

Abstract
Det er blevet mere og mere tydeligt, at den rumlige distribution og bevægelsen af membran komponenter som lipider og proteiner er vigtige faktorer i reguleringen af mange cellulære funktioner. Men på grund af de hurtige dynamik og de små strukturer, der er involveret, er en meget høj rumlig-tidslig opløsning kræves for at fange den virkelige opførsel af molekyler. Her præsenterer vi forsøgsprotokollen for at studere dynamikken i fluorescens-mærket plasma-membran proteiner og lipider i levende celler med høj Spatiotemporal opløsning. Især er denne fremgangsmåde ikke brug for at spore hver enkelt molekyle, men det beregner befolkning adfærd ved hjælp af alle molekyler i en given region af membranen. Udgangspunktet er en hurtig billeddannelse af en given region på membranen. Bagefter er en komplet spatio-temporale autokorrelationsfunktion beregnet korrelere erhvervede billeder med stigende forsinkelser, for eksempel hver 2, 3, n gentagelser. Det er muligt at påvise, at breddenaf toppen af den rumlige autokorrelationsfunktion stiger ved stigende tidsforsinkelse som en funktion af partikel bevægelse som følge af diffusion. Derfor montering af rækken af autokorrelationsfunktioner gør det muligt at udtrække den faktiske protein mean square forskydning fra imaging (iMSD), her præsenteret i form af tilsyneladende diffusivitet vs gennemsnitlige forskydning. Dette giver en kvantitativ betragtning af de gennemsnitlige dynamik i enkelte molekyler med nanometer nøjagtighed. Ved at anvende en GFP-mærket variant af transferrin-receptor (TFR) og en ATTO488 mærket 1-palmitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamin (PPE), er det muligt at observere spatiotemporale regulering af protein og lipid udbredelse på um-størrelse membran regioner i mikro-til-milli-sekund tidsinterval.
Introduction
Startende fra den oprindelige "flydende mosaik" model ved sanger og Nicolson, har billedet af cellulære plasmamembran løbende blevet opdateret i løbet af de seneste årtier for at medtage den nye rolle Cytoskeleton og lipid domæner 1,2.
De første observationer blev opnået ved fluorescerende bedring efter fotoblegning (FRAP) afsløringen, at en betydelig del af membranproteiner er immobil 3-5. Disse banebrydende undersøgelser, selv om meget informativ, lidt fra den relativt dårlige opløsning i rummet (mikron) og tid (sekunder) i FRAP opsætninger. Også bliver et ensemble gennemsnit måling FRAP mangler i at give oplysninger om enkelt molekyle adfærd.
I denne sammenhæng muligheden specifikt mærke et enkelt molekyle med meget lyse tags (tillader studiet af diffusionsprocessen et molekyle ad gangen) har været meget vellykket. Især ved at skubbetidsopløsning af det indre Particle Tracking (SPT) tilgang til mikrosekunder tidshorisont, Kusumi, et al. fået adgang til ukendte egenskaber af lipid og protein dynamik, der i høj grad har bidraget til anerkendelsen af den rolle, skelet membran actin-baserede i membran fysiologi 6 7. Disse fund genereret den såkaldte den "picket og hegn 'model, hvor lipid og protein diffusion er reguleret af actin-baserede skelet. Men for at få adgang til den enorme mængde af oplysninger fra SPT mange eksperimentelle spørgsmål har, som skal løses. Især er fremgangsmåden mærkning typisk sammensat af mange trin som produktion, oprensning og indførelse af de mærkede arter til systemet. Desuden store etiketter, ligesom kvantepunkter eller metal nanopartikler, der ofte kræves for at nå sub-millisekund tidshorisont og tværbinding af målmolekylerne af miljømærket ikke kan undgås i mange tilfælde. Endelig er mange banerskal registreres til at passe statistiske kriterier og samtidig en lav densitet af etiketten er forpligtet til at tillade sporing.
Sammenlignet med SPT, fluorescenskorrelationsspektroskopi (FCS), overvinde mange af disse ulemper, repræsenterer en meget lovende tilgang til at studere molekylær dynamik. Faktisk FCS fungerer godt også med dæmpet og tætte etiketter, gør det muligt at studere dynamikken af fluorescerende protein-mærkede molekyler i transient transficerede celler. Også, gør det muligt at nå høje statistik i en begrænset mængde tid. Endelig, på trods af "høj" densiteten for etiketter FCS giver enkelte molekyler oplysninger. Tak til alle disse egenskaber, FCS repræsenterer en meget enkel tilgang og er blevet grundigt anvendt til at studere lipid og protein dynamik både i modelmembraner og i levende celler 8-10. Mange forskellige fremgangsmåder er blevet foreslået at gøre det muligt for FCS at afsløre detaljerne i molekylær diffusion. For eksempel var det shegne, ved at udføre FCS på forskellige størrelser observation områder man kan definere en "FCS diffusion law" oplysende skjulte funktioner i molekylær bevægelse 11,12. Udover varieres i størrelse, blev indsatsområdet også duplikeres 13, flyttede i rummet langs linjerne 14-20 eller konjugerede med hurtige kameraer 21,22. Ved hjælp af disse 'spatio-temporale' korrelation tilgange blev relevante biologiske parametre for flere membrankomponenter kvantitativt beskrevet på begge modelmembraner og faktiske biologiske dem og dermed et indblik i membran rumlige organisering.
Men i alle FRAP og FCS-applikationer beskrevet hidtil størrelsen af det prioriterede område udgør en grænse i rumlig opløsning, der ikke kan overvindes. Adskillige super opløsning billeddannelse metoder er for nylig blevet udviklet til at omgå denne grænse. Nogle er baseret på lokalisering præcision, såsom stokastiske optisk genopbygning mikroskopi (STORM) <sup> 23,24, fotoaktiveringsprogrammet lokalisering mikroskopi (PALM) 25, fluorescens PALM (FPALM) 26, og single-partikel sporing PALM (sptPALM) 27: den relativt store mængde af fotoner, der kræves på hvert snapshot begrænser imidlertid tidsopløsning disse metoder til mindst flere millisekunder, hvilket hæmmer deres anvendelighed in vivo.
