

Summary
Proteínas tau não modificados e hiperfosforiladas foram utilizados em dois ensaios in vitro de agregação para revelar a cinética de agregação rápida dependente de hiperfosforila�o. Estes ensaios pavimentar o caminho para futuras telas para os compostos que podem modular a propensão de tau hyperphosphorylated para formar fibrilas que fundamentam a progressão da doença de Alzheimer.
Introduction
Doença de Alzheimer (DA) é uma de uma grande colecção de desordens neurodegenerativas conhecidos como tauopatias. O tauopathy patologia de base por excelência é os emaranhados neurofibrilares, NFTs, nos neurônios, astrócitos e microglia 1-4. A densidade NFT se correlaciona com comprometimento cognitivo 3,5 e neurônio perda 6. NFT contém proteína tau hiperfosforilada principalmente (referido como "p-tau" daqui em diante), que forma filamentos helicoidais emparelhados (linear ou APC) 7,8. Tau é uma proteína associada à microtúbulos pensado para facilitar o transporte axonal, que é essencial para a sinalização neuronal e tráfico de 9,10. Cada molécula de tau contém 2 a 3 fosfatos em cérebro normal, mas o conteúdo de fosforilo aumenta em várias dobras em pacientes taupatia 11. Várias cinases são susceptíveis de contribuir para a tau hyperphosphorylation incluindo GSK3β (glicogênio sintase-quinase 3β) e CDK5 (ciclina-dequinase pendente 5) 12,13, mas o gatilho direto para a fosforilação patológica permanece indefinida 14. Fosforilação anormal em ou perto dos motivos de ligação de microtúbulos dissocia tau do microtúbulos 15, e faz com que tau mis-localização para o compartimento somatodendrítico, onde p-tau oligomerizes em filamentos helicoidais retas ou emparelhados que pode, eventualmente, polimerizam em inclusões NFT. A relação estreita hyperphosphorylation tau, formação NFT, e neurodegeneração levou a uma hipótese prevalente de que emaranhados p-tau extrair respostas citotóxicas apoptóticos e outros, e, portanto, é a causa subjacente para tauopathy neurodegeneração 16,17. Telas de drogas e testes clínicos iniciais com base nesta premissa, foram lançados 18. No entanto, esta hipótese enfrenta desafios 19,20. Por exemplo, SantaCruz et al., Mostraram que as funções cognitivas de ratinhos transgénicos pode ser melhorada através da supressão da expressão de um mutantetau humana, embora NFTs continuou a formar a partir de moléculas de tau existentes 21. Em um modelo de Drosophila, NFT foi mostrado para sequestrar a proteína tau citosólica tóxico para proteger as células neuronais subjacentes 22,23. Claramente, o papel patogénese de NFT, se houver, serão de grande impacto no sentido do desenvolvimento de agentes terapêuticos taupatia.
Em concentrações elevadas, a proteína tau recombinante de cérebro normais ou espontaneamente, mas lentamente polimeriza numa estrutura semelhante a PHF in vitro, tal como indicado pela ligação de vários corantes β folhas preferidos fluorescentes, microscopia electrónica, e espectroscopia de dispersão de luz 24-27. Adicionando heparina ou ácido araquidônico, um ácido graxo abundante no cérebro humano, acelera drasticamente a formação de PHF em isoform- tau e indutor dependentes da concentração maneiras 28-32. Curiosamente, tau hyperphosphorylated purificada a partir de cérebros AD ou preparados por exaustiva em reações de fosforilação in vitro umggregates mais rápida e eficiente 26,33-35. Estes resultados estão em concordância excelente com os papéis patológicos de p-tau. Um sistema in vitro baseado na agregação de p-tau pode, assim, servir como uma ferramenta poderosa para o rastreio de drogas AD.
Dada a estreita associação entre a agregação tau ea neurodegeneração progressiva da AD, bem como o recente fracasso no desenvolvimento de medicamentos visando a placa de Ap, outro marcador histológico chave de AD 36-38, o interesse em descobrir medicamentos que controlam a agregação tau está subindo. De facto, vários grupos começaram já telas de droga no caudal diferente, utilizando, em reacções de agregação de tau in vitro como o ensaio primário. Uma série de compostos exibiram actividades inibitórias sobre a agregação de reversão ou a tau in vitro 39-42. No entanto, todas as telas atuais regulador agregação tau tau usar inalterado que erra o alvo patológica chave de fósforoylation, levantando uma preocupação para a especificidade e eficácia da utilização destes compostos no tratamento AD.
