

Abstract
De neuromusculaire junctie (NMJ) is de grote, cholinerge relais synaps waardoor zoogdieren motorische neuronen controle vrijwillige spiersamentrekking. Structurele veranderingen op de NMJ kan resulteren in neurotransmissie falen, wat resulteert in zwakheid, atrofie en zelfs de dood van de spiervezels. Vele studies hebben onderzocht hoe genetische modificaties of aandoening de structuur van de muis NMJ kan veranderen. Helaas kan het moeilijk zijn direct vergelijken resultaten van deze studies, omdat ze vaak gebruikt verschillende parameters en analysemethoden. Drie protocollen worden hier beschreven. De eerste maakt gebruik van maximale intensiteit projectie confocale beelden op het gebied van acetylcholine receptor (AChR) te meten rijke postsynaptische membraan domeinen bij de eindplaat en het gebied van de synaptische blaasjes kleuring in de bovenliggende presynaptische zenuwuiteinde. Het tweede protocol vergelijkt de relatieve intensiteiten van immuunkleuring voor synaptische eiwitten in het postsynaptische membraan. De derde protocol gebruikt Fluorescence Resonance Energy Transfer (FRET) veranderingen in de verpakking van postsynaptische AChRs aan de eindplaat detecteren. De protocollen zijn ontwikkeld en verfijnd over een reeks van studies. Factoren die de kwaliteit en de consistentie van de resultaten beïnvloeden worden besproken en normatieve gegevens zijn bedoeld voor NMJs bij gezonde jonge volwassen muizen.
Introduction
De neuromusculaire junctie (NMJ) is de kritische relais synaps dat de communicatie tussen het zenuwstelsel en de skeletspieren bemiddelt. Het is vereist voor alle vrijwillige beweging. Fluorescentie microscopie al lang gebruikt om de effecten van transgenen bestuderen op muis NMJ 1-3 of de effecten van leeftijd, dieet, lichaamsbeweging en ziekte vergelijk bij knaagdieren NMJs 4-11. Dergelijke studies hebben ons veel geleerd over de fysiologie en pathofysiologie van de NMJ, maar de diverse parameters gerapporteerd (bv AChR gebied, eindplaat oppervlakte, omtrek lengte, fragmentatie indices) maken het vaak moeilijk om de resultaten van deze studies te vergelijken. Er is een toenemende verwachting voor preklinische onderzoekers kunnen reproduceerbaar zijn, vooral in studies met diermodellen voor ziekten 12. De hier beschreven protocollen werden verfijnd door een reeks studies die ontwikkeling, fysiologie en pathofysiologische ch onderzochtanges naar de NMJ. Dergelijke studies vereisen meting van het gebied van de synaptische specialisaties op de muis motorische eindplaat en de relatieve dichtheid van de verpakking van synaptische eiwitten binnen postsynaptische specialisaties 13-15.
De bruikbaarheid van deze methode wordt geïllustreerd door recente studies in een muismodel van anti-MuSK myasthenia gravis. Dagelijkse injecties van IgG uit anti-MuSK-positieve myasthenia gravis patiënten in volwassen muizen zorgde ervoor dat ze zwak worden binnen 2 weken 16. Confocale maximale projectie beelden spier secties werden dubbel gelabeld voor synaptofysine (in zenuw-terminals) en postsynaptische AChRs toonde een geleidelijke afname van het gebied van AChR kleuring als primaire verandering. Belangrijk is het tempo van de daling was voldoende om vergelijkbare dalingen in de amplitude van synaptische potentialen, falen van synaptische transmissie en spierzwakte 17,18 verklaren. Kwalitatief vergelijkbaar bevindingen werden gerapporteerd door andere onderzoeksgroepen10,19. Dezelfde NMJ meetmethoden zijn sindsdien gebruikt om het effect van drie geneesmiddelen te beoordelen voor het behandelen van anti-MuSK myasthenia gravis in dit muismodel 20,21.
Sedentaire veroudering kan leiden tot verlies van de neuromusculaire verbindingen. De hier beschreven protocollen hebben een leeftijdsgebonden achteruitgang op het gebied van zenuwuiteinde synaptofysine bij motorische eindplaten geopenbaard als muizen vooruitgang tot op hoge leeftijd. Dezelfde methoden bleek dat vrijwillige uitoefening grotendeels kon voorkomen dat de vermindering van de presynaptische zenuwuiteinde gebied 22, in overeenstemming met eerder werk van andere groepen 4. Verlies van neuromusculaire verbindingen komt ook voor in de SOD1G93A muismodel van amyotrofische lateraal sclerose 9,23.
De bovengenoemde studies tonen aan dat een verscheidenheid van gezondheidsproblemen kan leiden tot vermindering van het gebied van een van beide pre- of post-synaptische specialisaties op het NMJ. Dit kan resulteren in een verminderde synaptische functie of kunnen inluiden volledig verlies van de neuromusculaire verbinding. Drie protocollen worden beschreven dat kwantificering van het gebied en de dichtheid van synaptische specialisaties mogelijk te maken. Het doel van het eerste protocol is een praktische en reproduceerbare meting van de oppervlakken van pre- en postsynaptische specialisaties en hun aanpassing aan zoogdieren NMJs indienen waarbij fluorescentiemicroscopie. Tweedimensionale maximum projectie confocale beelden en beeldanalyse met NIH ImageJ om wijzigingen in het gebied van synaptophysine kleuring (synaptische blaasjes), postsynaptische AChRs en synaptische overlappingsgebied detecteren. Confocale beeldvorming parameters (gain en offset niveau) worden geoptimaliseerd voor elke NMJ om de visuele informatie gebruikt om het gebied van synaptische specialisatie onderscheiden maximaliseren. Neuromusculaire mislukking kan ook resulteren uit veranderingen in de dichtheid van postsynaptische AChR en / of andere synaptische eiwitten. Het tweede protocol kan worden toegepast om de veranderingen in de relatieve dichtheid van postsynaptische eiwitten detecterenals muskus, rapsyne, dystroglycan, gefosforyleerd Src kinase en gefosforyleerd AChR 18,21.
In gravis myasthenie, een verminderde dichtheid van AChR binnen de postsynaptische membraan is de directe oorzaak van synaptische mislukking en spierzwakte. Het derde protocol beschrijft een Fluorescence Resonance Energy Transfer (FRET) gebruikt om veranderingen in de nabijheid van aangrenzende AChRs op postsynaptische membranen 14,15 beoordelen. Deze methode detecteert energieoverdracht tussen naburige AChRs gelabeld met fluorescerende-α-bungarotoxin (BGT). FRET alleen gebeurt wanneer de fluorescerende donor en acceptor probes minder dan 10 nm uit elkaar. Dit kan (submicroscopic) veranderingen in de dichtheid van AChR verpakking die rechtstreeks betrekking kunnen hebben op de amplitude van synaptische potentialen onthullen.
Deze drie protocollen, verfijnd in het afgelopen decennium, bieden aanvullende maatregelen van NMJ integriteit op een consistente en reproduceerbare wijze. Het gebruik van gestandaardiseerde protocollen eennd parameters dienen vergelijking van de effecten van genen en milieu-ingrepen op het zoogdier NMJ vergemakkelijken.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
OPMERKING: Ontwerp, uitvoering en rapportage van dierproeven moeten rekening houden met de huidige richtlijnen 24. Dergelijke werkzaamheden moeten vooraf worden goedgekeurd door de lokale dierenwelzijn gezag (in ons geval de Animal Ethics Committee van de Universiteit van Sydney).
1. Euthanasie van de dieren en de Muscle Dissection
- Breng de muis van het dierenverblijf naar een aparte kamer waar het geëuthanaseerd met een intraperitoneale injectie van pentobarbiton oplossing (30 mg / kg) met de muiswerkzaamheden methode beschreven door Shimizu 25. Plaats de muis weer in de kooi.
- Zodra de ademhaling van de muis is gestopt voor meer dan 1 min, test de voet-terugtrekking reflex door zachtjes te knijpen de voet, en de cornea reflex door licht borstelen het hoornvlies. Alleen als reflex reacties afwezig zijn kan de muis worden voorbereid voor dissectie.
