

Abstract
Den neuromuskulære junction (NMJ) er den store, cholinerge relæ synapse hvorigennem pattedyr motoriske neuroner styrer frivillige muskelsammentrækning. Strukturelle ændringer på NMJ kan resultere i neurotransmission fiasko, hvilket resulterer i svaghed, atrofi og endda død af muskelfibrene. Mange undersøgelser har undersøgt, hvordan genetiske ændringer eller sygdom kan ændre strukturen af muse NMJ. Desværre kan det være vanskeligt direkte at sammenligne resultaterne fra disse undersøgelser, da de ofte anvendes forskellige parametre og analysemetoder. Tre protokoller er beskrevet her. Den første anvender maksimale intensitet projektion konfokale billeder at måle arealet af acetylcholin-receptor (AChR) -rige postsynaptiske membran domæner på endepladen og området af synaptisk vesikel farvning i overliggende præsynaptiske nerve terminal. Den anden protokol sammenligner de relative intensiteter af immunfarvning for synaptiske proteiner i den postsynaptiske membran. Den tredje protocol bruger fluorescensresonansenergioverførsel (FRET) til at detektere ændringer i pakning af postsynaptiske AChRs på endepladen. Protokollerne er blevet udviklet og forfinet gennem en række undersøgelser. Faktorer, der har indflydelse på kvalitet og sammenhæng i resultaterne diskuteres og normative data leveres for NMJs hos raske unge voksne mus.
Introduction
Den neuromuskulære junction (NMJ) er den kritiske relæ synapse, der medierer kommunikation mellem nervesystemet og skeletmuskulatur. Det er nødvendigt for alle frivillige bevægelser. Fluorescensmikroskopi har længe været anvendt til at undersøge virkningerne af transgener på musen NMJ 1-3 eller for at sammenligne virkningerne af alder, kost, motion og sygdom ved gnavere NMJs 4-11. Sådanne undersøgelser har lært os meget om fysiologi og patofysiologi af NMJ, men de forskellige parametre rapporteret (f.eks AChR område, endepladens område, perimeter længde, fragmentering indeks) ofte gør det vanskeligt at sammenligne resultaterne af disse undersøgelser. Der er en stigende forventning til prækliniske forskere til at kunne påvise reproducerbarhed, især i studier med gnavermodeller på sygdom 12. De her beskrevne protokoller blev forfinet gennem en række undersøgelser, der undersøgte udviklingsmæssige, fysiologiske og patofysiologiske lmanges til NMJ. Sådanne undersøgelser kræver måling af arealet af synaptiske specialiseringer på musen motor endeplade og den relative tæthed af pakning af synaptiske proteiner i postsynaptiske specialer 13-15.
Anvendeligheden af disse metoder er illustreret af nylige undersøgelser i en musemodel af anti-moskus myasthenia gravis. Daglige injektioner af IgG fra anti-Musk-positive myasthenia gravis patienter i voksne mus fik dem til at blive svag inden for 2 uger 16. Konfokale maksimal projektion billeder af muskel sektioner, der var dobbelt-mærket til synaptophysin (i nerve-terminaler) og postsynaptiske AChRs viste en gradvis nedgang i området AChR farvning som det primære ændring. Vigtigere hastigheden af faldet var tilstrækkelig til at forklare sammenlignelige fald i amplituden af synaptiske potentialer, svigt af synaptisk transmission og muskelsvaghed 17,18. Kvalitativt lignende resultater blev rapporteret af andre forskergrupper10,19. Er siden blevet brugt De samme NMJ målemetoder til at vurdere virkningen af tre lægemidler til behandling af anti-Musk myasthenia gravis i denne musemodel 20,21.
Stillesiddende aldring kan føre til tab af neuromuskulære forbindelser. De her beskrevne protokoller har afsløret en alder-associeret nedgang inden for nerve terminal synaptophysin ved motoriske endeplader som mus fremskridt i alderen. De samme metoder viste, at frivillig øvelse i høj grad kunne forhindre reduktion i præsynaptiske nerve terminalområde 22, i overensstemmelse med tidligere arbejde med andre grupper 4. Tab af neuromuskulære forbindelser forekommer også i SOD1G93A musemodel for amyotrofisk lateral sklerose 9,23.
Ovennævnte undersøgelser viser, at en række forskellige sundhedsmæssige betingelser kan føre til en reduktion i det område, enten før eller efter synaptiske specialiseringer på NMJ. Dette kan resultere i nedsat synaptisk sjovIndsatsen eller kan varsle fuldstændigt tab af den neuromuskulære forbindelse. Tre protokoller beskrives som tillader kvantificering af området og tæthed af synaptiske specialer. Formålet med den første protokol er at give en praktisk og reproducerbar måling af områderne før og efter synaptiske specialiseringer og deres tilpasning ved pattedyrs NMJs, anvendelse af fluorescens mikroskopi. Todimensionale maksimale projektion konfokale billeder og billedanalyse med NIH ImageJ bruges til at påvise ændringer i området synaptophysin farvning (synaptiske vesikler), postsynaptiske AChRs og synaptisk overlappende område. Konfokale imaging parametre (gain og offset-niveau) er optimeret for hver NMJ således at maksimere den visuelle information til at skelne det område af synaptisk specialisering. Neuromuskulær fiasko kan også skyldes ændringer i tætheden af postsynaptiske AChR og / eller andre synaptiske proteiner. Den anden protokol kan anvendes til at detektere ændringer i den relative massefylde af postsynaptiske proteiner, såsomsom moskus, RapSyn, dystroglycan, phosphoryleret Src kinase og phosphoryleret AChR 18,21.
I myasthenia gravis, en reduceret tæthed af AChR i den postsynaptiske membran er den umiddelbare årsag til synaptisk svigt og muskelsvaghed. Den tredje protokol beskriver en fluorescensresonansenergioverførsel (FRET) metode til at vurdere ændringer i nærhed af tilstødende AChRs inden postsynaptiske membraner 14,15. Denne metode detekterer energioverførsel mellem tilstødende AChRs mærket med fluorescerende-α-bungarotoxin (BGT). FRET forekommer kun, når de fluorescerende donor- og acceptor-prober er mindre end 10 nm fra hinanden. Dette kan afsløre (submikroskopiske) ændringer i tæthed AChR emballage, der direkte vedrører amplituden af synaptiske potentialer.
Disse tre protokoller, raffinerede i det seneste årti, giver supplerende foranstaltninger af NMJ integritet i en konsekvent og reproducerbar måde. Anvendelse af standardiserede protokoller and parametre bør lette sammenligning af virkningerne af gener og miljømæssige tiltag upon pattedyrs NMJ.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
BEMÆRK: Design, gennemførelse og anmeldelse af dyreforsøg bør tage hensyn til gældende retningslinjer 24. Dette arbejde skal godkendes på forhånd af den lokale dyrevelfærd myndighed (i vores tilfælde Animal Ethics Committee fra University of Sydney).
1. Aflivning af dyret og Muscle Dissection
- Overfør musen fra bedriften plads til et separat rum, hvor det er aflivet med en intraperitoneal injektion af pentobarbiton opløsning (30 mg / kg) ved hjælp håndtering musen metode beskrevet af Shimizu 25. Placer musen tilbage i sit bur.
- Når vejrtrækning af musen er stoppet i mere end 1 min, teste mund- tilbagetrækning refleks ved forsigtigt at klemme foden, og hornhindens refleks ved let børstning hornhinden. Først når refleks reaktioner er fraværende, kan musen være forberedt på dissektion.
