

Abstract
Die neuromuskulären Synapse (NMJ) ist die große, cholinergen Synapse Relais durch die Säugetiermotoneuronen steuern freiwilligen Muskelkontraktion. Bauliche Veränderungen an der NMJ kann bei der Neurotransmission Versagen führen, was zu Schwäche, Atrophie und sogar zum Tod der Muskelfaser. Viele Studien haben untersucht, wie genetische Veränderungen oder Krankheit kann die Struktur der Maus NMJ ändern. Leider kann es schwierig sein, direkt Erkenntnisse aus diesen Studien zu vergleichen, weil sie oft verwendet verschiedene Parameter und analytische Methoden. Drei Protokolle werden hier beschrieben. Der erste verwendet Maximum Intensity Projection konfokalen Bildern, die Fläche der Acetylcholin-Rezeptor (AChR) -reichen messen postsynaptischen Membran-Domänen an der Endplatte und der Fläche der synaptischen Vesikel-Färbung in der darüberliegenden präsynaptischen Nervenende. Das zweite Protokoll vergleicht die relativen Intensitäten der Immunfärbung für synaptische Proteine in der postsynaptischen Membran. Die dritte protocol verwendet Fluorescence Resonance Energy Transfer (FRET), um Veränderungen in der Packung der postsynaptischen AChR an der Endplatte zu detektieren. Die Protokolle sind entwickelt worden, und über eine Reihe von Studien verfeinert. Faktoren, die die Qualität und Konsistenz der Ergebnisse beeinflussen werden diskutiert und normative Daten für NMJs bei gesunden jungen erwachsenen Mäusen zur Verfügung gestellt.
Introduction
Die neuromuskuläre Verbindung (NMJ) ist die kritische Relais Synapse, die eine Kommunikation zwischen dem Nervensystem und im Skelettmuskel vermittelt. Es wird für alle willkürlichen Bewegungen erforderlich. Die Fluoreszenzmikroskopie ist seit langem verwendet, um die Auswirkungen von Transgenen auf der Maus NMJ 1-3 studieren oder um die Auswirkungen von Alter, Ernährung, Bewegung und Krankheit auf Nagetier NMJs 4-11 vergleichen. Solche Studien haben uns viel über die Physiologie und Pathophysiologie der NMJ gelehrt, aber die verschiedenen Parameter angegeben (zB AChR Bereich Endplatte Fläche, Umfang Länge, Fragmentierung Indizes) machen es oft schwierig, die Ergebnisse dieser Studien zu vergleichen. Es gibt eine steigende Erwartung für die präklinische Forscher in der Lage, um die Reproduzierbarkeit zu demonstrieren, insbesondere in Studien mit Tiermodellen der Krankheit 12 sein. Die hier beschriebenen Protokolle wurden durch eine Reihe von Studien, die entwicklungsbedingten, physiologischen und pathophysiologischen ch sucht verfeinertanges auf die NMJ. Erfordern solche Untersuchungen Messung der Fläche von synaptischen Spezialisierungen an der Maus motorischen Endplatte und die relative Packungsdichte der synaptischen Proteinen innerhalb postsynaptischen Spezialisierungen 13-15.
Die Nützlichkeit dieser Verfahren wird durch jüngste Studien in einem Mausmodell von anti-MuSK Myasthenia gravis dargestellt. Tägliche Injektionen von IgG von Anti-Moschus positive Myasthenia gravis Patienten in erwachsenen Mäusen veranlasste sie, innerhalb von 2 Wochen 16 schwach werden. Die konfokale maximale Projektionsbilder von Muskelpartien, die doppelt markierten wurden Synaptophysin (in Nerven Terminals) und postsynaptischen AChRs ergab einen allmählichen Rückgang im Bereich der AChR-Färbung als primäre Veränderung. Wichtig ist der Rückgang war ausreichend, um vergleichbare Rückgänge in der Amplitude des synaptischen Potentiale, Versagen der synaptischen Übertragung und Muskelschwäche 17,18 erklären. Qualitativ ähnliche Ergebnisse wurden von anderen Arbeitsgruppen berichtet10,19. Die gleichen NMJ Meßmethoden seit verwendet worden, um die Auswirkungen der drei Medikamente zur Behandlung von anti-MuSK Myasthenia gravis Beurteilung in diesem Mausmodell 20,21.
Sitzende Alterung können zum Verlust der neuromuskulären Verbindungen führen. Die hier beschriebenen Protokolle haben eine altersbedingte Rückgang im Bereich der Nervenendigung Synaptophysin bei motorischen Endplatten zeigte Mäuse Fortschritt ins hohe Alter. Die gleichen Methoden ergab, dass freiwillige Übung konnte weitgehend verhindern, dass die Reduzierung der präsynaptischen Nervenende Bereich 22, im Einklang mit früheren Arbeiten anderer Gruppen 4. Der Verlust der neuromuskulären Verbindungen kommt auch in der SOD1G93A Mausmodell für Amyotrophe Lateralsklerose 9,23.
Die genannten Studien zeigen, dass eine Vielzahl von gesundheitlichen Bedingungen kann zu Kürzungen im Bereich der entweder vor oder nach der synaptischen Spezialisierungen an der NMJ führen. Dies kann zu einer Beeinträchtigung der synaptischen Spaß führenction oder vollständigen Verlust der neuromuskulären Verbindung ankündigen. Drei Protokolle beschrieben, die Quantifizierung der Fläche und der Dichte der synaptischen Spezialisierungen ermöglichen. Der Zweck des ersten Protokolls besteht darin, eine praktische und reproduzierbare Bestimmung der Flächen der prä- und postsynaptischen Spezialisierungen und deren Ausrichtung in Säuger NMJs bereitzustellen, unter Verwendung von Fluoreszenzmikroskopie. Zweidimensionale maximale Projektions konfokalen Bildern und Bildanalyse mit NIH ImageJ wird verwendet, um Änderungen im Bereich des Synaptophysin-Färbung (synaptische Vesikel) postsynaptischen AChR und synaptische Überlappungsbereich zu detektieren. Konfokales Abbildungsparameter (Verstärkung und Offset-Pegel) werden für jede NMJ optimiert, um die visuellen Informationen verwendet werden, um den Bereich der synaptischen Spezialisierung erkennen maximieren. Neuromuscular Ausfall kann auch von Änderungen der Dichte des postsynaptischen AChR und / oder andere synaptische Proteine zur Folge haben. Das zweite Protokoll kann angewandt, um Änderungen in der relativen Dichte der postsynaptischen Proteine, nachzuweisenwie Moschus, Rapsyn, dystroglycan, phosphoryliert Src-Kinase phosphoryliert und AChR 18,21.
In Myasthenia gravis, ist eine reduzierte Dichte von AChR in der postsynaptischen Membran die unmittelbare Ursache der synaptischen Versagen und Muskelschwäche. Das dritte Protokoll beschreibt ein Fluoreszenz-Resonanz-Energie-Transfer (FRET), um die Veränderungen in der Nähe der benachbarten AChR innerhalb postsynaptischen Membranen 14,15 beurteilen. Diese Methode erkennt Energietransfer zwischen benachbarten AChRs mit fluoreszenz α -Bungarotoxin (BGT) gekennzeichnet. FRET tritt auf, wenn die fluoreszierenden Donor und Akzeptor-Sonden sind weniger als 10 nm auseinander liegen. Dies kann (submikroskopische) Veränderungen in der Dichtheit der AChR Verpackung, die direkt auf die Amplitude der synaptischen Potentiale beziehen können offenbaren.
Diese drei Protokolle, in der letzten Dekade raffiniert, bieten ergänzende Maßnahmen der NMJ Integrität in einer konsistenten und reproduzierbar. Verwendung standardisierter Protokolle einnd Parameter sollten Vergleich der Wirkungen von Genen und Umweltmaßnahmen auf die Säugetier NMJ erleichtern.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
HINWEIS: Design, Durchführung und Berichterstattung von Tierversuchen zu berücksichtigen, aktuellen Leitlinien 24 zu nehmen. Diese Arbeit ist im Voraus durch die lokale Tierschutzbehörde (in unserem Fall die Tierethikkommission der Universität Sydney) genehmigt werden.
1. Euthanasie der Tiere und Muscle Dissection
- Übertragen Sie die Maus von der Halteraum an einem separaten Raum, wo sie mit einer intraperitonealen Injektion von Pentobarbital-Lösung (30 mg / kg) mit der Maus Behandlungsmethode von Shimizu 25 detaillierte euthanasiert. Platzieren Sie die Maus wieder in seinen Käfig.
- Sobald die Atmung der Maus ist seit mehr als 1 min gestoppt, testen Sie die Fuß-Rückzugreflex durch leichtes Zusammendrücken des Fußes und der Hornhautreflex durch leichtes Streichen Sie die Hornhaut. Nur wenn Reflexe fehlen kann die Maus für die Präparation vorbereitet werden.
