

Abstract
Bu uzaysal dağılımı ve lipit ve protein gibi zar bileşenlerinin hareketi pek çok hücresel fonksiyonlarının düzenlenmesinde önemli bir etken olduğu giderek daha belirgin hale gelmiştir. Bununla birlikte, hızlı dinamikleri ve ilgili küçük yapılar için, çok yüksek uzay-zaman çözünürlüklü moleküllerin gerçek davranışlarını yakalamak için gereklidir. Burada yüksek uzaysal çözünürlüklü Canlı hücrelerdeki floresan etiketli plazma zar proteinleri ve lipidler dinamiğini çalışmak için deneysel protokol mevcut. Özellikle, bu yaklaşım, her bir molekülün izlendiği gerek yoktur, fakat zarın belirli bir bölgedeki tüm molekülleri kullanılarak nüfus davranışı hesaplar. Başlangıç noktası, zar üzerinde belirli bir bölgenin hızlı görüntülenmesidir. Daha sonra, tam bir uzay-zaman otomatik bağıntı kurma fonksiyonu, örneğin, her biri 2, 3, n, tekrarları zaman gecikmeleri arttırmayı alınan görüntüleri ilişkilendirilmesi hesaplanır. Bu genişlik göstermek mümkündürdifüzyona bağlı parçacık hareketinin bir fonksiyonu olarak zaman gecikmesini arttırmayı mekansal otokorelasyon fonksiyonu artar pik. Bu nedenle, otokorelasyon fonksiyonların dizi montaj gerçek protein burada ortalama yer değiştirme karşı belirgin yayılma biçiminde, görüntüleme (iMSD) için ortalama kare deplasman elde etmek için olanak sağlar. Bu nanometre hassasiyetle tek moleküllerin ortalama dinamiklerinin kantitatif bir görünüm verir. Etiketli Transferin Reseptör (TfR) ve ATTO488 bir GFP etiketli varyantı kullanarak 1 Palmitoyl- 2-hidroksi sn -glisero-3-fosfoetanolamin (KKE) o üzerinde protein ve lipit difüzyon Spatiotemporal düzenlenmesini gözlemlemek mümkündür mikro kadar mili saniyelik bir zaman aralığında um boyutlu zar bölgeleri.
Introduction
Singer ve Nicolson tarafından orijinal "sıvı mozaik" modeli başlayarak, hücre plazma zarı resim sürekli iskeleti ve lipit etki 1,2 yükselen rolünü kapsayacak şekilde son yıllarda güncellendi.
İlk gözlemler membran proteinlerinin önemli bir kısmı 3-5 hareketsiz olduğunu (sıkı bağlamak) açıklanması photobleaching sonra floresan iyileşme elde edilmiştir. Bu öncü çalışmalar, çok bilgilendirici olmasına rağmen, sıkı bağlamak kurulumları nispeten zayıf alan (mikron) olarak çözünürlük ve zaman (saniye) muzdarip. Ayrıca, bir topluluk ortalama ölçümü olmak, sıkı bağlamak tek molekül davranışı hakkında bilgi veren yoksun.
Bu bağlamda, spesifik olarak bir olasılık (aynı anda bir difüzyon işlemi bir molekülün çalışma sağlayan) çok parlak etiketleri ile tek bir molekül etiketlemek için oldukça başarılı olmuştur. Özellikle, iterekve mikrosaniye zaman çizelgesi, Kusumi, büyük ölçüde membran fizyolojisi 6 aktin esaslı membran iskeletinin rolünün tanınması katkıda lipid ve protein dinamikleri bilinmeyen özellikleri ark. kazandı erişim için Tek Parçacık Takibi (SPT) yaklaşımı zaman çözünürlüğü , 7. Bu bulgular, lipid ve protein difüzyon aktin bazlı bir iskelet tarafından ayarlandığı, "kazık ve çit" modeli olarak adlandırılan oluşturulur. Ancak, SPT birçok deneysel konularda sağlanan bilgilerin büyük miktarda erişimi için ele alınması gerekir. Özellikle, etiketleme prosedürü genellikle sisteme etiketli türlerin üretimi, saflaştırılması ve tanıtım gibi birçok adımlarla oluşur. Ayrıca, büyük bir etiket, kuantum noktaları veya metal nanopartiküller gibi, çoğu zaman alt-milisaniye süreler ve bir çok durumda önlenebilir edilemeyen etiket ile hedef moleküllerinin çapraz bağlanmasını ulaşmak için gereklidir. Son olarak, birçok yörüngelerİstatistiksel kriterlerine uygun kaydedilecek olan ve eş zamanlı olarak etiketin bir düşük yoğunluklu izleme izin vermek için gereklidir.
SPT ile karşılaştırıldığında, bu sakıncaları birçok üstesinden flüoresan korelasyon spektroskopisi (FCS), moleküler dinamiklerini incelemek için çok umut verici bir yaklaşımı temsil etmektedir. Aslında, FCS geçici transfekte hücrelerinde floresan protein etiketli moleküllerinin dinamiklerini incelemek sağlayan, loş ve yoğun etiketleri ile iyi de çalışıyor. Ayrıca, zaman sınırlı bir miktarda yüksek istatistikleri ulaşan verir. Son olarak, etiket "yüksek" yoğunluğuna rağmen FCS tek molekülleri bilgi sağlar. Tüm teşekkürler, bu özelliklere FCS çok düz bir yaklaşımı temsil etmektedir ve yoğun bir model membranlar ve canlı hücreler 8-10 hem lipid ve protein dinamikleri üzerinde çalışmak için uygulanmıştır. Birçok farklı yaklaşımlar moleküler difüzyon ayrıntılarını ortaya çıkarmak için FCS yeteneğini artırmak için önerilmiştir. Örneğin, sh wasKendi farklı büyüklükteki gözlem alanları FCS gerçekleştirerek bir molekül hareketlerinin 11,12 bir "FCS difüzyon yasası" aydınlatıcı gizli özellikleri tanımlayabilirsiniz. Boyutu değişmiştir olmanın yanı sıra, odak alanı da, 13 yinelenmiş oldu hızlı kameralarla 21,22 hatlarda 14-20 veya konjuge boyunca uzayda taşındı. Bu 'uzay-zamansal' korelasyon yaklaşımlar kullanılarak, çeşitli membran bileşenlerinin ilgili biyolojik parametreler nicel modeli membranlar ve gerçek biyolojik olanlar, membran mekansal organizasyon haline tutulmakta ve böylece içgörü hem tarif edildi.