Um dos principais obstáculos para desenvolver ensaios de agregação para caracterização bioquímica e rastreio de drogas AD é a produção de quantidades suficientes de proteína tau hiperfosforilada patofisiologicamente relevantes. Usando o sistema de Catálise Zippers assistidas em que a isoforma 1N4R de tau e da quinase GSK-3β são co-expressos em E. coli como proteínas de fusão de fecho de leucina, temos que superar esse desafio (. Sui et al, submetido; veja a Figura 1 para os produtos finais de tau e p-tau; veja também 43 para caracterização de espectrometria de massa preliminar do p-tau). A partir de um painel de nove anticorpos específicos para diferentes locais de fosforilação de tau, sinais positivos foram observados em oito posições (dados não mostrados). A seguir, descrevemos os protocolos e instrumentações que podem diferenciar a agregação d cinéticaIFERENÇAS entre espécies de tau não modificado e p-tau. Estes ensaios foram modificados a partir de protocolos publicados que mediram o aumento da fluorescência de tioflavina T (ThT) ou tioflavina S (THS) sobre amilóides (agregados tau) vinculativo 26. No primeiro "terminal", abordagem sem corante, reacções de agregação são montados e incubadas na ausência do corante amilóide. Em diferentes pontos de tempo, uma aliquota de cada reacção é removido e misturado com um volume igual de tampão de ThT contendo a parar a agregação e permitir ThT para ligar agregados tau. A fluorescência é medida por um IAP FluoroMax-2 fluorómetro. Na segunda "com corante" ensaio de monitorização contínua, ThT ou THS está incluído nas reacções de agregação. A fluorescência pode ser medida de forma contínua ao longo de toda a experiência manualmente ou utilizando um leitor de placas múltiplas. Além disso, descreve-se um ensaio que utiliza uma concentração quase fisiológica de tau e p-tau para a agregação na mo medição contínuade. O efeito de fosforilação permanece facilmente detectável. A seguir, vamos descrever os procedimentos passo-a-passo da operação, e mostrar resultados representativos destes ensaios. Discussão de alguns dos prós e contras de cada abordagem, bem como potenciais aplicações de rastreio de drogas vai seguir.
Numa elevada concentração, a tau se agrega em estruturas amilóides como espontaneamente. No entanto, no laboratório, tau fibrillization é tipicamente acelerada por indutores tais como heparina (peso molecular médio, 6.000 g / mol) e ácido araquidónico. Exemplos aqui apresentados incluem 30? M de heparina. A formação de agregados amilóides tau é monitorizada pela fluorescência resultante da sua ligação por tioflavina T (ThT) ou tioflavina S (THS) amilóide. Após a ligação aos agregados de tau, ThT exibe um desvio para o vermelho de fluorescência (excitação: 450 nm; emissão de pico: 485 nm). THS, por outro lado, tem fraca emissão a 510 nm (excitação a 450 nm) antes da ligação de amilóide, mas este fluorescence aumenta significativamente na presença de uma proteína amilóide, tais como a tau agregada 44. Ambos os corantes funcionam bem na detecção de tau e p-tau agregação. Devido à forte pico de emissão relativamente amplo e de ThT (ver Figura 2), existe apenas 30% de redução na unidade de fluorescência a 510 nm. Por conveniência, utilizar a mesma combinação de comprimentos de onda de excitação / emissão (isto é, 450 nm / 510 nm), para controlar a agregação tau quando utilizado um ou corante.
Agregação de tau pode ser realizada na presença ou ausência do corante, dependendo do objetivo do ensaio e a disponibilidade da proteína tau. Ambos os modos de reacções são apresentados abaixo. Além disso, podemos demonstrar o funcionamento de dois instrumentos diferentes - um fluorometer single-amostra (ISA-SPEX FluoroMax-2) e um leitor multi-plate (SpectraMax M2). Os leitores devem ser capazes de se adaptar estes protocolos para atender às suas necessidades específicas e disponibilidade de instrumentos.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Preparação dos Reagentes
- Preparar tampão de agregação (20 mM Tris, pH 7,4, NaCl 100 mM, EDTA 1 mM). Armazenar à temperatura ambiente, estável por meses. Suplemento 1 mM de ditiotreitol (DTT) antes da utilização.
NOTA: um tampão à base de HEPES (HEPES 10 mM, pH 7,5, EDTA 0,1 mM, DTT 5 mM) também produz resultados semelhantes em agregação de tau. - Prepare tioflavina T ou S tioflavina solução estoque (3 mM, dissolvido em tampão de agregação), e um filtro de 0,22 um por unidade de filtro estéril. Armazenar a -20 ° C em um tubo coberto por folha de alumínio, estável durante meses.
- Preparar a solução de heparina (300 uM, dissolvidos em tampão de agregação). Armazenar a -20 ° C, estável durante meses.
- Prepare ditiotreitol (DTT) estoque (1 M, dissolvido em água). Alíquota em tubos de 1,5 ml. Armazenar a -20 ° C. Antes de ensaios de agregação, descongelar a solução a 1 M à temperatura ambiente. A partir deste 1.000x estoque, prepare uma alíquota de 100 mM estoque que trabalham com água deionizada. Deixar no geloaté que esteja pronto.
- Remover tau a partir de -80 ° C congelador. Descongele no gelo. Ajustar a concentração predeterminada de tau com o tampão de agregação. Centrifugação numa microcentrífuga a 20.800 xg durante 10 min a 4 ° C para remover grandes agregados pré-formados. Este passo de pré-fiação aumenta a consistência da medição da fluorescência de cada lote subsequente da preparação de proteína. Transferir o sobrenadante para outro tubo; deixar no gelo até que esteja pronto para montar a reação de agregação.
