

Abstract
הצומת העצבית-שהרירית (NMJ) היא סינפסה הגדולה, כולינרגית הממסר שדרכו הנוירונים מוטוריים יונקים לשלוט התכווצות שרירים מרצון. שינויים מבניים בNMJ יכולים לגרום לכישלון עצבית, וכתוצאה מכך חולשה, ניוון ואף למוות של סיבי השריר. מחקרים רבים בחנו כיצד שינויים או מחלה גנטיים יכולים לשנות את המבנה של NMJ העכבר. למרבה הצער, זה יכול להיות קשה כדי להשוות ישירות ממצאים ממחקרים אלה כי לעתים קרובות הם מועסקים פרמטרים שונים ושיטות אנליטיות. שלושה פרוטוקולים מתוארים כאן. הראשון משתמש בתמונות confocal הקרנת עוצמה מקסימלית כדי למדוד את השטח של קולטן האצטילכולין (AChR) -rich תחומים קרום postsynaptic בendplate והאזור של מכתים שלפוחית הסינפטית במסוף עצב presynaptic שמעליה. הפרוטוקול השני משווה את העוצמות היחסית של immunostaining לחלבונים סינפטיים בקרום postsynaptic. יחסי הציבור השלישיotocol משתמש העברת הקרינה תהודה אנרגיה (סריג) כדי לזהות שינויים באריזה של AChRs postsynaptic בendplate. הפרוטוקולים פותחו ושוכללו על סדרה של מחקרים. גורמים המשפיעים על האיכות ועקביות של תוצאות נדונים ונתונים נורמטיביים מסופקים NMJs בעכברים צעירים למבוגרים בריאים.
Introduction
הצומת העצבית-שהרירית (NMJ) היא סינפסה הממסר הקריטית שמתווכת תקשורת בין מערכת העצבים ושרירי שלד. היא נדרשת לכל התנועה רצונית. מיקרוסקופ פלואורסצנטי משמש כבר זמן רב כדי לחקור את ההשפעות של transgenes על העכבר NMJ 1-3 או להשוות את ההשפעות של גיל, תזונה, פעילות גופנית ומחלות על NMJs מכרסמים 4-11. מחקרים מסוג זה לימדו אותנו הרבה על הפיסיולוגיה ופתופיזיולוגיה של NMJ, אבל הפרמטרים המגוונים דיווחו (למשל, אזור AChR, אזור endplate, אורך היקפי, מדדי פיצול) לעתים קרובות לעשות את זה קשה להשוות את הממצאים של מחקרים אלה. יש ציפייה גוברת לחוקרים פרה-קליניים כדי להיות מסוגלים להפגין שחזור, במיוחד במחקרים עם מודלים של מכרסמים של מחלה 12. הפרוטוקולים שתוארו כאן היו מעודנים באמצעות סדרה של מחקרים שחקרה ch התפתחותית, פיסיולוגי וpathophysiologicalAnges לNMJ. מחקרים מסוג זה דורשים מדידה של השטח של התמחויות הסינפטי בendplate מנוע עכבר והצפיפות היחסית של אריזה של חלבונים סינפטיים בתוך התמחויות postsynaptic 13-15.
השירות של שיטות אלה בא לידי ביטוי במחקרים שנעשה לאחרונה במודל של עכברים של אנטי-מושק מיאסטניה גרביס. זריקות יומיות של IgG ממיאסטניה אנטי-מושק חיובית גרביס חולים לעכברים בוגרים שנגרמו להם להיות חלשים בתוך 2 שבועות 16. תמונות מרבי הקרנת Confocal סעיפים שריר שכפול שכותרתו עבור synaptophysin (בעצבי מסופים) וAChRs postsynaptic הציגו ירידה הדרגתית באזור של מכתים AChR כשינוי היסודי. חשוב לציין את קצב הירידה היה די כדי להסביר ירידות דומות במשרעת של פוטנציאלים סינפטיים, כישלון של העברת הסינפטית וחולשת השרירים 17,18. איכותי ממצאים דומים דווחו על ידי קבוצות מחקר אחרות10,19. באותן שיטות מדידת NMJ ומאז נהגו להעריך את ההשפעה של שלוש תרופות לטיפול במיאסטניה גרביס אנטי-המושק ב20,21 מודל העכבר הזה.