- Wenden Sie sich an einen Atlas von Nagetier Anatomie wie Chiasson 26 und / oder suchen die Hilfe eines experienced Anatom, bevor Dissektion der Muskeln von Interesse. Jeweils entfernen Haare von der Haut unter Verwendung eines kleinen elektrischen Rasierers vor dem Öffnen der Haut, um den Muskel freizulegen.
HINWEIS: Die Präparation wird für jede anatomisch ausgeprägte Muskel abweichen. - Mit stumpfen Pinzette befreien den Muskel vom darüberliegenden Membranen und das umgebende Gewebe. Fassen Sie und schneiden Sie die distale Sehne um den Muskel von seinem Ansatz zu trennen.
- Sanft zu necken und schneiden den Muskel frei von umgebenden Gewebe gleich wieder zu seinem Ursprung. Kurz gesagt stellen die neu präpariert Muskel in 0,1 M phosphatgepufferter Salzlösung (PBS) oder Ringerlösung vor der weiteren Verarbeitung.
2. Vorbereiten der Muskel für Kryoschneiden
HINWEIS: Die optimale strukturelle Haltbarkeit lässt sich durch ganze Tier Perfusion erzielt wie zuvor beschrieben 27, oder Tauchfixierung (für kleine Muskeln), wie in optionalen Schritt 2.1 beschrieben werden. Jedoch4% Paraformaldehyd Fixierung anschließender Färbung mit vielen Antikörpersonden mit fluoreszierend BGT beeinträchtigen. Glutaraldehyd sollte insbesondere vermieden werden. Wenn Muskeln nicht behoben werden, müssen sie sofort eingefroren werden schnappt (fahren Sie mit 2.3).
- Optional Tauch Fixierung: Pin der Muskel zu Wachs in einer Petrischale bei Ruhelänge. Decken den Muskel mit 2% w / v Paraformaldehyd (frisch gelöst in PBS) für 2 h bei RT. Waschen Sie es mit 3 Änderungen von PBS über 30 Minuten (3 x 10 min) und setzen Sie die PBS mit 30% w / v Sucrose in PBS und Inkubation O / N bei 4 ° C.
- Stellen Formen ("Booten") vorab durch Falten von 2 cm x 1,5 cm große Stücke aus Aluminiumfolie, wie sie in 1 gezeigt ist. Ein Stück Nitrocellulose-Membran in dem Boden des Bootes. Gießen Sie vorsichtig Kryostat Einbettung Matrix (Material Tabelle) ins Boot zu einer Tiefe von 2 mm, die Pflege, um Luftblasen zu vermeiden. Setzen Sie den Muskel in das Boot, dass es in die Kugelschreiber Linien aufdie Nitrocellulose. Noch ein Einbetten Matrix, um die Muskel (Abbildung 1) vollständig bedecken.
- Pre-Label Polypropylen-Röhrchen mit einer unauslöschlichen Markierung. Geben Sie einen Tropfen Wasser in jedes Röhrchen und Chill das Röhrchen in flüssigem Stickstoff.
HINWEIS: Die gefrorenen Wassertropfen hält den Dampfdruck und verhindert Austrocknen bei längerer Lagerung -80 ° C - Mit einem Gesichtsschutz, dicke Schutzhandschuhe und ein großes Paar stumpfen Pinzette, teilweise senken eine kleine Metallbecher (3 cm Durchmesser, 8 cm tief), die 2 cm Tiefe von Isopentan in einen Behälter mit flüssigem Stickstoff für 30 Sekunden. Das Becherglas und legen Sie sie auf der Tischplatte. Mit einem kleineren Paar stumpfen Pinzette legen Sie die Form, die die Muskeln und die Einbettung in die Matrix gekühlt Isopentan. Achten Sie darauf, flüssigem Stickstoff Vermischung mit dem Isopentan zu vermeiden.
- Lassen Sie 2 Minuten für den Block, um vollständig, bevor Sie stumpfen Pinzette, um den gefrorenen Block herausheben und verschließen Sie diese in der richtigen pr einfrierene-markierte und vorgekühlte Rohr (Schritt 2.3).
- Die Röhrchen speichern vorübergehend in der flüssigen Stickstoff vor bis -80ºC übertragen. Melden Sie alle Proben in einer Tabelle der Inhalt Gefrierschrank.
3. Kryoschneiden und Fluoreszenzfärbung für En Face Bilder NMJs
- Abziehen der Aluminiumform. Im -20ºC Kryostatkammer befestigen Eisblocks am Kryostaten Futter so geschnitten, um 20 & mgr; m Kryoschnitte parallel zu der langen Achse der Muskelfasern (Abbildung 1). Pick-up die Abschnitte von Poly-L-Lysin oder Gelatine beschichteten Objektträgern.
- HINWEIS: Lassen Sie diesen Schritt aus, wenn das Gewebe vor dem Einfrieren fixiert. Nachdem man 30 Minuten für Abschnitte, um auf die Objektträger trocknen, beheben Sie diese mit einem Tropfen von 2% Paraformaldehyd in PBS über jedem Abschnitt für 15 min bei RT.
- Wäsche gleitet 3 x 10 min in PBS in einer Coplin-Gefäß und dann tauchen die Objektträger in PBS mit 0,1 M Glycin für 30 Minuten, um restliche Aldehydgruppen zu blockieren.
- Für 10 min in PBS waschen gleitet, dann Eintauchen in Methanol für 7 min (bis -20 ° C gekühlt). Diese Permeabilisierung Schritt ist eine Routine Teil Doppelmarkierung mit Fluoreszenz-BGT und Anti-Synaptophysin aber es kann sich negativ auf die Immunfärbung für einige andere Proteine.
- Wäsche gleitet 2 x 10 min in PBS dann legen Sie jede Folie in einer stabilen und nivelliert feuchten Kammer. Jeder Abschnitt mit 20 ul Blockierungslösung sofort Bedeckung (0,2% Triton X-100, 2% Rinderserumalbumin (BSA) in PBS) für 1 h bei RT. Schnitte dürfen in jeder Phase der Immunprozess austrocknen.
- Führen Sie die primären Inkubation: Unter einem Objektträger vorsichtig die überschüssige Blockierungslösung aus über jeden Abschnitt und ersetzen sie durch 20 ul Kaninchen-anti-Synaptophysin (1: 200 verdünnt in Blockierungslösung).
- Fügen Sie eine Untersteuerschieber, die mit Sperr einzige Lösung inkubiert wird. Diese "nicht-primären Antikörper Steuer'Ist wichtig, in jedem Immunfärbung laufen.
- Darauf achten, dass der primäre Antikörper bleibt an Ort und Stelle über jeden Abschnitt, schließen Sie das befeuchtete Kammer und Inkubation für 1-2 Tage bei 4 ° C.
- Überprüfen Sie jeden Abschnitt, um zu bestätigen, dass der primäre Antikörper an Ort und Stelle bleibt. Verwenden Sie eine Pasteurpipette vorsichtig spülen jede Folie mit PBS und legen Sie sie in einem Coplin-Gefäß. Alle Folien 3 x 10 min in PBS waschen.
- Führen Sie sekundäre Inkubation. Unter einem Objektträger, entfernen Sie vorsichtig überschüssiges PBS, legen Sie sie in der feuchten Kammer und können daher auf jedes Teil mit 20 ul einer Mischung, die FITC-konjugiertem Esel-Anti-Kaninchen-IgG und BGT konjugiert mit Rhodamin Tetramethyl oder einen anderen roten Fluorophor (TRITC- / redBGT; 5 g / ml) in Blockierlösung verdünnt. Inkubiere bei Raumtemperatur für 2 Std.
- Wäsche gleitet 3 x 10 min in PBS in Coplin-Glaszylinder.
- Unter einem Objektträger, entfernen Sie vorsichtig überschüssiges PBS und montieren mit einem Deckglas mit einem minimalen Volumen, glycerol-basierte, verblassen-Wider Eindeckmedium. Dichten Sie die Ränder der Deckgläser mit klarem Nagellack. Lassen Sie es schwer zu trocknen.
- Bewahren Sie die Folien im Dunkeln bei 4 ° C für bis zu einer Woche, oder bei -20 ° C für längere Lagerung (bis zu mehreren Monaten).
4. Unvoreingenommene Probenahme und En Face Imaging der motorischen Endplatten
- Blenden Sie die Folien durch die Kennzeichnung jedes Dia mit einem zufälligen Code-Nummer, die nur zu einem zweiten Forscher (nicht in die Analyse einbezogen) bekannt bleibt. Als Ergebnis der Bediener blind bleibt Behandlungsgruppen bis Quantifizierung NMJ Parameter vollständig für alle Proben ist.
- Stellen Sie den Schieber auf dem Mikroskoptisch, und zeigen Sie unter Weitfeldbeleuchtung mit TRITC-Filtersatz (63X Öl 1,3 NA-Objektiv). Bewegen Sie schrittweise (Feld für Feld) von links nach rechts und wieder zurück, bis eine Endplatte im Bereich (2A) wird angezeigt.