2. Não-dye, Ensaio Terminal
NOTA: A reacção de agregação deste ensaio é realizado na ausência do corante fluorescente. Depois de misturar todos os componentes, a reacção é deixada prosseguir a pontos de tempo pré-determinados. As aliquotas são então retiradas da reacção de agregação e misturado com ThT ou THS para a ligação antes da leitura da fluorescência amilóide. O volume inicial da reacção de agregação depende do número de pontos de tempo necessários. Esta abordagem pode require uma grande quantidade de proteína tau, mas é rápido, simples, e pode ser feito de um fluorímetro ou um leitor de placa de multi-poços (ver discussão). Abaixo está o funcionamento passo-a-passo, usando o ISA SPEX FluoroMax-2 espectrofluorómetro compacto para quantificação de fluorescência.
- Defina-se a mistura de agregação em tubos de 1,5 ml de Eppendorf como na Tabela 1. Cada coluna representa os ingredientes necessários para uma reacção de 100 ul, o que é suficiente para uma medição de ponto de momento. Ajuste a quantidade de todo o mix de agregação com base nos pontos de tempo necessário para a experiência particular. Adicionar adicional de 10% de cada componente para dar espaço para a pipetagem erro. A reacção contendo heparina típica mostrada aqui pode ser substituído por ácido araquidónico ou tampão de agregação. Adicionar DTT a 1 mM à mistura de reacção. Se toda a reacção demora mais do que um dia, complementar fresco todos os dias DTT (1 mM) para garantir um ambiente redutor.
- Inverter o tubo algumas vezes para misturar. Place cada reacção numa incubadora de água ou banho de 37 ° C. A agitação não é necessária para a agregação de tau.
- Antes de medir a fluorescência, ligue o espectrofluorómetro (lâmpada em primeiro lugar, em seguida, computador).
NOTA: A lâmpada de arco de xenon que pode ser utilizado de imediato. No entanto, para obter melhores resultados permitem que a máquina aquecer por cerca de 10 min antes da leitura de fluorescência. - Inicie o software no computador.
- Escolha o modo de visualização em tempo real em Instrument Control Center, definir comprimento de onda de excitação de 450 nm (fenda de 2 nm) e comprimento de onda de emissão de 510 nm (fenda de 5 nm). Fechar a janela Modo de exibição Tempo real para retornar ao Centro de Controle de Instrumento.
- Escolha Constante chave Análise Wavelength, prima Add >> na estrutura superior para adicionar conjuntos de comprimentos de onda. Defina os parâmetros de aquisição de erro padrão para 1 e Trials máxima a 3, em seguida, clique em Adicionar. Clique em Go! Para abrir a tela de dadosjanela.
- Na janela exibir os dados, clique em Iniciar Acq para abrir a caixa de diálogo Sample Novo. Escolha "desconhecido" para o tipo de amostra.
- Para cada mistura de agregação de 100 ul, adicionar 98 ul de tampão de agregação e 2 ul de 3 mM de tioflavina T. pipeta várias vezes para misturar.
- Transfira a totalidade da mistura a uma cuvete (FCA3, dimensão exterior, CxLxH = 12,5 mm x 12,5 milímetros x 45 mm). Coloque a cuvete no porta-amostras em compartimento de amostra a e feche a tampa. Clique em Executar para coletar os dados de fluorescência. Grave os dados.
- Retirar a cuvete e decantar a solução. Lavar o cadinho por água destilada 3 vezes. Dry, soprando ar para dentro e fora da tina.
3. Com-dye, Modo contínuo Assay num leitor de placas SpectraMax M2
NOTA: Este ensaio difere do anterior em que o corante fluorescente ou ThT THS está incluído no aggregareação ção. Isto permite a medição contínua do mesmo conjunto de reacções. Devido à utilização repetitiva de reacção, este método é melhor feita com um leitor de placas multi-poços automática (como mostrado a seguir a operação de SpectraMax M2). A fluorometer regular também funciona, mas a freqüência de medição de reações rápidas de agregação é um pouco limitado, devido à natureza manual da operação.
- Defina-se a mistura de agregação em uma placa de 96 poços (placa de 96 poços preta sólida, bem volume de 360 ul, fundo plano) como na Tabela 2. Cada coluna representa os ingredientes necessários para uma reacção de 200 ul, o que é suficiente para uma medição do tempo de ponto. Misture bem por pipetagem várias vezes. Suplemento 1 mM de DTT fresco todos os dias ao longo do decorrer de experiências.
- Incubar a placa de 96 poços a 37 ° C.
- Em cada ponto de tempo, antes da medição de fluorescência, ligar o leitor de microplacas multi-modo e ao computador. Dê tempo suficiente para o machine a se estabilizar, cerca de 10 min.
- Inicie o software no computador. Ajuste da temperatura a 37 ° C e escolher o modo de intensidade de fluorescência (FI-Top Ler), definido pelo comprimento de onda de excitação de 450 nm e comprimento de onda de emissão a 510 nm.