- Raadpleeg een atlas van de anatomie van knaagdieren zoals Chiasson 26 en / of zoeken naar de hulp van een experienced anatoom voordat dissectie van de spier van belang. In elk geval te verwijderen haren uit de bovenliggende huid met behulp van een kleine elektrische scheerapparaat voor het openen van de huid om de spieren bloot.
OPMERKING: De dissectie zal verschillen per anatomisch afzonderlijke spier. - Met behulp van botte tang gratis de spier van bovenliggende membranen en omliggende weefsels. Pak en snijd de distale pees om de spier te scheiden van haar inbrengen.
- Zachtjes plagen en knip de spier vrij van omliggend weefsel direct terug naar zijn oorsprong. Kort plaats de nieuwe spier ontleed in 0,1 M fosfaat gebufferde zoutoplossing (PBS) oplossing of Ringer's oplossing voorafgaand aan verdere verwerking.
2. Voorbereiden van de Spier voor cryosectioning
OPMERKING: Optimale structurele conservering kan worden bereikt door volledige dier perfusie zoals eerder beschreven 27 of onderdompeling fixatie (kleine spieren) zoals beschreven in optionele stap 2.1. Echter,4% paraformaldehyde fixatie daaropvolgende kleuring beïnvloeden vele antilichaam probes en met fluorescerende-BGT. Glutaraldehyde bijzonder moet worden vermeden. Als spieren niet vast te stellen moeten ze onmiddellijk worden snel bevroren (ga naar 2.3).
- Optioneel onderdompeling fixatie: Speld de spier om de was in een petrischaal bij rust lengte. Bedek de spier met 2% w / v paraformaldehyde (vers opgelost in PBS) gedurende 2 uur bij KT. Wassen met 3 veranderingen van PBS gedurende 30 min (3 x 10 min) plaats de PBS met 30% w / v sucrose in PBS en geïncubeerd O / N bij 4 ° C.
- Voeg mallen ("boten") vooraf door vouwen 2 cm x 1.5 cm stukjes aluminiumfolie zoals weergegeven in figuur 1. Leg een vel nitrocellulosemembraan in de bodem van de boot. Giet langzaam een cryostaat inbedden matrix (Materials tabel) in de boot tot een diepte van 2 mm, de zorg om luchtbellen te voorkomen. Plaats de spier in de boot, gelijk met de balpen lijnen opde nitrocellulose. Voeg meer inbedden matrix teneinde volledig te bedekken de spier (figuur 1).
- Pre-label polypropyleen buizen met een onuitwisbare stift. Plaats een druppel water in elke buis en relaxen de buis in vloeibare stikstof.
OPMERKING: De bevroren waterdruppel handhaaft de dampdruk en voorkomt uitdroging tijdens langdurige -80 ° C bewaren - Met behulp van een vizier, dikke beschermende handschoenen en een grote paar stompe tang, gedeeltelijk verlagen van een kleine metalen beker (3 cm doorsnede, 8 cm diep) met 2 cm diepte van isopentaan in een container van vloeibare stikstof voor 30 sec. Verwijder het bekerglas en zet hem op de bank top. Met behulp van een kleinere paar stompe pincet plaats de mal met de spieren en het inbedden van matrix in de gekoelde isopentaan. Zorg ervoor dat vloeibare stikstof mengen met de isopentaan te voorkomen.
- Laat 2 min voor het blok volledig bevriezen voordat u botte tang om de bevroren blok uit te tillen en sluit deze in de juiste pre-gelabelde en voorgekoelde buis (stap 2.3).
- Bewaar de buizen tijdelijk in de vloeibare stikstof alvorens naar -80 ° C. Log alle monsters in een spreadsheet van de inhoud vriezer.
3. cryosectioning en Fluorescentie kleuring voor En Face afbeeldingen van NMJs
- Schil de aluminium mal weg. Binnen de -20 ° C cryostaatkamer bevestigen het bevroren blok aan de cryostaat klem om snijden 20 urn vriescoupes evenwijdig aan de lange as van de spiervezels (figuur 1). Pak de secties op poly-L-lysine of gelatine gecoate microscoop dia's.
- OPMERKING: Sla deze stap over als het weefsel voorafgaand aan het bevriezen is bevestigd. Nadat men 30 min bij gedeelten drogen op de glijbanen, bevestig ze door een druppel van 2% paraformaldehyde in PBS gedurende elke sectie gedurende 15 minuten bij kamertemperatuur.
- Was slides 3 x 10 min in PBS in een Coplin jar, en dompel de objectglaasjes in PBS met 0,1 M glycine gedurende 30 min om resterende aldehydegroepen blokkeren.
- Was objectglaasjes gedurende 10 minuten in PBS, vervolgens ondergedompeld in methanol (gekoeld tot -20 ° C) gedurende 7 min. Dit permeabilisatie stap is een routine-onderdeel van dubbele labeling met tl-BGT en anti-synaptofysine maar het kan een negatieve invloed hebben immuunkleuring voor een aantal andere eiwitten.
- Was glijdt 2 x 10 minuten in PBS plaats dan elke dia in een stabiele en geëgaliseerd bevochtigde kamer. Die onmiddellijk elke sectie met 20 ui blokkerende oplossing (0,2% Triton X-100, 2% runderserumalbumine (BSA) in PBS) gedurende 1 uur bij KT. Secties mag niet uitdrogen in elk stadium van de immunokleuring proces.
- Het uitvoeren van de primaire incubatie: Het nemen van een dia in een tijd de overtollige blokkerende oplossing te verwijderen voorzichtig uit meer dan elke sectie en te vervangen door 20 ul van konijnen anti-synaptofysine (verdund 1: 200 in de blokkerende oplossing).
- Neem een negatieve controle dia die worden geïncubeerd met alleen blokkeringsoplossing. Deze 'no-primaire antilichaam controle'Is essentieel in elk immunokleuren run.
- Zorg ervoor dat het primaire antilichaam blijft op zijn plaats dan elke sectie, sluit de vochtige kamer en incubeer gedurende 1-2 dagen bij 4 ºC.
- Inspecteer elke sectie om te bevestigen dat het primaire antilichaam op zijn plaats blijft. Gebruik een Pasteur pipet om voorzichtig te spoelen elke dia met PBS en plaats deze in een Coplin pot. Was de schuiven 3 x 10 min in PBS.
- Het uitvoeren van secundaire incubatie. Het nemen van een dia op een moment, verwijder voorzichtig overtollige PBS, leg het in de vochtige kamer en bedek elke sectie met 20 pi van een mengsel dat FITC-geconjugeerd ezel anti-konijn IgG en BGT geconjugeerd aan rhodamine Tetramethyl of een andere rode fluorofoor (TRITC- / redBGT, 5 g / ml) verdund in blokkerende oplossing. Incubeer bij kamertemperatuur gedurende 2 uur.
- Was glijdt 3 x 10 min in PBS in Coplin potten.
- Het nemen van een dia op een moment, verwijder voorzichtig overtollige PBS en monteer met een dekglaasje met behulp van een minimaal volume, glycero-l gebaseerde, fade-weerstand montage medium. Dicht de randen van de dekglaasjes met heldere nagellak. Laat het moeilijk drogen.
- Bewaar de objectglaasjes in het donker bij 4 ° C gedurende maximaal een week, of bij -20 ° C voor langere opslagtijden (tot enkele maanden).
4. Aanbevelingen bemonstering en En Face Imaging van Motor Eindplaten
- Verblindt de dia's door het labelen van elk glaasje met een willekeurige code die alleen bekend is blijft een tweede onderzoeker (niet betrokken in de analyse). Hierdoor blijft de exploitant blind behandelingsgroepen tot kwantificering van NMJ parameters is voltooid voor alle monsters.
- Plaats de dia op de microscoop podium en bekijk het onder brede veld belichting met de TRITC filter set (63x olie 1.3 NA objectief). Verplaats progressief (veld voor veld) van links naar rechts en weer tot een eindplaat verschijnt in het veld (Figuur 2A).