I modsætning hertil er et lovende alternativ til super opløsning billeddannelse blevet åbnet af rumligt modulere fluorescensemissionen med stimuleret emission depletion metoder (STED eller reversible mætbare optiske fluorescens overgange (RESOLFT)) 28,29. Disse metoder kombineres udformningen af observationsvolumenet godt under diffraktionsgrænsen med muligheden for at anvende hurtig scanning mikroskoper og detektionssystemer. I kombination med fluorescens udsving analyse STED mikroskopi lov til direkte sonde nanoskala spatiotemporale dynamik lipider og proteins i levende cellemembraner 30,31.
De samme fysiske mængder af STED-baserede mikroskopi kan opnås ved en modificeret spatio-temporale billede korrelation spektroskopi (STICS 32,33) metode, der er egnet til undersøgelse af dynamikken i fluorescens-kodede membranproteiner og / eller lipider i levende celler og af en kommerciel mikroskop. Den eksperimentelle protokol præsenteres her består af få trin. Den første kræver en hurtig billeddannelse af regionen af interesse på membranen. Derefter bliver den resulterende stabel af billeder anvendes til at beregne de gennemsnitlige rumlige-tidsmæssige korrelationsfunktioner. Ved at montere den række af korrelationsfunktioner, kan den molekylære "diffusion ret« hentes direkte fra billeddannelse i form af en tilsyneladende diffusivitet (D app) - vs -Gennemsnitlig forskydning plot. Dette plot kritisk afhænger af omgivelserne udforsket af molekylerne og tillader direkte anerkende de faktiske diffusion tilstandeaf lipid / protein af interesse.
I flere detaljer, som tidligere vist 34, det spatio-temporale autokorrelationsfunktion af den erhvervede billedserier kritisk afhænger af dynamikken i molekylerne bevæger sig i den indsamlede billede serien (bemærk, at det samme ræsonnement kan anvendes i en linje erhvervelse hvor kun én dimension i rummet betragtes). Især definerer vi korrelationsfunktionen som:
 (1)
(1)
hvor  repræsenterer den målte fluorescensintensitet i positionen x, y og på tidspunkt t,
repræsenterer den målte fluorescensintensitet i positionen x, y og på tidspunkt t, ![]() og
og ![]() repræsenterer afstanden i x ogy-retningerne henholdsvis
repræsenterer afstanden i x ogy-retningerne henholdsvis ![]() repræsenterer tidsforsinkelsen, og
repræsenterer tidsforsinkelsen, og ![]() repræsenterer gennemsnittet. Denne funktion kan udtrykkes som:
repræsenterer gennemsnittet. Denne funktion kan udtrykkes som:
 (2)
(2)
hvor »N« repræsenterer det gennemsnitlige antal af molekyler i området observation, ![]() repræsenterer foldning drift i rummet, og
repræsenterer foldning drift i rummet, og  repræsenterer autokorrelation instrumentale taljen. Sidstnævnte kan tolkes som et mål for, hvor fotonerne i en enkelt emitter er spredt ud i rummet på grund af den optiske / indspilning opsætningen (den såkaldte punktspredningsfunktion, PSF, genRALLY godt tilnærmes ved en Gaussisk funktion). Endelig
repræsenterer autokorrelation instrumentale taljen. Sidstnævnte kan tolkes som et mål for, hvor fotonerne i en enkelt emitter er spredt ud i rummet på grund af den optiske / indspilning opsætningen (den såkaldte punktspredningsfunktion, PSF, genRALLY godt tilnærmes ved en Gaussisk funktion). Endelig  repræsenterer sandsynligheden for at finde en partikel i en afstand
repræsenterer sandsynligheden for at finde en partikel i en afstand ![]() og
og ![]() efter en forsinkelse
efter en forsinkelse ![]() . Hvis vi overveje et diffusive dynamik, hvor partikler bevæger sig tilfældigt i alle retninger, og netto flux ikke er til stede, er denne funktion også godt tilnærmes ved en Gaussisk funktion, hvor variansen kan identificeres som Mean Square Displacement (MSD) i partikel den bevægelige . Således talje korrelationsfunktionen (også benævnt
. Hvis vi overveje et diffusive dynamik, hvor partikler bevæger sig tilfældigt i alle retninger, og netto flux ikke er til stede, er denne funktion også godt tilnærmes ved en Gaussisk funktion, hvor variansen kan identificeres som Mean Square Displacement (MSD) i partikel den bevægelige . Således talje korrelationsfunktionen (også benævnt ![]() ), Kan defineres som summen af partikel sygdomme i bevægeapparatet og instrumentale taljen og kan måles ved en gaussisk pasformTing af korrelationen funktion for hver tidsforsinkelse. Den målte jeg MSD kan bruges til at beregne en tilsyneladende diffusionsevne de bevægelige molekyler
), Kan defineres som summen af partikel sygdomme i bevægeapparatet og instrumentale taljen og kan måles ved en gaussisk pasformTing af korrelationen funktion for hver tidsforsinkelse. Den målte jeg MSD kan bruges til at beregne en tilsyneladende diffusionsevne de bevægelige molekyler ![]() og en gennemsnitlig forskydning
og en gennemsnitlig forskydning ![]() som:
som:
 (3)
(3)
 (4)
(4)
Par overvejelser om forsøgsopstillingen anvendes kan vejlede læseren gennem de følgende afsnit. For selektivt at excitere fluoroforer på basalmembranen af levende celler, vil vi benytte en total intern refleksion (TIR) belysning ved hjælp af en kommerciel TIR-fluorescens (TIRF) mikroskop (detaljer kan findes i materialet afsnit). Desuden, for at indsamle the fluorescens vil vi bruge en høj forstørrelse målsætning (100X NA 1,47, er høj numerisk blænde til TIRF belysning) og en EMCCD kamera (fysiske størrelse af pixel på chippen 16 um). For at nå en pixelstørrelse på 100 nm anvender vi en yderligere forstørrelse linse 1,6X. Som omtalt nedenfor, vil en tidsopløsning under 1 msek kræves til korrekt at beskrive dynamikken af hurtige membranlipider under 100 nm. For at nå denne tidsmæssige opløsning vi nødt til at vælge et område af interesse (ROI) er mindre end hele chippen på kameraet (512 x 512). På denne måde, vil kameraet læse et reduceret antal linjer øger tidsopløsning. Men i denne udlæsning regime rammen gang ville være begrænset af den tid, der kræves for at flytte afgifter fra eksponeringen for udlæsning chip på kameraet og er normalt i størrelsesordenen millisekunder til 512 x 512 pixel EMCCD. At slå denne grænse, en ny teknologi gør det muligt at flytte ROI-linjer kun i stedet for hele rammen, wed en praktisk effektiv reduktion af den eksponerede chip størrelse (kaldet Beskåret Sensor tilstand i vores EMCCD). Til denne konfiguration skal være effektiv, skal chippen uden ROI være dækket af et par slidser monteret i den optiske vej. Takket være denne opsætning en tidsopløsning ned til 10 -4 sekunder kan opnås. Bemærk dog, at denne fremgangsmåde kan kobles med mange forskellige forsøgsopstillinger, som forklaret i "diskussion" sektionen.