HINWEIS: Sampling Kriterium: Jede AChR-gefärbten Struktur, die relativ istflach und steht das Ziel (dh erstreckt <15 m in der z-Dimension) wird als eine Endplatte und für die Analyse abgebildet (Sicheln AChR Färbung darstellen Querschnitte durch Endplatten und werden deshalb ausgeschlossen). - Mit der konfokale Blende auf 1,0 Airy-Einheit und niedriger Laserleistung die Optimierung der Verstärkung und Offsetwerte für TRITC / red-BGT (532-nm-Laser) an der Endplatte, die abgebildet werden soll. Nächste optimieren FITC / Synaptophysin Fluoreszenz mit dem 488 nm-Laser. Sammeln Sie eine Z-Stapel der Endplatte mit einem 0,7 um Intervall zwischen den einzelnen optischen Scheibe. Speichern Sie die Bilder mit einem Dateinamen, die das Datum des Imaging-Sitzung, den Codenamen des Schlittens und die Anzahl der Endplatte umfasst.
HINWEIS: Die Scans unter Verwendung der 488 nm und 532 nm-Laser (FITC und TRITC) sollten gesammelt nacheinander (nicht gleichzeitig), um eine Kontamination des Kanals durch FITC Fluoreszenz von dem roten Fluoreszenzfarbstoff und umgekehrt (Durchbluten) zu vermeiden. - Wiederholen Sie die Probennahme einnd Abbildung von Schritten von 4,2 bis 4,3, bis 20 Endplatten werden aus der Folie / Probe gesammelt.
- Wechseln Sie in den nächsten codierten Rutsche und wiederholen 4,2-4,4. Wiederholen Sie dies für jedes der kodierten Objektträger.
- Sammeln Sie ein paar Bilder der Endplatten von der Steuerschieber (nicht-primären Antikörper Steuerung) mittels konfokaler Einstellungen, die optimal für die experimentellen Rutschen (die FITC-Fluoreszenz-Kanal sollte dunkel erscheinen) gefunden wurden.
- Am Ende des konfokalen Sitzungsübertragung die Bilddateien auf einen anderen Computer und eine Sicherungskopie der Originaldateien auf einer externen Festplatte oder einem Server.
5. Messung der Fläche der synaptischen Spezialisierungen in En Face Bilder
- Verwenden NIH ImageJ Freeware (http://imagej.nih.gov/ij/), um eine maximale Projektion (MIP) Bilder von jedem Z-Stapel vorzubereiten. Speichern Sie sie als TIFF-Dateien (2A & B). Dateinamen sollten das Bild Sitzungsdatum, Beispielcode Endplatte Anzahl und Fluoreszenzkanal (zB 060414_57 gehören23_7_FITC.tiff).
- Öffnen Sie die z-Projektionsbild in ImageJ. Wählen Sie den Acetylcholin-Rezeptor-Bildkanal (3A), und wählen Sie Bild> Typ> 8-Bit, um die 24-Bit-RGB-Farbbild in drei 8-Bit-Graustufenbilder auf dem Bildschirm zu konvertieren.
- Mit dem ImageJ Polygon-Werkzeug zeichnen Sie eine grobe Skizze auf der Endplatte des Interesses an der redBGT gefärbt (ACHR) Kanal Damit Sie alle offensichtlich Bunt Regionen des jeweiligen Einzel Endplatte, während unter Ausschluss jeglicher Färbung, die nicht von der Endplatte von Interesse stammt, zu schließen ( 3C).
- Tragen Sie eine minimale Intensitätsschwelle auf das Bild, indem Sie Bild> Anpassen> Schwellenwert (3E und zugehörige ImageJ Screenshots).
- Stellen Sie den Schwellenwert, um die AChR-gefärbten Teile zu isolieren, während ohne umgebenden Hintergrundsignal als Untergrenze (3E). Öffnen Sie eine zweite Window mit dem Original (Halbton-) Bild direkt neben dem Fenster für den Vergleich, um die Entscheidung über den Schwellenwert zu erleichtern. Notieren Sie den Schwellenwert für die spätere Verwendung in Kolokalisationsanalysen.
- Beibehaltung der Polygonumrisslinie um die Endplatte aus: Analysieren> Analysieren Partikel. Im Popup-Menü geben Sie die Auswahl von Größen wie: 50 bis unendlich Pixel (dies beseitigt kleine Artefakte aus elektrisches Rauschen im Photomultiplier).
- Analysieren Partikel Befehl erzeugt ein Fenster mit einer Liste von diskreten supra Schwelle Bereiche und deren Fluoreszenzintensität Werte gezählt, wie sie in dem binären Bild (Abbildung 3G und zugehörige ImageJ Screenshot) angezeigt. Kopieren Sie diese Daten in einer markierten Tabelle.
- Messen Sie die Gesamt Endplatte Bereich (Bereich innerhalb des Polygons), indem Sie: Analysieren> Messen. Daraus ergibt sich die Gesamt Endplatte Bereich. Kopieren Sie die Daten für AChR Bereichen und Intensitäten ineine Tabellenkalkulation und achten Sie auf Spalten entsprechend zu kennzeichnen, werden Zeilen für einzelne Endplatten für bestimmte Folien verwendet werden.
- Wechseln Sie auf die anti-Synaptophysin Fluoreszenzkanal und wiederholen Sie die Schritte 5,1 bis 5,5, aber für die FITC-Kanal (3B, D und F). Das Ziel ist, den Schwellenwert so einzustellen, daß es ein binäres Bild, das so eng wie möglich, mit den Begrenzungen der Färbung von Augen wahrgenommen schafft. Notieren Sie den Schwellenwert.
- Messen Sie den Überlappungsbereich durch die Anwendung wie folgt vor: Öffnen Sie die Originaldatei mit den beiden Kanal Bilder und teilen Sie es in zwei separate Bilder durch: Bild> Stapel> Stapel zu Bildern.
- Mit der Kolokalisation Plugin (heruntergeladen und aus dem ImageJ Webseite installiert) Wählen Sie: pluggin> Kolokalisation und geben die Schwellenwerte zuvor für die AChR und Nervenbahnen in den jeweiligen Kanal Abfrage b aufgezeichnetOchse. Dies wird eine Überlappung Bild in weiße Pixel (3H und zugehörige ImageJ Screenshots) zu erhalten.
- Konvertieren Sie die neu erstellte Bildüberlappung in ein Graustufenformat und Anwendung einer Schwelle auf den Maximalwert. Der maximale Schwellenwert wählt nur die weißen Pixel, die dem Überlappungsbereich der zwei vorherigen Kanälen. Nehmen Sie in der Tabelle der resultierende Bereich Wert von 'Co-Lokalisation ", die den Überlappungsbereich in Pixel darstellt.
- Bereiten Sie eine Tabelle der Datenprobe Mittel, Berechnung und Darstellung Standardabweichungen und Standardfehler als Histogramme oder Streudiagramme 20,22. Man beachte, dass der Wert von n in der Regel die Anzahl der Mäuse pro Testgruppe für statistische Zwecke.
- Plot Endplatte AChR Bereichen Streudiagramme oder Frequenzhistogramme, um zu bestimmen, ob die Daten in der Regel vor der statistischen Tests (6) verteilt sind.
6. Relative FärbungIntensitäten im Vergleich Mit Quer optische Schnitte
HINWEIS: Für dieses Protokoll verarbeiten alle Muskelproben zusammen und Bild in einem konfokalen Sitzung. Bei der Planung eines Experiments kann bis zu 30 Minuten Aufnahmezeit pro Muskelprobe.
- Schneiden 15 um Kryoschnitte quer zu der langen Achse der Muskelfasern und sammeln auf Objektträger, wie in Schritt 3.1 beschrieben.
- Führen Fluoreszenzfärbung wie in Schritten von 3,2 bis 3,13 beschrieben.
- Code die gefärbten Folien, dass die Bildgebung und Analysen werden mit dem Betreiber blind Behandlungsgruppe durchgeführt, wie in Schritt 4.1 beschrieben.
- Mit einem 40-fach-Fluoreszenz Ziel (NA 0,75) einen Abschnitt von jedem Objektträger kurz überblicken, um einen einzelnen Gewinn bestimmen und Offset-Einstellungsebene für AChR, die sich für alle die Endplatten in allen Proben Folien sein wird. Der hellste Endplatte sollte dann knapp unter 256 Grau auf der Skala sein. Diese Optimierung sollten getrennt für das zweite fluorescenc erfolgenE-Kanal (sukzessive gesammelt). Notieren Sie sich die festgelegten Verstärkungs- und Offset-Level-Einstellungen und nicht in der gesamten Imaging-Sitzung ändern sie.
- Sammeln von Bildern einer Fluoreszenzstandard Schieber (zB nicht-Bleich fluoreszierenden Perlen), mit den gleichen Parametern, zu Beginn und am Ende des konfokalen Sitzung, um mögliche Schwankungen in der Laserintensität zu erfassen.
