölçümlere dayanan verilerden elde edilmiştir:
F (t) = F post + (F rec-F sonrası) (1-e-t / τ)
F sonrası photobleaching sonra floresan sinyal nerede, F rec kayıt t zaman, ağartma sonra ulaşmak ve zaman sabiti τ olan maksimum floresan kurtarma değerdir.
Floresan kurtarma değişmesine neden olur ve sonuç olarak, protein hareketlilik analizleri olabilir, doymuş piksel olmadan görüntü edinme önemini dikkat edin. Bu iher zaman odak düzleminde görüntü elde etme ya da küçük değişiklikler sırasında ağartma dolayı floresan yoğunluğu değişimleri dikkate almak amacıyla, aynı hücre toplam floresan ağartılmış ROI floresans sinyalinin normalize etmek de önemlidir s.
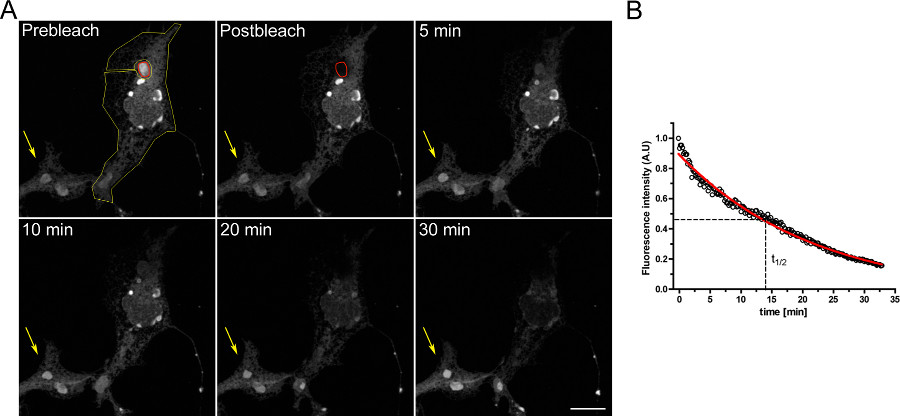
Hücre içi bölmeler arasında sürekliliği incelemek için bir deney FLIP bir örneği, Şekil 3'te gösterilmiştir. OSER alan sürekli olarak beyazlatılır ER'nin ilerleyen boşaltma gösterildiği gibi OSERs fiziksel olarak ER geri kalanı ile bağlanır.
Uygun bir analiz için, doymuş piksel edinimi (yukarıya bakınız) kaçınılmalıdır; ayrıca, satın alma parametreleri görüntü toplama nedeniyle photobleaching önlemek için mümkün olduğunca düşük lazer güçleri ile ayarlanması gerekir. Bu nedenle kuvvetli bir görüntü için ağartılmış c floresan sinyalinin normalize etmek için kullanılacak olan aynı alanda bir beyazlatılmamış hücre önerilirell.
Bütün deneyler nedeniyle protein biyosentezi için bir ER flüoresan sinyalinde bir artış (ve dolayısıyla toplam floresan) önlemek amacıyla, sikloheksimid, bir çeviri inhibitörün varlığında yapılması gerekir.
Transmisyon elektron mikroskopisi d EGFP-ER olan kültürden hücrelerinde gözlenen floresan agrega pürüzsüz yamalar temsil gösterdi ve mekansal onların desen bazında sınıflandırılarak iyi tanımlanmış 3D geometriler kendilerini organize ER sisternaları basık: genellikle doğrusal veya kavisli yığınları ( sinüzoidal ER bölgeleri ile sürekli olabilir çekirdek zarfına, gösterilmemiş olan) (Şekil 4A ve B) (Şekil 4A) ile bağlantılı, bazı bölgelerde zarlar bir kare ya da altıgen simetri (kristaloid ER ile örgüleri halinde düzenlenir, gösterilmemiştir .) Bitişik sisternaları ince bir tabaka ile ayrılır hafif elektronyoğun sitoplazmaları kalınlığında yaklaşık 11 nm bu agrega çevreleyen sitoplazma ile süreklidir.