OPMERKING: Sampling criterium: Elke AChR-gekleurd structuur die relatiefvlak en tegenover het objectief (dwz uitstrekt <15 m in de z-dimensie) wordt beschouwd als een eindplaat en wordt afgebeeld voor analyse (halvemaantjes van AChR kleuring doorsneden door eindplaten en worden daarom uitgesloten). - Met de confocale pinhole op 1,0 Airy eenheid en lage laservermogen optimaliseren versterking en offset niveaus TRITC / rood-BGT (532 nm laser) en de eindplaat dat moet worden afgebeeld. Volgende optimaliseren FITC / synaptofysine fluorescentie met behulp van de 488 nm laser. Verzamel een z-stack van de eindplaat met een 0,7 urn interval tussen elke optische segment. Sla de beelden met een bestandsnaam die de datum van de opnamesessie, de codenaam van de glijbaan en het nummer van de eindplaat omvat.
OPMERKING: De scans met de 488 nm en 532 nm lasers (FITC en TRITC) worden verzameld opeenvolgend (niet tegelijkertijd) verontreiniging van het FITC kanaal fluorescentie van het rode fluorofoor en vice versa (doorbloeding) voorkomen. - Herhaal de bemonstering eennd beeldvorming van stappen 4,2-4,3 tot 20 eindplaten worden verzameld uit de dia / monster.
- Ga naar de volgende gecodeerde glijbaan en herhaal 4,2-4,4. Herhaal deze procedure voor elk van de gecodeerde objectglaasjes.
- Verzamel een paar beelden van de eindplaten van de regelschuif (no-primair antilichaam controle) met behulp van confocale instellingen die optimaal zijn voor de experimentele dia's (de FITC fluorescentie kanaal moet donker zijn) werden gevonden.
- Aan het einde van de confocale sessie overdracht van de beeldbestanden naar een andere computer en een back-up van de originele bestanden op een externe schijf of server.
5. Het meten van de ruimte van Synaptic Specialisaties in En Face Afbeeldingen
- Gebruik NIH ImageJ freeware (http://imagej.nih.gov/ij/) voor maximale projectie (MIP) beelden te bereiden van elkaar z-stack. Opslaan als tiff-bestanden (Figuur 2A & B). Bestandsnamen moeten de datum afbeelding sessie, voorbeeldcode, eindplaat aantal en tl-kanaal (bijv 060414_57 omvatten23_7_FITC.tiff).
- Open de z-projectie afbeelding in ImageJ. Selecteer de acetylcholine receptor afbeelding kanaal (figuur 3A) en selecteer: Image> Type> 8-bits naar 24-bits RGB-afbeelding gekleurd om te zetten in drie 8-bits grijswaarden beelden op het scherm.
- Met behulp van de ImageJ polygoon gereedschap trek een ruwe schets rond de eindplaat van belang in de redBGT gekleurd (AVRM) kanaal, zodat alle schijnbare gebrandschilderde gebieden van de specifieke individuele eindplaat, met uitsluiting van alle vlekken die niet afkomstig zijn van de eindplaat van belang zijn ( Figuur 3C).
- Breng een minimale intensiteit drempel om het beeld door het selecteren van: Image> Adjust> Threshold (Figuur 3E en bijbehorende ImageJ screenshots).
- Pas de drempel om zo de AChR-gekleurd gedeelten te isoleren terwijl exclusief omliggende achtergrond signaal als sub-drempel (Figuur 3E). Open een tweede window met het originele beeld (continue toon) direct naast het raam ter vergelijking, om de beslissing over de drempelwaarde te vergemakkelijken. Noteer de drempelwaarde voor later gebruik in colokalisatie analyses.
- Behoud van de veelhoek omtrek rond de eindplaat selecteren: Analyseren> Analyseren Deeltjes. In het pop-up menu geeft u het bereik van maten: 50 tot oneindig pixels (dit elimineert kleine artefacten als gevolg van elektrische ruis in de photomultiplier).
- Analyseer Deeltjes commando genereert een venster met een lijst van discrete supraliminaire gebieden en hun fluorescentie-intensiteit waarden genummerd zoals ze in het binaire beeld (Figuur 3G en bijbehorende ImageJ screenshot). Kopieer deze gegevens in een gelabelde spreadsheet.
- Meet de totale eindplaat (gebied binnen de polygoon) door het selecteren van: Analyze> Meet. Dit levert de totale eindplaat gebied. Kopieer en plak de gegevens voor AChR gebieden en intensiteiten ineen spreadsheet en zorg ervoor dat de kolommen op de juiste wijze etiketteren, zal rijen worden gebruikt voor individuele eindplaten voor specifieke dia's.
- Schakelen naar de anti-synaptofysine fluorescentie kanaal en herhaal de stappen 5,1-5,5, maar voor de FITC kanaal (figuur 3B, D en F). Het doel is de drempel aan te passen zodat een binair beeld dat zo dicht mogelijk overeenkomt met de grenzen van kleuring zoals waargenomen door het oog ontstaat. Noteer de drempelwaarde.
- Meet het gebied van overlap door toepassing van de volgende stappen: Open het oorspronkelijke bestand met de tweekanaals afbeeldingen en opgesplitst in twee afzonderlijke afbeeldingen door het selecteren van: Image> Stapels> Stapel naar afbeeldingen.
- Met behulp van de colokalisatie plugin (gedownload en geïnstalleerd vanaf de ImageJ webpagina) Selecteer: pluggin> colocalization en input van de drempelwaarden eerder opgenomen voor de AChR en zenuwbanen in het desbetreffende kanaal query-bos. Dit zal een overlap afbeelding in witte pixels (Figuur 3H en bijbehorende ImageJ screenshots) opleveren.
- Deze afbeelding converteren nieuw gecreëerde overlap in een grijstinten formaat en een drempel van toepassing op de maximale waarde. De maximumgrens alleen selecteert de witte pixels, corresponderend met het overlappende gedeelte van de twee voorgaande kanalen. Record in de spreadsheet de resulterende gebied waarde van 'colokalisatie', die het gebied van overlap in beeldpunten.
- Bereid een spreadsheet van data steekproefgemiddelden, berekenen en plotten standaarddeviaties en standaard fouten histogrammen of scatterplots 20,22. Ook de waarde van n in het algemeen het aantal muizen per steekproef voor statistische doeleinden.
- Plot eindplaat AChR gebieden scatterplots of frequentiehistogrammen te bepalen of de gegevens normaal verdeeld voor statistische test (figuur 6).
6. Relatieve kleuringIntensiteiten vergeleken met behulp van Dwarse optische secties
LET OP: Voor dit protocol proces worden alle spieren monsters bij elkaar en het beeld in een enkele confocale sessie. Bij de planning van een experiment te laten tot 30 min beeldvorming keer per spier monster.
- Snij 15 urn vriescoupes dwars op de lange as van de spiervezels en het verzamelen op objectglaasjes zoals beschreven in stap 3.1.
- Het uitvoeren van fluorescentie kleuring, zoals beschreven in de stappen 3,2-3,13.
- Code de gekleurde coupes, zodat beeldvorming en analyse wordt uitgevoerd met de exploitant blind voor de behandeling groep, zoals beschreven in stap 4.1.
- Met behulp van een 40X fluorescentie doelstelling (NA 0,75) enquête kort een deel van elke dia om een enkele winst te bepalen en offset-niveau-instelling voor AChR die geschikt zijn voor alle eindplaten in alle sample dia's zullen zijn. De helderste eindplaat moet dan net onder 256 grijs op de schaal. Deze optimalisatie moet afzonderlijk worden gedaan voor de tweede fluorescence kanaal (achtereenvolgens verzameld). Noteer de vaste gain en offset instellingen van het niveau en ze niet veranderen gedurende de opnamesessie.
- Verzamel beelden van een fluorescentie standaardglaasje (bijvoorbeeld niet-blekende fluorescerende beads) met dezelfde parameters aan het begin en einde van de confocale sessie eventuele fluctuatie in laserintensiteit detecteren.
- Gebruik de AChR kanaal naar de dia progressief scannen naar eindplaten lokaliseren.