- Verwenden Sie die AChR Kanal, um die Folie schrittweise zu scannen, um Endplatten zu lokalisieren.
- Konzentrieren Sie sich auf das einzige optische Schnittebene in jedem Mikroskop Feld, das die größte Anzahl von AChR-gefärbten Endplatten enthält zu finden.
- Fotografiere diesen einzigen optischen Abschnitt zweimal und speichern Sie die gemittelten Bild (4G).
- Mit der selben Brennebene Schalter zum zweiten Fluoreszenzkanal (Protein von Interesse) und sammeln Sie das Bild wie in Schritt 6.8. Speichern Sie die Bilddatei, einschließlich des Dateinamens: Datum der Imaging-Sitzung, Beispielcode, Bildnummer und ein Symbol, um die Leuchtstoffkanal anzuzeigen.
- Bewegen Sie die Stufe zur nächsten Feld, das einen oder mehrere Endplatten enthält und wiederholen Sie Schritt 6,8 bis 6,9. Wiederholen Sie diesen Vorgang, bis insgesamt 60 Endplatten abgebildet werden.
- Am Ende des Bildgebungssitzung Übertragung aller Dateien auf einen anderen Computer und sichern Sie sie.
- Öffnen Sie jede Original-Bilddatei und beim Betrachten des AChR Kanal, wählen Sie Bild> Stapel> Stapel auf Bilder, um die Kanäle aufgeteilt.
- Wählen Sie Bild> Typ> 8bit bis zu 8-Bit-Graustufen-Format auf dem Bildschirm zu konvertieren. Tun Sie dies für beide Fluoreszenzkanäle.
- Wählen Sie Bild> Stapel> Bilder zu stapeln. Öffnen Sie ein neues Stapel von zwei bisher getrennten 8-Bit-Bilder. Man kann dann bequem wechseln zwischen den beiden Fluoreszenzkanäle innerhalb der einzelnen Fenster.
- Verwenden Sie das Polygon-Werkzeug, um einen lin ziehene eng um den Rand des AChR Färbung (Figur 4E).
- Wählen Sie: Analysieren> Messen Sie um die durchschnittliche Pixelintensität für AChR im geschlossenen Bereich zu messen (beachten Sie die Bedeutung Zeichnen der Linie fest). Kopieren Sie diesen Wert in einer markierten Tabelle.
- Beibehaltung der gleichen Polygon-Umrisslinie (zur Definition der Fläche gemessen werden soll), auf die zweite Leuchtstoffkanals (beispielsweise 4B, D, F) zu wechseln und wählen Sie: Analysieren> Maßnahme. Dadurch wird die durchschnittliche Intensität der Färbung, die das Protein von Interesse in dem durch AChR Anfärbung definierten synaptischen Bereich ergeben.
- Wählen Sie einen Bereich von sichtbaren Endplatte Färbung wählen: Analysieren> Messen Sie die durchschnittliche Hintergrundfluoreszenzintensität zu messen. Wiederholen Sie diesen Vorgang für die andere Fluoreszenzkanal / s und kopieren Sie die Hintergrundwerte in die Tabelle der Fluoreszenzwerte.
- UnterTrakts des durchschnittlichen Hintergrundwerte aus Endplatte Werte, um die korrigierten Intensitäten für AChR und das Protein von Interesse in jeder Endplatte zu erhalten.
- Dividieren der korrigierten Endplatte Intensitätswerte für das Protein von Interesse durch den korrigierten BGT Fluoreszenzintensität, um die Fluoreszenzintensitätsverhältnisse 14,21 Ausbeute
7. Vergleich der postsynaptischen Membran AChR Dichte Mit FRET
Hinweis: Dieses Protokoll bewertet, inwieweit die AChR sind eng gepackt (<10 nm Abstand) in der postsynaptischen Membran. Die genaue Donor und Akzeptor-Fluorophor Kombination kritisch ist dieser FRET Assay. Namen und Details der Fluorophore sind in der Tabelle angegebenen Materialien. Ihre spektralen Eigenschaften in Bezug auf FRET, werden in unseren früheren Arbeiten 14,15 diskutiert.
- Bereiten festen Quergefrierschnitten, wie in Abschnitt 6.1 beschrieben. Alle Probengruppen müssen zusammen und Bild verarbeitet werdend in der gleichen konfokalen Sitzung.
- Gut mischen 2,5 g / ml Rote-BGT (FRET-Donor) mit 10 g / ml bis rot-BGT (FRET-Akzeptor) mit Blockierlösung in einem kleinen Plastikschlauch durch Auf- und Abpipettieren 12 mal. Dieses 1: 4 molare Mischung maximiert die Effizienz des FRET 14.
- Platzieren Sie jede Folie in einer feuchten Kammer, sorgfältig auf jedes Kapitel mit einem Tropfen (12 ul) der obigen Mischung und Inkubation für 1,5 Stunden bei Raumtemperatur.
- Kontrollschnitte: decken eine kleine Anzahl von Abschnitten mit 2,5 g / ml Rote-BGT (Donor nur, beschriftet C1 Kontrollen), und auch einige Abschnitte mit 10 g / ml dunkelrotes BGT (nur Akzeptor; bezeichnet C2 Kontrollen). Diese Kontrollen Inkubieren wie in Schritt 7.3.
- Wäsche gleitet 3 x 10 min in PBS und -halter Glycerinbasis, lichtbeständige Eindeckmedium (siehe Schritt 3.12).
- Führen Probenahme von Endplatten, wie in Schritt 6.7. Fluoreszenz von der Donor und Akzeptor sollte zumin Endplatten perfekt kolokalisiert aufgrund der zufälligen Bindung der Fluoreszenzmoleküle BGT.
- Steuer Bilder: Mit dem 40X-Objektiv und niedrige Laserleistung optimieren redBGT Verstärkungs- und Offset-Level-Einstellungen für Endplatten aus einem Steuerschieber C1. Optimieren weit redBGT Verstärkungs- und Offset-Ebenen für Endplatten von Steuerschieber C2. Bestätigen Sie das Fehlen einer Fluoreszenz Durchschlagen.
- Ohne Änderung der Laserleistung, Gain oder Offset-Level-Einstellungen zu bewegen, um den experimentellen Folien und sammeln Bilder (pre-Photobleich) für beide Fluoreszenzkanäle.
- Selektiv photobleach die dunkelrotes BGT über einen Teil eines einzigen Endplatte durch das Zoomen in der Scanbereich der Scanvorgang 10 mal mit dem 633-nm-Laser bei 100% Leistung. Die Fluoreszenz im gescannten Bereich sollte blass.
- Setzen Sie die Laserleistung und zu vergrößern und sammeln nach Bleich Bilder auf beiden Fluoreszenzkanäle mit den in 7.7 fest konfokalen Einstellungen.
- Berechnen Sie die FRET-Effizienz (E) aus der prozentualen Zunahme der Spender (rot-BGT) Fluoreszenz nach Photobleich der Akzeptor (dunkelrotes BGT) nach der folgenden Formel *:

* Für alle Fälle, in denen die Fluoreszenz des Donor-after photo Acceptors zunimmt.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Messung von Synaptic Umgebung am NMJ
Jede Schätzung Bereich stützt sich auf die Zeichnung einer Grenz um das Ausmaß der synaptischen Spezialisierungen definieren. Bei gesunden jungen Erwachsenen Muskeln sollten NMJ Bilder gut definierten Grenzen sowohl für AChR und Synaptophysin-Färbung (2A und B) angezeigt werden sollen. Fluoreszenzintensität sowohl für AChR und Synaptophysin steigt scharf an der Grenze zwischen dem peri-synaptischen und synaptische Abschnitt der motorischen Endplatte (5A 'und B'). Für solche Bilder ein Schwellenwert (direkt über extrasynaptischen Hintergrundfluoreszenz) leicht isolieren AChR-reich oder Synaptophysin-reichen Bereich der Endplatte (horizontale gestrichelte Linien (5A 'und B')). Im Alter von Mäusen und in einem gewissen Krankheitszuständen Endplatte Färbung für AChR weniger intensiv sein kann AChR Cluster Kanten unscharf wird und dieWieder können höhere extrasynaptische Autofluoreszenz (; 17,22 5D) sein. Fluoreszenzfärbung mit undeutlichen Grenzen können Fehler in Schätzungen der synaptischen Bereich einzuführen. In allen Fällen das Ziel ist, eine Schwelle, die ein binäres Bild in Form und Größe dem AChR- oder Synaptophysin reichen Gebieten ergibt, wie sie in der ursprünglichen, Halbtonbild erscheinen dem Auge zu wählen. Durchführung der Analyse blind gegenüber der Behandlungsgruppe sollte das Risiko einer subjektiven Bias in der Schwellenwertschritt (Schritt 4.1) zu reduzieren. Manchmal Ohnmacht oder verschwommenes Endplatte Bilder können vom suboptimale Verarbeitungs führen. 2C und D zeigt ein Beispiel für ein Bild mit niedriger Qualität Endplatte aus einem gesunden 2 Monate alten Maus. Unscharfe Kanten und schwache Synaptophysin Färbung kann in diesem Fall von partiellen Auftauen und Wiedereinfrieren des Muskels vor Kryoschneiden entstanden sind. Einige Abschnitte der jungen gesunden (Positivkontrolle) Muskel sollte geschnitten werden, einnd parallel Versuchsproben verarbeitet werden, um sicherzustellen, dass jegliche Verschlechterung zeigt sich in den Bildern NMJ nicht aufgrund von Problemen mit Immunfärbung. Chargen von Bildern durch suboptimale Verarbeitungs kompromittiert sollte von Analysen ausgeschlossen werden.