Şekil 1. Deney prosedürünün akış şeması. Kültürlenmiş hücreler önce, ilgi konusu floresan füzyon proteinini aşırı ifade etmek için jetPEI (Protokolü bakınız) ile transfekte edilir. 24 saat sonra, transfekte edilen hücreler canlı görselleştirilebilir ve sıkı bağlamak ve FLIP deneyler, sıcaklığı kontrol edilen bir CO2 inkübatöründe ile donatılmış bir konfokal mikroskop kullanılarak gerçekleştirilir ve kaydedilen görüntüler dışa ve uygun yazılımı (örneğin, ImageJ) kullanılarak analiz edilmiştir. Ultrastrüktürel analizi için, transfekte edilmiş hücreler, sabit topak ve EPON epoksi reçine blok gömülür. İnce kesitler c toplanan, bir elmas bıçak kullanılarak elde ediliropper ızgaraları ve transmisyon elektron mikroskobu altında gözlendi. büyük rakam görmek için buraya tıklayın .

Şekil 2. FRAP deney geçici d EGFP-ER ile transfekte edilmiş COS-7 hücreleri kullanılarak. A) İki OSER yapılar (kırmızı ROI) ağartılmış edildi ve floresan kurtarma zamanla kaydedildi. Temizle floresan kurtarma 1 dk sonrası ağartma tespit edilebilir ve sinyal daha da artar 4 dakika sonra (ölçek çubuğu 10 mikron) B):. Kurtarma yarım saatini gösteren sıkı bağlamak deney kantitatif analiz ve d EGFP'nin mobil fraksiyonu -ER proteini. Clbüyük rakam görmek için buraya ick

Şekil 3,. Geçici d EGFP-ER ile transfekte edilmiş COS-7 hücreleri kullanılarak FLIP deney. A) kırmızı ROI ile gösterilen bir OSER () 'in sürekli ağartma sarı ROI ile gösterilen ER'nin geri kalanında ve aynı hücre içindeki diğer OSER yapılarda floresan ilerleyen bir azalmaya () neden olur. Sarı ok floresans sinyali zaman içinde sabit olduğu bir ağartılmamış hücrenin bir kısmını gösterir. (Ölçek çubuğu 10 mikron). B) FLIP deney kantitatif analizi. büyük rakam görmek için buraya tıklayın
Şekil 4. Sabitleme ve gömülen, OSER yapılar floresan optik mikroskopi vasıtasıyla tespit edilebilir ki d EGFP-ER yüksek seviyelerde ifade eden hücreler, bir transmisyon elektron mikroskobu ile tespit edilmiştir. Yığılmış sisternalarında ve sinüs dalgalı membran içeren bir OSER ihtiva eden bir hücrenin sitoplazması içinde bir bölümü) düşük büyütme görünüşüdür. Ribozomlar dış sisternalarında (ok başları ve ek) sadece zarlarının dekore ise mitokondri (M), OSER yapıların çevresinde kümelenmiş görülebilir. Zarlar arasında 11 nm kalınlığında elektron-yoğun alan sitoplazması (ok ve ek) (L = lizozomlar / (auto-) phagosomes) ile sürekli (ölçek çubuğu 1.5 mikron; içerlek 0.25 mikron). B) Bir OSER oluşturulabilir Sürekli veya bp parçası olabilir düzleştirilmiş ER sisternalarında yani yığınları: Yapraklı ER tarafındanİnce kesitlerde kendi görünümünü nted. Yığının dış sisternalarında gelen tomurcuklanan veziküller bazen (yıldız) (PM, plazma zarı) (Ölçek çubuğu 150 nm) görülebilir.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Bu yazıda anlatılan protokoller ve görüntüleme yaklaşımlar canlı hücrelerin ER ikamet transfekte TA floresan proteinlerinin dağılımını ve hareketliliği araştırmak için kullanılmıştır. Ayrıca mikroskobik analizler vasıtasıyla bu hücre içi bölmesinin mimarisi bu proteinlerin aşırı ifadesinin etkisini analiz ettik.