- Insira a placa de 96 poços na gaveta e pressione a tecla de leitura para iniciar a medição.
- Após a leitura, retire a placa e devolvê-lo de volta para o 37 ° C incubadora. Copie os dados e cole em uma planilha do Excel para análise de dados e plotagem.
4. Com-dye, Modo contínuo Ensaio sobre uma Espectrofluorímetro Compact
- Defina-se a mistura de agregação em tubos de 1,5 ml de Eppendorf como na Tabela 3. Cada coluna representa os ingredientes necessários para uma reacção de 200 ul, o que é suficiente para uma medição de ponto de momento.
- Inverter o tubo algumas vezes para misturar.
- Ligue o espectrofluorómetro e definir o software como nos passos 2.3 e 2.4.
- Transferir toda a mistura toa cuvete. Coloque a cuvete no porta-amostras em compartimento de amostra a e feche a tampa. Clique em Executar para coletar os dados de fluorescência. Grave os dados.
- Continue lendo a intervalos apropriados, clicando em Executar e gravar os dados. Se a agregação é para ser monitorizada, com uma frequência elevada (por exemplo, todos os 30 ou 60 seg), deixar a reacção na cuvete e na máquina até que a medição está concluída, ou quando não há tempo suficiente para trocar reacções ou cuvetes.
- Retirar a cuvete e decantar a solução. Lavar o cadinho por água destilada 3 vezes. Dry, soprando ar para dentro e fora da tina.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Usando tau recombinante e p-tau (Figura 1), que estabeleceu dois protocolos diferentes para comparar a cinética de agregação de tau e p-tau, aproveitando a forte emissão de fluorescência de ThT e THS por ligação a agregados de proteína amilóide, incluindo tau e p-tau (Figura 2). Com ou sem o corante fluorescente na reacção de agregação, observou-se aumento consistente de agregação de tau por hiperfosforilação (Figuras 3-5). Esta estimulação é independente de heparina (dados não mostrados). Numa reacção, tau-tau e p oligomerizar típico na taxa rápida dentro dos primeiros 30 min antes de abrandar (Figuras 3 e 5), com p-tau exibindo unidades de fluorescência mais elevados ao longo do curso do experimento significativamente. Incluindo ThT nas reacções de agregação provoca atraso significativo na velocidade de agregação (Figura 4). Ambas as isoformas plat abordadoeau 160 horas após as reacções tinha começado. THS, por outro lado, não causa desaceleração significativa da agregação (Figura 5).

Figura 1. purificada tau e tau hiperfosforilado (tau-p) usado neste estudo. As amostras foram resolvidas por 10% de um gel de SDS-PAGE e coradas por azul de Coomassie R250 (esquerda) ou sondado por um anticorpo monoclonal anti-tau (direita painel). Pista M, marcador de peso molecular; raias 1 e 3, tau fosforilada; lane2 e 4, tau hyperphosphorylated.

Figura 2. espectros de emissão para ThT (30 M) com ou sem agregados tau. Aquisição de emissão foi s enlatado de 460 nm a 600 nm (incrementos de 1 nm, a integração de 0,1 seg, 5 nm de largura de fenda), excitação a 450 nm. Agregados tau foram obtidos por 50 uM permitindo a agregação tau-se a 37 ° CO / N (ver Protocolo 2 para detalhes).
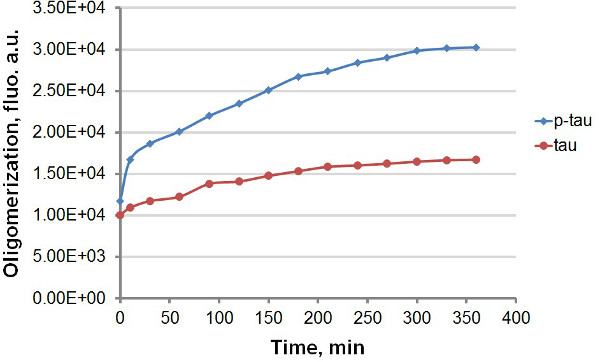
Figura 3. Curvas de agregação de tau e p-tau em ensaio terminal. Agregação de 50 uM de tau e p-tau foi completado com 30 uM de heparina como o indutor. Em diferentes momentos após o início da reacção, a 100 ul da reacção foi removido e misturado com o mesmo volume de 60 uM ThT antes das medições de fluorescência. A fluorescência foi medida a 450 nm de excitação, 510 nm de emissão. , unidades arbitrárias "au". Note-se que a escala de tempo é em minutos.