Für en face Z-Stapel-Bildern ist 15-20 Endplatten eine angemessene Stichprobenumfang zur Abschätzung synaptischen Bereichen. Eine breite Vielfalt in den Formen und Größen von NMJs innerhalb einer bestimmten Muskel gefunden. Streudiagramme zeigen einen beträchtlichen Bereich der AChR reichen Gebiet zwischen Endplatten des vorderen Schienbeinmuskel eines einzelnen Maus (6A). Dennoch ist die durchschnittliche AChR Bereich (auf der Basis von 15 bis 20 en face Endplatte Bilder) war ähnlich wie bei den sieben Proben Mäuse (~ 200 m 2; 6A). Die Fläche der Endplatte Synaptophysin Färbung ebenfalls deutlich unter den Endplatten aus einem bestimmten Muskel variiert. Wiederum unter Verwendung einer Probengröße von 15-20 Endplatten die mittlere Fläche der Endplatte Synaptophysin war Similar unter den 7 Mäusen untersucht (~ 170m 2, 6B). Frequenz-Histogramme von gepoolten Daten zeigten etwa Normalverteilungen für den Bereich der Endplatte AChR und Synaptophysin (6A 'und B'). Jedoch eine Normalverteilung der synaptischen Bereiche nicht bei Krankheitszuständen, wie angenommen werden Myasthenia gravis 16,20. Dies kann die Wahl der statistischen Test beeinflussen.
Tabelle I listet Bereiche vor und nach dem synaptischen Spezialisierungen für NMJs für gesunde 2 Monate alt (junge Erwachsene) weibliche C57BL / 6J-Mäusen aus früheren Studien. Die Bereiche der sowohl vor als auch postsynaptischen Spezialisierungen sank mit sitzender Alterungs 22. AChR-Bereich wurde auch deutlich bei Mäusen mit IgG von Anti-MuSK Myasthenia gravis Patienten injiziert reduziert 17,21. Myasthenie Mäuse mit der Cholinesterase-Inhibitor Drogen, Pyridostigmin behandelt, erscheint eine weitere signifikante Reduktion der Endplatte AChR Bereich 20 </ Sup>.
Relative Intensität der Endplatte Fluoreszenzmarkierung
Die relative Intensität der Immunfluoreszenz-Markierung kann Veränderungen in der Dichte der synaptischen Protein von Interesse mit dem Alter, Genotyp und / oder Krankheitszustand zu offenbaren. AChR Fluoreszenz (rot-BGT oder dunkelrotes BGT) zuerst verwendet wird, um die Position des NMJ definieren. Dann wird die Helligkeit der Fluoreszenz in der 8-Bit-AChR-reichen Bereich verwendet werden, um Veränderungen in der Konzentration des Proteins von Interesse, gegenüber den Kontrolltieren zu bewerten. Im Querschnitt die Endplatte AChR werden normalerweise als halbmondförmige Gestalt, aber diese Form ist oft unregelmäßig (4A, C, E, H). Niedrige Intensität der Hintergrundfluoreszenz zeigt in der Regel, ob ein Patch von AChR Färbung stellt einen einzelnen Endplatte oder zwei getrennte Endplatten auf benachbarten Muskelfasern. Viele synaptische Proteine (wie Rapsyn, Moschus und SRK)mit AChR kolokalisiert an der Endplatte (4A - D). Immunfluoreszenzfärbung mit phospho-spezifischen Antikörper können auch verwendet werden, um die Wirkung der Versuchs Eingriffe am Phosphorylierungsstatus insbesondere postsynaptischen Membranproteine 21 zu vergleichen.
Die Zuverlässigkeit und Reproduzierbarkeit der Messung der Fluoreszenzintensität hängt stark von der Unversehrtheit des gefrorenen Muskel und die Qualität der Immunfärbung. Muskeln sollten seziert und sofort schockgefroren oder Parafest (innerhalb von Minuten nach dem Tod des Tieres) zu degenerativen Veränderungen an den NMJ vermeiden. Immunfärbung hängt stark von der Qualität der Reagenzien und die Optimierung der Färbeprotokoll auf spezifische Antikörper. Für jede neue Charge von primärer Antikörper-Immunfärbung Pilotversuche sind erforderlich. Frisch geschnittene Gefrierschnitten von gesunden jungen Muskeln mit serieller 2-fache Verdünnungen des primären Antikörpers inkubiert. Abekannt zuverlässige sekundäre Antikörper verwendet wird, und die Ergebnisse werden verglichen. Wenn das Protein von Interesse bekannt ist, um zu der NMJ beschränkt werden dann der beste Antikörper-Konzentration ist diejenige, die das höchste Verhältnis von NMJ Fluoreszenzintensität relativ zu der in extrasynaptischen Teilen des Muskels (Hintergrundfluoreszenz) gefunden ergibt. Extrasynaptische (vermutlich unspezifische) Fluoreszenzintensität sollte in der Regel höchstens 15% der Endplatte Fluoreszenzintensität. Ebenso "kein Primärantikörperkontrolle 'Abschnitte (nur mit sekundären Antikörper) sollte dunkel erscheinen, die bestätigt, dass der sekundäre Antikörper nicht unspezifisch binden. Die Qualität der verschiedenen Chargen (polyklonalen) Sekundärantikörper merklich so alternative sekundäre Antikörper Chargen variieren sollte vor Festlegung eines Standardprotokolls verglichen werden. Die ideale Test für die Spezifität der Immunfluoreszenz einen direkten Vergleich der Abschnitte von Wildtyp-Mäusen und Negativ-Kontrollabschnitte von Mäusen tHut fehlt das Protein von Interesse (Gen-Knockout). Die folgenden Referenzen beschreiben die Quantifizierung der Endplatte Fluoreszenzfärbung für MuSK, Rapsyn, dystroglycan, src und AChR 13,14,18,21.
Kleine Probengrößen einzuführen Fehler in Schätzungen der relativen Fluoreszenzintensität. Einzelne Endplatten wesentlich in der Fluoreszenzintensität variiert. Vermutlich Variabilität zwischen den Endplatten innerhalb einer bestimmten Muskel spiegelt die Vielfalt der NMJ Struktur und Zufall Unterschiede in der optischen Strecke abgetastet. Eine Erhöhung der Anzahl von Stirnplatten abgetastet ergibt eine stabilere Schätzung der durchschnittlichen Fluoreszenzintensität (Abbildung 7). Um die durchschnittliche Endplatte Fluoreszenzintensität 40-60 Endplatten voneinander Muskelprobe sollte gemittelt werden zu schätzen.
AChR-AChR FRET
Jede AChR ist ein Pentamer mit zwei Bindungsstellen für BGT (jeweils auf einer alpha-Untereinheit). Die Bindung vonred-BGT und dunkelrotes BGT an diesen beiden Standorten würde eine Donor-Akzeptor-Abstand von etwa 9 nm 28 bis 30 ergeben. So geringe Effizienz FRET nachgewiesen werden kann, noch bevor AChRs montieren in Cluster 14. Allerdings ist die Effizienz von FRET bei murinen Endplatten etwa verdoppelt postnatal, im Einklang mit der effizienteren Einbeziehung AChRs in einen dicht gepackten Gitter postsynaptischen Membran 14. Verwendung der roten BGT und dunkelrotes BGT als FRET-Donor und Akzeptor (jeweils), die Endplatten von 1-2 Monate alten Mäusen durchschnittlich FRET Wirkungsgrade im Bereich von 20 bis 37% (Tabelle 2). FRET Effizienzen von 20% oder mehr sind gedacht, um eine dichte Packung des AChR 14 darstellen. Endplatte FRET-Effizienz wurde leicht nach Denervation 14 reduziert und deutlich reduziert, nachdem Mäuse wurden mit IgG von Anti-Moschus positive Myasthenia gravis Patienten injiziert 18. Dies sind Bedingungen, bei denen einzelne AChR sind weniger dicht in die postsy gepacktnaptic Membrangerüst von der postsynaptischen MuSK / Rapsyn System 31.