Ancak, tüm FRAP ve FCS uygulamalarda şimdiye kadar tarif edilen fokal alanının boyutu aşılamayan uzamsal çözünürlüklü bir sınırı temsil eder. Çeşitli süper çözünürlüklü görüntüleme yöntemleri son zamanlarda bu sınırı aşmak için geliştirilmiştir. Bazıları böyle Stokastik optik yeniden mikroskobu (STORM) gibi, yerelleştirme hassas dayanmaktadır <sup> 23,24, foto-aktifleştirme lokalizasyon mikroskobu (PALM) 25, flöresanlı PALM (FPALM) 26 ve tek parçacık izleme PALM (sptPALM) 27: her anlık gerekli foton nispeten büyük miktarda bununla birlikte, zaman çözünürlüğü sınırlar En az birkaç milisaniye için bu yöntemler, bu nedenle in vivo olarak uygulanabilirliğini engellemektedir.
Buna karşılık, süper çözünürlük görüntüleme için umut verici bir alternatif mekansal uyarılmış emisyon tükenmesi yöntemleri (STED'in veya geri dönüşümlü doyabilen optik floresan geçişler (RESOLFT)) 28,29 ile floresan emisyonu modüle tarafından açılmıştır. Bu yaklaşımlar hızlı tarama mikroskopları ve algılama sistemleri kullanmaya imkanı ile gözlem hacminin şekillenmesini çok altında kırılma sınırını birleştirir. Flüoresan iniş-çıkışlarını analizi ile birlikte, doğrudan mikroskopi STED'in lipidler ve p nano uzaysal dinamikleri prob bırakıldıcanlı hücre membranları içinde 30,31 roteinler.
STED'in bazlı mikroskopi aynı fiziksel miktarlar Canlı hücrelerdeki floresan etiketli zar proteinleri ve / veya lipidler dinamiklerinin çalışma için uygun modifiye edilmiş bir uzay-zaman görüntü korelasyon spektroskopisi (H TATİSTİKLERİ 32,33) yöntemi ile elde edilebilir ve ticari bir mikroskop ile. Burada sunulan deneysel protokol birkaç adımda oluşur. Birincisi, zar üzerinde ilgili bölgenin hızlı bir görüntüleme gerektirir. Daha sonra, elde edilen görüntülerin yığın ortalama uzaysal ve zamansal ilişki işlevleri hesaplamak için kullanılır. Bağıntı fonksiyonlarının bir dizi montaj olarak, molekül "difüzyon kanunu 'belirgin bir difüzyon (D uygulaması) şeklinde görüntüleme şirketinden elde edilebilir - -Ortalama değiştirme grafiğinin. Bu komplo eleştirel molekülleri tarafından keşfedilmeyi ortama bağlıdır ve doğrudan gerçek difüzyon modları tanıma sağlarilgilenilen lipit / protein elde edilir.
Daha önce 34 görüldüğü gibi ayrıntılı olarak, elde edilen görüntü serisi uzay-zamansal otomatik korelasyon fonksiyonu eleştirel toplanan resim serisi hareketli moleküllerin dinamiklerine bağlıdır (Aynı mantık bir hat satın uygulanabilir lütfen unutmayın Uzayda sadece tek boyut) olarak kabul edilir burada. Özellikle, korelasyon fonksiyonu olarak tanımlar:
 (1)
(1)
nerede  , x konumuna, ölçülen floresans yoğunluğu temsil etmektedir, y-ve t de,
, x konumuna, ölçülen floresans yoğunluğu temsil etmektedir, y-ve t de, ![]() ve
ve ![]() temsil x mesafe vesırasıyla y yönlerinde,
temsil x mesafe vesırasıyla y yönlerinde, ![]() zaman aralığını temsil eder ve
zaman aralığını temsil eder ve ![]() ortalamasını temsil eder. Bu fonksiyon, şu şekilde ifade edilebilir:
ortalamasını temsil eder. Bu fonksiyon, şu şekilde ifade edilebilir:
 (2)
(2)
'N', gözlem alanında moleküllerin ortalama sayısı temsil ettiği, ![]() Uzayda büklüm operasyon temsil eder ve
Uzayda büklüm operasyon temsil eder ve  enstrümantal bel otokorelasyonu temsil eder. Bu ikinci durum optik / kayıt kurulum için tek bir emitörün fotonlar uzayda yayılır nasıl bir önlem olarak yorumlanabilir (sözde Noktası Yayılması Fonksiyonu, PSF, geniyi) bir Gauss fonksiyonu tarafından yaklaşık ralli. Son olarak,
enstrümantal bel otokorelasyonu temsil eder. Bu ikinci durum optik / kayıt kurulum için tek bir emitörün fotonlar uzayda yayılır nasıl bir önlem olarak yorumlanabilir (sözde Noktası Yayılması Fonksiyonu, PSF, geniyi) bir Gauss fonksiyonu tarafından yaklaşık ralli. Son olarak,  bir mesafede bir parçacık bulmak için olasılığını temsil eder
bir mesafede bir parçacık bulmak için olasılığını temsil eder ![]() ve
ve ![]() bir süre geçtikten sonra
bir süre geçtikten sonra ![]() . Biz parçacıkların her yöne rastgele hareket ve net tozları mevcut olmayan hangi bir uzadıya dinamikleri, düşünürsek, bu fonksiyon da iyi varyans hareketli parçacığın ortalama Meydanı Tehcirine (MSD) olarak tespit edilebilir bir Gauss fonksiyonu ile yaklaşılır . Bu durumda, korelasyon fonksiyonunun bel (ayrıca şu şekilde de ifade
. Biz parçacıkların her yöne rastgele hareket ve net tozları mevcut olmayan hangi bir uzadıya dinamikleri, düşünürsek, bu fonksiyon da iyi varyans hareketli parçacığın ortalama Meydanı Tehcirine (MSD) olarak tespit edilebilir bir Gauss fonksiyonu ile yaklaşılır . Bu durumda, korelasyon fonksiyonunun bel (ayrıca şu şekilde de ifade ![]() ), Partikül İskelet toplamı ve enstrümantal bel olarak tanımlanabilir ve bir Gauss uyum ile ölçülebilirHer bir zaman gecikmesi için bağıntı fonksiyonunun anyonlar olabilecektir. Ölçülen ı MSD hareket eden moleküllerin belirgin bir yayılma hesaplamak için kullanılabilir
), Partikül İskelet toplamı ve enstrümantal bel olarak tanımlanabilir ve bir Gauss uyum ile ölçülebilirHer bir zaman gecikmesi için bağıntı fonksiyonunun anyonlar olabilecektir. Ölçülen ı MSD hareket eden moleküllerin belirgin bir yayılma hesaplamak için kullanılabilir ![]() ve bir ortalama yer değiştirme