- Kontakt en atlas af gnaver anatomi såsom Chiasson 26 og / eller søge hjælp hos en experienced anatom inden du forsøger dissektion af musklen af interesse. I hvert tilfælde fjerne hår fra den overliggende huden ved hjælp af en lille elektrisk barbermaskine før åbning af huden for at blotlægge musklen.
BEMÆRK: dissektion vil variere for hver anatomisk distinkt muskel. - Ved stump pincet frigøre muskel fra overliggende membraner og omgivende væv. Gribe og skære den distale senen at adskille muskel fra dets insertion.
- Forsigtigt drille og snip musklen fri omgivende væv helt tilbage til sin oprindelse. Kort placere nyligt dissekeret muskel i 0,1 M phosphatpufret saltvand (PBS) opløsning eller Ringers opløsning forud for yderligere forarbejdning.
2. Forberedelse muskel for Cryosectioning
BEMÆRK: Optimal strukturel konservering som tidligere beskrevet 27, eller nedsænkning fiksering (til små muskler) som beskrevet i valgfri trin 2.1 kan opnås ved hele dyret perfusion. Imidlertid4% paraformaldehyd fiksering kan forringe efterfølgende farvning med mange antistofprober og med fluorescerende-BGT. Glutaraldehyd især bør undgås. Hvis der ikke skal fastsættes muskler, skal de straks lynfrosset (fortsæt til 2.3).
- Valgfri nedsænkning fiksering: Pin musklen til voks i en petriskål på hvile længde. Dæk musklen med 2% w / v paraformaldehyd (frisk opløst i PBS) i 2 timer ved stuetemperatur. Vask det med 3 skift af PBS i 30 minutter (3 x 10 min) og derefter erstatte PBS med 30% vægt / volumen sucrose i PBS og inkuberes O / N ved 4 ° C.
- Foretag forme (»både«) i forvejen ved at folde 2 cm x 1,5 cm stykker af aluminiumsfolie, som vist i figur 1. Anbring et stykke nitrocellulosemembran i bunden af båden. Hæld forsigtigt cryostat indlejring matrix (Materialer tabel) i båden til en dybde på 2 mm, idet man undgår luftbobler. Placer musklen ind i båden, som bringer den med kuglepen linjer pånitrocellulose. Tilføj flere indlejring matrix så fuldstændigt dækker musklen (figur 1).
- Pre-label polypropylen rør med et uudsletteligt markør. Anbring en dråbe vand i hvert rør og chill røret i flydende nitrogen.
BEMÆRK: frosne vanddråbe opretholder damptrykket og forhindrer udtørring under langvarig -80 ° C opbevaring - Ved hjælp af en ansigtsmaske, tykke beskyttelseshandsker og et stort par stumpe pincet, delvist sænke en lille metal bægerglas (3 cm i diameter, 8 cm dyb) indeholdende 2 cm dybde af isopentan i en beholder med flydende nitrogen i 30 sek. Derefter fjernes bægerglasset og placere den på bænken toppen. Ved hjælp af en mindre par stumpe pincet placere formen indeholdende muskler og indlejring matrix i den kølede isopentan. Sørg for at undgå flydende nitrogen blanding med isopentan.
- Tillad 2 min for blokken til at fryse helt, før du bruger stumpe pincet til at løfte den frosne blok ud og forsegle det i den rigtige pre-mærket og præ-kølet rør (trin 2.3).
- Opbevar rørene midlertidigt i flydende nitrogen før overførsel til -80 ° C. Log alle prøver i et regneark for indhold fryser.
3. Cryosectioning og fluorescens farvning for en Face billeder af NMJs
- Skræl væk aluminium skimmel. Inden for -20 ° C kryostatkammeret vedhæfte den frosne blok til kryostaten chuck således at skære 20 um kryosektioner parallelt med den lange akse af muskelfibre (figur 1). Saml afsnittene om poly-L-lysin eller gelatineovertrukne objektglas.
- BEMÆRK: Udelad dette trin, hvis vævet er fastgjort inden frysning. Efter at have ladet 30 min for de dele tørre på dias, rette dem ved at placere en dråbe 2% paraformaldehyd i PBS over hver sektion i 15 minutter ved stuetemperatur.
- Wash slides 3 x 10 minutter i PBS i en Coplin krukke, og derefter nedsænkes objektglassene i PBS indeholdende 0,1 M glycin i 30 minutter for at blokere tilbageværende aldehydgrupper.
- Vask slides i 10 minutter i PBS, derefter nedsænkes i methanol (afkølet til -20 ° C) i 7 min. Denne permeabilization trin er en rutinemæssig del af dobbelt mærkning med fluorescerende-BGT og anti-synaptophysin men det kan påvirke immunfarvning for nogle andre proteiner.
- Vask slides 2 x 10 minutter i PBS derefter placere hvert dias i en stabil og nivelleret fugtigt kammer. Dække Umiddelbart hver sektion med 20 pi blokerende opløsning (0,2% Triton X-100, 2% bovint serumalbumin (BSA) i PBS) i 1 time ved stuetemperatur. Sektioner må ikke få lov til at tørre ud på noget tidspunkt under den immunfarvning proces.
- Udfør den primære inkubation: Tager et dias ad gangen forsigtigt fjerne det overskydende blokeringsopløsning fra over hver sektion og erstatte det med 20 pi kanin anti-synaptophysin (fortyndet 1: 200 i blokerende opløsning).
- Medtag en negativ kontrol dias, der vil blive inkuberet med kun blokerende opløsning. Denne "no-primært antistof kontrol'Er afgørende i alle immunfarvning løb.
- At tage sig at det primære antistof forbliver på plads i løbet af hvert afsnit, lukke fugtigt kammer og inkuberes i 1-2 dage ved 4 ºC.
- Undersøg hver sektion for at bekræfte, at det primære antistof forbliver på plads. Brug en Pasteur-pipette til forsigtigt skylle hver slide med PBS og læg den i en Coplin-skål. Vask alle dias 3 x 10 minutter i PBS.
- Foretag sekundær inkubation. Tager et dias ad gangen, forsigtigt fjerne overskydende PBS, lægge den i fugtigt kammer og dækker hver sektion med 20 pi af en blanding indeholdende FITC-konjugeret æsel anti-kanin IgG og BGT konjugeret til tetramethylrhodamin eller en anden rød fluorophor (TRITC- / redBGT; 5 g / ml) fortyndet med blokerende opløsning. Inkuber ved stuetemperatur i 2 timer.
- Vask glider 3 x 10 min i PBS i Coplin-skåle.
- Tager et dias ad gangen, forsigtigt fjerne overskydende PBS og montere med et dækglas under anvendelse af en minimal mængde af, glycerol-baseret, fade-modstand montering medium. Seal kanter dækglassene med klar neglelak. Lad det tørre hårdt.
- Opbevar dias i mørke ved 4 ºC i op til en uge, eller ved -20 ºC i længere perioder opbevaring (op til flere måneder).
4. Unbiased Sampling og En Face Imaging af motoriske endeplader
- Blind objektglassene ved mærkning hvert objektglas med en tilfældig kodenummer, forbliver kun kendes en anden forsker (ikke involveret i analysen). Som et resultat er operatøren fortsat blind for behandlingsgrupper indtil kvantificering af NMJ parametre er komplet for alle prøverne.