- Focus op de enkele optische sectie vliegtuig in elke microscoop gebied dat het meeste aantal AChR-gekleurd eindplaten bevat te vinden.
- Scan deze enkele optische gedeelte twee keer en bespaart de gemiddelde afbeelding (figuur 4G).
- Met behoud van hetzelfde focal plane schakelaar naar het tweede fluorescentie kanaal (eiwit van belang) en het verzamelen van het beeld als bij stap 6.8. Sla het image-bestand, ook in de bestandsnaam: datum van de opnamesessie, voorbeeldcode, afbeelding nummer en een symbool voor de tl-kanaal aan te geven.
- Verplaats de etappe naar het volgende veld dat een of meer eindplaten bevat en herhaal stap 6,8-6,9. Herhaal dit tot een totaal van 60 eindplaten worden afgebeeld.
- Aan het einde van de opnamesessie overdracht van alle bestanden naar een andere computer en een back-up.
- Open elke originele beeldbestand en tijdens het bekijken van de AChR kanaal, selecteert: Image> Stapels> Stapel naar afbeeldingen, om kanalen te splitsen.
- Selecteer: Afbeelding> type> 8bit te converteren naar 8-bits grijstinten formaat op het scherm. Doe dit voor beide fluorescentie kanalen.
- Selecteer: Beeld> Stapels> afbeeldingen te stapelen. Open een nieuwe stapel van twee eerder gescheiden 8-bits afbeeldingen. Men kan dan overschakelen gunstig tussen de twee fluorescentie kanalen binnen de enkel venster.
- Gebruik de polygoon gereedschap om een lin trekkene strak rond de grens van de AChR kleuring (figuur 4I).
- Selecteer: Analyze> Meet de gemiddelde pixelintensiteit voor AChR binnen het afgesloten gebied te meten (let op het belang van het tekenen van de lijn strak). Kopieer deze waarde in een gelabelde spreadsheet.
- Behoud van dezelfde veelhoek omtrek (om het gebied te definiëren te meten), over te schakelen naar de tweede fluorescerende kanaal (bijvoorbeeld figuur 4B, D, F) en selecteer: Analyze> maatregel. Dit zal de gemiddelde kleurintensiteit oplevert voor het eiwit van belang in de synaptische gedefinieerd door AChR kleuring.
- Kies een gebied uit de buurt van zichtbare eindplaat vlekken selecteer vervolgens: Analyze> Meet de gemiddelde achtergrond fluorescentie-intensiteit te meten. Herhaal dit voor de andere fluorescentiekanaal / s en kopieer de achtergrond waarden in de spreadsheet van fluorescentie waarden.
- Subdarmkanaal de gemiddelde achtergrondwaarden van eindplaat waarden om de gecorrigeerde intensiteiten voor AChR en het eiwit van belang op elke eindplaat verkrijgen.
- Deel de gecorrigeerde eindplaat intensiteitswaarden voor het eiwit van belang door de gecorrigeerde BGT fluorescentie intensiteit van de fluorescentie-intensiteit ratio 14,21 opleveren
7. Vergelijking van de Postsynaptische Membrane AChR Dichtheid Met behulp van FRET
Opmerking: Dit protocol beoordeelt de mate waarin AChRs nauw verpakt (<10 nm gemeten) in het postsynaptische membraan. De precieze donor en acceptor fluorofoor combinatie is essentieel voor dit FRET assay. Namen en gegevens van de fluoroforen worden gegeven in de tabel Materials. De spectrale eigenschappen met betrekking tot FRET, worden besproken in onze vorige papieren 14,15.
- Bereid je vast dwars cryocoupes zoals beschreven in paragraaf 6.1. Alle sample groepen moeten samen en imago worden verwerktd dezelfde confocale sessie.
- Meng 2,5 g / ml rood-BGT (FRET donor) met 10 g / ml ver rood-BGT (FRET acceptor) met blokkerende oplossing in een kleine plastic buis pipetteren en neer 12 keer. Deze 1: 4 molair mengsel maximaliseert de efficiëntie van FRET 14.
- Plaats elke dia in een bevochtigde kamer zorgvuldig bedekken elke sectie met een druppel (12 pl) van het bovenstaande mengsel en incubeer gedurende 1,5 uur bij KT.
- Controle secties: dekking klein aantal secties met 2,5 g / ml rood-BGT (alleen donor, gelabeld C1 controles) en ook enkele secties met 10 g / ml ver-rood-BGT (alleen acceptor; gemerkte C2 controles). Incubeer deze controles als bij stap 7.3.
- Was glijdt 3 x 10 min in PBS en monteren in-glycerol gebaseerde, fade-weerstand montage medium (zie stap 3.12).
- Voeren bemonstering van eindplaten als in stap 6.7. De fluorescentie van de donor en acceptor dienen perfect op elkaar gelokaliseerd op eindplaten door de willekeurige binding van de fluorescerende moleculen BGT.
- Controle beelden: Met behulp van de 40x objectief en lage laservermogen optimaliseren redBGT gain en offset niveau instellingen voor eindplaten uit een controle dia C1. Optimaliseer ver-redBGT gain en offset niveaus voor eindplaten van controle dia C2. Bevestigen de afwezigheid van fluorescentie doorbloeding.
- Zonder wijziging van het laservermogen, winnen of offset-niveau-instellingen te verplaatsen naar de experimentele dia's en het verzamelen van beelden (pre-photobleach) voor zowel fluorescentie kanalen.
- Selectief photobleach de ver-rode-BGT over een gedeelte van een eindplaat van inzoomen het scangebied dan scannen 10 maal met de 633 nm laser bij 100% vermogen. De fluorescentie in het gescande gebied moet gedimd worden.
- Reset het laservermogen en zoom en het verzamelen van post-bleach beelden op beide fluorescerende kanalen met de confocale instellingen op 7,7 vastgesteld.
- Bereken de FRET efficiëntie (E) van de procentuele stijging van de donor (rood-BGT) fluorescentie na photobleach van de acceptor (ver-rood-BGT) volgens de volgende formule *:

* Voor alle situaties waarin de fluorescentie van de donor vergroot na fotobleken de acceptor.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Meting van Synaptic Area op de NMJ
Elk geraamde gebied vertrouwt op de tekening van een grens aan de mate van synaptische specialisaties definiëren. Bij gezonde jonge volwassen spieren moeten NMJ beelden goed gedefinieerde grenzen voor zowel AChR en synaptofysine kleuring (Figuur 2A en B) weer te geven. Fluorescentie-intensiteit voor zowel AChR en synaptofysine stijgt scherp aan de grens tussen de peri-synaptische en synaptische gedeelte van de motor eindplaat (figuur 5A en B). Voor dergelijke beelden een minimumdrempel (net boven extrasynaptic achtergrond fluorescentie) zal gemakkelijk isoleren AChR-rijk of synaptophysine-rijk gebied van de eindplaat (horizontale streeplijnen in (figuur 5A en B)). In de leeftijd van muizen en in sommige ziektebeelden eindplaat kleuring voor AChR kan minder intens zijn, kan AChR cluster randen verschijnen wazig en dere kan zijn hogere niveaus van extrasynaptic autofluorescentie (Figuur 5D; 17,22). Fluorescentie kleuring met onduidelijke grenzen kunnen fouten introduceren in schattingen van synaptische gebied. In alle situaties is het doel om een drempel die een binair beeld gelijk in vorm en grootte van de AChR- of synaptofysine-rijke gebieden oplevert als ze lijken voor het oog in de originele, afbeelding de continue toon te kiezen. Het uitvoeren van de analyse blind voor de behandelingsgroep moet het risico van persoonlijke bias de drempelwaarden (stap 4.1) verminderen. Soms vaag of wazig eindplaat beelden kunnen het gevolg zijn van sub-optimale verwerking. Figuur 2C en D toont een voorbeeld afbeelding van een lage kwaliteit eindplaat van een gezonde 2 maand oude muis. Onscherpe randen en vage synaptofysine vlekken kunnen zijn ontstaan in dit geval van gedeeltelijke ontdooien en opnieuw bevriezen van de spieren voorafgaand aan cryosectioning. Sommige delen van jonge gezonde (positieve controle) spier moet worden gesegmenteerd eennd parallel verwerkt met proefmonsters zodat eventuele bijzondere zichtbaar in de NMJ beelden niet vanwege problemen met immunokleuring. Batches van beelden gecompromitteerd door sub-optimale verwerking moet uit analyses worden uitgesloten.