"Figura 4" src = "/ files / ftp_upload / 51537 / 51537fig4highres.jpg" />
Figura 4. As curvas de agregação para TT por si só, tau-tau e p no modo de medição contínua na presença de ThT. Cada reacção consistiu em 50 ou 0 pM ou p tau-tau, 30 uM de heparina e 30 | iM em tampão de ThT agregação . As reacções foram incubadas a 37 ° C em uma placa de 96 poços. Em diferentes pontos de tempo, a placa foi removida da incubadora e carregado para o leitor de placas para leitura da fluorescência (excitação 450 nm, emissão 510 nm). Entre as leituras, a placa foi mantida em incubadora sem agitação sob uma cobertura. A presença de ThT abrandou consideravelmente a agregação, mas, mais importante, a tau hiperfosforilada ainda exibiu uma taxa mais rápida de agregação do que a sua contraparte não modificada. Note-se que a escala de tempo é em horas.
pload / 51537 / 51537fig5highres.jpg "/>
Figura 5. Pequenos ensaios de agregação de tau na presença de THS. Na agregação induzida por heparina in vitro de 6 | iM de tau e p-tau foi avaliada no modo de medição contínua com tioflavina S como o corante indicador. Para além da proteína, cada reacção continha 30 | iM de heparina e 20 | iM no tampão de THS agregação de HEPES (10 mM HEPES pH 7,5, DTT 5 mM, EDTA 0,1 mM). Todos os ingredientes, excepto a heparina foram misturados e equilibrou-se à temperatura ambiente. Após a adição de heparina, a reacção foi transferida para a cuvete e colocadas no suporte de amostra. A fluorescência foi registada imediatamente a T 0, e continuou durante cerca de 2 horas ou até que o aumento de fluorescência retardada para perto de zero. Por causa do tempo de reacção relativamente curto, toda a reacção foi realizada à temperatura ambiente na mesma cuvette.
| tau | p-tau | |
| 60-100 uM tau | 50 ul | 0 ul |
| 60-100 uM p-tau | 0 ul | 50 ul |
| 300 uM heparina | 10 ul | 10 ul |
| tampão agregação | 39 ul | 39 ul |
| DTT 100 mM | 1 ul | 1 ul |
Tabela 1. componentes da mistura de agregação para não-dye, ensaio terminal.
| tau | p-tau | Tingir sozinho | |
| 60-100 uM tau | 50 ul | 0 ul | 0 ul |
| 60-100 uM p-tau | 0 ul | 50 ul | |
| 300 uM heparina | 20 ul | 20 ul | 20 ul |
| 3 mM de tioflavina T | 2 ul | 2 ul | 2 ul |
| Tampão Agregação | 126 ul | 126 ul | 176 ul |
| DTT 100 mM | 2 ul | 2 ul | 2 ul |
Tabela 2. componentes da mistura de agregação para com-dye, ensaio contínuo em um leitor de placas.
| tau | p-tau | Tingir sozinho | |
| 60-100 uM tau | 5 ul | 0 ul | 0 ul |
| 60-100 uM p-tau | 0 ul | 5 ul | 01; l |
| 300 uM heparina | 20 ul | 20 ul | 20 ul |
| 3 mM de tioflavina S | 1,5 l | 1,5 l | 1,5 l |
| Tampão Agregação | 171,5 ul | 171,5 ul | 176,5 ul |
| DTT 100 mM | 2 ul | 2 ul | 2 ul |
Tabela 3. componentes da mistura de agregação para com-dye, ensaio contínuo em um espectrofluorómetro compacto.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Este protocolo demonstra diferentes condições e instrumentos que detectam as tau rápida cinética de agregação dependente da fosforilação de ensaio. No ensaio de terminal, o corante de fluorescência ThT é adicionada a uma porção da reacção foi removida a partir da mistura principal em cada ponto de tempo. Amilóide A ligação induzida por fluorescência é então medida 26. Na segunda, com corante modo, a agregação tau é levada a cabo na presença de ThT ou THS, tornando este tipo de reacção adequados para a avaliação automática em tempo real do crescimento de agregados tau. Cada um desses métodos tem seus prós e contras.
A reacção de modo terminal é realizado apenas com os ingredientes necessários para a agregação tau. A diluição e a mistura da reacção com tioflavina T diminui drasticamente a taxa de aumento de fluorescência, essencialmente pára a reacção para a quantificação de fluorescência. Este método também é, portanto, compatível com o funcionamento manual. No entanto, porque o reaction está praticamente extinto após ThT disso, uma grande quantidade de tau pode ser necessária para traçar uma curva de agregação. Outra ressalva potencial para este método é que o acesso frequente à mistura de reacção pode introduzir contaminação microbiana ou proteolítica, ou oxidação de proteínas. Em contraste, o modo com corante permite a geração de amilóide na presença de ThT ou THS. O avanço de agregação pode ser monitorizada continuamente sem perturbar a reacção. Este recurso é particularmente atraente ao estabelecer uma plataforma de teste automatizado. No entanto, diferentes corantes podem induzir respostas específicas. Na verdade, ThT retarda tau e p-tau agregação significativamente, mas THS tem efeitos pouco (comparar Figuras 3 e 5). Existem vários outros corantes de fluorescência, incluindo Congo red e tiazinas, que têm sido utilizadas em estudos histológicos e biologia celular para a formação de PHF. Pelo menos um relatório indica que alguns desses corantes podem induzir a agregação de tau em tecidoscélulas de cultura de 45. Portanto, quando a escolha destes compostos para estudos cinéticos amiloidogese, cuidado deve ser praticado e diferentes corantes que podem ter que ser comparados.