Demonstration af fremgangsmåden vil blive givet i levende celler, ved hjælp af både en ATTO488 mærket 1-palmitoyl-2-hydroxy-sn-glycero-3-phosphoethanolamin (ATTO488-PPE) og GFP-mærket variant af transferrin-receptor (GFP- TFR). I tilfælde af ATTO488-PPE denne fremgangsmåde med succes kan gendanne en næsten konstant D app som en funktion af gennemsnitlig forskydning indikerer en hovedsagelig fri diffusion, som tidligere rapporteret 30,35. Derimod TFR-GFP viser en faldende D
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. systemkalibrering
- Punktspredningsfunktion (PSF) med kalibrering
- Fortynd 10 pi 30 nm fluorescerende vulst opløsning (ca. 5 uM) i 90 ul destilleret vand og derefter sonikeres opløsningen i 20 minutter. Skær en firkant (1 cm x 1 cm) stykke agarosegel (3%) og deponering 10 ul af opløsningen på toppen af gelen. Vælte stykke gel på bunden glas af en 2 cm petriskål og presse dråbe på glasset.
- Tænd setup erhvervelse, sætte prøven i holderen, skal du indstille kameraets eksponering og EMgain (100 ms og 1000 er gode parametre, men optimere ifølge systemet) og vente til kameraet for at køle ned.
- Indstil kamera eksponering til 100 ms, kamera EMgain til 1.000, dataopsamling til Frame Transfer, 100 gentagelse og auto gem indstilling.
- Brug af okularet og transmitteret lys fokus på grænsen af gelen og derefter flytte det mål til midten af gelen, justere fokus og start proceduren laserjustering (i LAS AF, vælg 'TIRF setup', og følg den automatiske procedure justering).
- Find et synsfelt med isolerede enkelt pletter, nøjagtigt fokus på lysere plet (der normalt repræsenterer perler aggregat) som en reference, erhverve 100 billeder, og gentag trin 5-6 gange for at erhverve flere enlige pletter.
- Importer den erhvervede serie til et databehandling program, og gennemsnittet stakken i tiden (figur 1A), og vælg et enkelt isoleret perle. Vær omhyggelig med at vælge de mindste at undgå partikelaggregater.
- Passer til den valgte intensitet distribution (et eksempel på en enkelt perler profil er præsenteret i figur 1B) med en Gaussisk funktion ved hjælp af kommandoen "gaussfit" (i ICS-Matlab værktøjer i Materialer i Matlab). Kontroller godhed fit ved at inspicere de opnåede residualer (et eksempel på udstyret Gaussian profil med de tilsvarende residualer præsenteres in Figur 1B).
- Kamera kalibrering
- Tænd kameraet, og vent til kameraet for at køle ned. Indstil indstilling kamera køb, (dvs. for den anvendte kamera vi indstille eksponeringen til 0,5 ms, kamera EMgain til 1.000, dataopsamling til Beskåret Mode størrelse ROI til 32 x 128 10.000 gentagelser) og start købet af kameraets baggrund signalet.
- Import erhvervede ramme serie til et databehandling program. Beregn og inspicere den gennemsnitlige intensitet i hver pixel for at kontrollere, at kameraet baggrund er omtrent fladt i det valgte område af chippen. I Beskåret tilstand, fjerne den første og de sidste par vandrette linjer (3 til 10 afhængigt af størrelsen af ROI) for hver ramme fordi kameraet baggrund sædvanligvis er forudindtaget i grænseområderne linjer.
- Oprette et histogram af værdierne (også defineret Digital Niveau, DL) i erhvervede billeder stabel (ved hjælp af kommandoen 'historik' i Matlab), og plotte logaritmenaf resulterende frekvens (ved hjælp semilogy kommando i Matlab). Et eksempel DL fordeling til kamera baggrund er vist i figur 2.
BEMÆRK: Hvis kameraet fungerer godt, vil plottet viser en tilnærmelsesvis Gaussisk top (en parabolsk profil i log-skala), der repræsenterer fordelingen af værdier er forbundet til nul foton efterfulgt af en eksponentiel henfald (en linie med negativ hældning i log-skala ), der repræsenterer fordelingen af værdier, der er forbundet til en foton (figur 2). Især midten og variansen af den gaussiske funktion repræsenterer kameraet offset og fejl, henholdsvis mens henfaidskonstant eksponentielle del repræsenterer et estimat af DL tildelt af kamera til hver enkelt foton. I Matlab bruge afsnittet "CalibrateCamera" i Script støtte materialer. - Gentag handlingen for alle de valgte kamera EMGain og Gain.