ve bir ortalama yer değiştirme ![]() gibi:
gibi:
 (3)
(3)
 (4)
(4)
Aşağıdaki bölümlerde boyunca okuyucuya rehberlik Kullanılan deney düzeneği üzerinde birkaç hususlar. Seçici ticari bir TIR floresan kullanarak, bir toplam iç yansıma (TIR) aydınlatma kullanacak yaşayan hücrelerin bazal membran üzerinde florosforlar heyecanlandırmak için (TIRF) mikroskop (detaylar malzeme bölümünde bulunabilir). Ayrıca, sırayla th toplamakE floresan bir yüksek büyütmeli objektif kullanın ve (çip 16 mikron üzerinde piksel fiziksel boyutu) bir EMCCD kamera (100X NA 1.47, yüksek sayısal açıklık TIRF aydınlatma için gereklidir). 100 nm piksel boyutu ulaşmak için biz 1.6x ek bir büyütme merceği geçerlidir. Aşağıda tartışıldığı gibi, 1 ms altında bir zaman çözünürlüğü doğru 100 nm altında hızlı bir membran lipidlerinin dinamiklerini tanımlamak için gerekli olacaktır. Bu zamansal çözünürlüğe ulaşmak için biz kamera (512 x 512) bütün çip daha küçük faiz (ROI) bir bölgeyi seçmeniz gerekir. Bu şekilde, kamera zaman çözünürlüğü artırmak satır, daha az sayıda okuyacaktır. Ancak, bu okuma rejimde çerçeve zaman kamera üzerinde okuma çip maruz ücretleri değişiklikleri için gerekli ve 512 x 512 piksel EMCCD için milisaniye sırayla genellikle zaman ile sınırlı olacaktır. Bu sınırı yenmek için, gelişmekte olan bir teknoloji w, ROI-hatları, sadece yerine tüm çerçeveyi edilebiliyor(bizim EMCCD kırpılmış Sensör Modu denir) maruz çip boyutu bir pratik ve etkili azalma ITH. Bu yapılandırma, etkili olması için, dış ROI çip, optik bir yoldaki monte edilmiş delikli bir çift ile örtülmesi gerekir. Bir zaman çözünürlüğü aşağı 10 -4 saniye bu kurulum sayesinde elde edilebilir. 'Tartışma' bölümünde açıklandığı gibi bu yaklaşım, birçok farklı deney düzeneklerinin ile birleştiğinde olabilir, ancak unutmayın.
Yöntemin Gösterme GFP- bir ATTO488 etiketli hem 1-palmitoil-2-hidroksi-SN -glisero-3-fosfoetanolamin (ATTO488-PPE) ve transferrin alıcı bir GFP-etiketli varyantı kullanarak (canlı hücrelerde sağlanacaktır TfR). Daha önce 30,35 bildirildiği gibi ATTO488-KKD'nin durumunda bu yaklaşım başarılı bir çoğunlukla ücretsiz difüzyon gösteren ortalama değiştirmesinin bir fonksiyonu olarak neredeyse sabit bir D uygulaması kurtarabilirsiniz. Buna karşılık, TfR-GFP azalan D göstermektedir
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Sistem Kalibrasyon
- Nokta yayılma fonksiyonu (PSF) kalibrasyon
- Damıtılmış su içinde 90 ul, 30 ile floresan boncuk çözeltisi (yaklaşık 5 uM), 10 ul seyreltilir ve daha sonra 20 dakika boyunca ultrasonik titremeye maruz bırakılır çözeltisi. Bir kare (1 cm x 1 cm) agaroz jeli (% 3) ve depozit jel üstündeki çözeltisinin 10 ul parça kesin. 2 cm Petri kabı alt camına jel parçası devirmek ve camına damla sıkın.
- , Satın alma kurulum açın tutucuya örnek koymak, kamera pozlama ve EMgain set (100 msn ve 1000 sistemine göre iyi parametrelerdir ama optimize) ve kamera soğumasını bekleyin.
- 100 milisaniye, 1,000 kamera EMgain, edinim moduna ayarlayın kamera pozlama ayarı kaydetmek Transferi, 100 tekrarı ve otomatik Frame.
- Jelin sınırında mercek ve iletilen ışık odaklama kullanılarak ve daha sonra, jel merkezine amacı hareket odaklanma ve sta ayarlamaklazer hizalama prosedürü rt (LAS AF, 'TIRF kurulum' seçin ve otomatik hizalama prosedürünü izleyin).
- Bir referans olarak (ki genellikle boncuk bütünü temsil) parlak noktaya odaklanmak doğru, izole tek noktalar ile bir görüş alanı bulun 100 kare kazanmak ve birkaç tek noktalar kazanmak için adımı 5-6 kez tekrarlayın.
- Bir veri işleme programına edinilen serisi İthalat ve zaman içinde (Şekil 1A) yığını ortalama ve tek bir izole boncuk seçin. Parçacık agrega önlemek için küçük olanları seçmek için özen gösterin.
- (Matlab Malzemelerinde ICS-Matlab araçları) komutu "gaussfit" kullanarak bir Gauss fonksiyonu ile seçilen yoğunluk dağılımı (tek boncuk profilinin bir örneğini Şekil 1B sunulmuştur) takınız. Elde edilen artıklar inceleyerek uyum iyiliğini doğrulayın (gelen artıklar ile donatılmış Gauss profilinin bir örneği i sunulmuşturn Şekil 1B).
- Kamera kalibrasyonu
- Kamerayı açın ve kamera soğumasını bekleyin. Set kamera alma ayarı, (yani, kullanılan kamera için biz 0.5 msn, 1.000 kamera EMgain, Kesilmiş Modu, 32 x 128, 10.000 tekrarlar için ROI boyutu edinim moduna pozu ayarlamak) ve kamera arka alımı başlatabilirsiniz sinyali.
- İthalat bir veri işleme programı çerçeve serisi satın aldı. Hesaplayın ve kamera arka çipin seçilen bölgede yaklaşık düz olduğunu doğrulamak için her pikselin ortalama yoğunluğunu kontrol edin. Kamera plan genellikle sınır hatlarında önyargılı çünkü kırpılmış modunda, her çerçeve için ilk ve (ROI boyutuna bağlı olarak 3-10) son birkaç yatay satırları kaldırın.
- Değerlerinin histogramı oluşturun (Matlab komut 'hist' kullanarak) yığını elde edilen görüntü (aynı zamanda Dijital Seviye DL tanımlanan) ve logaritmasını arsafrekansını sonuçtaki (Matlab semiloloji komutunu kullanarak). Kamera arka DL dağılımının bir örneği, Şekil 2'de gösterilmiştir.