הזדקנות כרוך בישיבה יכולה להוביל לאובדן של קשרים עצביים-שרירית. הפרוטוקולים שתוארו כאן חשפו ירידת גיל הקשורים בתחום synaptophysin מסוף העצב בendplates המוטורי כמו עכברים להתקדם בגיל מבוגר. באותן השיטות גילו כי פעילות גופנית במידה רבה מרצון יכולה למנוע את הירידה בשטח מסוף עצב presynaptic 22, עולה בקנה אחד עם מחקרים קודמים על ידי קבוצות אחרות 4. אובדן של קשרים עצביים-שרירית מתרחש גם במודל עכבר SOD1G93A של טרשת לרוחב amyotrophic 9,23.
המחקרים שהוזכרו לעיל מראים כי מגוון רחב של מצבים בריאותיים עלול להוביל לירידה בשטח של שני התמחויות לפני או אחרי-סינפטי בNMJ. הדבר עלול לגרום כיף הסינפטי לקויction או עשוי לבשר אובדן הקשר העצבי-שהרירית מלא. שלושה פרוטוקולים מתוארים המאפשרים לכמת את האזור והצפיפות של התמחויות הסינפטי. המטרה של הפרוטוקול הראשון היא לספק מידה מעשית ושחזור של התחומים לפני והתמחויות פוסט-סינפטי ויישורם בNMJs היונקים, באמצעות מיקרוסקופ פלואורסצנטי. תמונות דו-ממדיות המרביות confocal ההקרנה וניתוח תמונה עם NIH ImageJ משמש כדי לזהות שינויים באזור של מכתים synaptophysin (שלפוחית סינפטית), AChRs postsynaptic ואזור חפיפה הסינפטי. פרמטרים Confocal הדמיה (רווח לקזז רמה) מותאמים לכל NMJ על מנת למקסם את המידע החזותי המשמש להבחין תחום התמחות הסינפטי. כישלון Neuromuscular יכול לנבוע גם משינויים בצפיפות של AChR postsynaptic ו / או חלבונים סינפטיים אחרים. הפרוטוקול השני ניתן ליישם כדי לזהות שינויים בצפיפות היחסית של חלבוני postsynaptic כזהכמושק, rapsyn, dystroglycan, קינאז Src phosphorylated וAChR phosphorylated 18,21.
במיאסטניה גרביס, צפיפות מופחתת של AChR בתוך הקרום postsynaptic היא הגורם המיידי לכישלון הסינפטי וחולשת שרירים. הפרוטוקול השלישי מתאר שיטת העברת הקרינה תהודה אנרגיה (סריג) כדי להעריך את השינויים בקרבת AChRs הסמוך בתוך ממברנות postsynaptic 14,15. שיטה זו מזהה העברת אנרגיה בין AChRs השכן שכותרתו עם הניאון-α-bungarotoxin (BGT). סריג מתרחש רק כאשר בדיקות תורם acceptor ניאון בנפרד פחות מ -10 ננומטר. זה יכול לגלות שינויים (תת-מיקרוסקופים) באטימות של אריזת AChR שעשוי להיות קשור ישירות להמשרעת של פוטנציאל הסינפטי.
שלושת פרוטוקולים אלה, מעודנים בעשור האחרון, מספקים אמצעי משלימים של יושרת NMJ באופן עקבי לשחזור. שימוש בפרוטוקולים סטנדרטייםפרמטרים nd צריכים לאפשר השוואה של ההשפעות של גנים והתערבויות סביבתיות על NMJ של היונקים.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
הערה: עיצוב, התנהגות ודיווח של ניסויים בבעלי חיים צריכה לקחת בחשבון את ההנחיות הנוכחיות 24. עבודה כזו חייבת להיות מאושרת מראש על ידי הרשות המקומית לרווחת בעלי החיים (במקרה שלנו לועדת האתיקה בבעלי חיים מאוניברסיטת סידני).
1. המתת חסד של החיה וDissection השריר
- העבר את העכבר מחדר ההמתנה לחדר נפרד שבו הוא מורדמים עם זריקת intraperitoneal של פתרון pentobarbitone (30 מ"ג / קילוגרם) תוך שימוש בשיטת טיפול עכבר מפורט על ידי שימיזו 25. מניחים את העכבר חזרה לכלוב שלה.
- ברגע שהנשימה של העכבר הפסיקה דקות יותר מ 1, לבדוק את רפלקס רגל-הנסיגה בעדינות על ידי צובט את הרגל, ורפלקס מצמוץ על ידי מלטפת קלה את הקרנית. רק כאשר תגובות הרפלקס נעדרות יכול העכבר להיות מוכן לנתיחה.