Voor en face z-stack beelden, 15-20 eindplaten is een redelijke steekproefomvang voor het schatten van synaptische gebieden. Een grote diversiteit in de vormen en afmetingen van NMJs worden gevonden binnen een bepaalde spier. Scatterplots onthullen een groot bereik in de AChR-rijk gebied tussen eindplaten van de tibialis anterior spier van elke individuele muis (Figuur 6A). Toch is de gemiddelde AChR gebied (gebaseerd op 15-20 en face eindplaat beelden) vergelijkbaar in zeven monster muizen (-200 m 2. Figuur 6A). Het gebied van de eindplaat synaptofysine vlekken aanzienlijk varieerde tussen eindplaten van een bepaalde spier. Nogmaals, het gebruik van een steekproef van 15-20 eindplaten de gemiddelde oppervlakte van de eindplaat synaptofysine was simiLAR onder de 7 muizen bestudeerd (~ 170m 2; Figuur 6B). Frequentiehistogrammen van gepoolde gegevens bleek ruwweg normale verdelingen voor het gebied van de eindplaat AChR en synaptofysine (Figuur 6A 'en B'). Maar een normale verdeling van synaptische zones niet bij ziektetoestanden kan worden aangenomen zoals myasthenia gravis 16,20. Dit kan de keuze van de statistische toets beïnvloeden.
Tabel I geeft gebieden van pre- en post-synaptische specialisaties voor NMJs voor gezonde 2 maanden oud (jong volwassen) vrouwelijke C57Bl / 6J muizen uit eerdere studies. De gebieden van zowel pre- en postsynaptische specialisaties daalde met sedentaire veroudering 22. AChR was ook duidelijk bij muizen geïnjecteerd met IgG van anti-MuSK myasthenia gravis patiënten verminderde 17,21. Myastheen muizen behandeld met de cholinesterase inhibitor geneesmiddel, pyridostigmine, toonde een verdere aanzienlijke vermindering eindplaat AChR gebied 20 </ Sup>.
Relatieve intensiteit van Kopplaat fluorescentielabeling
De relatieve intensiteit van immunofluorescentie labeling kan veranderingen in de dichtheid van een synaptisch eiwit van belang met de leeftijd, genotype en / of ziektetoestand onthullen. AChR fluorescentie (rood-BGT of ver-rood-BGT) wordt eerst gebruikt om de locatie van de NMJ definiëren. De helderheid van fluorescentie in de 8-bit AChR-rijk gebied wordt vervolgens gebruikt om veranderingen in de concentratie van het eiwit van belang in verhouding tot controledieren beoordelen. In dwarsprofielen de eindplaat AChRs typisch weergegeven als een halve maan vorm, maar deze vorm is vaak onregelmatig (Figuur 4A, C, E, H). Lage intensiteit achtergrond fluorescentie brengt gewoonlijk of een patch van AChR kleuring een enkele eindplaat, of twee dekplaten gelegen op aangrenzende spiervezels. Veel synaptische eiwitten (zoals rapsyne, muskus en SRC) zijncolocalized met AChR op de eindplaat (Figuur 4A - D). Immunofluorescentie kleuring met fosfo-specifieke antilichamen kunnen ook worden gebruikt om het effect van experimentele ingrepen op de fosforylering status van bepaalde postsynaptische membraan eiwitten 21 vergelijken.
De betrouwbaarheid en reproduceerbaarheid van de fluorescentie-intensiteit meting sterk afhankelijk is van de integriteit van de bevroren spieren en de kwaliteit van de immunokleuring. Spieren moeten worden ontleed en onmiddellijk ingevroren of-paraformaldehyde gefixeerd (binnen enkele minuten na de dood van het dier) om degeneratieve veranderingen in de NMJ voorkomen. Immunokleuring hangt sterk af van de kwaliteit van de reagentia en het optimaliseren van de kleuring protocol voor specifieke antilichamen. Voor elke nieuwe partij van primair antilichaam proef immunokleuring experimenten nodig. Vers gesneden cryosecties van gezonde jonge spieren geïncubeerd met seriële 2-voudige verdunningen van het primaire antilichaam. Eenbekend betrouwbare secundair antilichaam gebruikt en de resultaten vergeleken. Als het proteïne van interesse bekend te worden beperkt tot de NMJ dan de beste antilichaamconcentratie is dat met de hoogste verhouding van NMJ fluorescentie-intensiteit ten opzichte van die in extrasynaptic delen van de spier (achtergrond fluorescentie) oplevert. Extrasynaptic (vermoedelijk niet-specifieke) fluorescentie-intensiteit normalerwijze niet langer dan 15% van de eindplaat fluorescentie-intensiteit. Zo ook 'nee primaire antilichaam control' secties (alleen geïncubeerd met secundair antilichaam) moeten donker lijken, wat bevestigt dat het secundaire antilichaam niet niet-specifiek bond. De kwaliteit van de verschillende partijen (polyklonale) secundaire antilichamen kunnen sterk zodat alternatieve secundair antilichaam batches variëren moeten worden vergeleken vóór de oprichting van een standaard protocol. De ideale test voor de specificiteit van immunofluorescentie gaat om een directe vergelijking van de secties van wild-type muizen en negatieve controle secties van muizen thoed missen de eiwit-of-interest (knockout). De volgende referenties beschrijven kwantificering van eindplaat fluorescentie kleuring voor muskus, rapsyne, dystroglycan, src en AChR 13,14,18,21.
Kleine steekproeven introduceren fout in de schattingen van het relatieve fluorescentie-intensiteit. Individuele eindplaten aanzienlijk varieerde in fluorescentie-intensiteit. Vermoedelijk deze variabiliteit tussen eindplaten binnen een bepaalde spier weerspiegelt de diversiteit van NMJ structuur en de kans op verschillen in het optische gedeelte bemonsterd. Echter, het verhogen van het aantal eindplaten bemonsterde resulteert in een stabiele schatting van de gemiddelde fluorescentie-intensiteit (Figuur 7). Een schatting van de gemiddelde eindplaat fluorescentie-intensiteit 40-60 eindplaten van elke spier monster moet worden gemiddeld.
AChR-AChR FRET
Elke AChR een pentameer met twee bindingsplaatsen voor BGT (gelegen aan elke alfa-subeenheid). Binding vanrood-BGT en ver-rood-BGT op deze twee locaties zou een donor-acceptor scheiding van ongeveer 9nm 28-30 opleveren. Zo laag rendement FRET kan worden gedetecteerd, zelfs voordat AChRs assembleren in clusters 14. De efficiëntie van FRET bij muizen eindplaten ruwweg verdubbeld postnataal overeenstemming met de efficiëntere integratie van AChRs in een overvolle postsynaptische membraan rooster 14. Met rood-BGT en far-red-BGT als FRET-donor en acceptor (respectievelijk), de eindplaten van 1-2 maanden oude muizen geproduceerd gemiddelde FRET efficiëntie variërend 20-37% (tabel 2). FRET efficiëntie van 20% of meer worden gedacht aan strakke verpakking van AChRs 14 vertegenwoordigen. Eindplaat FRET efficiëntie werd enigszins verminderd na denervatie 14, en duidelijk verminderd na de muizen werden geïnjecteerd met IgG van anti-MuSK-positieve myasthenia gravis patiënten 18. Bij deze aandoeningen waarbij afzonderlijke AChRs minder strak verpakt in de postsynaptic membraan steiger door de postsynaptische MuSK- / rapsyne systeem 31.