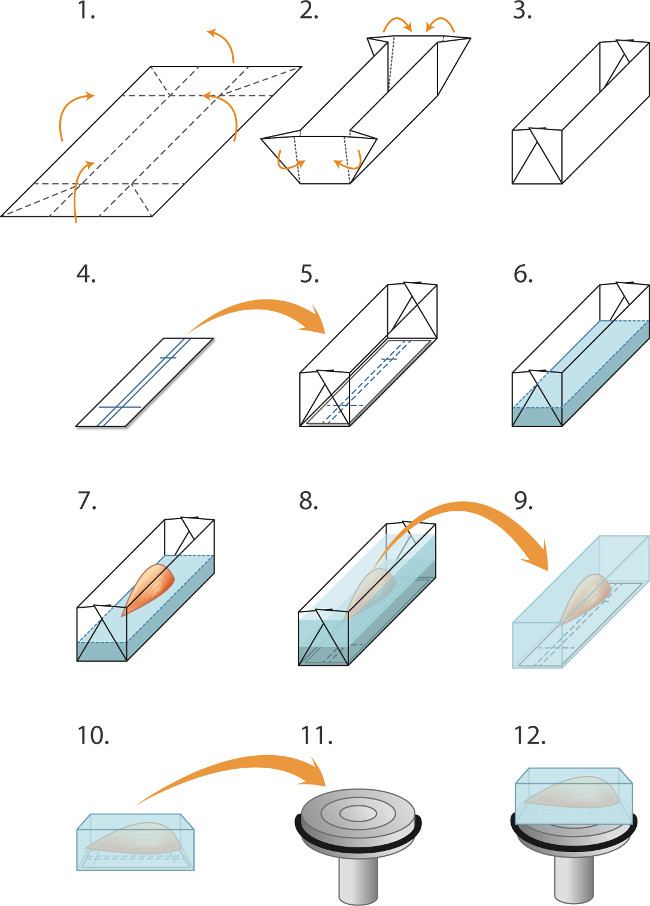
Abbildung 1. Embedding und Gefrier Muskeln für Kryoschneiden (1-5) vorbereiten Formen ('Boote') vor dem Einfrieren einen Stapel von Muskeln:. (1) Aluminiumfolie wird in Rechtecke (2,0 x 3,0 cm) und schneiden (2). (3) gefaltet, wie angeklagt, eine Form / Boot erstellen. (4) Rechtecke aus Nitrocellulose-Immunoblotting Papier werden geschnitten, um in die Form passen und ein Kugelschreiber verwendet wird, um Linien zur Orientierung des Muskels zu regieren. (5) Die Nitrozellulose Rechteck wird in der Form angeordnet. (6-8) Das Einbetten und Einfrieren der Muskeln: (6) Kryostat Einbettungsmatrix Fluid sanft in die Form (auf der Oberseite der Nitrocellulose) gegossen, um eine Tiefe von 2 mm. (7) einer feinen Pinzette mit dem nitrocellu in die Einbettungsmatrix verwendet, um den Muskel zu senken, durch seine Sehne, fluchtendverlieren Entscheidung (8) Zusätzliche Einbettung Matrix wird sanft in die Form gegossen, um den Muskel zu decken, wobei darauf zu vermeiden, Blasen. Der eingebettete Muskel wird dann schockgefroren, in Röhrchen versiegelt und bei -80 ° C gelagert, wie im Text beschrieben. (9-12) Vorbereitung Kryoschneiden: (9) bildenden Zangen werden verwendet, um abziehen der Aluminiumform und der gefrorene Block wird dann in der -20 ° C Kryostatkammer, (10) Die Kennzeichnung im Nitrocellulose werden verwendet, um ausgerichtet platziert der Muskel parallel zur Fläche des Futters (11) zur Längsschneide (12). Ein Flüssigkeitstropfen Kryo-Einbettungsmedium wird verwendet, um den Block auf die gekühlte Spannfutter zu befestigen. Stumpfen Pinzette verwendet, um das gefrorene Muskel Block manipulieren. Um Querschnitte zu erhalten, wird der Block statt montiert, dass der Muskel ist senkrecht zu der Oberfläche des Spannfutters (nicht gezeigt).

Fild 2. En face Bilder NMJs vom M. tibialis anterior von 2 Monate alten weiblichen C57BL6J Mäusen. Die frische Muskel war Schnappeingefroren und die Abschnitte fest auf der Folie wie in Schritt 3.2 beschrieben. MIP-Abbildungen der z-Stapel wurden erhalten, wie in dem ersten Protokoll beschrieben. (A) der roten BGT Färbung zeigt einen einzigen motorischen Endplatte, die aus zwei Sätzen von AChR-reiche Primär postsynaptischen Rinnen (en face Ansicht). (B) Synaptophysin Färbung mit FITC-konjugierten Sekundärantikörper offenbart die präsynaptischen Nervenende, besetzen die primäre synaptische Dachrinnen. Ein Teil des pre-terminalen Axon ist auch in dem oberen Teil der Tafel sichtbar. (C & D) Ein Beispiel für eine schlechte Qualität NMJ Bildes von einem jungen gesunden Maus. Die Färbung ist schwach und die Grenzen der prä- und postsynaptischen Spezialisierungen sind unscharf. Dies wurde von Mängeln in den t zurückzuführenfür den primären Antikörper Inkubation er Verarbeitung des Gewebes und / oder zu wenig Zeit erlaubt. Maßstabsbalken in Panel D für 10 & mgr; m. Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.

Abbildung 3: Die Schritte in der Verarbeitung von en face NMJ Bilder (wie in Protokoll 1 beschrieben). (A & B) Original Dauerton MIP-Bilder zeigen rot-BGT Fluoreszenz enthüllt AChR und grüne Immunfluoreszenz für Synaptophysin auf. (C) AChR-Färbung nach der Umwandlung in eine 8-Bit Graustufenbild und die Nutzung des Polygon-Werkzeug, um die Endplatte abzugrenzen (dünne gelbe Linie). (D) Endplatte Grenzlinie über dem grünen Bild Synaptophysin durchgeführt. (E) Application eines minimalen Intensitätsschwellen Befehl, um ein binäres Bild, das überschwelligen roten BGT (AChR) Fluoreszenz isoliert wird. Die Sequenz von ImageJ Benutzeroberfläche Screenshots auf der linken Seite der Bilder angezeigt. (F) Binary Synaptophysin Bildes nach Anlegen eines separaten Schwellen. (G) Identifikation der diskreten überschwelligen AChR-reiche Domänen innerhalb der Endplatte durch die Anwendung der Partikel Befehl Analysieren auf den binären roten BGT Bildes. Die entsprechende ImageJ (links von Tafel (G) zeigt die Eingangsdaten erforderlich. Die Mindestgröße der überschwelligen erforderliche Pixelbereiche eingegeben werden soll. (H) Identifizierung der Überlappungsbereiche der binären Synaptophysin und AChR Bilder. Overlap ist von weißen Pixeln dargestellt . ImageJ Benutzeroberfläche Screenshots (unten Panel H) zeigen die Schritte auf dem binären Bild Überschneidungen kommen. Die gewählte minimale Intensität Grenzwerte für jeden Fluoreszenz channel muss innerhalb Fenster 'Kolokalisation' eingegeben werden. Maßstabsbalken entspricht 10 & mgr; m. Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.

Abbildung 4. Beispiele für optische Querschnitte, die den relativen Fluoreszenzintensitäten zu vergleichen (Protokoll 2). Der Muskel wurde schockgefroren und die auf der Folie fixierten Schnitten, wie in Schritt 3.2. (A & B) eine einzige Endplatte mit weit doppelt markierten beschrieben -rot-BGT (AChR, hier in blau-Pseudo gezeigt) und FITC-anti-Rapsyn zeigt die Co-Lokalisation der beiden interagierenden Proteine in der postsynaptischen Membran (C & D) zwei Endplatten benachbarter Muskelfasern Anzeige kolokalisiert AC. hR und Moschus. (E & F) eine Endplatte für AChR und -dystroglycan (-DG) doppelt markierten. Die -DG sich gleich um die Muskelfaser Umfang aber ist an der Endplatte angereichert (Maßstab in F, für Platten AF: 25 um) (G - I) Isolieren einer Endplatte zur Intensitätsmessung (G) Ein typisches Mikroskopfeld,.. mit drei dunkelrotes BGT-gefärbten Endplatten (Maßstab in Platte G: 40 & mgr; m (H) Ein vergrößertes Bild des Box-Endplatte (I) Das gleiche Endplatte zu einem 8-Bit-Graustufenbild umgewandelt und abgegrenzt mit dem Polygon.. . Werkzeug von ImageJ (dünne gelbe Linie) Durchschnittliche Fluoreszenzintensität innerhalb dieser Grenzlinie gemessen. (Maßstab für die K & E: 10 um) Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.