- התייעץ עם אטלס של האנטומיה של מכרסמים כגון Chiasson 26 ו / או לבקש את עזרתו של experienאנטומיה CED לפני שתנסה לנתיחה של השריר של עניין. בכל מקרה להסיר שיער מהעור המכסה באמצעות מכונת גילוח חשמלי קטנה לפני שפתחת את העור כדי לחשוף את השרירים.
הערה: לנתיחה תהיה שונה לכל שריר אנטומית-מובחן. - בעזרת מלקחיים בוטים לשחרר את השרירים שממעל הקרומים ורקמות הסובבים. לתפוס ולחתוך את הגיד הדיסטלי להפריד את השריר מההכנסה שלה.
- בעדינות להקניט ולגזור את השריר חופשי מרקמות זכות חזרה למקורו. בקצרה למקם את השרירים גזורים החדש לפתרון 0.1 M פוספט שנאגרו מלוח (PBS) או תמיסת רינגר לפני עיבוד נוסף.
2. הכנת השריר לCryosectioning
הערה: שימור מבני אופטימלי יכולה להיות מושגת על ידי זלוף בעלי החיים שלם כמפורט בעבר 27, או קיבעון טבילה (לשרירים קטנים) כמתואר בשלב אופציונאלי 2.1. עם זאת,4% קיבעון paraformaldehyde יכול לפגוע בצביעה לאחר מכן עם הרבה בדיקות נוגדנים ועם ניאון BGT. Glutaraldehyde במיוחד יש להימנע. אם שרירים לא להיות קבועים הם חייבים להיות מייד צמד קפוא (המשך 2.3).
- קיבעון טבילה אופציונאלי: Pin שריר השעווה בצלחת פטרי באורך מנוחה. מכסה את השריר עם 2% w / v paraformaldehyde (טרי מומס בPBS) לשעה 2 ב RT. לשטוף אותו עם 3 שינויים של PBS על 30 דקות (3 x 10 דקות) ולאחר מכן להחליף את PBS עם 30% w / v סוכרוז ב PBS ו דגירה O / N ב 4 ° C.
- להפוך את תבניות ("סירות") מראש על ידי קיפול 2 סנטימטרים X 1.5 חתיכות של נייר אלומיניום סנטימטר כפי שמוצג באיור 1. מניחים פיסת קרום nitrocellulose בקרקעית הסירה. בעדינות יוצק מטריצת cryostat הטבעה (שולחן חומרים) לסירה עד לעומק של 2 מ"מ, לטפל כדי למנוע בועות אוויר. הנח את השריר לתוך הסירה, יישורו עם קווי העט הכדוריים עלnitrocellulose. להוסיף מטריצת הטבעה יותר כדי לכסות את השריר (איור 1) לחלוטין.
- צינורות פוליפרופילן מראש תווית עם סמן בל יימחה. מניחים ירידה של מים בצינור אחד ומקרר את הצינור בחנקן נוזלי.
הערה: טיפת המים הקפואה שומרת על לחץ האדים ומונעת התייבשות במהלך ממושך -80 ° C אחסון - שימוש במגן פנים, כפפות מגן עבות וזוג גדול של מלקחיים בוטים, באופן חלקי להפחית כוס קטנה מתכת (קוטר 3 סנטימטר, 8 סנטימטר עמוק) המכילה 2 סנטימטר עומק של איזופנטאן לתוך מיכל של חנקן נוזלי למשך 30 שניות. הסר את הכוס ומניח אותו על גבי הספסל. בעזרת זוג קטן של מלקחיים בוטים למקם את התבנית המכילה את השריר והטבעת מטריצה לאיזופנטאן המצונן. תשמור על עצמך כדי למנוע ערבוב חנקן נוזלי עם איזופנטאן.
- לאפשר 2 דקות לבלוק להקפיא לחלוטין לפני שימוש מלקחיים בוטים להרים את הגוש הקפוא בחוץ ולאטום אותו ביחסי הציבור הנכוניםכותרת דואר וצינור מראש צונן (שלב 2.3).
- אחסן את הצינורות באופן זמני בחנקן הנוזלי לפני העברה ל-80 ° C. רשום את כל הדגימות בגיליון אלקטרוני של תוכן מקפיא.
3. Cryosectioning וקרינת מכתים לתמונות En פנים של NMJs
- לקלף עובש האלומיניום. בתוך 20 ° C קאמרי cryostat לצרף בלוק הקפוא לצ'אק cryostat כדי לחתוך 20 מיקרומטר cryosections מקביל לציר הארוך של סיבי השריר (איור 1). להרים את הסעיפים על poly-L- ליזין או שקופיות מיקרוסקופ ג'לטין מצופים.