Canlı hücre konfokal görüntüleme ve elektron mikroskobu birleşimi proteinlerin dinamik özelliklerini araştıran bir çok güçlü bir araç olduğunu temsil eder, ve protein fonksiyonu ile ilgili önemli bilgiler sağlayabilir. Anlatılan yöntemlerin zaman (tipik olarak işin üç gün) tüketen değil, ve görüntü toplama ve analiz için birçok kullanıcı dostu yazılım uygulamalarının geliştirilmesi ışıkla-tabanlı, canlı hücre görüntüleme nispeten basit yapar.
Bu tekniklerin ana sınırlama floresan füzyon proteinlerinin kullanılmasıdır, çünküfloresan etiket, ilgi konusu proteinin doğru katlanmasını ve / veya montaj etkileyebilir. Buna ek olarak, fazla sentezlenme transfekte edilmiş olarak, flüoresan olarak işaretlenmiş olan proteinlerin davranışını değiştirebilir ve bu nedenle endojen proteinlerin gerçek özelliklerini yansıtmıyor olabilir, ancak bu ekspresyon seviyesi hangi indüklenebilir ve kararlı biçimde transfekte edilmiş hücrelerin kullanılarak aşılabilir tam endojen protein 1,7 olanlarla karşılaştırılabilir düzeyde elde edilmesi için modüle edilmiş. Oligomerize fps eğilimi yaygın olarak belgelenmiştir ve önemli ölçüde kimerik proteinlerin davranışı (yani kinetik, istenmeyen protein-protein etkileşimleri ve agrega oluşumu) değiştirebilir. Optimize monomerik floresan proteinleri kullanımı bu nedenle 17 düşünülmelidir.
Floresan ve photobleaching kullanarak dinamik görüntüleme çalışmalarının bir diğer önemli yönü verimli floresan çamaşır suyu ve floresan yeniden ölçmek için gerekli zamanıPasa (ve bu nedenle protein hareketlilik) tam da ROI ve yerel hücre kalınlığı alanına bağlıdır edilmektedir. Belirli bir GFP-etiketli protein, yüksek difüzyon katsayısı ise, difüzyon ağartma sırasında meydana gelen ve bu nedenle iyileşme süresi ölçümleri engelleyebilir. Hızlı ve verimli bir ağartma elde etmek için, bu güçlü bir işlev "yakınlaştırma" (varsa) ve birden fazla lazer hattı kullanılması tavsiye edilir. Hızlı bir tarama modülü (yani rezonans tarayıcı) kullanımı büyük ölçüde deney kurtarma aşamasında görüntüleme hızını artırabilir rağmen, elimizde bu da oldukça beyazlatma verimini düşürür. Bununla birlikte, (örneğin, özel bir ışıkla ağartma cihazı ile donatılmış bir eğirme disk gibi) alternatif tarama sistemleri, ve daha güçlü lazerler ağartma etkinliği ve hem de satın alma hızını artırabilir.
Sıkı bağlamak ve FLIP deneylerde kullanılan floresan proteinleri çevrilebilir ağartmanın ve BLI bazı derecesini göstermektedirKantitatif analizler gerçekleştirirken yanaşma olduğu göz önünde bulundurulmalıdır. Floresan ve karanlık devletler arasındaki dalgalanmalar dakikalık zaman ölçeğine ikinci oluşur. EGFP için, bu deneyler sırasında ağartma, floresan değişimleri böylece mevcut protokol bu olay ihmal edilebilir, moleküllerin en az% 10 dahil olabilir gösterilmiştir. Tüm koşullar sabit tutulduğu, bu sonuçlar sürekli bir önyargı tanıtacak. Diğer floresan proteinleri tersinir fraksiyonu (örneğin YFP) önemli ölçüde daha yüksek, ya da ışıkla ağartma tersinirlik tespit etmek ve değerlendirmek için hangi kullanılır ise, bu tüm canlı hücrenin içinde ışıkla ağartma sonra floresan kurtarma ölçülerek yapılabilir; geri görülmektedir, bu can Sadece geri döndürülme oranı 18 ışıkla ağartma sonucu olabilir.