- Placer dias på mikroskopet scenen og se den under bredt felt belysning med TRITC filtersæt (63X olie 1,3 NA mål). Flyt gradvist (felt for felt) fra venstre til højre og tilbage, indtil en endeplade vises i feltet (figur 2A).
BEMÆRK: Sampling kriterium: Hver AChR-farvede struktur, der er relativtflad og vender mod målet (dvs. strækker <15 m i z-dimension) betragtes som en endeplade og afbildes til analyse (halvmåner af AChR farvning repræsenterer tværsnit gennem endeplader og kan derfor ikke). - Med det konfokale pinhole sæt til 1,0 Airy enhed og lav lasereffekt optimere gain og offset niveauer for TRITC / rød-BGT (532 nm laser) på endepladen, der skal afbildes. Næste optimere FITC / synaptophysin fluorescens under anvendelse af 488 nm laser. Saml en z-stak af endepladen med en 0,7 um interval mellem hver optisk skive. Gemme billederne med et filnavn, der indeholder datoen for imaging session, kodenavnet af slæden, og antallet af endepladen.
BEMÆRK: scanninger ved hjælp af 488 nm og 532 nm lasere (FITC og TRITC) skal indsamles sekventielt (ikke samtidigt) for at undgå forurening af FITC-kanalen ved fluorescens fra den røde fluorofor og omvendt (bløder-through). - Gentag prøveudtagning ennd billeddannelse af trin 4,2-4,3 indtil den 20. endeplader indsamles fra slide / prøve.
- Skift til den næste kodet dias og gentag 4,2-4,4. Gentag dette for hver af de kodede objektglas.
- Saml et par billeder af endeplader fra dias kontrollen (no-primært antistof kontrol) ved brug konfokale indstillinger, der blev fundet optimale for de eksperimentelle dias (FITC fluorescens kanal skal vises mørke).
- Ved afslutningen af det konfokale overførsel session billedfilerne til en anden computer, og tilbage op de originale filer på et eksternt drev eller server.
5. Måling af området af synaptisk Specialiseringer i en Face Images
- Brug NIH ImageJ freeware (http://imagej.nih.gov/ij/) at forberede maksimal projektion (MIP) billeder fra hver z-stak. Gem dem som TIFF-filer (figur 2A & B). Filnavne bør omfatte dato billedet session, kodeeksempler, endepladen nummer og fluorescerende kanal (f.eks 060414_5723_7_FITC.tiff).
- Åbn z-projektion billede i ImageJ. Vælg acetylcholin receptor billede kanal (figur 3A) og vælg: Billede> type> 8-bit til at konvertere 24-bit RGB farvet billede i tre 8-bit gråtonebilleder på skærmen.
- Brug af ImageJ polygon værktøj tegne et groft omrids omkring endepladen af interesse i redBGT farves (AChR) kanal til også at omfatte alle tilsyneladende farvede områder af særlige individuelle endepladen, og udelukker ethvert farvning, som ikke stammer fra endepladen af interesse ( figur 3C).
- Påfør et minimum intensitet tærskel til billedet ved at vælge: Image> Juster> Threshold (figur 3E og tilhørende ImageJ screenshots).
- Juster tærskelværdien for at isolere AChR-farvede dele samtidig udelukke omgivende baggrund signal som sub-tærskel (figur 3E). Åbn en anden vindow med originalen (kontinuerlig-tone) billede umiddelbart ved siden af vinduet til sammenligning, for at lette afgørelsen om tærskelværdien. Optag tærskelværdien til senere brug i colokalisering analyser.
- Bevarelse af den polygon kontur omkring endepladen vælge: Analyse> Analyser Partikler. I pop-up-menuen angive udvalg af størrelser som: 50 til uendeligt pixels (dette fjerner små artefakter skyldes elektrisk støj i fotomultiplikator).
- Analyser Partikler kommandoen genererer et vindue med en liste over diskrete overtærskel- områder og deres fluorescensintensitet værdier nummereret som de vises i det binære billede (figur 3G og tilhørende ImageJ screenshot). Kopier denne data i en mærket regneark.
- Mål Samlet endepladen (område inden for polygon) ved at vælge: Analyze> Mål. Dette giver det samlede endepladen område. Kopier og indsæt data for AChR områder og intensiteter iet regneark og sørg for at mærke kolonner korrekt, vil rækker blive brugt til enkelte endeplader for specifikke dias.
- Skift til det anti-synaptophysin fluorescens kanal og gentag trin 5,1-5,5, men for FITC kanal (figur 3B, D og F). Målet er at justere tærsklen, så den skaber et binært billede, som så tæt som muligt matcher grænserne for farvning som opfattes af øjet. Optag tærskelværdien.
- Mål det område, overlapning ved anvendelse af følgende fremgangsmåde: Åbn den oprindelige fil, der indeholder de to kanals billeder og opdele den i to separate billeder ved at vælge: Image> Stakke> Stack til billeder.
- Brug af colokalisering plugin (downloades og installeres fra ImageJ hjemmeside) Vælg: pluggin> colokalisering og input tærskelværdierne tidligere optaget for AChR og nerve-kanaler ind i den respektive kanal forespørgslen Box. Dette vil give et overlap billede i hvide pixels (Figur 3H og tilhørende ImageJ screenshots).
- Konverter den nyoprettede overlap billede i en gråtoner format og anvender en tærskel til den maksimale værdi. Den maksimale grænse vil kun vælge de hvide pixels, der svarer til det overlappende område af de to foregående kanaler. Optag i regnearket den resulterende område værdien »colokalisering ', der repræsenterer området af overlapning i pixels.
- Forbered et regneark med data prøve midler, beregne og plotte standardafvigelser og standardfejl som histogrammer eller scatterplots 20,22. Bemærk, at værdien af n betegner generelt antallet af mus pr prøve gruppe til statistiske formål.
- Plot endepladen AChR områder som scatterplots eller frekvens histogrammer at afgøre, om data er normalfordelt inden statistisk test (figur 6).
6. Relativ FarvningIntensiteter sammenlignet ved anvendelse Tværgående Optiske Sektioner
BEMÆRK: For denne protokol proces alle muskelgrupper prøver sammen og billede i en enkelt konfokal session. Ved planlægningen et eksperiment tillader op til 30 min imaging pr muskel prøve.
- Skær 15 um kryosektioner på tværs af længdeaksen af muskelfibrene og indsamle på objektglas som beskrevet i trin 3.1.
- Udfør fluorescensfarvning som beskrevet i trin 3,2-3,13.
- Kode de farvede objektglas, så billedbehandling og analyser gennemføres med operatøren blind for behandlingsgruppen, som beskrevet i trin 4.1.
- Ved hjælp af en 40X fluorescens mål (NA 0,75) kortvarigt overskue et afsnit fra hver slide for at bestemme en enkelt forstærkning og offset niveau indstilling for AChR der vil være egnet til alle endepladerne på tværs af alle prøve dias. Den lyseste endeplade skal da være lige under 256 grå på skalaen. Denne optimering bør ske særskilt for det andet fluorescence kanal (opsamlet successivt). Optag den faste forstærkning og offset indstillinger niveau og ikke ændrer dem i hele imaging session.
- Saml billeder af en fluorescens standard slide (f.eks, ikke-blegning fluorescerende perler) under anvendelse af de samme parametre i begyndelsen og slutningen af det konfokale session til at detektere eventuelle udsving i laser intensitet.
- Brug AChR kanal til at scanne dias gradvis at lokalisere endeplader.
- Fokus for at finde den eneste optiske sektion fly i hvert mikroskop felt, der indeholder de mest antallet af AChR-farvede endeplader.