- הערה: השמט שלב זה אם הרקמה קבועה לפני ההקפאה. אחרי 30 דקות המאפשרים לחלקים להתייבש על השקופיות, לתקן אותם על ידי הנחת ירידה של 2% בparaformaldehyde PBS על כל חלק במשך 15 דקות ב RT.
- לשטוף שקופיות 3 x 10 דקות ב PBS בצנצנת Coplin, ולאחר מכן לטבול את השקופיות בPBS המכיל 0.1 M גליצין למשך 30 דקות כדי לחסום קבוצות אלדהיד שייר.
- לשטוף שקופיות למשך 10 דקות ב PBS, ולאחר מכן לטבול במתנול (מקורר ל -20 מעלות צלזיוס) במשך 7 דקות. צעד permeabilization זה חלק משגרה של תיוג כפול עם ניאון BGT ואנטי-synaptophysin אבל זה עלול להשפיע לרעה immunostaining לכמה חלבונים אחרים.
- לשטוף שקופיות 2 x 10 דקות ב PBS ואז למקם את כל שקופית בתא humidified יציב ומפולס. מייד לכסות כל חלק עם 20 μl של פתרון חסימה (0.2% Triton X-100, 2% אלבומין בסרום שור (BSA) בPBS) במשך שעה 1 ב RT. סעיפים אסור להתייבש בכל שלב של תהליך immunostaining.
- לבצע את הדגירה העיקרית: לקיחת שקופית אחת בכל פעם להסיר בזהירות את פתרון החסימה העודף מעל כל סעיף ולהחליף אותו עם 20 μl של אנטי-synaptophysin ארנב (בדילול 1: 200 בפתרון החסימה).
- כולל שקופיות שלילית-שליטה שיהיה הודגרו עם חסימת פתרון היחידה. 'לא-יסודי שליטת נוגדן זה"הוא חיוני בכל ריצת immunostaining.
- טיפול שהנוגדן הראשוני נשאר במקום מעל כל קטע, לסגור את תא humidified ודגירה של 1-2 ימים על 4 מעלות צלזיוס.
- בדוק כל סעיף כדי לוודא שהנוגדן הראשוני נשאר במקום. השתמש פיפטה פסטר לשטוף בעדינות כל שקופית עם PBS ולמקם אותו בצנצנת Coplin. לשטוף את כל השקופיות 3 x 10 דקות ב PBS.
- לבצע דגירה משנית. לוקח שקופית אחת בכל פעם, להסיר בזהירות PBS העודף, ולהניח אותה בתא humidified ולכסות כל חלק עם 20 μl של תערובת המכילה IgG FITC מצומדות- החמור נגד הארנב וBGT המוצמד לtetramethyl rhodamine או אחר fluorophor אדום (TRITC- / RedBGT; 5 גר '/ מיליליטר) בדילול מלא בפתרון חסימה. לדגור על RT עבור שעה 2.
- לשטוף שקופיות 3 x 10 דקות ב PBS בצנצנות Coplin.
- לוקח שקופית אחת בכל פעם, להסיר בזהירות PBS העודף והר עם coverslip באמצעות נפח מינימאלי של, glyceroמבוסס l, הרכבה התנגדות בינונית-לדעוך. לאטום את הקצוות של coverslips עם לכה מסמר ברור. לאפשר לו להתייבש קשה.
- אחסן את השקופיות בחושך ב 4 מעלות צלזיוס עד שבוע אחד, או ב -20 ºC לתקופות ארוכות יותר אחסון (עד מספר חודשים).
4. משוחדת דגימה וEn Face הדמיה של הרכב endplates
- עיוור שקופיות על ידי תיוג כל שקופית עם מספר קוד אקראי שנשאר ידוע רק לחוקר שני (שלא היה מעורב בניתוח). כתוצאה מכך המפעיל נשאר עיוור לקבוצות טיפול עד לכמת את הפרמטרים NMJ הוא מלאה עבור כל הדגימות.
- מניחים את השקף על הבמה מיקרוסקופ ולצפות בו תחת תאורת שדה רחבה עם מערכת סינון TRITC (שמן 1.3 אובייקטיבי NA 63X). העבר בהדרגה (שדה על ידי שדה) משמאל לימין ובחזרה עד endplate מופיעה בשדה (איור 2 א).