NOT: kamera iyi çalışıyorsa, komplo üslü (log ölçekli negatif eğimli bir çizgi izledi sıfır foton ilişkili değerlerin dağılımını temsil eden yaklaşık Gauss tepe (log-ölçekte bir parabolik profili) gösterecektir ) foton 1 (Şekil 2) ile ilişkili değerlerinin dağılımını temsil eder. Üstel kısmının çürümesi sürekli tek tek her foton kamera tarafından atanan DL bir tahmin temsil ederken, özellikle merkezi ve Gauss fonksiyonunun varyans, sırasıyla, ofset kamera ve hata temsil eder. Matlab malzemeleri destekleyen Script bölümünde "CalibrateCamera" kullanın. - Tüm seçilen Kamera EMGain ve Kazancı için işlemi tekrarlayın.
2. EtiketliHücre Hazırlanması
- (1,2-dioleoyl- SN -glisero-3-fosfoetanolamin), lipid dahil 36 için gerekli olan lipozomları hazırlamak DOPE ayrı ayrı olarak 1 mg giderilmesi, DOTAP (1,2-dioleoil-3-trimetilamonyum propan-), 1 mg, ve kloroform, 1 ml PPE ATTO488 1 mg. 24 saat boyunca vakum altında bir araya 0,5 DOPE solüsyonu, DOTAP çözeltiden 0.5 ml ve PPE ATTO488 çözeltisi 25 ul kuru karıştırılır. 40 ° C'de 15 dakika boyunca ultrasonik titremeye maruz bırakılır ve 15 dakika boyunca 20 mM HEPES tamponu, 0.5 ml, vorteksleyin.
- , Hücre hazırlanması PBS ile birleşik CHO-K1 (Çin Hamsteri Yumurtalığı) bir p 100 tabak 3 kez yıkama, 5 dakika boyunca inkübatör içinde tripsin ve mağaza 1 ml ekleyin. % 10 FBS ile desteklenmiş DMEM / F12 ortamında 9 ml ilave müstakil hücrelerin süspansiyonu ve aynı ortamın 800 ul ihtiva eden bir Petri kabındaki hücre çözeltisinin 150 ul tohum.
- 37 ° C'de 24 saat boyunca kuluçka makinesi içinde depolayın ve% 5 CO2. Lipid eklenmesi için, 500 hücre orta yerineserumsuz ortam ul; 30 dakika sonra, lipozomlar çözeltisi 2 ul; ve PSB 15 dakika yıkamadan sonra görüntüleme için yeni DMEM / F12 orta ekleyin.
- Transfaksiyon için, transfect hücreler görüntüleme önce inkübatör TfR-GFP plazmid ve mağaza 24 saat kullanarak Lipofektamin protokolü (Üretici talimatları) göre.
3. Veri Toplama
- Kurulum hazırlığı
- Mikroskop termostat amacıyla, deneyden önce 24 saat kuluçka açın.
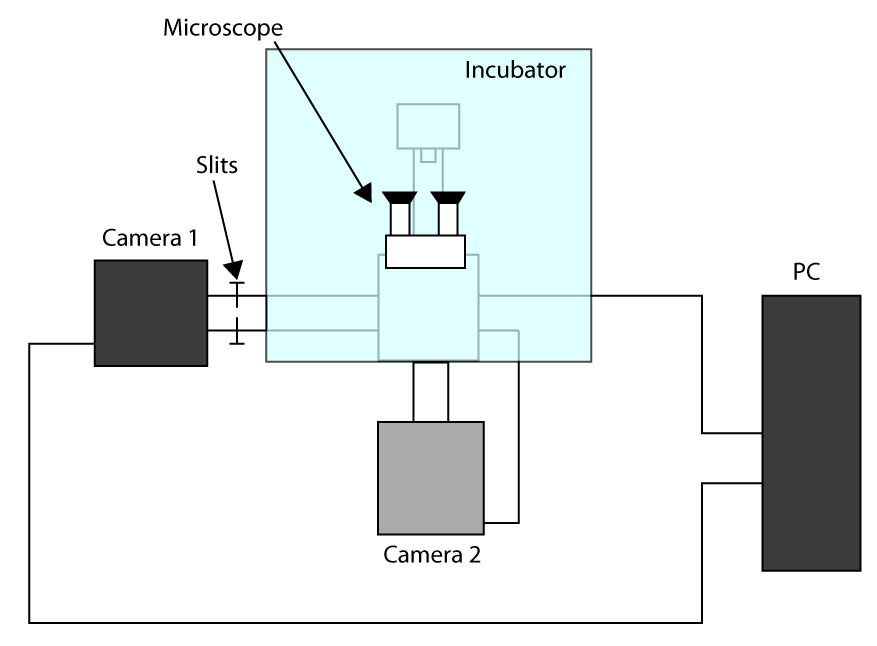
- Hızlı ulaşılabilir edinimi süresi uygulamak amacıyla, Kesilmiş Sensör Modu iş (giriş) ve hücre (kamera 2) seçmek için görüntüleme (kamera 1) ve ikinci kamera için bir ilk kamera kullanmak. Kurulum yapılandırmasına bir şeması Ek Şekil S1 sunulmuştur. Ardından, mikroskop iki kamera dönüşü uyum ve kameralar soğumasını bekleyin.
- Iki kamera iletilen ışık görüntüleme parametreleri ayarlamak (yani
- Sadece (burada 32 x 32 piksel ROI) hücre görüntüleme için kullanılan ROI ışık sağlayan yarıklar hafifçe itin tutucu örnekleri koyun ve mercek kullanarak odak, fotoğraf makinesi 1 ışık göndermek ve.
- Seçilen bölgedeki bir hücreyi taşımak ve kamera 2 ışık göndermek, sonra bir başvuru olması için kamerayı 2. kontrol yazılımı bir ROI çizin.
- Görüntüleme (Şekil 3A)
- Öncelikle, sizin kurulum prosedürüne göre TIRF lazer hizalama. Bizim kurulumunda, 'TIRF kurulum' seçin ve otomatik hizalama işlemini başlatmak. Lazer hizalandığında (yaklaşık 70 °) penetrasyon derinliği 70 nm ayarlayın.
- Hem kamera 1 ve kamera 2 100 EMGain 70 msn ve pozlama süresi ayarlayın; Daha sonra, kamera 1 kullanarak bir hücreyi seçin, ardından kamera 2 ışık göndermek ve doğru hücre zarı odaklanın. Ca üzerinde asgari pozu ayarlamakmera 2, 1000 EMGain, Kesilmiş Sensör Modu, 10 5 tekrar ve set kaydetme dosyaları (Esnek Görüntü Nakil Sistemi, kolayca yönetilen bir biçimi) uygun olarak.
- Görüntü serilerini kaydetmek için satın alma başlayın. Kazanç ve sonra, yeni bir hücre almadan önce sıcaklık stabilizasyonu sağlamak 8-10 hücrelerini elde etmek için son iki adımları tekrarlayın kırpılmış Modu bırakın.