No que diz respeito à escolha do instrumento, o fluorómetro única amostra utilizada na primeira abordagem é altamente fiável, mas a operação pode ser trabalhoso quando mais do que algumas reacções estão a ser comparadas. Usando vários cuvetes pode ajudar a evitar a contaminação cruzada entre as reações, embora o custo destas cuvetes de quartzo frágeis pode ser proibitivo para alguns. Por outro lado, o leitor de microplacas com poços múltiplos pode examinar várias reacções ao mesmo tempo. A utilização de placas de 96 poços descartáveis é vantajosa também. Com um elemento de aquecimento, um leitor de microplacas pode ser um dispositivo dedicado para monitorizar reacções múltiplas a 37 ° C durante um período de tempo prolongado. No entanto, a evaporação pode ser uma preocupação. DiNitto et al. Sobreposta uma reacção semelhante com óleo mineral para evitarevaporação 46.
Algumas precauções devem ser tomadas para os protocolos acima, para garantir resultados consistentes e quantitativos. Em primeiro lugar, tau-tau e p formam espontaneamente agregados amilóides ao longo do tempo, particularmente quando numa concentração elevada. É, portanto, imperativo para congelar todas as alíquotas de preparações protéicas, e descongelar apenas o volume necessário antes dos experimentos. Mesmo assim, alguns agregados, incluindo os intermediários granulares detectáveis por ThT 47 pode ter formado durante a preparação de proteínas recombinantes. A leitura de fluorescência inicial substancial de uma reação típica de agregação é assim comum. No entanto, a adição de um passo de pré-fiação e transferir o sobrenadante para um tubo separado, mesmo sem uma pílula de proteína visível, pode reduzir, e manter consistente fluorescência, inicial do mesmo lote de tau e p-tau prep. Em segundo lugar, a solução de estoque de trabalho de ThT (isto é, 60 uM) é estável à temperatura ambiente durante menos do que uma semana antes the fluorescência diminui. É, portanto, aconselhável a re-fazer a 60 mM ThT cada vários dias.
Uma razão importante estudos de agregação de p-tau subjacente é o desenvolvimento de novos agentes terapêuticos e de diagnóstico da DA. Os compostos que inibem ou reverter a agregação tau recombinante foram identificados a partir de telas de alto rendimento e testes direcionados 18,40,41,48. A eficácia destes compostos para a agregação p-tau permanece por esclarecer. Estas telas foram realizadas no modo terminal, dispensando uma mistura de agregação comum sem o corante para poços de múltiplas placas individuais com diferentes compostos. Tipicamente, depois de O / N de incubação, ThT ou THS é adicionado para a medição de fluorescência, revelando o poder inibitório de vários compostos. A abordagem com-dye mencionado acima e por Rankin et al. 49 ainda tem que ser incorporada às telas de alto rendimento. Agora com tau hyperphosphorylated disponível para estudos cinéticos e farmacêuticas do emparelhadoformação de filamentos helicoidal, a descoberta de medicamentos da doença de Alzheimer é provável que avançar mais longe.
Por fim, vale ressaltar que o estudo da agregação p-tau é de fundamental importância, não só para tauopatias, mas também pode afetar uma população ainda maior. Por exemplo, há relatos de que os emaranhados neurofibrilares são detectáveis em alguns pacientes de encefalopatia traumática crônica, como jogadores profissionais de futebol americano e boxeadores 50-52. Correlação semelhante também foi relatada por pacientes com lesão cerebral simples ou repetitivos traumáticas incluindo soldados 53. Protocolos descritos neste trabalho podem, assim, ajudar a descoberta e desenvolvimento de novos agentes terapêuticos que visam agregados tau-p em células neuronais.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Trizma base | Sigma | T1503 | |
| NaCl | Macron Fine Chemicals | MAL-7581-06 | |
| Ethylenediaminetetraacetic Acid (EDTA) | Invitrogen | 15576-028 | |
| Thioflavin T | Sigma | T3516 | Stored in dark |
| Thioflavin S | Sigma | T1892 | Stored in dark |
| heparin | Sigma | H3393 | |
| DL-Dithiothreitol (DTT) | Sigma | D9779 | Stored at 4 °C |
| 96-well plate | Corning | 3917 | |
| ISA SPEX FluoroMax-2 | Horiba | ||
| SpectraMax M2 Multi-Mode Microlate Reader | Molecular Devices | ||
| Mouse Anti-Tau Monoclonal Antibody | R&D Systems | MAB3494 | Stored at –80 °C |
References
- Lee, V. M., Goedert, M., Trojanowski, J. Q. Neurodegenerative tauopathies. Annu Rev Neurosci. 24, 1121-1159 (2001).
- Ballatore, C., Lee, V. M., Trojanowski, J. Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 8 (9), 663-672 (2007).