Deneyler sırasında ışık potansiyel toksisitesi photobleaching güçlü aydınlatma gerektirir özellikle çünkü, başka önemli faktördür. Bu well heyecanlı flüoroforlar çeşitli hücre içi süreçleri ve hatta hücre canlılığı 19. etkileyebilecek serbest radikaller üretmek için oksijen ile reaksiyona girme eğiliminde olduğu bilinmektedir ve bu nedenle verimli ağartma ve minimal fototoksisite arasında bir denge kurmak için gerekli olan, ayrıca hücre canlılığı her zaman kontrol edilmelidir Canlı hücre görüntüleme deneylerinden sonra. Kısa kayıt süresi göz önüne alındığında, biz bu yazıda anlatılan örnekte kısa dalga boylu ışık (405 nm) genotoksik etkisini dikkate vermedi ama daha uzun bir deneme gerekiyorsa, 405 nm lazer hattı kullanılmamalıdır.
Biz bu OSER mimarisi heterojen doğası ve mümkün olduğu kadar çok hücreler (ve yapıları) gözlemlemek istedim aslında transmisyon elektron mikroskobu için bağdaşık bir yaklaşım kullanmayı tercih etti. Hücrelerdeki protein agrega ince yapısının çeşitliliği, farklı hastalıkların önemli bir özellik olabilir ve biz sa geniş bir yelpazede elde ilgilenmişlerdirörnek olarak şunlar verilebilir, bağdaşık bir yaklaşım, aynı zaman süreci sırasında daha az olayların gözlem sağlar oysa. Kolayca (örneğin daha az belirgin ER alt etki alanları gibi) ya da (örneğin mikro-enjekte hücreleri gibi) sınırlı sayıda tespit edilemeyen yapıların olayları araştıran Ancak, bağlaşık ışık-elektron mikroskobu (CLEM) ilk seçenek olmalıdır. Bu başka noncorrelatively OSER yapıları gözlemleme olasılığı oldukça sınırlıdır, (hücrelerin en az% 30 transfekte edilmiştir) Deneylerimiz transfeksiyon verimliliği yüksek bir derecesi ile karakterize olduğu kayda değerdir.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarlar ifşa hiçbir şey yok.
Acknowledgments
Yazarlar bu yazının yayınlanmasından onun yardım ve destek için Fondazione Filarete minnettarız. Biz de Tecnai G2 transmisyon elektron mikroskobu kullanımı için Centro Europeo di Nanomedicina teşekkür etmek istiyorum.
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s Modified Eagle Medium (DMEM) | Invitrogen | 41966029 | |
| Dulbecco’s Modified Eagle Medium (DMEM) w/o phenol red | Invitrogen | 31053028 | |
| Fetal Bovine Serum (FBS) | Invitrogen | 10270106 | |
| Pen/Strep | Invitrogen | 15140-122 | |
| L-Glutamine 200 mM solution | Invitrogen | 25030-024 | |
| jetPEI | Polyplus Transfection | PP10110 | |
| OxyFluor | Oxyrase Inc. | OF-0005 | |
| Glutaraldehyde Grade I | Sigma Aldrich | G5882 | |
| Sodium Cacodylate Trihydrate | Sigma Aldrich | C0250 | |
| Osmium Tetroxide 4% solution | Electron Microscopy Science | 19150 | |
| Uranyl Acetate Dihydrate | Sigma Aldrich | 73943 | slightly radioactive |
| Propylene Oxide | Sigma-Aldrich | 82320 | |
| EPON embedding medium kit | Sigma-Aldrich | 45359-1EA-F | |
| Lead Citrate | Electron Microscopy Science | 17800 | |
| Bench top centrifuge | Eppendorf | 5415 D | |
| Spectral Confocal Microscope | Leica Microsystems | TCS SP5 | |
| CO2 Microscope Cage Incubation System | OkoLab | ||
| Ultramicrotome | Leica Microsystems | UC6 | |
| Diamond knife | Diatome | Ultra 45 ° | |
| Transmission Electron Microscope | FEI | Tecnai G2 | |
| GraphPad Prism Software | GraphPad Software, Inc | ||
| Steel culture cell chamber for 24 mm coverslip | Bioscience Tools | CSC-25 | |
| Electron Microscopy grids | Electron Microscopy Science | G300Cu |
References
- Fasana, E., et al. A VAPB mutant linked to amyotrophic lateral sclerosis generates a novel form of organized smooth endoplasmic reticulum. FASEB J. 24, 1419-1430 (2010).