2. MærketCelle Forberedelse
- Til fremstilling af liposomer, der kræves for lipid inkorporering 36 opløses separat 1 mg DOPE (1,2-dioleoyl--sn-glycero-3-phosphoethanolamin), 1 mg DOTAP (1,2-dioleoyl-3-trimethylammonium-propan), og 1 mg PPE-ATTO488 i 1 ml kloroform. Bland 0,5 ml DOPE-opløsning, 0,5 ml DOTAP opløsning, og 25 ul af PV-ATTO488 opløsning og tørres under vakuum i 24 timer. Tilsæt 0,5 ml HEPES-buffer 20 mM, vortex i 15 minutter og sonikeres i 15 minutter ved 40 ° C.
- For at forberede cellen, vaskes 3 gange med PBS en P100 skål af sammenflydende CHO-K1 (kinesisk hamster ovarie), tilsættes 1 ml trypsin og butik i kuvøse i 5 min. Suspender løsnede celler tilføje 9 ml DMEM / F12-medium suppleret med 10% FBS og frø 150 pi celle opløsning i en petriskål indeholdende 800 ul af det samme medium.
- Opbevares i inkubator i 24 timer ved 37 ° C og 5% CO2. For lipid inkorporering, erstatte cellemedium med 500ul serum-frit medium; efter 30 min, tilsættes 2 pi af liposomer opløsning; efter 15 min vask med PSB og tilføje nyt DMEM / F12 medium for billeddannelse.
- Til transfektion, transfektion celler ifølge Lipofectamin protokol (fabrikantens anvisninger) ved hjælp af TFR-GFP plasmid og butik 24 timer i inkubator før billeddannelse.
3. Data Acquisition
- Klargøring Opsætning
- For at termostat mikroskop 24 timer før forsøget tænde inkubatoren.
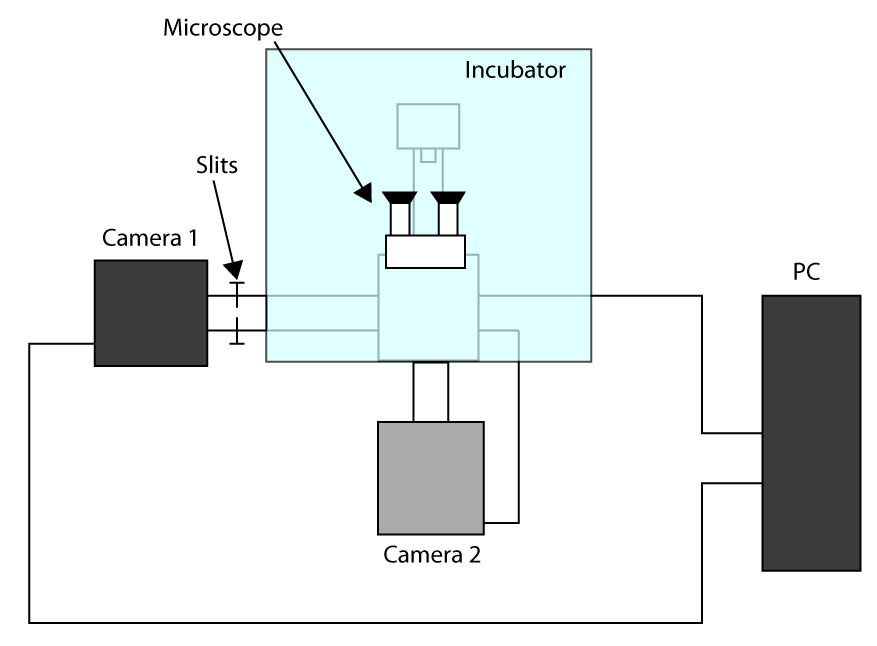
- For at kunne anvende den hurtigste opnåelige erhvervelse tid, arbejde i Beskåret Sensor tilstand (se indledning) og bruge en første kamera til billedbehandling (kamera 1) og det andet kamera til at markere cellen (2 kamera). En ordning af opsætningskonfigurationen er præsenteret i supplerende Figur S1. Så, for at tilpasse to kameraer tur på mikroskopet og vente på kameraer til at køle ned.
- Set på begge kameraer Parametrene for transmitteret lys billeddannelse (dvs.
- Sæt prøverne i holderen og fokusere ved hjælp okular, sende lys til kamera 1 og skub forsigtigt spalterne tillader lys kun på ROI bruges til cell imaging (her en 32 x 32 pixels ROI).
- Flytte en celle i det valgte område og sende lyset til kameraet 2, derefter tegne en ROI i den software, der styrer kamera 2 for at få en henvisning.
- Imaging (figur 3A)
- Først og fremmest, skal du justere TIRF laser ifølge proceduren i din opsætning. I vores setup, skal du vælge 'TIRF setup' og start proceduren auto justering. Når laseren er justeret sæt 70 nm indtrængningsdybde (ca. 70 °).
- Indstil eksponeringstid til 70 ms og EMGain til 100 på både kamera 1 og kamera på 2; derefter vælge en celle ved hjælp af kamera 1 og derefter sende lyset på kameraet 2 og præcist fokus cellemembranen. Indstil den minimale eksponering på camera 2, 1000 EMGain, Beskåret Sensor Mode 10 5 gentagelser og sæt autosave som passer filer (fleksibelt image Transport System, et format, der kan være let at administrere).
- Start købet optage billedet serien. Slip Gain og det beskårne tilstand at tillade stabilisering temperatur, før erhverve en ny celle, derefter gentage de sidste to trin for at opnå 8-10 celler.