Imaging'e Mean Square Yerinden 4. Hesaplama (i MSD)
Not: Aşağıdaki protokol, doğrudan ham verilere uygulanabilir. Aynı zamanda, tüm protokol Matlab ve SimFCS hem de simüle edilmiş veri satın alma için de geçerlidir. Gelen öğreticiler için bağlantı 'Malzeme' bölümünde bulunabilir.
- Matlab ile Hesaplama
- ImportImageSeries script kullanarak Matlab'a edinilen serisi aktarın. Co kullanarak zaman içinde her görüntünün ortalama yoğunluğunu hesaplamakmmand ilk 2 boyutlarına ortalama ve elde edilen vektör görmek için arsa kullanın.
- Photobleaching% 10'dan fazla varsa, dizi atmak veya onların ilk kısmını kaldırın. Daha düşük ise, 37 önce gösterildiği gibi, her bir görüntü için ortalama yoğunluğunu çıkarılarak korelasyon fonksiyonu üzerinde etkisini düzeltmek için çalışın.
- Üçüncü boyut ortalamasını kullanarak her pikselin ortalama yoğunluğunu hesaplamak ve çıkan görüntü görüyorum.
NOT: Özellikle dikkat artifaktüel korelasyon önlemek için gereklidir. Aslında, daha önce benzer teknikler 38, hücre sınırları yanı sıra odaklama veziküllerin üzerinden gösterildiği gibi bir güçlü bir korelasyon getirebilir. Ortalama görüntünün muayene hücre sınırları veya odak kesecikler dışında ortaya çıkarsa, aksi takdirde satın atmak dahil bölgeyi dışlamaya çalışıyorlar. Bu hareketsiz yapıların etkisini düzeltmek için her piksele 39 dan ortalama zamansal yoğunluğunu çıkarmak. - T hesaplayınişlevini CalculateSTICScorrfunc kullanarak o zamanmekansal korelasyon (G (ξ, χ, τ)). G (ξ, χ, 0) nedeniyle, düşük ışık rejimi atış gürültü korelasyon G hakim çünkü (0,0,0) sökün; nedeniyle detektöre korelasyon ölçülen bel artırarak τ = 0 g (ξ, χ, τ) deforme olabilir, bu süre boyunca G (± 1,0,0), ve parçacık hareketi hakimdir (bu etki, τ kaybolur > 0) 34.
- Daha sonra Malzeme desteklenmesinde "LogBinStack" fonksiyonunu kullanarak gürültüyü azaltmak ve bir logaritmik zaman-bin kullanılarak Ortalama G (ξ, χ, τ> 0) (ξ, χ, τ) fonksiyonu kullanılarak "gaussfit doğan" G sığdırmak Malzemelerinde ICS-Matlab araçları i MSD (çıkan dizinin ikinci sütun) kurtarmak için.
- Zamanın bir fonksiyonu olarak elde edilen bel σ (τ) 2, (i MSD) çizilir. Veri çok gürültülü ise, Edinim sayısını artırmak için çalışınIRED çerçeveler, birlikte lazer gücünü, ortalama daha fazla G (ξ, χ, τ) artırmak.
- SimFCS tarafından Hesaplama
- ImageJ BioFormat ithalatçı eklentisi kullanarak edinilen dosyalarını açın ve TIFF dizisi olarak iktisap dizi kaydedebilirsiniz.
- Aç SimFCS seçin RICS aracı seçin ve Dosya> İçe Çoklu Görüntüler (Ek Şekil S2).
- Uygun pencereyi (Ek Şekil S3), Fit seçin doğru toplama parametreleri yerleştirin ve kapatın.
- Ekran> Ortalama Yoğunluk> CH1 seçin ve photobleaching varlığını (Ek Şekil S4) doğrulayın.
- Photobleaching% 10'dan fazla mevcut ATIN serisi ise veya mümkün tekrar yük serisinin ilk bölümünü kaldırarak görüntü dizisi ise.
- Ağartılması daha düşük% 10 Araçlar> Eğer i MSD> Set Parametreleri, sol bir on ROI panelinde set, 'kullan hareketli ortalama' kontrolmuhabir zaman karakteristik difüzyon zaman daha yüksek olduğunu hareketli ortalama ödeme İlginiz için çerçeve koyu kahverengi (1 mikron 2 sn hareketli parçacık için -1 10 saniyelik bir zaman iyi bir hareketli ortalama)
- Araçlar> iMSD> i MSD (Ek Şekil S5) ve zinde hesaplayın ve bloknot gelen i MSD ihracat (Ek Şekil S6).
I MSD gelen Difüzyon Kanunu 5. Hesaplama
- Kesişim (σ 0 2) hesaplamak için ilk birkaç puan Fit (5 puan genelde yeterli olur ama daha fazla puan onlar doğrusal bir davranış gösterir eğer takılabilir) ve daha önce ölçülen PSF 2 ile bu değeri karşılaştırın. Onlar karşılaştırılabilir iseniz, izole Flüoroforlann dinamikleri takip edilmektedir. Buna karşılık, σ 0 2 >> PSF 2 denemek sağlamak için daha hızlı elde etmek için isehiçbir gizli dinamikleri 34 mevcuttur.
- Belirgin yayılımını (D app) denklem ve 3 ve 4 (bkz) kullanılarak, ortalama yer değiştirme (R) hesaplayın.
- 12 FCS (Şekil 3D) göre yer değişimi ölçülür ile karşılaştırılabilir bir difüzyon kanunu elde etmek üzere Rl bir fonksiyonu olarak Arsa M app.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Protokol aşama 1.1 'de tarif edildiği gibi etkili bel kalibre etmek için, tek bir floresan nano boncuk görüntü ölçü olabilir. Bu tür boncukların tipik floresan görüntü, Şekil 1 'de sunulmuştur. 2B Gauss fonksiyonuyla yoğunluğu dağılımının uygun olarak geri artıkları verir ve 270 nm'de ölçülmesi etkili bel verir. Bu değer Rayleigh denklemi tahmin beklenen kırılma sınırı ile iyi bir anlaşma olduğunu. Bu kalibrasyon parçacık dinamiği ölçümü için gerekli değildir, ancak bu durum, görünür parçacık boyutunu ölçmek için gereklidir.
Kamera arka tipik bir frekans dağılımı yaklaşık 180 DL tepe noktasının herhangi bir foton için kamera tepkisine bağlıdır. Şekil 2'de sunulmuştur ve bu Analog Sayısal (AD) çevirici katkısını temsil etmektedir. Bu katkı ofset tahmin etmek için bir Gauss dağılımı ve varyans gibi tahmin edilebilirSinyal kaydı ile tanıtıldı. 200 DL Yukarıda dijital seviye dağılımı üstel (logaritmik ölçekte doğrusal) olur ve tek bir foton ortalama kamera yanıttır. Üstel dağılımı ile bu kısmını uydurma, tek tek her bir fotonun atanan ortalama DL ölçümler sağlar. Daha yüksek her foton atanan ortalama DL ve AD dönüştürücü hata oranıdır alt hesaplanan korelasyon fonksiyonu olarak gürültü olacaktır. Ayrıca, ortalama tek foton yanıt kamera dinamik aralık tahminini sağlar.