- Skan denne enkelt optisk afsnit to gange og gemme den gennemsnitlige billede (figur 4G).
- Holde det samme brændplan skifte til den anden fluorescens kanal (protein) og indsamle billedet som i trin 6.8. Gem billedfilen, herunder filnavnet: dato for billeddannelse session, kodeeksempler, billede nummer og et symbol for at angive fluorescerende kanal.
- Flyt scenen til det næste felt der indeholder en eller flere endeplader og gentag trin 6,8-6,9. Gentag dette, indtil i alt 60 endeplader er afbildet.
- Ved afslutningen af overførslen imaging session alle filer til en anden computer og bakke dem op.
- Åbn hver original billedfil og mens du ser på AChR kanal, skal du vælge: Image> Stakke> Stack til billeder, for at opdele kanaler.
- Vælg: Billede> type> 8bit at konvertere til 8-bit gråtoner format på skærmen. Gør dette for begge fluorescens kanaler.
- Vælg: Billede> Stakke> Billeder til at stable. Åbn en ny stak fra to tidligere adskilte 8-bit billeder. Man kan så skifte bekvemt mellem de to fluorescens-kanaler i det indre vindue.
- Brug polygon til at tegne en line stramt omkring grænsen af AChR farvning (figur 4I).
- Vælg: Analyze> Mål for at måle den gennemsnitlige pixel for AChR intensitet i det lukkede område (bemærk at det er vigtigt at trække linjen stramt). Kopier denne værdi i en mærket regneark.
- Bevarelse af den samme polygon skitse (at definere det område, der skal måles), skifte til den anden fluorescerende kanal (f.eks figur 4B, D, F) og vælg: Analyze> foranstaltning. Dette vil give den gennemsnitlige farvningsintensitet for proteinet af interesse i den synaptiske defineret af AChR farvning.
- Vælg et område væk fra synligt endeplade farvning derefter vælge: Analyze> Mål for at måle den gennemsnitlige baggrund fluorescensintensitet. Gentag dette for de andre fluorescens kanal / s og kopier baggrundsværdier i regnearket af fluorescens værdier.
- SubTract de gennemsnitlige baggrundsværdier fra endepladen værdier for at opnå de korrigerede intensiteter for AChR og proteinet af interesse på hver endeplade.
- Opdel korrigerede endepladen intensitetsværdier for proteinet af interesse ved den korrigerede BGT fluorescensintensitet til opnåelse af fluorescensintensitetsforhold 14,21
7. Sammenligning af postsynaptiske membran AChR Density Brug FRET
BEMÆRK: Denne protokol vurderer hvilket omfang AChRs er tæt pakket (<10 nm mellemrum) i den postsynaptiske membran. Den præcise donor- og acceptor fluorophor kombination er kritisk for FRET assay. Navne og adresser på fluoroforerne er angivet i Materials tabellen. Deres spektrale egenskaber, i forhold til at ærgre sig, diskuteres i vores tidligere papirer 14,15.
- Forbered faste tværgående kryosektioner som beskrevet i afsnit 6.1. Alle prøve grupper skal behandles sammen og billeded i samme konfokal session.
- Bland grundigt 2,5 g / ml røde BGT (FRET donor) med 10 g / ml langt røde BGT (FRET acceptor) med blokerende opløsning i en lille plastikrør ved pipettering op og ned 12 gange. Dette 1: 4 molær blanding maksimerer effektiviteten af FRET 14.
- Placer hver slide i et fugtigt kammer, omhyggeligt dække hver sektion med en dråbe (12 pi) af den ovennævnte blanding og inkuberes i 1,5 timer ved stuetemperatur.
- Kontrol sektioner: dækker et lille antal sektioner med 2,5 g / ml rød-BGT (kun donor, mærket C1 kontroller), og også nogle sektioner med 10 g / ml langt rød-BGT (kun acceptor, mærkede C2 kontroller). Inkuber disse kontroller som ved trin 7.3.
- Vask glider 3 x 10 min i PBS og montere i glycerol-baseret, fade-modstand montering medium (se trin 3.12).
- Udfør prøvetagning af endeplader som i trin 6.7. Fluorescens fra donoren og acceptoren skal være fuldkommen co-lokaliseret ved endeplader grund af den tilfældige binding af fluorescerende BGT molekyler.
- Kontrol billeder: Brug af 40X objektiv og lav laser power optimere redBGT gain og offset indstillinger niveau for endeplader fra et dias kontrol C1. Optimer langt redBGT gain og offset niveauer for endeplader fra slide kontrol C2. Bekræft mangel af fluorescens gennemblødning.
- Uden at ændre laser magt, vinde eller indstillinger forskudt niveau flytte til de eksperimentelle dias og indsamle billeder (pre-photobleach) for begge fluorescens kanaler.
- Selektivt photobleach den langt-rød-BGT over en del af en enkelt endeplade ved at zoome ind scanningsområdet derefter scanne 10 gange med 633 nm laser ved 100% effekt. Den fluorescens i det scannede område bør blive dim.
- Nulstil lasereffekten og zoome og indsamle post-blegemiddel billeder på begge fluorescerende kanaler ved hjælp af konfokale indstillinger, der er etableret på 7,7.
- Beregn FRET effektivitet (E) fra procentvise stigning i donor (rød-BGT) fluorescens efter photobleach af acceptor (langt-rød-BGT) i henhold til følgende formel *:

* For alle situationer, hvor fluorescens af donoren stiger efter fotoblegning acceptoren.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Måling af Synaptic Area på NMJ
Ethvert skøn over område afhængig tegningen af en grænse for at definere omfanget af synaptiske specialer. Hos raske unge voksne muskler NMJ billeder skal vise veldefinerede grænser for både AChR og synaptophysin farvning (figur 2A og B). Fluorescens for både AChR og synaptophysin intensitet stiger kraftigt ved grænsen mellem den peri-synaptiske og synaptiske del af motoren endeplade (figur 5A 'og B'). For sådanne billeder en minimumstærskel (lige over ekstrasynaptiske baggrundsfluorescens) vil let isolere AChR-rige eller synaptophysin-rigt område af endepladen (vandret punkterede linier i (figur 5A 'og B')). I ældre mus og i nogle sygdomstilstande endepladen farvning for AChR kan være mindre intens, kan AChR cluster kanter er sløret, ogre kan være højere niveauer af ekstrasynaptiske autofluorescens (figur 5D; 17,22). Fluorescens farvning med utydelige grænser kan introducere fejl i skøn over synaptisk område. I alle situationer er målet at vælge en tærskel, der giver et binært billede svarer i form og størrelse til AChR- eller synaptophysin-rige områder, som de vises for øjet i den oprindelige, halvtonebillede. Udførelse af analysen blind for behandlingsgruppen bør reducere risikoen for subjektive forudindtagethed i tærskelværdier (trin 4.1). Undertiden svage eller sløret Endepladeforandringer billeder kan resultere fra sub-optimal behandling. Figur 2C og D viser et eksempel på en lav kvalitet endeplade billede fra en sund 2 måned gammel mus. Slørede kanter og svag synaptophysin farvning kan være opstået i dette tilfælde fra delvis optøning og genindfrysning af musklen før cryosectioning. Dele af unge sunde (positiv kontrol) muskel bør sektioneret ennd behandles parallelt med eksperimentelle prøver at sikre, at enhver svækkelse tydeligt i de NMJ billeder er ikke på grund af problemer med immunfarvning. Hold af billeder kompromitteret af suboptimal behandling bør udelukkes fra analyserne.