4. Beregning af Mean Square Forskydning fra Imaging (i MSD)
BEMÆRK: Følgende protokol kan anvendes direkte til rådata. På samme tid er hele protokollen er gyldig for erhvervelser simulerede både i Matlab og SimFCS data. Linket til de tilsvarende tutorials kan findes i "Materials 'sektionen.
- Beregning af Matlab
- Importer den erhvervede serie til Matlab hjælp ImportImageSeries script. Beregn den gennemsnitlige intensitet af hvert billede i tid ved hjælp af den fællesmmand betyde på de første 2 dimensioner og bruge plot for at se den resulterende vektor.
- Hvis mere end 10% af fotoblegning er til stede, skal du kassere den serie eller fjerne den første del af dem. Hvis det er lavere, forsøge at afhjælpe effekten på korrelationsfunktionen ved at trække til hvert billede dens gennemsnitlige intensitet, som vist før 37.
- Beregn den gennemsnitlige intensitet af hver pixel ved hjælp af middelværdi på den tredje dimension og se resulterende billede.
BEMÆRK: Særlig opmærksomhed er påkrævet for at undgå artefakt-korrelationer. I virkeligheden kunne som tidligere vist for lignende teknikker 38, celle grænser samt ude af fokus vesikler indføre en stærk korrelation. Hvis inspektionen af det gennemsnitlige billede afslører celle grænser eller ude af fokus vesikler, så prøv at udelukke involverede ellers kassere købet regionen. For at rette effekten af denne immobile strukturer trække den gennemsnitlige tidsmæssige intensitet fra hver pixel 39. - Beregn than spatiotemporale korrelation (G (ξ, χ, τ)) ved hjælp af funktionen CalculateSTICScorrfunc. Fjern G (ξ, χ, 0), fordi korrelationen på grund af skud støj i svagt lys regime dominerer G (0,0,0); korrelationen grund detektoren dominerer G (± 1,0,0) og partikel bevægelse under eksponeringen tid kunne deformere G (ξ, χ, τ) til τ = 0 ved at øge den målte taljen (denne effekt forsvinder for τ > 0) 34.
- Gennemsnit G (ξ, χ, τ> 0) ved hjælp af en logaritmisk tid-bin for at reducere støjen ved hjælp af "LogBinStack" funktion i støttemateriale og derefter passe det resulterende G (ξ, χ, τ) ved hjælp af funktionen "gaussfit" af ICS-Matlab værktøjer i materialerne til at inddrive i MSD (anden kolonne i den resulterende array).
- Plot opnåede taljen σ (τ) 2 (i MSD) som en funktion af tiden. Hvis dataene er for støjende, så prøv at øge antallet af acquIRED rammer, øge laser magt, gennemsnit mere G (ξ, χ, τ) sammen.
- Beregning af SimFCS
- Åbn de erhvervede filer med ImageJ hjælp BioFormat importør plugin og gem erhvervet serier som Tiff sekvens.
- Åbent SimFCS og vælg RICS værktøj og vælge Filer> Importer flere billeder (supplerende figur S2).
- Vælg Tilpas, skal du indsætte de korrekte erhvervelse parametre og luk pasform vindue (supplerende figur S3).
- Vælg Skærm> gennemsnitlig intensitet> CH1 og kontrollere tilstedeværelsen af fotoblegning (supplerende figur S4).
- Hvis mere end 10% af fotoblegning er til stede udsmid serien, eller hvis det er muligt belastning igen billedet sekvens fjerne den første del af serien.
- Hvis blegning det er mindre end 10% vælge Værktøjer> i MSD> indstille parametre, tjek 'Brug glidende gennemsnit «, der i panelet ROI til venstre ensortjord af ramme til det glidende gennemsnit opmærksom at korrespondent tiden er højere end den karakteristiske diffusion (for partikel bevæger sig ved 1 um 2 sek -1 en tid på 10 sekunder er en god glidende gennemsnit)
- Vælg Værktøjer> iMSD> Beregn I MSD (supplerende figur S5) og pasform og eksportere jeg MSD fra notesblok (supplerende figur S6).
5. Beregning af diffusion lov fra I MSD
- Monter de første par punkter at ekstrapolere skæringspunkt (σ 0 2) (5 point er som regel nok, men flere punkter kan monteres, hvis de viser en lineær adfærd) og sammenligne denne værdi med den tidligere målt PSF 2. Hvis de er sammenlignelige, er dynamikken i isolerede fluoroforer følges. Hvis derimod σ 0 2 >> PSF 2 forsøge at erhverve hurtigere at sikre, atingen skjulte dynamik er til stede 34.
- Beregn den tilsyneladende diffusivitet (D app) og den gennemsnitlige forskydning (R) under anvendelse af ligningerne 3 og 4 (se indledningen).
- Plot D app som en funktion af R for at opnå en spredning lov sammenlignelig med, hvad der måles med spot variation baseret FCS 12 (figur 3D).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
For at kalibrere den instrumentale taljen, kan billedet af en enkelt fluorescerende nano-kugle være foranstaltning som beskrevet i protokol trin 1.1. En typisk fluorescerende billede af disse perler er vist i figur 1. Monteringen af intensitet fordeling af en 2D Gaussisk funktion giver tilbage gode rester og tillader måling af instrumentale talje ved 270 nm. Denne værdi er i god overensstemmelse med det forventede diffraktionsgrænsen anslået Rayleigh ligningen. Denne kalibrering er ikke nødvendig for måling af partikel dynamik, men det er nødvendigt at måle den tilsyneladende partikelstørrelse.