- Arriagada, P. V., Marzloff, K., Hyman, B. T. Distribution of Alzheimer-type pathologic changes in nondemented elderly individuals matches the pattern in Alzheimer's disease. Neurology. 42 (9), 1681-1688 (1992).
- Arriagada, P. V., Growdon, J. H., Hedley-Whyte, E. T., Hyman, B. T. Neurofibrillary tangles but not senile plaques parallel duration and severity of Alzheimer's disease. Neurology. 42 (3 Pt 1), 631-639 (1992).
- Bancher, C., Braak, H., Fischer, P., Jellinger, K. A. Neuropathological staging of Alzheimer lesions and intellectual status in Alzheimer's and Parkinson's disease patients. Neurosci Lett. 162 (1-2), 179-182 (1993).
- Guillozet, A. L., Weintraub, S., Mash, D. C., Mesulam, M. M. Neurofibrillary tangles, amyloid, and memory in aging and mild cognitive impairment. Arch Neurol. 60 (5), 729-736 (2003).
- Hasegawa, M., et al. Protein sequence and mass spectrometric analyses of tau in the Alzheimer's disease brain. J Biol Chem. 267 (24), 17047-17054 (1992).
- Matsuo, E. S., et al. Biopsy-derived adult human brain tau is phosphorylated at many of the same sites as Alzheimer's disease paired helical filament tau. Neuron. 13 (4), 989-1002 (1994).
- Bamburg, J. R., Bloom, G. S. Cytoskeletal pathologies of Alzheimer disease. Cell Motil Cytoskeleton. 66 (8), 635-649 (2009).
- Denk, F., Wade-Martins, R. Knock-out and transgenic mouse models of tauopathies. Neurobiol Aging. 30 (1), 1-13 (2009).
- Gong, C. X., Iqbal, K. Hyperphosphorylation of microtubule-associated protein tau: a promising therapeutic target for Alzheimer disease. Curr Med Chem. 15 (23), 2321-2328 (2008).
- Mazanetz, M. P., Fischer, P. M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat Rev Drug Discov. 6 (6), 464-479 (2007).
- Brunden, K. R., Trojanowski, J. Q., Lee, V. M. Advances in tau-focused drug discovery for Alzheimer's disease and related tauopathies. Nat Rev Drug Discov. 8 (10), 783-793 (2009).
- Querfurth, H. W., LaFerla, F. M. Alzheimer's disease. N Engl J Med. 362 (4), 329-344 (2010).
- Buee, L., Bussiere, T., Buee-Scherrer, V., Delacourte, A., Hof, P. R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev. 33 (1), 95-130 (2000).
- Lee, V. M., Brunden, K. R., Hutton, M., Trojanowski, J. Q. Developing therapeutic approaches to tau, selected kinases, and related neuronal protein targets. Cold Spring Harb Perspect Med. 1 (1), 006437 (2011).
- Mandelkow, E. M., Mandelkow, E. Biochemistry and cell biology of tau protein in neurofibrillary degeneration. Cold Spring Harb Perspect Med. 2 (7), a006247 (2012).
- Bulic, B., Pickhardt, M., Mandelkow, E. Progress and Developments in Tau Aggregation Inhibitors for Alzheimer Disease. J Med Chem. 56 (11), 4135-4155 (2013).
- Cowan, C. M., Quraishe, S., Mudher, A. What is the pathological significance of tau oligomers. Biochem Soc Trans. 40 (4), 693-697 (2012).
- Spires-Jones, T. L., Kopeikina, K. J., Koffie, R. M., de Calignon, A., Hyman, B. T. Are tangles as toxic as they look. J Mol Neurosci. 45 (3), 438-444 (2011).
- SantaCruz, K., et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 309 (5733), 476-481 (2005).
- Wittmann, C. W., et al. Tauopathy in Drosophila: neurodegeneration without neurofibrillary tangles. Science. 293 (5530), 711-714 (2001).
- Shulman, J. M., Feany, M. B. Genetic modifiers of tauopathy in Drosophila. Genetics. 165 (3), 1233-1242 (2003).
- Wille, H., Drewes, G., Biernat, J., Mandelkow, E. M., Mandelkow, E. Alzheimer-like paired helical filaments and antiparallel dimers formed from microtubule-associated protein tau in vitro. J Cell Biol. 118 (3), 573-584 (1992).
- Alonso, A., Zaidi, T., Novak, M., Grundke-Iqbal, I., Iqbal, K. Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci U S A. 98 (12), 6923-6928 (2001).
- Barghorn, S., Biernat, J., Mandelkow, E. Purification of recombinant tau protein and preparation of Alzheimer-paired helical filaments in vitro. Methods Mol Biol. 299, 35-51 (2005).
- Wilson, D. M., Binder, L. I. Polymerization of microtubule-associated protein tau under near-physiological conditions. J Biol Chem. 270 (41), 24306-24314 (1995).
- Wilson, D. M., Binder, L. I. Free fatty acids stimulate the polymerization of tau and amyloid beta peptides. In vitro evidence for a common effector of pathogenesis in Alzheimer's disease. Am J Pathol. 150 (6), 2181-2195 (1997).