For en face z-stack billeder, 15-20 endeplader er en rimelig stikprøvestørrelse til estimering synaptiske områder. En bred mangfoldighed i former og størrelser af NMJs findes under en bestemt muskel. Scatterplots afslører en betydelig rækkevidde i AChR-rige område blandt endeplader af tibialis anterior muskel af individuelle mus (figur 6A). Ikke desto mindre er den gennemsnitlige AChR området (baseret på 15-20 en face endepladen billeder) var ens blandt syv prøve mus (~ 200 m 2, figur 6A). Arealet af endepladen synaptophysin farvning også varierede betydeligt blandt endeplader fra en given muskel. Endnu en gang ved hjælp af en stikprøve på 15-20 endepladerne det gennemsnitlige areal på endepladen synaptophysin var SimiLAR blandt 7 mus undersøgt (~ 170m 2, figur 6B). Frekvens histogrammer for poolede data viste nogenlunde normalfordelinger for området med endepladen AChR og synaptophysin (figur 6A 'og B'). Men en normal fordeling af synaptiske områder kan ikke antages i sygdomstilstande såsom myasthenia gravis 16,20. Dette kan påvirke valget af den statistiske test.
Tabel I viser områderne forudgående og postsynaptiske specialiseringer for NMJs til sund 2 måned gammel (unge voksne) kvindelig C57BL / 6J mus fra tidligere undersøgelser. Områderne både præ- og postsynaptiske specialiseringer faldt med stillesiddende aldring 22. AChR området blev også reduceret markant i mus injiceret med IgG fra anti-Musk myasthenia gravis patienter 17,21. Muskelsvaghed mus behandlet med kolinesterasehæmmer narkotika, pyridostigmin, viste en yderligere væsentlig reduktion i endeplade AChR area 20 </ Sup>.
Relative intensitet af Gavlsæt fluorescensmærkning
Den relative intensitet af immunfluorescens mærkning kan afsløre ændringer i massefylden af en synaptisk protein af interesse med alderen, genotype og / eller sygdomstilstand. AChR fluorescens (rød-BGT eller langt-rød-BGT) først til at definere placeringen af NMJ. Lysstyrken af fluorescens inden for 8-bit AChR-rigt område er derefter anvendt til at vurdere ændringer i koncentrationen af proteinet af interesse, i forhold til kontroldyr. I tværsnit endepladen AChRs vises typisk som en halvmåne form, men denne form er ofte uregelmæssig (figur 4A, C, E, H). Lav intensitet baggrundsfluorescens afslører sædvanligvis om et plaster på AChR farvning betegner en enkelt endeplade, eller to separate endeplader placeret på tilstødende muskelfibre. Mange synaptiske proteiner (såsom RapSyn, moskus og SRC) ercolocalized med AChR på endepladen (figur 4A - D). Immunofluorescens-farvning med phospho-specifikke antistoffer kan også anvendes til at sammenligne effekten af eksperimentelle interventioner på phosphorylering status af bestemte postsynaptiske membranproteiner 21.
Pålidelighed og reproducerbarhed af fluorescensintensitet måling afhænger stærkt afhængig af integriteten af den frosne muskler og kvaliteten af immunfarvning. Muskler skal dissekeres og straks lynfrosset eller paraformaldehyd-fast (inden for minutter af dyrets død) for at undgå degenerative ændringer i NMJ. Immunfarvning afhænger stærkt af kvaliteten af de reagenser og optimere farvningsprotokol for specifikke antistoffer. For enhver ny batch af primært antistof pilot Immunofarvning eksperimenter er nødvendige. Frisk skåret snit af raske unge muskler inkuberet med serielle 2-folds fortyndinger af det primære antistof. Akendt pålidelig sekundært antistof anvendes, og resultaterne sammenlignes. Hvis proteinet af interesse er kendt for at være begrænset til NMJ så den bedste antistofkoncentrationen er den, der giver det højeste forhold af NMJ fluorescensintensitet i forhold til den, der findes i ekstrasynaptiske dele af musklen (baggrundsfluorescens). Ekstrasynaptiske (formentlig ikke-specifik) fluorescensintensitet bør normalt ikke overstige 15% af endepladen fluorescensintensitet. Tilsvarende nej primære antistof kontrol «sektioner (inkuberet kun med sekundært antistof) skal vises mørkt, der bekræfter, at det sekundære antistof ikke bandt ikke-specifikt. Kvaliteten af forskellige partier af (polyklonale) sekundære antistoffer kan variere markant så alternative sekundære antistof partier bør sammenlignes før etablere en standard-protokol. Den ideelle test for specificitet immunfluorescens indebærer en direkte sammenligning af sektioner fra vildtype mus og negativ-kontrol sektioner fra mus that mangler proteinet af interesse (gen knockout). De følgende referencer beskriver kvantificering af endepladen fluorescens farvning for moskus, RapSyn, dystroglycan, src og AChR 13,14,18,21.
Små stikprøvestørrelser introducere fejl i skøn over relativ fluorescensintensitet. Individuelle endeplader varierede betydeligt i fluorescensintensitet. Formentlig denne variation mellem endeplader inden for en given muskel afspejler mangfoldigheden i NMJ struktur og tilfældige forskelle i den optiske del i stikprøven. Men forøgelse af antallet af endeplader stikprøven resulterer i en mere stabil anslåede gennemsnitlige fluorescensintensitet (figur 7). For at estimere den gennemsnitlige endepladen fluorescensintensitet 40-60 endeplader fra hver muskel prøve skal beregnes.
AChR-AChR FRET
Hver AChR er en pentamer med to bindingssteder for BGT (placeret én på hver alfa-underenhed). Binding afrød-BGT og langt-rød-BGT til disse to steder ville give en donor-acceptor adskillelse af omkring 9nm 28-30. Således lav effektivitet FRET kan påvises, selv før AChRs samle i klynger 14. Men effektiviteten af FRET i murine endeplader groft fordoblet postnatal overensstemmelse med den mere effektiv inkorporering af AChRs i en tætpakket postsynaptiske membran gitter 14. Brug røde BGT og langt røde BGT som FRET donor og acceptor (henholdsvis), endepladerne på 1-2 måneder gamle mus producerede gennemsnitlige FRET effektivitet spænder fra 20 til 37% (tabel 2). FRET virkningsgrad på 20% eller mere menes at repræsentere stram pakning af AChRs 14. Gavlsæt FRET effektivitet blev reduceret en smule efter denervering 14 og markant reduceret efter mus blev injiceret med IgG fra anti-Musk-positive myasthenia gravis patienter 18. Disse er tilstande, hvor de enkelte AChRs er mindre tætpakkede i postsynaptic membran stillads ved den postsynaptiske moskus / RapSyn systemet 31.