Bütün deney bir usul diyagramı Şekil 3'te özetlenmiştir ve zar içine Atto488-PPE ekleme bir resmi Şekil 4A'da temsil edilmektedir. Atto488-PPE ile etiketlenmiş bir CHO hücrelerinin bazal membranın bir temsilcisi TIRF görüntü Şekil 4B'de sunulmaktadır. Birkaç parlak noktalar li nedeniyle hücre dışında mevcut olabilmektedirposomes camın üzerine yığılmış. Bir membran kısmı üzerinde floresan çoğunlukla muntazam bir ROI seçilerek iptal edilebilir (yani., Hücresel plazma membran). Beklendiği gibi, bu lipid için ölçülen difüzyon kanunu (Şekil 4C), daha önce STED'in-FCS ölçümleri 30,35 ile gösterildiği gibi, bir çok serbest dolaşımını gösterir düzdür. Açıkça bu yaklaşımın yeteneğini gösteren, tüm gösterilen deplasman değerlerinin kırılma sınırının altında olduğunu belirtmekte yarar var nanometre birkaç on sıra kırılma sınırının altında ve aşağı ortalama moleküler deplasmanları süper-gidermek.
Zarına TfR GFP dimer takılmasının Şemalaştınldığmdan Şekil 5A'da gösterilmiştir. Birçok çalışma, bu reseptörün sitoplazmik kuyruğunun, bu da alıcı hareketlilik 12,40 bir çit olarak hareket eden membran iskelet ile etkileşim olduğunu göstermiştir. TfR-GFP'yi bir CHO hücre temsilcisi TIRF görüntü prese olduğunuŞekil 5B'de nted. Membran, doğal durumu ve en aza aşırı ifadesi ile ilgili eserleri olasılıklarına yakın olduğu gibi floresan yoğunluğu düşük hücreler tercih edilmelidir. Buna ek olarak, hücrenin merkezi kısmı mevcut olabilir (sitoplazmadan, örneğin) ayarsız floresans etkileri olarak kaçınılmalıdır. Beklendiği gibi, ölçülen difüzyon kanunu TfR-GFP (Şekil 5C) 0.2 um 2'ye kadar belirgin yayılma gücünde hızlı bir azalma sonucu ile, ardından yaklaşık 0.7 um -1 2 sn arasında bir ortalama D uygulaması ile, 100 nm altında bir birinci düz davranışı gösterir sn -1 (değer genellikle ölçülen kırınım sınırlı FCS 12). Bu sonuç yaklaşım kolayca nanometre birkaç on çözünürlüğe GFP etiketli proteinlerin ortalama yer değiştirmeyi ölçmekte göstermektedir. Ayrıca D uygulaması başlayacaktır hangi mekansal ölçek belirledi¤inin karakteristik azaltmak içinönceki tahminlerden 6 ile tutarak yaklaşık 120 nm membran iskelet protein kısmi hapsi mekansal ölçek,.

Nokta Fark Fonksiyonu Şekil 1. Kalibrasyon. (A), bir izole edilmiş ve boncuk boncuk agrega Pseudocolor görüntüsü. Izole edilmiş bir tanenin, yoğunluk profilinin (B) 3D arsa iyi tanımlanmış bir Gauss profilini gösterir. (C) bir Gauss fonksiyonuyla yoğunluğu dağılımının Fit (üst panel) karşılık gelen artıklar (alt panel). Monte dağıtım ve ölçülen yoğunluk profilinin arasındaki iyi Anlaşma aynı zamanda enstrümantal PSF bir Gauss fonksiyonu ile tahmin edilebilir bir kanıtıdır. vi için tıklayınız Bu rakamın büyük bir versiyonunu ew.

Tek fotonlardan Kamera tepki Şekil 2. Kalibrasyon. Rakam kırpılmış Sensör Modu 32 x 128 ROI kamera arka, pozlama 0.5 milisaniye için Dijital Seviye (DL) dağılımını göstermektedir. Yaklaşık 180 dl tepe hiçbir fotonlardan kamera yanıttır. Özellikle, Analog Dijital (AD) çevirici katkısını temsil eder ve ofset ve sinyal kaydı ile tanıtıldı varyansını tahmin etmek bir Gauss fonksiyonu ile tahmin edilebilir. 200 DL Yukarıdaki dijital düzeylerinin dağılımı üstel olur ve tek bir foton ortalama kamera yanıttır. Bu parametrelerin ölçümü edinimi sırasında kaydedilir foton yoğunluğunu tahmin sağlar.s / ftp_upload / 51994 / 51994fig2highres.jpg "target =" _blank "> Bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.

Yöntemin Şekil 3. Şemalaştınldığmdan. (A) 'EMCCD kamera tarafından geniş alan görüntüleme TIRF küçük kopya plazma zarının doğru optik kesitlerinin sağlamak için kullanılabilir durumdaysa, milisaniyenin çözüme ulaşmak için uygulanır. (B)' Elde edilen görüntü yığını ortalama uzaysal hesaplamak için otomatik korelasyonu yapılmış olduğu -temporal korelasyon fonksiyonu. Bu ilişki fonksiyonu iyi bir Gauss fonksiyonu ile yaklaşılır (bkz) ve partikül değiştirmelere göre zaman içinde yayılır. (C) Böylece nedeniyle moleküler değiştirmeye korelasyon fonksiyonunun yayılmasını ölçmek amacıyla, bir Gauss uydurma ian işlev gerçekleştirilir. Bu ortalama deplasman arsa vs belirgin yayılma şeklinde, doğrudan görüntüleme gelen moleküler 'difüzyon yasası' ölçme verir. (D) sayesinde, bu arsa için, moleküler difüzyon modları doğrudan ilgili bir yorumlayıcı modeli veya varsayımları gerek ile tespit edilebilir zarın mekansal organizasyonu. Hareketlilik ölçme alansal ölçekte bağlı değildir Aslında, serbest difüzyon molekülleri, sabitin yayılma gösterir. Buna göre, kısmi sınırlı moleküller hapsi boyutundan daha küçük yer değiştirmeler için oldukça sabit belirgin yayılma gösterecektir, sonra lohusalık boyutundan daha büyük mekansal ölçekler için azalan bir yayınım. Ilgili uzamsal ölçü lohusalık Mekansal tahmin etmek için kullanılabilir Böylece, yayılma belirgin bir azalma görünümü, geçici kısıtlanmasının bir parmak izi olarak yorumlanabilir. .jove.com / files / ftp_upload / 51994 / 51994fig3highres.jpg "target =" _blank "> Bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.