En typisk frekvens fordeling af kamera baggrund er vist i figur 2. Toppen ved omkring 180 DL skyldes kameraets reaktion på ingen foton, og det repræsenterer bidraget Analog Digital (AD) konverter. Dette bidrag kan tilnærmes som en Gaussisk fordeling at estimere offset og variansindført ved optagelse af signaler. Over 200 DL det digitale niveau fordeling bliver eksponentiel (lineær i logaritmisk skala), og repræsenterer den gennemsnitlige kamera reaktion på en enkelt foton. Montering denne del med en eksponentiel fordeling tillader målinger af den gennemsnitlige DL tildelt hver enkelt foton. Jo højere forholdet mellem den gennemsnitlige DL tildelt til hver foton, og AD konverter fejl, vil den nederste være støj i den beregnede korrelationsfunktionen. Desuden er den gennemsnitlige enkelt foton reaktion tillader estimeringen af kameraets dynamikområde.
Et diagram af det komplette eksperimentelle procedure er opsummeret i figur 3, og et billede af Atto488-PPE indsættelse i membranen er vist i figur 4A. Et repræsentativt TIRF billede af basalmembranen af en CHO-celler mærket med Atto488-PV er præsenteret i figur 4B. Adskillige lyspunkter kan være til stede uden for cellen på grund af liposomes stablet på glasset. De kan kasseres ved at vælge en ROI på en membrandel hovedsagelig ensartet i fluorescens (dvs.. Det cellulære plasmamembran). Som forventet den målte diffusion lov (figur 4C) for denne lipid er flad, hvilket indikerer en overvejende fri diffusion som tidligere vist ved STED-FCS målinger 30,35. Det er værd at nævne, at alle de viste forskydning værdier ligger under diffraktionsgrænsen med tydelig angivelse af evne til denne tilgang til super-løse Middelmolekylvægtene forskydninger godt under diffraktionsgrænsen og ned til nogle få snese nanometer.
En skematisering af TFR-GFP-dimer indsættelse i membranen er repræsenteret i figur 5A. Mange undersøgelser har vist, at den cytoplasmatiske hale af denne receptor interagerer med skelet membranen, hvilket fungerer som et hegn for receptoren mobilitet 12,40. Et repræsentativt TIRF billede af en CHO-celle udtrykker TFR-GFP er presented i figur 5B. Lav fluorescensintensitets celler bør foretrækkes, da membranen er tættere på den native tilstand og sandsynligheden for artefakter relateret til overekspressionen minimeres. Desuden bør den centrale del af cellen undgås, da virkningerne af ude-af-fokus fluorescens (fra cytoplasmaet, for eksempel) kan være til stede. Som forventet den målte diffusion lov (figur 5C) for TFR-GFP viser en første flad adfærd under 100 nm, med en gennemsnitlig D app omkring 0,7 um 2 sek -1, efterfulgt af efterfølgende hurtige fald i tilsyneladende diffusivitet ned til 0,2 um 2 sek -1 (værdien typisk ved diffraktionsbegrænset FCS 12). Dette resultat viser, at vores fremgangsmåde nemt kan måle den gennemsnitlige forskydning af GFP-mærkede proteiner med en opløsning på nogle få nanometer. Desuden den rumlige skala, hvor D app begynder at falde sæt karakteristiskerumlig skala af protein delvis indespærring af skelet membranen på omkring 120 nm, i overensstemmelse med tidligere skøn 6.

Figur 1. Kalibrering af punktspredningsfunktion. (A) pseudo-billede af en isoleret perle og perler aggregater. (B) 3D-plot af intensiteten profil af en isoleret perle viser en veldefineret gaussiske profil. (C) Fit af intensiteten fordeling med Gaussisk funktion (øverste panel) med de tilsvarende residualerne (nederste del). Den gode aftale mellem monteret fordeling og den målte intensitet profil er også et bevis på, at den instrumentelle PSF kan tilnærmes med en Gaussisk funktion. Klik her for at besà ew en større version af dette tal.

Figur 2. Kalibrering af kamera reaktion på enkelte fotoner. Figuren viser den digitale Niveau (DL) distribution af kamera baggrund i et 32 x 128 ROI, eksponering 0,5 msek i Beskåret Sensor tilstand. Toppen ved omkring 180 DL repræsenterer kameraet reaktion på ingen fotoner. Især det repræsenterer bidrag Analog Digital (AD) konverter og kan tilnærmes med en Gaussisk funktion til at estimere offset og variansen indført af signalet optagelse. Over 200 DL distribution af digitale niveauer bliver eksponentiel og repræsenterer den gennemsnitlige kamera reaktion på en enkelt foton. Målingen af disse parametre giver estimere tæthed af fotoner, der er optaget i løbet af købet.s / ftp_upload / 51994 / 51994fig2highres.jpg "target =" _blank "> Klik her for at se en større version af dette tal.

Figur 3. skematisering af metoden. (A) Bredt felt billeddannelse af EMCCD kamera anvendes til at nå sub-millisekund, mens TIRF mikroskopi udnyttes til at give præcise optiske sektionering af plasmamembranen. (B) Den resulterende stak billeder er autokorrelerede med henblik på at beregne den gennemsnitlige rumlige -temporal korrelationsfunktionen. Denne sammenhæng funktion er godt tilnærmes ved en Gaussisk funktion (se Indledning), og det breder sig i tid i henhold til partikel forskydninger. (C) For således at kvantificere udbredelsen af korrelationen funktion på grund af molekylær forskydning, montering med en Gauss ian funktion udføres. Dette gør det muligt at måle den molekylære "diffusion lov« direkte fra billeddannelse, i form af tilsyneladende diffusivitet vs gennemsnitlige forskydning plot. (D) Takket være dette plot kan molekylær diffusion tilstande identificeres direkte uden behov for en fortolkende model eller antagelser om rumlige organisering af membranen. Faktisk vil frit spredende molekyler vise en konstant tilsyneladende diffusivitet da deres mobilitet ikke afhænger af den rumlige skala af målingen. Derimod vil delvist tillukkede molekyler vise en helt konstant tilsyneladende diffusivitet til forskydninger mindre end indespærring størrelse, så en faldende diffusionsevne for rumlige skalaer større end indespærring størrelse. Således kan forekomsten af en reduktion i den tilsyneladende diffusivitet fortolkes som et fingeraftryk for forbigående indeslutning, mens den tilhørende rumlig skala kan anvendes til at estimere den rumlige udvidelse af fødslen. .jove.com / filer / ftp_upload / 51994 / 51994fig3highres.jpg "target =" _blank "> Klik her for at se en større version af dette tal.