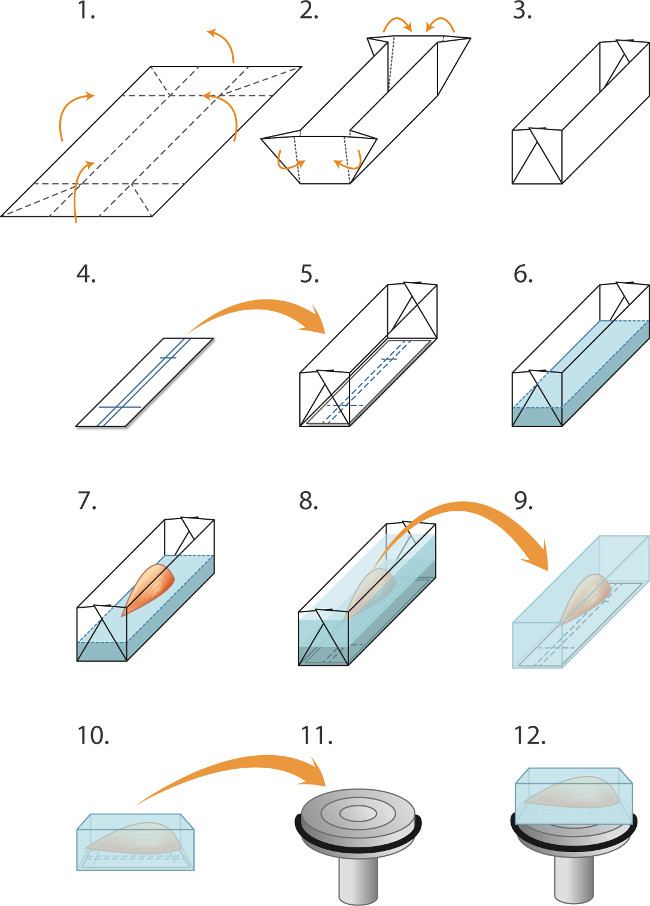
Figur 1. Indlejring og indefrysning muskler til cryosectioning (1-5) Forberedelse forme (»både«) inden frysning en batch af muskler:. (1) Aluminium folie skæres i rektangulær form (2,0 x 3,0 cm) og (2). Foldes som sigtet for at skabe en form / båd (3). (4) rektangler af nitrocellulose immunoblotting papir skæres til at passe i formen og en kuglepen anvendes til at udelukke linjer til orientering musklen. (5) nitrocellulose rektangel er anbragt i formen. (6-8) Embedding og frysning muskler: (6) Kryostat indlejring matrix væske forsigtigt hældes i formen (oven på nitrocellulose) til en dybde på 2 mm. (7) fin pincet bruges til at sænke muskel ved sin sene i indlejring matrix, på linje med nitrocellumister domme, (8) Yderligere indlejring matrix forsigtigt hældes i formen for at dække musklen, idet man undgå at skabe bobler. Den integrerede muskel derefter lynfrosset forseglet i rør og opbevaret ved -80 ° C som beskrevet i teksten. (9-12) Forberedelse cryosectioning: (9) fin pincet bruges til at skrælle væk aluminiumsform og den frosne blok anbringes derefter i -20 ° C kryostatkammeret (10) Markeringerne i nitrocellulose anvendes til at tilpasse musklen parallelt med forsiden af spændepatronen (11) til langsgående skæring (12). En dråbe væske cryo-indlejringsmedium bruges til at fastgøre blokken til den afkølede chuck. Blunt pincet bruges til at manipulere den frosne muskel blok. For at opnå tværgående sektioner, er blokken i stedet monteret således, at musklen er vinkelret på overfladen af patron (ikke vist).

Figur 2. En ansigtsbilleder af NMJs fra tibialis anterior muskel på 2 måneder gamle kvindelige C57BL6J mus. Den friske muskel var snap frosne og sektionerne fastgjort på dias som beskrevet i trin 3.2. Maksimal intensitet projektion billeder af z-stakke blev opnået som beskrevet i den første protokol. (A) rød-BGT farvning afslører en enkelt motor endeplade bestående af to sæt AChR-rige primære postjunctional tagrender (da ansigt visning). (B) synaptophysin farvning med FITC-konjugeret sekundært antistof viser den præsynaptiske nerve terminal, besætter de primære synaptiske tagrender. En del af den præ-terminale axon er også synlig i den øverste del af panelet. (C & D) Et eksempel på en dårlig kvalitet NMJ billede fra en ung sund mus. Farvning er svag og grænserne for de præ- og postsynaptiske specialer er slørede. Dette blev tilskrevet mangler i than behandlingen af væv og / eller utilstrækkelig tid tilladt for det primære antistof inkubation. Scale bar i panel D er 10 um. Klik her for at se en større udgave af dette tal.

Figur 3. Trin i behandlingen af en face NMJ billeder (som beskrevet i protokol 1). (A & B) Originale kontinuerlige tone MIP billeder viser røde BGT fluorescens afslørende AChR og grøn immunofluorescens for synaptophysin hhv. (C) AChR farvning efter konvertering til en 8-bit gråskalabilledet og brug af polygon værktøj til at afgrænse endepladen (tynd gul linje). (D) Gavlsæt grænselinje, der overføres til den grønne synaptophysin billede. (E) Applgement på mindst intensitet tærskel kommando til at oprette et binært billede, der isolerer suprathreshold røde BGT (AChR) fluorescens. Sekvensen af ImageJ brugergrænseflade screenshots vises til venstre for billederne. (F) Binary synaptophysin billede efter anvendelse af en separat tærskel. (G) Identifikation af diskrete suprathreshold AChR-rige domæner i endepladen ved anvendelse af analyse-partikler kommando til den binære røde BGT billede. Den tilsvarende ImageJ (til venstre for panelet (G) viser input nødvendige data. Skal indtastes Mindstemålet for suprathreshold pixel områder, der kræves. (H) Identifikation af områder af overlapning af det binære synaptophysin og AChR billeder. Overlapning er repræsenteret af hvide pixels . ImageJ brugergrænseflade screenshots (under panel H) viser trin for at nå frem til det binære overlap billede. De valgte minimum intensitet tærskelværdier for hvert fluorescens channel skal indtastes under "colokalisering 'vindue. Scale bar udgør 10 pm. Klik her for at se en større udgave af dette tal.

Figur 4. Eksempler på tværgående optiske sektioner anvendes til at sammenligne relative fluorescerende intensiteter (protokol 2). Musklen blev hurtigt nedfrosset og sektionerne fast på slæden som beskrevet i trin 3.2. (A & B) En enkelt endeplade dobbelt-mærket med langt -Red-BGT (AChR; her vist i blå pseudofarve) og FITC-anti-RapSyn illustrerer co-lokalisering af disse to interagerende proteiner i den postsynaptiske membran (C & D) To endeplader på tilstødende muskelfibre display co-lokaliserede AC. hR og moskus. (E & F) En endeplade dobbelt-mærket for AChR og -dystroglycan (-DG). Den -DG strækker lige rundt om muskelfibrene omkreds men er beriget på endepladen (skala bar i F, for paneler AF: 25 pm) (G - I) Isolering en endeplade for intensitet måling (G) En typisk mikroskop feltet.. indeholdende tre langt røde BGT-farvede endeplader (skala bar i panel G: 40 um (H) Et forstørret billede af boxed endeplade (I) Det samme endeplade konverteret til en 8-bit gråtoner billede og afgrænset ved hjælp af polygonen.. . redskab ImageJ (tynd gul linje) er Gennemsnitlig fluorescensintensitet målt inden for denne grænse linje. (skala bar for H & I: 10 pm) Klik her for at se en større udgave af dette tal.