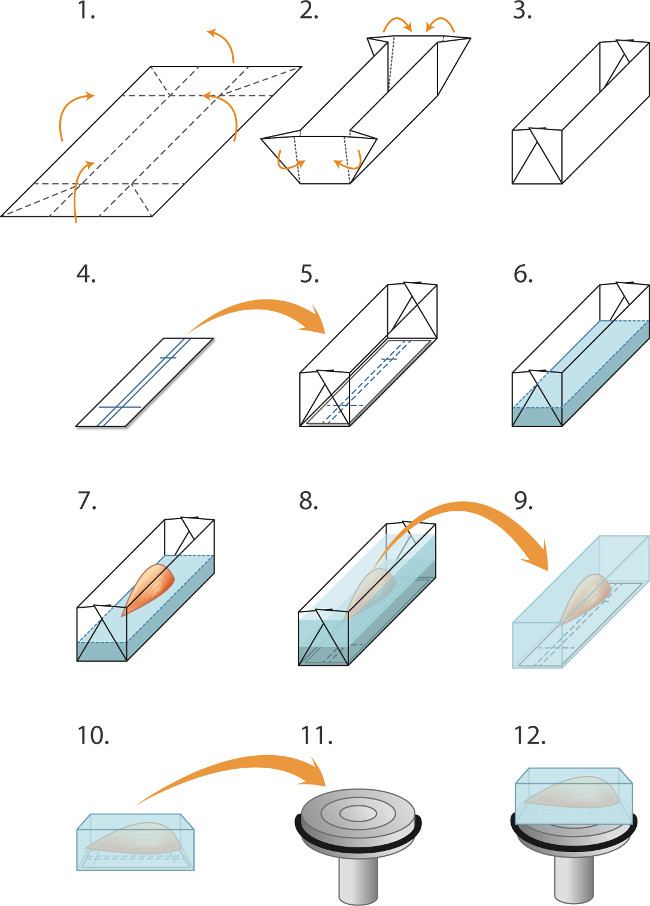
Figuur 1. inbedden en bevriezen spieren cryosectioning (1-5) Voorbereiden mallen ("boten") voor het invriezen een partij spieren. (1) Aluminium folie wordt gesneden in rechthoeken (2,0 x 3,0 cm) en (2). Gevouwen als aangeklaagd om een mal / boot te maken (3). (4) rechthoeken van nitrocellulose immunoblotting papier worden gesneden om te passen in de mal en een balpen wordt gebruikt om lijnen te oriënteren de spier regeren. (5) De nitrocellulose rechthoek wordt in de matrijs. (6-8) insluiten en bevriezen van de spieren: (6) cryostaat inbedding matrix vloeistof voorzichtig in de mal (bovenop de nitrocellulose) gegoten tot een diepte van 2 mm. (7) Schone tang worden gebruikt om de spier te verlagen, door de pees in de inbedding matrix, in lijn met de nitrocelluverliezen uitspraken, (8) Aanvullende inbedding matrix wordt voorzichtig in de mal gegoten om de spier te dekken, zorg om te voorkomen dat het creëren van bubbels. De geïntegreerde spier wordt vervolgens snel bevroren, verzegeld in buizen en opgeslagen bij -80 ° C zoals beschreven in de tekst. (9-12) Voorbereiding cryosectioning: (9) schone pincet gebruikt om afpellen de aluminium matrijs en het bevroren blok wordt vervolgens in de -20 ° C cryostaatkamer (10) De markeringen in het nitrocellulose gebruikt uitlijnen de spier evenwijdig aan het vlak van de klem (11) voor longitudinaal snijden (12). Een druppel vloeistof cryo inbeddingmedium wordt gebruikt om het blok te bevestigen aan de gekoelde boorkop. Stompe tang worden gebruikt om de bevroren spier blok manipuleren. Voor dwarsdoorsneden verkrijgen, wordt het blok plaats gemonteerd dat de spier loodrecht op het oppervlak van de boorkop (niet getoond).

Figure 2. En face beelden van NMJs van de tibialis anterior spier van 2 maand oud vrouwtje C57Bl6J muizen. De verse spier was snap bevroren en de secties gefixeerd op de dia zoals beschreven in stap 3.2. Maximale intensiteitsprojectie afbeeldingen z-stacks werden verkregen zoals beschreven in het eerste protocol. (A) rood-BGT kleuring onthult een enkele motor eindplaat uit twee sets van AChR-rijke primaire postjunctional goten (en vooraanzicht). (B) synaptophysine kleuring met FITC-geconjugeerd secundair antilichaam onthult de presynaptische zenuwuiteinde, het bezetten van de primaire synaptische goten. Een gedeelte van het pre-terminal axon is ook zichtbaar in het bovenste deel van het paneel. (C & D) Een voorbeeld van een slechte beeldkwaliteit NMJ vanaf jonge gezonde muis van. Kleuring is zwak en de grenzen van de pre- en postsynaptische specialisaties vervagen. Dit werd toegeschreven aan tekortkomingen in tHij verwerking van het weefsel en / of onvoldoende tijd voor het primaire antilichaam incubatie. Schaal bar in panel D staat voor 10 micrometer. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 3. Stappen in de verwerking van en face NMJ afbeeldingen (zoals beschreven in protocol 1). (A & B) Originele continue toon MIP beelden tonen rood-BGT fluorescentie onthullen AChR en groene immunofluorescentie voor respectievelijk synaptofysine. (C) AChR vlekken na conversie naar een Grijswaardenbeeld 8-bit en het gebruik van de veelhoek tool om de eindplaat af te bakenen (dunne gele lijn). (D) Kopplaat grenslijn overgedragen naar het groene synaptofysine. (E) Applicatie van een minimale intensiteit drempel opdracht om een binair beeld dat bovendrempelige rood-BGT (AChR) fluorescentie isoleert creëren. De sequentie van ImageJ gebruikersinterface schermafbeeldingen wordt weergegeven links van de beelden. Binaire beeld synaptophysine na toepassing van een afzonderlijke drempelwaarde (F). (G) Identificatie van discrete suprathreshold AChR-rijke domeinen in de eindplaat door toepassing van de deeltjes opdracht Analyseren naar het binaire beeld rood-BGT. De overeenkomstige ImageJ (links van paneel (G) toont de input data nodig. Dient de minimumomvang van het bovendrempelige pixelgebieden nodig worden ingevoerd. (H) Identificatie van gebieden van overlap van de binaire synaptofysine en AChR beelden. Overlap wordt vertegenwoordigd door witte pixels . ImageJ gebruikersinterface screenshots (hieronder paneel H) tonen de stappen om te komen tot het de binaire overlap. De gekozen minimale intensiteit drempelwaarden voor elke fluorescentie channel moet worden ingevoerd in het venster 'colokalisatie'. Schaal balk vertegenwoordigt 10 micrometer. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 4. Voorbeelden van dwarsrichting optische secties gebruikt om relatieve fluorescentie intensiteiten vergelijk (protocol 2). De spier werd snel bevroren en in de dia vaste delen zoals beschreven in stap 3.2. (A & B) Een enkele eindplaat dubbel gelabeld met ver -Red-BGT (AChR, hier afgebeeld in blauw pseudocolor) en FITC-anti-rapsyne illustreert de co-lokalisatie van deze twee interacterende eiwitten in het postsynaptische membraan (C & D) Twee eindplaten op aangrenzende spiervezels scherm co-gelokaliseerde AC. HR- en muskus. (E & F) Een eindplaat dubbel-gelabeld voor AChR en -dystroglycan (-DG). De -DG strekt zich rond de spiervezels perimeter, maar is verrijkt op de eindplaat (schaal bar in F, voor panelen AF: 25 pm) (G - I) isoleren van een eindplaat voor intensiteit meting (G) Een typische microscoop veld.. met drie ver-rood-BGT-gekleurd eindplaten (schaal bar in panel G: 40 micrometer (H) Een vergroot beeld van de boxed eindplaat (I) Hetzelfde eindplaat omgezet naar een 8-bits beeldverwerking grijstinten en afgebakend met behulp van de veelhoek.. . instrument van ImageJ (dunne gele lijn) wordt Gemiddeld fluorescentie-intensiteit gemeten binnen deze grenslijn. (schaal bar voor H & I: 10 pm) Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 5. Invloed van de beeldkwaliteit in de beoordeling synaptische gebied. (A & B) Hoge kwaliteit en gezichtsbeelden van een gezonde NMJ van een 2 maanden oude muis on the red-BGT en anti-synaptophysine fluorescentie kanalen. (A & B ) fluorescentie intensiteitprofielen overeenkomt met de lijn over de eindplaat respectievelijk A en B. De horizontale rode stippellijn geeft de minimale drempel wordt gebruikt om de binaire beeld te creëren. (C & D) Kopplaat van een oudere muis. Eindplaat synaptophysine kleuring algemeen minder intens. (C 'en D') Intensiteit profielen vertonen een hoog extrasynaptic (baseline) fluorescentie fluctuatie in de synaptofysine (FITC) kanaal (backgrond) waarbij de vaststelling van een geschikte drempel beïnvloedt. Een groot deel hiervan is breed-spectrum weefsel autofluorescentie. Schaalbalken vertegenwoordigen 10 um. Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 6. De variabiliteit van synaptische gebieden onder NMJs binnen een spier en tussen muizen. (A) scatterplots tonen de totale AChR-rijk gebied van eindplaten van de tibialis anterior spier van zeven naïeve 2 maand oud vrouwtje C57Bl / 6J muizen verkregen door auteur NT. Elk symbool staat voor een eindplaat. Elke staaf vertegenwoordigt het gemiddelde ± SD voor eindplaten bemonsterd uit een muis. (B) scatterplots met de synaptofysine-rijk gebied voor dezelfde eindplaten. (A ') (B ') Frequentie distributie voor de synaptofysine-rijk gebied van eindplaten (gepoolde gegevens). Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 7. Effect van monstergrootte op schattingen van eindplaat fluorescentie-intensiteit. Transversale optische coupes werden gebruikt om de fluorescentie-intensiteit (in willekeurige eenheden) 40 60 eindplaten van de tibialis anterior spier van een gezonde 2 maanden oude muis te meten. Cumulatieve gemiddelden wordt uitgezet tegen het aantal eindplaten opgenomen in het gemiddelde. (A) Kopplaat rood-BGT fluorescentie-intensiteit verkregen door auteur AV. (B) Anti-synaptophysin immunofluorescentie intensiteit voor dezelfde eindplaten als in panel A. (C) Kopplaat ver-rood-BGT fluorescentie-intensiteit verkregen uit een tweede spier monster door auteur NG. (D) Anti-rapsyne immunofluorescentie intensiteit van dezelfde eindplaten als in panel C.