Figur 4. ATTO488-PPE diffusion lov i levende cellemembraner (A) Skematisk fremstilling af ATTO488-PPE indsættelse i cellemembranen (B) TIRF billede af CHO basalmembran mærket med ATTO488-PPE:.. En ROI (rød boks) er valgt i en overvejende ensartet del af cellen, undgå cellekanten og stærkt fluorescerende pletter. (c) loven udbredelse målt i den valgte ROI viser en flad adfærd bekræfter en fri diffusion model for denne komponent. Klik her for at se en større version af dette tal.

Figur 5. TFR-GFP diffusion lov levende cellemembraner (A) Skematisk repræsentation af TFR-GFP indsættelse i cellemembranen:. Den cytoplasmatiske hale af receptoren interagerer med skelettet membran, der fungerer som et hegn til receptor mobilitet (B. ) TIRF billede af CHO udtrykker TFR-GFP: en ROI valgt foretrækker lave udtrykkende celler at undgå artefakter på grund af overekspression (C) Udbredelsen lov TFR (sorte prikker), i modsætning til PPE (grå linie, taget fra figur 4). viser den typiske adfærd delvist begrænset diffusion, hvor en første flade del er efterfulgt af et fald i D-app. Klik her for at se en større version af denne figur.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Enkelt partikel sporing (SPT) repræsenterer en af de mest almindelige strategier for at studere molekylær dynamik, og det har den store fordel at måle partikelbaner. Dette vil til gengæld giver mulighed for sondering adfærd selv få mærkede partikler i et komplekst system. For at nå denne fordel SPT typisk behov for en lav densitet af sonden og meget lyse etiketter. Især for at få høj tidslig opløsning (usek interval) uorganisk sonde kræves normalt (f.eks kvantepunkter eller metal nanopartikler): i dette tilfælde er det nødvendigt med en kompliceret procedure for produktion, mærkning og indføring i systemet. Sammenlignet med SPT den nuværende metode viser nogle af de vigtigste fordele. Først og fremmest kan denne fremgangsmåde anvendes i konjugation med fluorescerende proteiner. I forhold til SPT, er en højere tidsmæssig opløsning nået (på samme etiket) takket være det mindste beløb af fotoner krævede 34. Mere detaljeret denne egenskab tillader skubber den tidsmæssige resolution under 10-3 sek også når de bruger kodes fluorescerende proteiner, og denne tidsplan giver eksklusiv adgang til nanoskala dynamik membran vælgere. Endelig er det værd at bemærke, molekylær diffusion love er beskrevet ved at analysere det fulde rum-tid korrelationsfunktion, uden behov for at spore hvert molekyle.
Sammenligningen med STED-baserede FCS er også interessant. I en STED-FCS måling gennemsnitlige transittid molekyler til faldende observation volumen måles ved den tidsmæssige korrelation af fluorescenssignalet. Dette gør det muligt at opnå en lokal måling af molekylær dynamik også under diffraktionsgrænsen. I den præsenterede metode loven diffusion måles som gennemsnittet af alle de partikler, der bevæger sig i den valgte ROI, observeret ved hjælp af standard, diffraktionsbegrænset, observation volumen. Rapporterede imidlertid viser, at denne metode ikke er begrænset af diffraktion, men kun ved the tidsmæssig opløsning. Faktisk kan selv en diffraktionsbegrænset erhvervelse anvendes til at detektere udsving (analogt til, hvad der er gjort i andre super opløsning teknikker, såsom Palm og STORM), molekylære forskydninger godt under diffraktionsgrænsen være (direkte) beregnet, som allerede påvises ved hjælp af STICS at måle molekylære strømme 32. I modsætning STED-FCS, denne tilgang kan nemt anvendes til en bred vifte af kommercielle og eksisterende mikroskopi opsætninger, såsom raster scanning mikroskoper eller bredt felt kamera-baserede mikroskoper. Det er værd at nævne, at STED-FCS målinger af molekylær diffusion love strengt kræver en fluorofor-afhængig kalibrering af størrelsen af den instrumentale taljen. Omvendt, målingen præsenteres her ikke kræver et system kalibrering (kun nødvendig til vurdering af partikelstørrelse).
Den faktiske opløsning i målingen af partikel forskydninger ved præsenterede metode dehænger af, hvor nøjagtigt vi kan måle korrelationsfunktionen. Derfor er det ikke i sig selv begrænset af diffraktion, analogt med SPT tilfælde hvor opløsningen afhænger af, hvor præcist partikel "billede" måles. For at måle en signifikant sammenhæng i mindre end 1 min for de foreslåede eksperimenter, få fotoner (normalt under 10 fotoner) for hver partikel i hver ramme er nok. Faktisk er bidrag fra alle de observerede partikler gennemsnit sammen, når korrelationsfunktionen er beregnet, selv om partikler, der ikke er isoleret. Denne egenskab er iboende fluktuation korrelations- metoder og tillader brug dim og tætte mærker, såsom fluorescerende proteiner transficeret i levende celler.
Med dette i tankerne fremgår det klart, at den mindste målelige forskydning afhænger diffusiviteten af partiklen og på den tid, opløsning af den billeddannende opsætning. Som et eksempel, skal overveje diffusion af molekyler på cellemembranen,hvor den maksimale målte diffusiviteten for proteiner eller lipider er omkring 5 um 2 sek -1. Under disse betingelser har vi brug for en tidsopløsning på cirka 10 -4 sek at fange en gennemsnitlig forskydning på 50 nm. Denne gang opløsning kan opnås ved hurtig scanning mikroskoper langs enkelte linjer eller ved hurtig EMCCD kamera, hvor tiden opløsning falder til eksponeringstiden, som viste her.