Figur 5. Påvirkning af billedkvaliteten ved vurderingen synaptisk område. (A & B) Høj kvalitet en face billeder af en sund NMJ fra en 2 måned gammel mus ses på røde BGT og anti-synaptophysin fluorescens-kanaler. (A 'og B ') Fluorescens intensitetsprofiler svarende til linje på tværs endepladen for A og B henholdsvis. Den vandrette røde stiplede linje angiver minimumsgrænse bruges til at skabe det binære billede. (C & D) Gavlsæt fra en ældre mus. Gavlsæt synaptophysin farvning generelt er mindre intens. (C '& D') intensitetsprofiler viser en høj grad af ekstrasynaptiske (baseline) fluorescens udsving i synaptophysin (FITC) kanal (tilbagejorden), der påvirker bestemmelsen af en passende tærskel. Meget af dette er bredspektret væv autofluorescens. Scale søjler repræsenterer 10 um. Klik her for at se en større udgave af dette tal.

Figur 6. Variabilitet af synaptiske områder blandt NMJs inden for en muskel og mellem mus. (A) Scatterplots viser den samlede AChR-rige område endeplader fra tibialis anterior muskel i syv naive 2 måned gammel kvindelige C57BL / 6J mus opnået ved forfatteren NT. Hvert symbol repræsenterer en endeplade. Hver søjle repræsenterer middelværdien ± SD for endeplader udtaget fra en mus. (B) Scatterplots viser synaptophysin-rigt område for de samme endeplader. (A) (B ') Frekvens fordeling for synaptophysin-rige område endeplader (fælles data). Klik her for at se en større udgave af dette tal.

Figur 7. Virkning af prøvestørrelse på estimater af endepladen fluorescensintensitet. Tværgående optiske sektioner blev brugt til at måle fluorescensintensiteten (i arbitrære enheder) 40 60 endeplader fra tibialis anterior muskel af en sund 2 måned gammel mus. Kumulative gennemsnit afsættes som funktion af antallet af endeplader indgår i gennemsnittet. (A) Gavlsæt rød-BGT fluorescensintensitet opnået ved forfatter AV. (B) Anti-synaptophysi(C) Gavlsæt langt rød-BGT fluorescensintensiteten opnået fra en anden muskel prøve af forfatteren NG. (D) Anti-RapSyn immunofluorescens intensitet fra samme endeplader som i panel C n immunofluorescens intensitet for de samme endeplader som i panel A..
| Undersøgelse | Muskel | Antallet af mus | AChR område | Synaptophysin område | Overlappende område |
| middelværdi ± SD (pm2) | middelværdi ± SD (pm2) | middelværdi ± SD (pm2) | |||
| Morsch et al. (2012) 1 | gastrocnemius | 3 | 181 ± 7 | 163 ± 24 | |
| membran | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch et al. (2013) 1 | tibialis anterior | 3 | 166 ± 26 | 117 ± 21 | ND |
| Cheng et al. (2013) 2 | tibialis anterior | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| TSE (upubliceret) 2 | tibialis anterior | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| 1 afbildes på et Zeiss LSM 510 Meta mikroskop, men med fast forstærkning og offset niveauer. Tærskel og synaptiske områder målinger brugte Metamorph Software af MM. | |||||
| 2 afbildet på et Leica DM IRE2 mikroskop og analyseret efter gældende protokol den angivne første forfatter, blind for behandlingsgruppe. | |||||
| nd ikke bestemt. | |||||
Tabel 1. Synaptiske områder for NMJs af 2 måneder gammel kvindelige C57BL / 6J mus (raske kontroller)
| Undersøgelse | Muskel | FRET effektivitet (%) * | Range (%) |
| (Gennemsnit ± SEM) | |||
| Brockhausen et al. (2008) | tibialis anterior | 24 ± 1 | na |
| Cole et al. 2010 | tibialis anterior | 26 ± 1 | 22-30 |
| Morsch (ikke offentliggjort) | membran | 37 ± 1 | 24-47 |
| Ghazanfari (ikke offentliggjort) | tibialis anterior | 30 ± 1 | 20 - 45 |
| * FRET mellem rød-BGT og langt-rød-BGT (Förster radius for FRET par = 51A (Life Technologies). | |||
| NA data ikke tilgængelige | |||
Tabel 2. effektivitetsgevinster for AChR FRET fra unge voksne mus C57BL6J mus
Subscription Required. Please recommend JoVE to your librarian.
Discussion
De her beskrevne protokoller har gjort det muligt for os at pålideligt måle og kvantificere ændringer i egenskaberne af NMJ tværs af en række forhold, herunder normal aldring og sygdomstilstande. Beskrevet for en metoder står NMJ billeder vil tillade forskere at sammenligne området præ- og postsynaptiske specialiseringer og området af synaptisk overlap / justering. For at sammenligne den relative intensitet af præ- og postsynaptiske proteiner anden protokol, som bruger tværgående optiske sektioner, foretrækkes. Den tredje protokol specifikt tester for ændringer i nærheden af pakning af AChRs i den postsynaptiske membran.
Specificitet kontrol er afgørende i immunfluorescensmikroskopi. Ved anvendelse af et primært antistof til indirekte immunfluorescens er det nødvendigt først at sikre, at det binder specifikt til dens målprotein i musklen sektioner. Forskellige former for vævsbehandling og fiksering kan differentielt ændre specificitetantistoffer. Det er vigtigt at bekræfte, at immunofluorescens-farvning (for sige RapSyn) virkelig er koncentreret med AChR på motoren endeplade. Negative kontrol- sektioner skal også inspiceres for at sikre, at antistofbinding er specifik. For eksempel ville den bedste negativ kontrol for RapSyn immunfluorescens være sektioner fra RapSyn - / - mus. Disse bør ikke vise endepladen farvning med anti-RapSyn. Ikke-specifik fluorescens kan også opstå fra endogene fluorescerende kemikalier i vævet (autofluorescens) eller af ikke-specifik binding af det fluorescerende sekundært antistofkonjugat. En sådan fluorescens ofte forværret af aldehyd fiksering. Desuden kan TRITC-BGT farvning af endepladerne undertiden påvises i FITC fluorescens kanal og dette fluorescerende gennemblødning kan forveksles med specifikke FITC immunofluorescens. For at beskytte mod de sidste tre former for ikke-specifik fluorescens, skal hvert parti af dias, der farves omfatter nogle 'no-primært antistof kontrol sektioner (trin 3.7 og 4.6). Billeder af endeplader fra disse kontrolsektionerne bør sammenlignes med dem fra de eksperimentelle slides at sikre, at den indirekte immunfluorescensfarvning af NMJs virkelig afspejler binding af det primære antistof.
Tværgående konfokale sektioner er særligt anvendelige til at vurdere forskelle i den relative intensitet af immunfarvning ved synapsen. I tværgående konfokale sektioner er det lettere at bedømme præcis co-lokalisering af synaptiske proteiner. Den halvmåneformede endeplade profil udgør blot en prøve cut-gennem NMJ pågældende. Imidlertid er baggrunden (ekstrasynaptiske) fluorescens generelt lavere i forhold til en face z-projektion billeder. Således kan det være lettere at diskriminere 'rigtige' (specifik) immunfarvning og etablere fast konfokal gain og offset værdier ved hjælp af tværgående optiske sektioner 13-15,18. For eksempel, i en musemodel af myasthenia gravis (Where endeplade AChR farvning markant reduceret) endeplader var klart afgrænset i tværgående optiske sektioner 18,21. Forskelle i den gennemsnitlige fluorescensintensitet ved NMJ sandsynligvis afspejler ændret tæthed af målproteinet indenfor den synaptiske specialisering. En advarsel er, at der i visse situationer kan en strukturel ændring i målproteinet eller okklusion af antistof binding ved tilstødende proteiner forklare ændret farvning intensitet.