- Perez, M., Valpuesta, J. M., Medina, M., Montejo de Garcini, E., Avila, J. Polymerization of tau into filaments in the presence of heparin: the minimal sequence required for tau-tau interaction. J Neurochem. 67 (3), 1183-1190 (1996).
- Carlson, S. W., et al. A complex mechanism for inducer mediated tau polymerization. Biochemistry. 46 (30), 8838-8849 (2007).
- Goedert, M., et al. Assembly of microtubule-associated protein tau into Alzheimer-like filaments induced by sulphated glycosaminoglycans. Nature. 383 (6600), 550-553 (1996).
- King, M. E., Gamblin, T. C., Kuret, J., Binder, L. I. Differential assembly of human tau isoforms in the presence of arachidonic acid. J Neurochem. 74 (4), 1749-1757 (2000).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pseudo-phosphorylation of tau at Ser202 and Thr205 affects tau filament formation. Brain Res Mol Brain Res. 138 (1), 84-93 (2005).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Pre-assembled tau filaments phosphorylated by GSK-3b form large tangle-like structures. Neurobiol Dis. 31 (3), 368-377 (2008).
- Grundke-Iqbal, I., et al. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci U S A. 83 (13), 4913-4917 (1986).
- Castellani, R. J., Perry, G. Pathogenesis and disease-modifying therapy in Alzheimer's disease: the flat line of progress. Arch Med Res. 43 (8), 694-698 (2012).
- Green, R. C., et al. Effect of tarenflurbil on cognitive decline and activities of daily living in patients with mild Alzheimer disease: a randomized controlled trial. JAMA. 302 (23), 2557-2564 (2009).
- Gauthier, S., et al. Effect of tramiprosate in patients with mild-to-moderate Alzheimer's disease: exploratory analyses of the MRI sub-group of the Alphase study. J Nutr Health Aging. 13 (6), 550-557 (2009).
- Pickhardt, M., et al. Anthraquinones inhibit tau aggregation and dissolve Alzheimer's paired helical filaments in vitro and in cells. J Biol Chem. 280 (5), 3628-3635 (2005).
- Crowe, A., Ballatore, C., Hyde, E., Trojanowski, J. Q., Lee, V. M. High throughput screening for small molecule inhibitors of heparin-induced tau fibril formation. Biochem Biophys Res Commun. 358 (1), 1-6 (2007).
- Taniguchi, S., et al. Inhibition of heparin-induced tau filament formation by phenothiazines, polyphenols, and porphyrins. J Biol Chem. 280 (9), 7614-7623 (2005).
- Sigurdsson, E. M. Tau-focused immunotherapy for Alzheimer's disease and related tauopathies. Curr Alzheimer Res. 6 (5), 446-450 (2009).
- Tan, Y. J., et al. Phosphopeptide Enrichment with TiO-Modified Membranes and Investigation of Tau Protein Phosphorylation. Anal Chem. 85 (12), 5699-5706 (2013).
- Santa-Maria, I., Perez, M., Hernandez, F., Avila, J., Moreno, F. J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J Alzheimers Dis. 9 (3), 279-285 (2006).
- Lira-De Leon, K. I., et al. Molecular mechanism of tau aggregation induced by anionic and cationic dyes. J Alzheimers Dis. 35 (2), 319-334 (2013).
- DiNitto, J. P., Wang, L., Wu, J. C. Continuous fluorescence-based method for assessing dicer cleavage efficiency reveals 3' overhang nucleotide preference. BioTechniques. 48, 303-311 (2010).
- Maeda, S., et al. Granular tau oligomers as intermediates of tau filaments. Biochemistry. 46 (12), 3856-3861 (2007).
- Pickhardt, M., et al. Phenylthiazolyl-hydrazide and its derivatives are potent inhibitors of tau aggregation and toxicity in vitro and in cells. Biochemistry. 46 (35), 10016-10023 (2007).
- Rankin, C. A., Sun, Q., Gamblin, T. C. Tau phosphorylation by GSK-3beta promotes tangle-like filament morphology. Mol Neurodegener. 2, 12 (2007).
- McKee, A. C., et al. Chronic traumatic encephalopathy in athletes: progressive tauopathy after repetitive head injury. J Neuropathol Exp Neurol. 68 (7), 709-735 (2009).
- Herrup, K. Reimagining Alzheimer's disease--an age-based hypothesis. J Neurosci. 30 (50), 16755-16762 (2010).
- Gavett, B. E., Stern, R. A., McKee, A. C. Chronic traumatic encephalopathy: a potential late effect of sport-related concussive and subconcussive head trauma. Clin Sports Med. 30 (1), 179-188 (2011).
- Tsitsopoulos, P. P., Marklund, N. Amyloid-beta Peptides and Tau Protein as Biomarkers in Cerebrospinal and Interstitial Fluid Following Traumatic Brain Injury: A Review of Experimental and Clinical Studies. Front Neurol. 4, 79 (2013).