- Borgese, N., Francolini, M., Snapp, E. Endoplasmic reticulum architecture: structures in flux. Curr. Opin. Cell Biol. 18, 358-364 (2006).
- Ronchi, P., Colombo, S., Francolini, M., Borgese, N. Transmembrane domain-dependent partitioning of membrane proteins within the endoplasmic reticulum. J. Cell Biol. 181, 105-118 (2008).
- Lippincott-Schwartz, J., Snapp, E., Kenworthy, A. Studying protein dynamics in living cells. Nat. Rev. Mol. Cell Biol. 2, 444-456 (2001).
- Lee, M. C., Miller, E. A., Goldberg, J., Orci, L., Schekman, R. Bi-directional protein transport between the ER and. 20, 87-123 (2004).
- Snapp, E. L., et al. Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol. 163, 257-269 (2003).
- Papiani, G., et al. Restructured endoplasmic reticulum generated by mutant amyotrophic lateral sclerosis-linked VAPB is cleared by the proteasome. J. Cell Sci. 125, 3601-3611 (2012).
- Almsherqi, Z. A., Kohlwein, S. D., Deng, Y. Cubic membranes: a legend beyond the Flatland* of cell membrane organization. J. Cell Biol. 173, 839-844 (2006).
- Federovitch, C. M., Ron, D., Hampton, R. Y. The dynamic ER: experimental approaches and current questions. Curr. Opin. Cell Biol. 17, 409-414 (2005).
- Takei, K., Mignery, G. A., Mugnaini, E., Sudhof, T. C., De Camilli, P. Inositol 1,4,5-trisphosphate receptor causes formation of ER cisternal stacks in transfected fibroblasts and in cerebellar Purkinje cells. Neuron. 12, 327-342 (1994).
- Feldman, D., Swarm, R. L., Becker, J. Ultrastructural study of rat liver and liver neoplasms after long-term treatment with phenobarbital. Cancer Res. 41, 2151-2162 (1981).
- Sprocati, T., Ronchi, P., Raimondi, A., Francolini, M., Borgese, N. Dynamic and reversible restructuring of the endoplasmic reticulum induced by PDMP in cultured cells. J. Cell Sci. 119, 3249-3260 (2006).
- Costantini, L. M., Fossati, M., Francolini, M., Snapp, E. L. Assessing the tendency of fluorescent proteins to oligomerize under physiologic conditions. Traffic. 13, 643-649 (2012).
- Pyszniak, A. M., Welder, C. A., Takei, F. Cell surface distribution of high-avidity LFA-1 detected by soluble ICAM-1-coated microspheres. J. Immunol. 152, 5241-5249 (1994).
- Taylor, J. P., Hardy, J., Fischbeck, K. H. Toxic proteins in neurodegenerative disease. Science. 296, 1991-1995 (2002).
- Winklhofer, K. F., Tatzelt, J., Haass, C. The two faces of protein misfolding: gain- and loss-of-function in neurodegenerative diseases. EMBO J. 27, 336-349 (2008).
- Borgese, N., Gazzoni, I., Barberi, M., Colombo, S., Pedrazzini, E. Targeting of a tail-anchored protein to endoplasmic reticulum and mitochondrial outer membrane by independent but competing pathways. Mol. Biol. Cell. 12, 2482-2496 (2001).
- Chudakov, D. M., Matz, M. V., Lukyanov, S., Lukyanov, K. A. Fluorescent proteins and their applications in imaging living cells and tissues. Physiol Rev. 90, 1103-1163 (2010).
- Bancaud, A., Huet, S., Rabut, G., Ellenberg, J. Fluorescence perturbation techniques to study mobility and molecular dynamics of proteins in live cells FRAP, photoactivation, photoconversion, and FLIP. Cold Spring Harb. Protoc. 2010, (2010).
- Michida, T., et al. Role of endothelin 1 in hemorrhagic shock-induced gastric mucosal injury in rats. Gastroenterology. 106, 988-993 (1994).

{kind=link}