| Studie | Spier | aantal muizen | AChR gebied | Synaptofysine gebied | Overlapgebied |
| gemiddelde ± SD (pm2) | gemiddelde ± SD (pm2) | gemiddelde ± SD (pm2) | |||
| Morsch et al. (2012) 1 | gastrocnemius | 3 | 181 ± 7 | 163 ± 24 | |
| middenrif | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch et al. (2013) 1 | tibialis anterior | 3 | 166 ± 26 | 117 ± 21 | nd |
| Cheng et al. (2013) 2 | tibialis anterior | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| Tse (ongepubliceerd) 2 | tibialis anterior | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| 1 afgebeeld op een Zeiss LSM 510 Meta microscoop, maar met vaste versterking en offset niveaus. Thresholding en synaptische gebieden metingen gebruikt Metamorph software door MM. | |||||
| 2 afgebeeld op een Leica DM IRE2 microscoop en geanalyseerd volgende huidige protocol door de aangegeven eerste auteur, blind voor de behandeling groep. | |||||
| nd niet bepaald. | |||||
Tabel 1. Synaptic gebieden voor NMJs van 2 maand oud vrouwtje C57Bl / 6J muizen (gezonde controles)
| Studie | Spier | FRET efficiëntie (%) * | Bereik (%) |
| (Gemiddelde ± SEM) | |||
| Brockhausen et al. (2008) | tibialis anterior | 24 ± 1 | niet beschikbaar |
| Cole et al. 2010 | tibialis anterior | 26 ± 1 | 22 - 30 |
| Morsch (niet gepubliceerd) | middenrif | 37 ± 1 | 24-47 |
| Ghazanfari (niet gepubliceerd) | tibialis anterior | 30 ± 1 | 20-45 |
| * FRET tussen rood-BGT en ver-rood-BGT (Förster straal voor FRET paar = 51A (Life Technologies). | |||
| nvt gegevens niet beschikbaar | |||
Tabel 2. Rendementen voor AChR FRET van jong volwassen muizen C57Bl6J muizen
Subscription Required. Please recommend JoVE to your librarian.
Discussion
De hier beschreven protocollen hebben ons in staat om betrouwbaar te meten en te kwantificeren veranderingen in de eigenschappen van de NMJ in een heel scala van voorwaarden, met inbegrip van de normale veroudering en de ziekte van staten. De beschreven voor en methoden geconfronteerd NMJ beelden kunnen de onderzoekers het gebied van pre- en postsynaptische specialisaties en het gebied van de synaptische overlap / uitlijning te vergelijken. Het vergelijken van de relatieve intensiteit van pre- en postsynaptische eiwitten het tweede protocol, dat dwars optische secties gebruikt, de voorkeur. Het derde protocol test met name veranderingen in de nabijheid van de verpakking van AChRs in het postsynaptische membraan.
Specificiteit controles zijn van vitaal belang in immunofluorescentie microscopie. Bij gebruik van een primair antilichaam voor indirecte immunofluorescentie moet eerst zorgen dat het specifiek bindt aan het doeleiwit in de spier secties. Verschillende soorten weefsel verwerking en fixatie kan differentieel veranderen de specificiteit vanantilichamen. Het is belangrijk dat immunofluorescentie kleuring (voor zeggen rapsyne) echt is geconcentreerd bij AChR op de motor eindplaat bevestigen. Negatieve controle elementen moeten worden gecontroleerd om te verzekeren dat het antilichaam binding specifiek is. / - - Muizen bijvoorbeeld, zou de beste negatieve controle voor rapsyne immunofluorescentie secties van rapsyne zijn. Deze moeten niet eindplaat kleuring met anti-rapsyne tonen. Niet-specifieke fluorescentie kunnen ook voortvloeien uit endogene fluorescerende stoffen in het weefsel (autofluorescentie) of voor niet-specifieke binding van het fluorescerende secundaire antilichaam conjugaat. Deze fluorescentie wordt vaak verergerd door aldehyde fixatie. Daarnaast kunnen TRITC-BGT kleuring van eindplaten soms gedetecteerd in de FITC fluorescentie kanaal en deze fluorescente doorbloeding kan worden aangezien specifieke FITC immunofluorescentie. Om te waken tegen de laatste drie vormen van niet-specifieke fluorescentie, moet elke partij van dia's die worden gekleurd ook enkele 'no-primaire antilichaam control 'secties (stappen 3.7 en 4.6). Afbeeldingen van eindplaten van deze besturingssecties worden vergeleken met die van de experimentele dia te zorgen dat de indirecte immunofluorescentie kleuring van NMJs werkelijk weerspiegelt de binding van het primaire antilichaam.
Transversale confocale secties zijn bijzonder nuttig is om verschillen in de relatieve intensiteit van immunokleuring in de synaps. In dwarse confocale secties is gemakkelijker nauwkeurig co-localisatie van synaptische eiwitten beoordelen. De halvemaanvormige eindplaat profiel vertegenwoordigt slechts een voorbeeld cut-through van de NMJ in kwestie. Echter, de achtergrond (extrasynaptic) fluorescentie algemeen lager in vergelijking met een eigen gezicht z-projectie beelden. Zo kan het gemakkelijker zijn om 'echte' (specifieke) immunostaining discrimineren en stellen vast confocale gain en offset waarden met dwarse optische secties 13-15,18. Bijvoorbeeld, in een muismodel van myasthenia gravis (Where eindplaat AChR kleuring is duidelijk verminderd) eindplaten werden duidelijk afgebakend in transversale optische secties 18,21. Verschillen in de gemiddelde intensiteit van de fluorescentie in de NMS waarschijnlijk veranderde dichtheid van het doeleiwit in de synaptische specialisatie weerspiegelen. Een nadeel is dat, in sommige gevallen, een structurele verandering van het doeleiwit of occlusie van antilichaambinding door naburige eiwitten kunnen veranderde kleurintensiteit verklaren.