Abbildung 5. Einfluss der Bildqualität bei der Beurteilung der synaptischen Bereich. (A & B) Hochwertiges en face Bilder eines gesunden NMJ von einem 2 Monate alten Maus auf dem roten BGT und anti-Synaptophysin Fluoreszenzkanäle angesehen. (A '& B ') Fluoreszenzintensitätsprofile an die Linie über die Endplatte für A und B gezeichnet entspricht. Die horizontale rote gestrichelte Linie die Mindestschwelle verwendet, um die Binär-Bild zu erzeugen. (C & E) Endplatte aus einer alten Maus. Endplatte Synaptophysin Färbung im allgemeinen weniger intensiv ist. (C '+ E') Intensitätsprofile zeigen eine hohe extrasynaptischen (Baseline) Fluoreszenz Schwankung der Synaptophysin (FITC) Kanal (backMasse), die Bestimmung einer geeigneten Schwelle beeinflußt. Vieles davon ist Breitspektrum-Gewebeautofluoreszenz. Maßstabsbalken repräsentieren 10 um. Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.

Abbildung 6. Variabilität der synaptischen Bereichen unter NMJs innerhalb eines Muskels und von Mäusen. (A) Streudiagramme zeigen die Gesamt AChR reichen Bereich der Endplatten aus dem M. tibialis anterior von sieben naive 2 Monate alte weibliche C57BL / 6J-Mäusen nach Autor NT erhalten. Jedes Symbol stellt eine Endplatte. Jeder Balken stellt den Mittelwert ± Standardabweichung für die Endplatten von einer Maus entnommen. (B) Streudiagramme, die die Synaptophysin reiches Gebiet für die gleichen Endplatten. (A ') (B ') Häufigkeitsverteilung für die Synaptophysin reichen Bereich der Endplatten (gepoolte Daten). Bitte klicken Sie hier, um eine größere Version dieses Bild anzuzeigen.

Abbildung 7. Wirkung der Stichprobengröße nach Schätzungen der Endplatte Fluoreszenzintensität. Transversale optische Schnitte wurden verwendet, um die Fluoreszenzintensität (in beliebigen Einheiten) bei 40 60 Endplatten von der vorderen Schienbeinmuskel eines gesunden 2 Monate alten Maus messen. Kumulierte Durchschnittswerte werden gegen die Anzahl der Endplatten im Durchschnitt eingeschlossen aufgetragen. (A) Die Endplatte rot-BGT Fluoreszenzintensität nach Autor AV erhalten. (B) Anti-synaptophysin Immunfluoreszenzintensität für die gleichen Endplatten wie in Abbildung A. (C) Endplatte dunkelrotes BGT Fluoreszenzintensität von einem zweiten Muskelprobe nach Autor NG erhalten. (D) Anti-Rapsyn Immunfluoreszenzintensität aus den gleichen Endplatten wie in Panel C.
| Studie | Muskel | Anzahl der Mäuse | AChR Bereich | Synaptophysin Bereich | Überlappungsbereich |
| Mittelwert ± Standardabweichung (mgr; m 2) | Mittelwert ± Standardabweichung (mgr; m 2) | Mittelwert ± Standardabweichung (mgr; m 2) | |||
| Morsch et al. (2012) 1 | gastrocnemius | 3 | 181 ± 7 | 163 ± 24 | |
| Membran | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch et al. (2013) 1 | tibialis anterior | 3 | 166 ± 26 | 117 ± 21 | nd |
| Cheng et al. (2013) 2 | tibialis anterior | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| Tse (unveröffentlicht) 2 | tibialis anterior | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| 1 Aufdruck auf einem Zeiss LSM 510 Meta-Mikroskop, aber mit fester Verstärkung und Offset Ebenen. Schwellen und synaptische Bereichen Messungen Metamorph Software von MM. | |||||
| 2 Aufdruck auf einem Leica DM IRE2 Mikroskop und analysiert folgende aktuelle Protokoll, das von der angegebenen ersten Autor, blind gegenüber Behandlungsgruppe. | |||||
| nd nicht bestimmt. | |||||
Tabelle 1 Synaptic Bereiche für NMJs von 2 Monate alten weiblichen C57BL / 6J-Mäusen (gesunde Kontrollen)
| Studie | Muskel | FRET-Wirkungsgrad (%) * | Range (%) |
| (Mittelwert ± SEM) | |||
| Brockhausen et al. (2008) | tibialis anterior | 24 ± 1 | n / a |
| Cole et al. 2010 | tibialis anterior | 26 ± 1 | 22 - 30 |
| Morsch (unveröffentlicht) | Membran | 37 ± 1 | 24-47 |
| Ghazanfari (unveröffentlicht) | tibialis anterior | 30 ± 1 | 20 - 45 |
| * FRET zwischen Rot-BGT und dunkelrotes BGT (Förster-Radius für FRET-Paar = 51A (Life Technologies). | |||
| na Daten nicht verfügbar | |||
Tabelle 2. Wirkungsgrade für AChR FRET von jungen erwachsenen Mäusen C57BL6J Mäuse
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Die hier beschriebenen Protokolle konnten wir zuverlässig messen und zu quantifizieren Veränderungen in den Eigenschaften der NMJ in einer Reihe von Bedingungen, einschließlich der normalen Alterung und Krankheitszuständen. Die für en beschriebenen Methoden stellen NMJ Bilder wird es den Forschern ermöglichen, den Bereich der prä- und postsynaptischen Spezialisierungen und den Bereich der synaptischen Überlappung / Ausrichtung zu vergleichen. Um die relative Intensität der prä- und postsynaptischen Proteine das zweite Protokoll, das optische Querschnitte verwendet zu vergleichen, wird bevorzugt. Das dritte Protokoll spezifisch testet für Änderungen in der Nähe der Verpackung der AChR in der postsynaptischen Membran.
Spezifität Kontrollen sind unerlässlich in Immunfluoreszenzmikroskopie. Bei der Verwendung eines primären Antikörpers für die indirekte Immunfluoreszenz ist es notwendig, zunächst zu gewährleisten, daß es sich spezifisch an sein Zielprotein in den Muskelpartien. Verschiedene Arten von Gewebeverarbeitung und Fixierung kann die Spezifität der differentiell verändernAntikörper. Es ist wichtig, dass die Immunfluoreszenzfärbung (für sagen Rapsyn) wirklich mit AChR an der motorischen Endplatte konzentriert bestätigen. Negative Kontrollabschnitte müssen geprüft werden, um sicherzustellen, dass die Antikörper-Bindung spezifisch ist. - / - Mäusen Zum Beispiel würde die beste negative Kontrolle für Rapsyn Immunofluoreszenz Schnitte von Rapsyn sein. Diese sollten keine Endplatte Färbung mit anti-Rapsyn zeigen. Die unspezifische Fluoreszenz können auch durch endogene fluoreszierende Chemikalien im Gewebe (Autofluoreszenz) oder von nicht-spezifischen Bindung von dem fluoreszierenden Zweitantikörper-Konjugat auftreten. Solche Fluoreszenz wird oft von Aldehyd-Fixierung verschlechtert. Zusätzlich kann TRITC-BGT Anfärbung Endplatten manchmal im FITC Fluoreszenzkanal detektiert werden, und diese Fluoreszenzdurchschlagen könnte mit bestimmten FITC Immunofluoreszenz verwechseln. Zum Schutz vor den drei letztgenannten Formen der nicht-spezifischen Fluoreszenz zu schützen, sollte jede Charge von Folien, die gefärbt sind einige 'n sindo-Primärantikörper Kontrolle "Abschnitte (Schritte 3.7 und 4.6). Bilder der Endplatten aus diesen Steuerabschnitten sollten denen von den experimentellen Folien verglichen werden, um sicherzustellen, dass die indirekte Immunfluoreszenzfärbung von NMJs reflektiert wirklich die Bindung des primären Antikörpers.
Quer konfokale Schnitte sind besonders nützlich für die Beurteilung der Unterschiede in der relativen Intensität der Immunfärbung an der Synapse. In Quer konfokale Schnitte es einfacher ist, präzise Kolokalisation von synaptischen Proteinen beurteilen. Der halbmondförmige Endplatte Profil stellt nur ein Beispiel Ausschnitt durch die NMJ in Frage. Allerdings ist der Hintergrund (extrasynaptischen) Fluoreszenz in der Regel niedriger, um en face z-Projektionsbilder verglichen. So kann es einfacher sein, um "echte" (spezifischen) Immunfärbung unterscheiden und stellen fest konfokalen Verstärkungs- und Offset-Werte mit Quer optische Schnitte 13-15,18. Zum Beispiel wird in einem Mausmodell von Myasthenia gravis (where Endplatte AChR Färbung deutlich reduziert) Endplatten wurden deutlich in Quer optische Schnitte 18,21 abgegrenzt. Unterschiede in der durchschnittlichen Intensität der Fluoreszenz bei der NMJ dürften veränderter Dichte des Zielproteins im synaptischen Spezialisierung zu reflektieren. Ein Nachteil ist, dass in einigen Situationen ein Strukturwandel in das Zielprotein oder Verschluss der Antikörperbindung durch benachbarte Proteine könnten veränderte Färbeintensität erklären.