Et yderligere væsentlige krav til denne metode til præcist at beskrive molekylære dynamik er en korrekt rumlig sampling. Faktisk, for at passe korrelationsfunktionen vi har brug for en rumlig prøvetagning (pixelstørrelse) lavere end taljen af instrumentale polyesterfibre. I de fleste kommercielle mikroskoper (konfokal eller bred field), polyesterfibre talje spænder fra 200 nm til 500 nm (hovedsageligt afhængigt af den numeriske apertur for det valgte objektiv og den anvendte bølgelængde) og kan let måles ved en kalibrering eksperiment ved hjælp nano- mellemstore fluorescerende perler. Thus, kan en pixelstørrelse på 70-150 nm (3 gange lavere end den instrumentale taljen) være nok. Imidlertid kan pixelstørrelse tilpasses til systemet under undersøgelse under hensyntagen til en simpel regel: sænke pixelstørrelse højere nøjagtighed i beskrivelsen af korrelationsfunktionen. Desuden minimumsstørrelsen af billedet, der skal erhverves skal være mindst 3 gange større end den maksimale forskydning af interesse (plus den instrumentale talje). Dette er nødvendigt for at nå en god konvergens montering algoritme og en statistisk signifikant udsnit af molekylære forskydninger. Som et eksempel, for at studere middelmolekylvægte forskydninger mindre end få hundrede nanometer (fx 200 nm) en billedstørrelse på få mikrometer er nok. Desuden er det samlede antal pixels (under konstant pixelstørrelsen) virkninger på kvaliteten af korrelationsfunktionen. Faktisk giver et større billede gennemsnit mere information i korrelationsfunktionen, selvom på bekostning af den tid resolution. Med hensyn til kamera-baseret system, der anvendes her, bedes du bemærke, at den fysiske størrelse af pixel på chippen er fast. Følgelig faldende pixelstørrelse sænker signalet i hver pixel (der afhænger af kvadratet af pixelstørrelse) nedsætter synsfeltet og kræver højere forstørrelse. På den anden side, i et scanningssystem, hvor området observation er fast, faldende pixelstørrelsen normalt resulterer i et øget antal pixel på bekostning af tidsopløsning.
Par detaljer om detektoren anvendes, skal drøftes. I modsætning til enkelt-fotondetektorer måler EMCCD systemer en gennemsnitlig intensitet (digitale niveau, DL), der ikke er direkte proportional med det opsamlede lys på grund af tilstedeværelsen af en offset. Selv om dette offset er lavt i forhold til den dynamiske rækkevidde af kameraet (få hundrede sammenlignet med 2 16 i 16 bit udlæsning) og ubetydelig i eksperimenter, hvor mange fotoner indsamlet, skal det tages i betragtning vedopnå en korrekt normalisering af korrelationsfunktionen. Også den offset kan bruges som reference i svagt lys for at identificere mængden af signal indsamles. Desuden har man for at vurdere den gennemsnitlige mængde af fotoner, der er indsamlet i løbet af købet den gennemsnitlige digitale niveau er forbundet til hver indsamlet foton skal måles. Denne mængde kan hentes ved at udsætte kameraet til en meget lav lysintensitet (fx baggrunden lys i rummet); i virkeligheden, i dette tilfælde, kan vi med rimelighed antage, at bare enkelte fotoner nå kameraet, dvs den målte intensitet kan relateres til kun nul eller én foton.
Endelig lad os kommentere på, hvordan nogle alternative indkøbssystemer (dvs. forskellige mikroskopi opsætninger) kan bruges til at udføre de præsenterede målinger. Først og fremmest kan den "W'factor i ligning 2 (der repræsenterer autokorrelation instrumentale PSF) tilpasses til than især køb, der anvendes for at passe den eksperimentelle korrelationsfunktionen. Som tidligere vist 34, en let sag erhvervelse Whit et laserscanningmikroskop når scanningshastigheden er betydeligt højere end partikel dynamik. I et sådant tilfælde, i virkeligheden, flytning af partiklerne under købet tid (dvs. linie tid) kan betragtes som ubetydelig, og korrelationsfunktionen er godt tilnærmes ved en Gaussisk funktion. I forbindelse med de nye imaging teknologier, er en interessant tilgang baseret på muligheden for at producere meget tynde lys ark (1-2 um) gennem prøven 41. Lyset ark muliggør selektiv belysning af et enkelt plan (Single plan belysning mikroskopi, SPIM) i prøven, og kombineret med et kamera-baserede erhvervelse system hurtig optisk sektionering i 3D 42. På grund af disse egenskaber, er SPIM blevet konjugeret med FCS 43 og kunne udgøre en valid værktøj til at forlænge den præsenterede analyse 3D miljøer, såsom cytoplasmaet eller kernen i levende celler.
Sammenfattende fra en eksperimentel synspunkt denne fremgangsmåde kræver kun adgang til et mikroskop udstyret med en hurtig erhvervelse modul. Proteinet af interesse kan mærkes med noget fluorescerende protein eller organiske fluorofor, således at også flerfarvet billeddannelse. I denne forbindelse forestiller vi mulighed for at bruge på tværs i MSD-analyse for at vælge sub-populationer af molekyler og afslører interaktioner og samarbejde udbredelse af levende cellemembraner. Endelig mener vi, at denne tilgang kan udgøre et kraftfuldt værktøj til at studere proteiner og / eller lipider undergår dynamisk opdeling inden nanodomains på plasmamembranen. I dette tilfælde den meget forskellig størrelse og levetid nanodomains indføre et ekstra niveau af i det virkelige data kompleksitet, som vil kræve yderligere metodologiske implementeringer, herunder 2-farvebilledbehandlingsområdet, lokalanalyse (f.eks 2D par korrelation) og / eller fluorescensanisotropi.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}