Canlı hücre zarlarında Şekil 4. ATTO488-PPE difüzyon yasası (A) hücre zarındaki ATTO488-KKE yerleştirilmesi şematik gösterimi (B) ATTO488-KKD ile etiketlenmiş CHO bazal membran TIRF görüntü.:. ROI (kırmızı kutu) seçili Seçilen ROI ölçülen difüzyon yasası, bu bileşen için ücretsiz bir difüzyon modeli doğrulayan düz bir davranışı gösterir hücre sınırı ve yüksek floresan noktalar kaçınarak hücrenin bir çoğunlukla üniforma parçası olarak. (C). daha büyük bir versiyonunu görmek için buraya tıklayınız Bu rakam.

Canlı hücre zarlarında Şekil 5. TfR GFP difüzyon kanunu (A), hücre zarı içinde TfR GFP ekleme şematik gösterimi:. Alıcısının sitoplazmik kuyruk reseptör hareketlilik için bir çit olarak hareket eden membran iskelet ile etkileşime girer (B. ) CHO ifade TfR-GFP TIRF görüntü: Bir ROI nedeniyle aşın eserler önlemek için, düşük sentezleyen hücrelerin tercih seçilir (C) TfR (siyah noktaların difüzyon yasası), PPE aksine (gri çizgi) Şekil 4 den, alınan. İlk düz kısmı D app bir düşüş ardından, kısmen sınırlı difüzyon tipik bir davranıştır. gösterir bu fi büyük halini görmek için buraya tıklayınızGüre.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Tek parçacık izleme (SPT) moleküler dinamiklerini incelemek için en yaygın stratejilerden birini temsil ve parçacık yörüngeleri ölçüm büyük bir avantaja sahiptir. Bu da karmaşık bir sistemde bile birkaç etiketli parçacıkların davranışlarını sondalama sağlar. Bununla birlikte, tipik haliyle SPT sistemi ve çok parlak bir etiket düşük yoğunluklu ihtiyacı bu avantajı ulaşmak için. Sisteme, üretim, etiketleme ve yerleştirme karmaşık bir prosedür gerekli bu durumda: Özellikle, yüksek temporal çözünürlük (mikro saniye olduğu aralık) inorganik sonda genellikle gereklidir (örneğin, kuantum noktalar veya metal nanopartiküller) kazanmak. SPT karşılaştırıldığında, mevcut yöntem, bazı önemli avantajları gösterir. Her şeyden önce, bu yaklaşım, floresan proteinler ile birleşme kullanılabilir. Böylece, SPT kıyasla daha yüksek bir zamansal çözünürlüğü fotonların düşük miktarda sayesinde (aynı etiketinde) 34 elde edilir gerekmektedir. Daha ayrıntılı olarak, bu özellik zamansal resol iterek sağlar10 Aşağıda Katkı - 3 sn de encodable floresan proteinleri kullanırken, ve bu zaman ölçeğini membran bileşenlerinin nano dinamiklerine özel erişim sağlar. Son olarak, moleküler difüzyon yasaları, her molekülü izlemek gerek kalmadan, tam uzay-zaman korelasyon fonksiyonu analiz edilerek açıklanmıştır dikkati çekiyor.
STED'in tabanlı FCS ile karşılaştırılması da ilginç. Bir STED'in-FCS ölçüm gözlem hacim azalması moleküllerin ortalama geçiş süresi, floresan sinyalinin zamansal korelasyonu ile ölçülür. Bu aynı zamanda kırılma sınırının altında moleküler dinamiklerin yerel ölçümünü elde edilmesine izin verir. Sunulan yaklaşımda difüzyon kanunu standart sapması sınırlı, gözlem hacim vasıtasıyla gözlenen seçilmiş ROI hareket bütün parçacıkların ortalama olarak ölçülür. Bununla birlikte, sonuçlar, bu yöntem kırınımı ile sınırlı olmadığını göstermektedir, ancak th tarafından raporE zamansal çözünürlük yok. Bir kırınım-sınırlı kazanım iyi kırınım sınırının altında (örneğin benzer PALM ve FIRTINA gibi diğer süper çözünürlük teknikleri, yapılır ne) dalgalanmaları, moleküler deplasmanlar algılamak için kullanılır rağmen Aslında, zaten, (doğrudan) olabilir hesaplanan olabilir Moleküler ölçmek için ampüller kullanılarak gösterilebilmektedir ve 32 akar. Ayrıca, STED'in-FCS aksine, bu yaklaşım kolay gibi raster tarama mikroskopları veya geniş alan kamera tabanlı mikroskoplar gibi ticari ve mevcut mikroskopi kurulumları, geniş bir yelpazede uygulanabilir. Bu moleküler difüzyon yasaları STED'in-FCS ölçümleri kesinlikle enstrümantal bel boyutu flüorofor bağımlı kalibrasyon gerektiren söz layık olduğunu. Buna karşın, ölçüm (parçacık boyutunun hesaplanması için sadece gerekli) bir sistem kalibrasyon gerektirmez burada açıklanmıştır.
Sunulan bir yöntem de parçacık değiştirmeler ölçümünde gerçek çözünürlükBiz korelasyon fonksiyonu ölçmek için ne kadar doğru basımınızın. Sonuç olarak, doğal olarak benzer çözünürlük parçacık "görüntü" nasıl ölçüldüğü doğru bağlıdır SPT duruma, kırınımı ile sınırlı değildir. Önerilen deneyler için 1 dakikadan daha az önemli bir korelasyon ölçmek için, her bir çerçeve içinde, her bir parçacık için birkaç fotonlar (genellikle 10 altında bir foton) yeterlidir. Bağıntı fonksiyonu hesaplanır Aslında, gözlenen bütün parçacıkların katkı partikülleri izole olsa bile, birlikte ortalaması alınır. Bu özellik dalgalanma korelasyon yöntemleri içsel ve böyle canlı hücrelerde transfekte floresan proteinleri gibi loş ve yoğun etiketleri kullanarak verir.
Bu göz önüne alınarak, asgari ölçülebilir değiştirme parçacığın yayılma ve görüntüleme kurulum zaman çözünürlüğüne bağlı olduğu açıkça görülmektedir. Bir örnek olarak, hücre zarı üzerinde moleküllerin difüzyon dikkate alınız,proteinler ya da lipidler için maksimum ölçülen yayınım yaklaşık 5 mikron 2 sn -1 nerede. Bu koşullar altında, 50 nm'lik bir ortalama yer değiştirme yakalamak için yaklaşık 10 -4 sn'lik bir zaman çözünürlüğü gerekir. Burada gösterdiği gibi bu çözünürlüğü, tek hatları boyunca veya zaman çözünürlüğü pozlama süresi denk hızlı EMCCD kamera tarafından hızlı tarama mikroskopları ile elde edilebilir.