Die Versuchsplanung erfordert einige Überlegungen. In vielen Fällen wäre das Experiment sollen die Auswirkung eines Transgens, Knock-down oder Krankheitszustand auf der Größe des NMJ testen. Die Versuchsprobe Gruppe könnte dann zu gesunden jungen (Wildtyp) Mäuse des gleichen Geschlechts und genetischen Hintergrund zu vergleichen. Ausgangswerte für den Bereich der Endplatte Synaptophysin, AChR und synaptische Überschneidungen für mehrere Muskeln werden in Tabelle 1 angegeben. Probengröße hängt vom Grad des Tieres ab-bis-Tier Variation innerhalb Behandlungsgruppen und der Effektgröße (Differenz der Mittelwerte für die experimentellen gegenüber der Kontrollgruppen pro Standardabweichung). Wenn die Analyse auf gute Bildqualität beschränkt ein faires Maß an Übereinstimmung gefunden wurde in das Probenmittel zur Endplatte Bereichen bei gesunden 2 Monate alten weiblichen C57BL6J Mäuse (6A und B). Somit war es möglich, signifikante 30-40% Verringerung der synaptischen Bereich in Mäusen, die mit IgG von anti-MuSK-positive Myasthenia gravis Patienten injiziert zeigen, im Vergleich zu Kontrollen mit einem Stichprobenumfang von drei Mäusen 17,20,32. Ältere Mäuse zeigten größere Tier zu Tier Variation Endplatte Parameter als jungen Mäusen 22. Folglich Versuche mit älteren Mäusen könnte größeren Stichproben erforderlich.
Wenn das Hauptanliegen ist es, die Größe der en face Endplatte dann die Verstärkungs- und Offset-Level-Einstellungen messen sollte für jeden einzelnen NMJ optimiert werden. IndivIdual NMJs beträchtlich in der Helligkeit des AChR und Synaptophysin Färbung, insbesondere bei Krankheitszuständen untersucht werden variieren. Darüber hinaus ist die Intensität der außer synaptischen (unspezifisch) Fluoreszenz ist oft höher und Variable in Muskeln der älteren Tieren, im Vergleich zu denen von gesunden jungen Tieren (5C und D). Die 1-256 Grau sollten umfassend genutzt werden, um die klanglichen Informationen, die in den letzten Bildern erhalten wird, zu maximieren. Dies beinhaltet die Einstellung der Verstärkung und Offsetwerte für jeden NMJ für die ein Z-Stapel entnommen werden soll. 5D zeigt ein Beispiel eines Bildes, wo NMJ Tonwertinformationen könnte kritisch bei der Festlegung der Grenzen der Fläche von Pre-und Post sein synaptischen Spezialisierungen.
Messungen der synaptischen Bereiche können an verschiedene Muskelpräparaten und Experimente angewendet werden. Die meisten unserer Messungen der synaptischen Bereiche Längsgefrierschnitten von Schnapp beschäftigt gefrorenen Muskeln. Einfrieren des Muskel vor der Fixierung beibehält, das die Antigenität eines breiten Spektrums von Proteinen. Wenn mit dem Antigen, Para Fixierung und Saccharose Infiltration vor Kryoschneiden (Schritt 2.1) kann eine bessere Konservierung der NMJ Struktur bereitzustellen kompatibel. Optimaler Strukturerhaltung könnte durch kardiale Perfusion mit Para erhalten. Artefakte des Einfrierens und Sektionierung vollständig durch Markierung Endplatten auf der Oberfläche der intakten Muskeln und Abbildungs NMJs auf Faszikel vom festen Muskel 21 aufgezogen vermeiden. Unabhängig von der Vorbereitung, die Verfahren zur Probenahme, Bildgebung und Quantifizierung Bereich bleiben unverändert (Protokoll Schritte 4-5). Einheitliche Anwendung der Blindprobennahme, Abbildung und Analyse-Protokolle (mit verschiedenen Betreibern, verschiedene Proben von Mäusen und unterschiedlichen Zeiten), können in relativ reproduzierbare Durchschnittswerte (Cheng et al. Und Tse Ergebnisse in Tabelle 1 vergleichen) führen.
"> Die Endplatten haben als Werden" fragmentierte "in einer Vielzahl von Krankheitszuständen, beschrieben. Beispielsweise wird in alternden Maus Muskeln, sporadischen Degeneration einer Muskelfaser (gefolgt von der Regeneration) resultierte in Umbau der Brezel artigen Endplatte AChR Plaque bilden mehrere kleinere AChR Cluster 6. In Mäusen, die mit IgG von Anti-MuSK Myasthenia gravis Patienten injiziert, war die Fragmentierung der Endplatte und nicht anders. Die Endplatte AChR Brezel weitgehend verteilt, und hinterlässt eine Konstellation von kleinen (<4 m 2) AChR 'Mikroaggregaten '20,21. Diese beiden Beispiele machen deutlich, dass die Größenverteilungen für AChR Cluster an Endplatten der Kontrolle gegenüber Tierversuchen 21 zu vergleichen.Es wurden andere Verfahren zur Bewertung der synaptischen Bereich oder Färbeintensität im NMJ gemeldet. Motorischen Endplatten manchmal gefaltet, dass die z-Projektionsbilder zweidimensionale welche hier könnte syn unterschätzenaptic Bereichen. Dreidimensionale konfokalen Rekonstruktionen könnte genauere Maßnahmen bieten, wenn absolute synaptischen Bereich muss definiert 33 werden. Ein wesentlicher Vorteil des hier beschriebenen z Projektions-Protokoll ist jedoch seine relative Einfachheit, die eine große Anzahl von Abschlussplatten zugelassen wurde, um aus mehreren Behandlungsgruppen und zuverlässige Identifizierung von möglichen Veränderungen gemessen werden. Das Protokoll für den Vergleich Endplatte Färbeintensitäten kann zur Untersuchung der Veränderungen der Konzentrationen der verschiedenen synaptischen Proteinen angepaßt werden. Das Verfahren ist jedoch begrenzt durch die Forderung, dass alle Proben zur Immun dann während der gleichen Sitzung konfokalen abgebildet verarbeitet. Eine aktuelle Studie von Yampolsky et al. 5 beschrieben, ein Verfahren zur Messung Endplatte AChR Dichte, die einen Beitrag zur Überwindung dieser Einschränkung könnte. In dieser Studie wurden die gleichen Bereiche der Endplatten an mehreren unterschiedlichen Laserleistungseinstellungen abgebildet. Die Steigung der Beziehung zwischen Laserleistung und rhodamine-BGT Fluoreszenzintensität wurde verwendet, um relative Änderungen AChR Dichte an Endplatten in verschiedenen Mäuse 5 beurteilen. Dieses Verfahren könnte für den Vergleich AChR Intensität in Proben im Laufe eines längeren Studie zu verschiedenen Zeiten dargestellt werden.
AChR-AChR FRET bietet konkrete und ergänzende Informationen über die Organisation der Endplatte AChRs. Die elektronenmikroskopische Autoradiographie unter Verwendung von 125. I-α-BGT hat gezeigt AChR dicht mit einer planaren Dichte von 10 4 m -2 unmittelbar unter jedem präsynaptischen Ort des Senderfreigabe verpackt werden, während dann angrenzenden Membran Einfaltungen enthalten viel niedrigeren Dichten AChR 34. AChR-AChR FRET macht es relativ leicht, (submikroskopische) Änderungen AChR Verpackung zu detektieren. Eine Verringerung der FRET-Effizienz reflektiert eine submikroskopische Umverteilung von AChR in der postsynaptischen Membran, die nicht von einer Änderung der durchschnittlichen BGT Fluoreszenzintensität detektiert werden könnten. Mehrere fAkteure könnte zu einer Änderung in der Effizienz des FRET. Dazu gehören die Donor-Akzeptor-Abstand und die relative Orientierung sowie die molekulare Umgebung 35,36. Eine Verringerung der Endplatte FRET-Effizienz möglicherweise aus einer Änderung in der Geometrie des AChR Gitter entstehen. Jedoch wahrscheinlich wäre es aufgrund einer Verringerung des Prozentsatzes der AChR, der in die nanoskaligen postsynaptischen Molekülgitter 14 gepackt sind.
Verlust der Verbindung zwischen Motorneuronen und Muskelfasern scheint die unmittelbare Ursache der Muskelschwäche in Motoneuronerkrankung und in sitzende Alterung 9,22,23 sein. Gemeinsame Methoden und Parameter zur Messung NMJs sollte es einfacher für die verschiedenen Forschungsgruppen zu vergleichen und veröffentlichten Ergebnisse zu machen. Die gemeinsame Nutzung von detaillierten Protokolle (und zukünftige Verbesserungen auf sie) kann dazu beitragen, beschleunigen Fortschritte im Verständnis der Mechanismen der NMJ Wartung und wie sie in Krankheitszuständen beeinflusst werden.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