Het ontwerp van experimenten vereist enige overweging. In veel gevallen zou het experiment doel de invloed van een transgen gen knockdown of ziektetoestand van de grootte van de NMJ testen. De experimentele groep monster kan vervolgens worden vergeleken met gezonde jonge (wild-type) muizen van hetzelfde geslacht en genetische achtergrond. Basislijn waarden voor het gebied van eindplaat synaptofysine, AChR en synaptische overlap meerdere spieren worden gegeven in Tabel 1. Sample size zal afhangen van de graad van dierlijke-om-dier variatie binnen behandelgroepen en de grootte effect (verschil in middelen voor de experimentele versus controle groepen per standaarddeviatie). Wanneer de analyse wordt beperkt tot een goede kwaliteit foto's een redelijke mate van consistentie werd gevonden in de steekproef betekent voor eindplaat gebieden bij gezonde 2 maand oud vrouwtje C57Bl6J muizen (Figuur 6A en B). Aldus was het mogelijk om significante verlagingen 30-40% in synaptische gebied muizen geïnjecteerd met IgG van anti-MuSK-positieve myasthenia gravis tonen patiënten in vergelijking met controles met een steekproefomvang van drie muizen 17,20,32. Bejaarde muizen getoond groter dier op dier variatie in eindplaat parameters dan jonge muizen 22. Bijgevolg experimenten met verouderde muizen misschien grotere steekproefomvang nodig.
Als de primaire zorg is de grootte van de en face eindplaat dan de versterking en offset-instellingen gemeten moet worden geoptimaliseerd voor elk individu NMJ. Individual NMJs kan aanzienlijk de helderheid van AChR en synaptofysine kleuring, vooral wanneer ziektetoestanden onderzocht variëren. Bovendien is de intensiteit van extra-synaptische (aspecifiek) fluorescentie vaak hogere en variabele spieren van veroudering dieren vergeleken met die van gezonde jonge dieren (Figuur 5C en D). De 1-256 grijsschaal volledig worden benut om de tonale die in de uiteindelijke opnamen behouden maximaliseren. Dit zal betrekken aangepaste versterking en offset niveaus voor elke NMJ waarvoor een z-stapel wordt opgevangen. Figuur 5D toont een voorbeeld van een NMJ afbeelding waar tonale kritisch afbakening van het gebied van pre- en post kan synaptische specialisaties.
Metingen van synaptische zones kan worden toegepast op verschillende spier preparaten en experimenten. De meeste van onze metingen van synaptische gebieden zijn longitudinale cryocoupes in dienst van snap bevroren spieren. Bevriezing de spier vóór fixatie handhaaft de antigeniciteit van een groot aantal eiwitten. Voor zover verenigbaar met het antigeen, paraformaldehyde fixatie en sucrose infiltratie voorafgaand aan cryosectioning (stap 2.1) kan beter behoud van de NMJ structuur te bieden. Optimale structurele conservering kan worden verkregen door cardiale perfusie met paraformaldehyde. Artefacten van invriezen en snijden kan geheel worden vermeden door labeling eindplaten op het oppervlak van de intacte spieren en imaging NMJs op bundels geplaagd van de vaste spier 21. Ongeacht de voorbereiding, de procedures voor bemonstering, beeldvorming en omgeving kwantificatie blijven ongewijzigd (protocol stap 4-5). Consistente toepassing blinde bemonstering, beeldvorming en analyse protocollen (met verschillende medewerkers, verschillende monsters van muizen en verschillende tijden), kan leiden tot redelijk reproduceerbare waarden (vergelijk Cheng et al. En Tse resultaten in tabel 1).
"> Zijn Eindplaten omschreven als worden gefragmenteerd in diverse ziektetoestanden. Bijvoorbeeld in oude muizen spieren, sporadische degeneratie van een spiervezel (gevolgd door de regeneratie) resulteerde hermodellering van de pretzel-achtige eindplaat AChR plaque vormen meerdere kleinere AChR clusters 6. Bij muizen geïnjecteerd met IgG van anti-MuSK- myasthenia gravis patiënten, fragmentatie van de eindplaat was nogal verschillend. De eindplaat AChR krakeling grotendeels verspreid, met achterlating van een constellatie van kleine (<4 m 2) AChR 'micro-aggregaten '20,21. Deze twee voorbeelden benadrukken de noodzaak om de grootte verdelingen van AChR clusters vergeleken en eindplaten controle versus proefdieren 21.Andere werkwijzen zijn beschreven voor de beoordeling van synaptische gebied of kleuringsintensiteit in de NMS. Motor eindplaten kan soms worden gevouwen zodat de tweedimensionale z-projectie hier gebruikte beelden syn kunnen onderschattenaptic gebieden. Driedimensionale confocale reconstructies zou nauwkeuriger maatregelen verstrekken als absolute synaptische gebied moet worden afgebakend 33. Een belangrijk voordeel van de z-uitsteeksel hier beschreven protocol, echter de relatieve eenvoud, waarbij grote aantallen eindplaten heeft toegestaan gemeten vanaf verschillende behandelingsgroepen en betrouwbare identificatie van mogelijke veranderingen. Het protocol voor het vergelijken eindplaat kleuring intensiteit kan worden aangepast voor het bestuderen van veranderingen in de niveaus van verschillende synaptische eiwitten. De werkwijze is echter beperkt door het vereiste dat alle monsters bewerkt voor immunofluorescentie dan afgebeeld tijdens dezelfde confocale sessie. Een recente studie van Yampolsky et al. 5 een werkwijze beschreven voor het meten van eindplaat AChR dichtheid die kunnen helpen overwinnen van deze beperking. In deze studie werden dezelfde gebieden eindplaten afgebeeld op verschillende laservermogen instellingen. De helling van de relatie tussen laservermogen en rhodamine-BGT fluorescentie-intensiteit werd gebruikt om de relatieve veranderingen in AChR dichtheid bij eindplaten beoordelen in verschillende muizen 5. Deze methode zou nuttig zijn voor het vergelijken van AChR intensiteit monsters afgebeeld op verschillende tijdstippen in de loop van een langdurige studie.
AChR-AChR FRET biedt specifieke en aanvullende informatie over de organisatie van de eindplaat AChRs. Elektronenmicroscopische autoradiografie met 125. I-α-BGT heeft aangetoond AChRs stevig te verpakken met een vlakke dichtheid van 10 4 m -2 onmiddellijk onder elke presynaptische website van de zender release, terwijl aangrenzende membraan infoldings bevatten veel lagere dichtheden van AChR 34. AChR-AChR FRET maakt het relatief eenvoudig om (sub-microscopische) veranderingen in AChR verpakking te detecteren. Een vermindering FRET efficiëntie weerspiegelt een submicroscopische herverdeling van AChRs in het postsynaptische membraan die niet kunnen worden gedetecteerd door een verandering van de gemiddelde fluorescentie-intensiteit BGT. Meerdere factoren kan een wijziging in de efficiëntie van FRET leiden. Deze omvatten de donor-acceptor afstand en relatieve oriëntaties en de moleculaire omgeving 35,36. Een vermindering eindplaat FRET efficiëntie mogelijkerwijs voortkomen uit een verandering in de geometrie van de AChR rooster. Echter, waarschijnlijk zou worden veroorzaakt door een lager percentage van AChRs die zijn verpakt in nanoschaal postsynaptische moleculaire rooster 14.
Verlies van de verbinding tussen motor neuronen en spiervezels lijkt de directe oorzaak van spierzwakte in motorneuronziekte en in sedentaire veroudering 9,22,23 zijn. Gedeelde methoden en parameters voor het meten van NMJs moet het gemakkelijker maken voor verschillende onderzoeksgroepen te vergelijken en het contrast gepubliceerde bevindingen. Het delen van gedetailleerde protocollen (en toekomstige verbeteringen op hen) kan helpen versnellen vooruitgang in het begrip van de mechanismen van NMJ onderhoud en hoe het kan worden aangetast bij de ziekte van staten.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