Doğru moleküler dinamik tanımlamak için bu yöntem için ek bir temel gereksinimi doğru mekansal örnekleme. Aslında, bağıntı fonksiyonunu uyması için biz aracı PSF bel daha düşük uzaysal bir örnekleme (piksel boyutu) gerekir. En ticari mikroskopları (konfokal veya geniş alanı) olarak, PSF bel 200 nm ila 500 nm arasında olur (ikinci seçilen amacı, sayısal açıklığı üzerine ve dalgaboyu ağırlıklı olarak) ve kolay bir şekilde kalibrasyon deney kullanılarak nano ölçülebilir boy floresan boncuk. THus, (enstrümantal bele oranla 3 kat daha düşük) 70-150 nm bir piksel boyutu yeterli olabilir. Bununla birlikte, piksel boyutu dikkate alan basit bir kural çalışılan sisteme adapte edilebilir: bağıntı fonksiyonunun tarifinde piksel boyutunu daha düşük doğruluk. Bundan başka, elde edilecek görüntünün en küçük boyutu, ilgi konusu bir maksimum yer değiştirme (artı aletsel bel) en az 3-kat daha büyük olması gerekir. Bu uydurma algoritması iyi bir yakınsama ve moleküler yer değiştirme istatistiksel olarak anlamlı bir örnekleme ulaşmak için gereklidir. Bir örnek olarak, nanometre ile birkaç yüz (örneğin, 200 nm): birkaç mikron bir görüntü büyüklüğü kadar olduğundan daha küçük ortalama bir moleküler yer değiştirmeleri incelemek. Ayrıca, bağıntı fonksiyonunun kalitesi üzerindeki etkileri (sabit piksel boyutu alınarak) toplam piksel sayısı. Aslında, daha geniş bir resim dahi zaman rezolüsyonlu pahasına, bağıntı fonksiyonunun daha fazla bilgi sağlar ortalamaiyonu temsil etmektedir. Burada kullanılan kamera tabanlı sistemi ile ilgili çip üzerindeki pikselin fiziksel boyutunun sabit olduğunu lütfen unutmayın. Sonuç olarak, piksel boyutunu azaltarak, (piksel boyutunda kare bağlıdır) her piksel sinyali düşürür görüş alanını azaltır ve daha yüksek büyütme gücü gerektirir. Öte yandan, gözlem alanı piksel boyutu genellikle zaman çözünürlüğü pahasına artan piksel sayısının azalması ile sonuçlanan sabit bir tarama sistemi içinde.
Kullanılan dedektör hakkında birkaç bilgi tartışılması gerekir. Tek foton detektörlerin aksine, EMCCD sistemler, bir ofset varlığı toplanan ışık ile direkt olarak orantılı değildir ortalama yoğunluğu (dijital seviyesi, DL) ölçün. Offset bu kameranın dinamik aralık ile karşılaştırıldığında düşük olsa bile ve önemsiz birçok fotonlar toplanır deneylerinde (birkaç yüz 2 16 bit okuma 16 karşılaştırıldığında), bu dikkate alınmalıdırbağıntı fonksiyonunun doğru bir normalleşme elde edilir. Ayrıca, toplanan sinyalin miktarını belirlemek için düşük ışık koşullarında bir referans olarak kullanılabilir ofset. Ayrıca, satın alma sırasında toplanan fotonların ortalama miktarını tahmin etmek için, ortalama dijital seviye toplanan her foton ilişkili ölçülmelidir. Bu miktar çok düşük ışık yoğunluğu kamerayı açığa alınabilir (örneğin, oda arka ışık); Aslında, bu durumda, makul surette ölçülen yoğunluğu sıfır ya da bir foton sadece ilişkili olabilir, yani sadece tek bir fotonlar, kameraya ulaşıncaya varsayabiliriz.
Son olarak, bize bazı alternatif toplama sistemleri (örneğin, farklı mikroskopi kurulumları) sunulan ölçümleri gerçekleştirmek için kullanılabilir nasıl belirtmeme izin verin. Her şeyden önce, Denklem 2'de "W'factor (yani aletsel PSF otokorelasyon ifade etmektedir) t uyarlanabilirdeneysel korelasyon fonksiyonu sığdırmak için kullanılan o özellikle kazanım sistemi. Daha önce 34 gösterildiği gibi, kolay bir durum tarama hızı parçacık dinamiği anlamlı derecede yüksek olduğu zaman satın alma bir lazer tarama mikroskobu katiyen olduğunu. Böyle bir durumda, aslında, elde etme süresi boyunca parçacıkların hareketi (örneğin, hat zaman) ihmal edilebilir olarak kabul edilebilir ve korelasyon fonksiyonu iyi bir Gauss fonksiyonu tarafından yaklaşık olarak ifadelendirilmektedir. Ortaya çıkan görüntüleme teknolojileri bağlamında, ilginç bir yaklaşım, numunenin 41 üzerinden çok ince ışık tabakaları (1-2 um) üretmek için ihtimali dayanmaktadır. Işık tabaka 3D 42 kamera esaslı alıcı sistemi, hızlı optik olarak kısımlara ayırma ile birlikte, tek bir numune içinde düzlemi (düzlem tek aydınlatma mikroskopisi SPIM) ve selektif aydınlatılmasını sağlar. Bu özellikleri nedeniyle, SPIM başarılı FCS 43 ile konjuge edilmiş bir vali temsil edebilecekd aracı gibi sitoplazma ya da canlı hücre çekirdeği olarak 3D ortamlarda, için sunulan analiz uzatmak için.
Bakış deneysel açıdan Özetle, bu yaklaşım hızlı bir alıcı modülü ile donatılmış bir mikroskop yalnızca erişim gerektirir. İlgilenilen protein böylece, çok renkli görüntüleme imkan veren herhangi bir floresan proteini veya organik florofor ile etiketlenmiş olabilir. Bu bağlamda, moleküllerin alt popülasyonları seçip canlı hücre membranlarında etkileşimleri ve ko-difüzyon ortaya i MSD analiz çapraz kullanma imkanı öngörülüyor. Son olarak, bu yaklaşım, plazma zarı üzerinde nanodomains içinde dinamik bölümleme geçiren proteinler ve / veya lipidler incelemek için güçlü bir araç olabilir inanıyoruz. Bu durumda, nanodomains son derece değişken boyutu ve ömrü 2-renk görüntüleme gibi ileri metodolojik uygulamaları gerektirecek gerçek verileri karmaşıklık ek bir düzeyde tanıtmak, yerelanalizi (örn, 2 çifti korelasyon) ve / veya fosforlu anisotropi.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}