Udformningen af eksperimenter kræver en vis overvejelse. I mange tilfælde eksperimentet tage sigte på at teste virkningen af et transgen, gen knockdown eller sygdomstilstand af størrelsen af NMJ. Den eksperimentelle prøve gruppe kan så sammenlignes med raske unge (vildtype) mus af samme køn og genetiske baggrund. Baseline værdier for området af endepladen synaptophysin, AChR og synaptisk overlapning flere muskler er angivet i tabel 1. Prøvens størrelse afhænger af graden af dyr-til-dyr variation inden behandlingsgrupper og effekten størrelse (forskel i midler til eksperimentelle versus kontrol grupper pr standardafvigelse). Når analysen er begrænset til billeder af god kvalitet en rimelig grad af ensartethed blev fundet i prøven midler til endepladen områder blandt raske 2 måned gammel kvindelige C57BL6J mus (figur 6A og B). Det var således muligt at påvise væsentlige reduktioner i synaptisk område 30-40% i mus injiceret med IgG fra anti-Musk-positive myasthenia gravis patienter, sammenlignet med kontroller med en stikprøvestørrelse på tre mus 17,20,32. Ældre mus viste større dyr-til-dyr variation i endepladen parametre end unge mus 22. Derfor forsøg med alderen mus kan kræve større stikprøvestørrelser.
Hvis den primære bekymring er at måle størrelsen på en face endeplade så gevinsten og offsetindstillinger niveau bør være optimeret til hver enkelt NMJ. Indivelle NMJs kan variere betydeligt i lysstyrke AChR og synaptophysin farvning, især når sygdomstilstande undersøges. Desuden intensiteten af ekstra-synaptisk (ikke-specifik) fluorescens er ofte højere og mere variabel i musklerne aldrende dyr, sammenlignet med de raske unge dyr (figur 5C og D). De 1-256 grå-skala bør udnyttes fuldt ud for at maksimere den tonale oplysninger, der vil blive medtaget på den endelige billeder. Dette indebærer at justere gain og offset niveauer for hver NMJ for hvilke en z-stak der skal opsamles. Figur 5D viser et eksempel på et NMJ billede, hvor tonale oplysninger kan være afgørende for afgrænsningen af området før og efter synaptiske specialiseringer.
Målinger af synaptiske områder kan anvendes til forskellige muskelgrupper præparater og eksperimenter. De fleste af vores målinger af synaptiske områder har ansat langsgående kryosektioner fra snap frosne muskler. Frysning musklen før fiksering opretholder antigeniciteten af en bred vifte af proteiner. Når forenelig med antigenet, paraformaldehydfiksering og saccharose infiltration før cryosectioning (trin 2.1) kan give bedre bevarelse af NMJ struktur. Optimal strukturel konservering kan opnås ved kardiel perfusion med paraformaldehyd. Artefakter af frysning og sektionering helt kan undgås ved mærkning endeplader på overfladen af de intakte muskel- og billedbehandling NMJs om hæfterne drillet fra den faste muskler 21. Uanset præparatet, procedurerne for prøvetagning, billedbehandling og areal kvantificering uændret (protokol trin 4-5). Konsekvent anvendelse af blinde prøveudtagning, billedbehandling og analyse protokoller (ved hjælp af forskellige operatører, forskellige prøver af mus og forskellige tidspunkter), kan resultere i forholdsvis reproducerbare gennemsnitsværdier (sammenligne Cheng et al. Og TSE resultaterne i tabel 1).
"> Endeplader er blevet beskrevet som at blive" opsplittet "i en række sygdomstilstande. For eksempel i aldrende mus muskler, sporadisk degeneration af en muskel fiber (efterfulgt af regenerering) resulterede i ombygning af kringle-lignende endeplade AChR plaque til danne flere mindre AChR klynger 6. I mus injiceret med IgG fra anti-Musk myasthenia gravis patienter, fragmentering af endepladen var noget anderledes. endepladen AChR kringle stort set spredt, efterlader en konstellation af små (<4 m 2) AChR 'mikroaggregater '20,21. Disse to eksempler fremhæve behovet for at sammenligne størrelsesfordelinger for AChR klynger på endeplader af kontrol versus forsøgsdyr 21.Er blevet rapporteret Andre metoder til vurdering af synaptisk område eller farvningsintensitet på NMJ. Motor endeplader kan undertiden foldes således at de to-dimensionelle z-projektion billeder her anvendte måske undervurdere synaptic områder. Tre-dimensionelle konfokale rekonstruktioner kan give mere præcise foranstaltninger, hvis absolut synaptisk område skal defineres 33. En væsentlig fordel ved z-projektion protokollen beskrevet her, er imidlertid dens relative enkelhed, som har tilladt et stort antal endeplader skal måles fra flere behandlingsgrupper og pålidelig identifikation af mulige ændringer. Protokollen til sammenligning Endepladeforandringer farvningsintensiteter kan tilpasses til at studere ændringer i niveauerne af mange forskellige synaptiske proteiner. Metoden er imidlertid begrænset af kravet om, at alle prøver behandles til immunfluorescens derefter afbildes i samme konfokal session. En nylig undersøgelse fra Yampolsky et al. 5 beskrives en metode til måling af endeplade AChR tæthed, der kan hjælpe med at overvinde denne begrænsning. I denne undersøgelse blev de samme områder af endeplader afbildet på flere forskellige laser strømindstillinger. Hældningen af forholdet mellem lasereffekt og rhodamiNE-BGT fluorescensintensitet blev anvendt til at vurdere de relative ændringer i AChR densitet ved endeplader i forskellige mus 5. Denne metode kan være nyttig til at sammenligne AChR intensitet i prøver afbildes på forskellige tidspunkter i løbet af en langvarig undersøgelse.
AChR-AChR FRET indeholder specifikke og supplerende oplysninger om tilrettelæggelsen af endepladen AChRs. Electron mikroskopisk autoradiografi under anvendelse af 125. I-α-BGT har vist AChRs skal pakkes tæt med en plan tæthed på 10 4 m -2 straks under hver præsynaptiske sted for transmitterfrigivelse, mens tilstødende membran infoldings indeholder meget lavere tætheder af AChR 34. AChR-AChR FRET gør det relativt let at opdage (sub-mikroskopiske) ændringer i AChR emballage. En reduktion i FRET effektivitet afspejler en sub-mikroskopisk omfordeling af AChRs i den postsynaptiske membran, der måske ikke fundet af en ændring i gennemsnitlig BGT fluorescensintensitet. Multiple faktører kan forårsage en ændring i effektiviteten af FRET. Heriblandt donor-acceptor afstand og relative orienteringer såvel som den molekylære miljø 35,36. En reduktion i endepladen FRET effektivitet eventuelt opstå en ændring i geometrien af AChR gitter. Men mest sandsynligt ville det være på grund af et fald i procentdelen af AChRs der er pakket ind i nanoskala postsynaptiske molekylær gitter 14.
Tab af forbindelsen mellem motoriske neuroner og muskelfibre synes at være den umiddelbare årsag til muskelsvaghed i motorisk neuron sygdom og i stillesiddende aldring 9,22,23. Delte metoder og parametre til måling NMJs skulle gøre det lettere for forskellige forskningsgrupper at sammenligne offentliggjorte undersøgelsesresultater. Deling af detaljerede protokoller (og fremtidige forbedringer på dem), kan bidrage til at fremskynde udviklingen i forståelsen af mekanismerne i NMJ vedligeholdelse og hvordan det kan blive påvirket i sygdomstilstande.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