קריטריון דגימה:: הערה בכל מבנה הצבעוני AChR שהוא יחסיתשטוח ופונה למטרה (כלומר, משתרע <15 מ 'בz-הממד) נחשב endplate והוא צילם לניתוח (סהרונים של מכתים AChR מייצגים חתכים דרך endplates ולכן לא נכללו). - עם סט חריר confocal 1.0 יחידת Airy וכוח הלייזר נמוך לייעל את הרווח לקזז רמות לTRITC / אדום-BGT (לייזר 532 ננומטר) בendplate שהיא להיות צילמו. FITC לייעל הבא / הקרינה synaptophysin באמצעות לייזר 488 ננומטר. לאסוף Z- מחסנית של endplate עם מרווח של 0.7 מיקרומטר בין כל פרוסה אופטית. שמור את התמונות עם שם קובץ שכולל את התאריך של פגישת ההדמיה, שם הקוד של השקופית ומספר endplate.
הערה: הסריקות באמצעות ננומטר 488 ו 532 ננומטר לייזרים (FITC וTRITC) יש לאסוף ברצף (לא בו זמנית), כדי למנוע זיהום של ערוץ FITC על ידי הקרינה מfluorophore האדום ולהיפך (לדמם דרך). - חזור על הדגימהnd הדמיה של צעדים 4.2-4.3 עד 20 endplates נאספים מהשקופיות / המדגם.
- לשנות לשקופית מקודדת הבאה וחזור 4.2-4.4. חזור על פעולה זו עבור כל אחד מהשקופיות מקודדות.
- לאסוף כמה תמונות של endplates משקופית השליטה (לא-יסודי שליטת נוגדן) באמצעות confocal הגדרות שנמצאו אופטימלי לשקופיות ניסוי (ערוץ הקרינה FITC אמור להופיע כהה).
- בסופו של העברת הפעלת confocal קבצי תמונה למחשב אחר ולגבות את הקבצים המקוריים בכונן או בשרת חיצוני.
5. מדידת השטח של התמחויות Synaptic בתמונות En Face
- השתמש בתוכנה חופשית NIH ImageJ (http://imagej.nih.gov/ij/) להכין תמונות הקרנה מקסימלי (MIP) מכל Z- מחסנית. לשמור אותם כקבצי TIFF (איור 2 א & B). שמות קבצים צריכים לכלול את תאריך תמונת מושב, קוד לדוגמא, מספר endplate וערוץ ניאון (למשל, 060414_5723_7_FITC.tiff).
- פתח את תמונת Z- ההקרנה בImageJ. בחר את ערוץ תמונת קולטן האצטילכולין (איור 3 א) ובחר: תמונה> סוג> 8 סיביות להמיר תמונת RGB 24-bit צבע לשלוש תמונות בגווני אפור 8 סיביות על המסך.
- שימוש בכלי מצולע ImageJ לצייר קווי המתאר גסים סביב endplate עניין בredBGT מוכתמת ערוץ (ACHR) כך שיכלול את כל האזורים לכאורה הצבעוניים של endplate אדם המסוימת, תוך נטרול כל כתמים שאינו נובעים מendplate של עניין ( 3C איור).
- החל סף עוצמה מינימאלי לתמונה על ידי בחירה: תמונה> התאם> סף (איור 3E וצילומי מסך ImageJ קשור).
- התאם את רמת הסף כדי לבודד את החלקים הצבעוניים AChR תוך הדרת אות רקע שמסביב כמו תת-סף (3E איור). פתח סיבוב שניOW עם התמונה המקורית (רציף הטון) מייד ליד החלון להשוואה, כדי להקל על ההחלטה על ערך הסף. רשום את ערך הסף לשימוש מאוחר יותר בcolocalization ניתוחים.
- שמירה על המתווה המצולע סביב endplate לבחור: הניתוח> לנתח חלקיקים. בתפריט המוקפץ לציין את הטווח של גדלים כ: 50 עד אינסוף פיקסלים (זה מבטל חפצים זעירים הנובעים מרעש חשמלי במכפיל).
- לנתח חלקיקי הפקודה יוצרת חלון עם רשימה של אזורים על-סף בדידים וערכי עוצמת הקרינה שלהם ממוספרים כפי שהם מופיעים בתמונה בינארי (איור 3G ומסך ImageJ קשור). העתק את הנתונים לגיליון אלקטרוני שכותרתו.
- למדוד את שטח endplate סה"כ (האזור בתוך המצולע) על ידי בחירה: ניתוח> מדוד. זה מניב הכולל אזור endplate. העתק והדבק את הנתונים לאזורי AChR ועוצמות לגיליון אלקטרוני כדי לוודא לתייג עמודות כראוי, שורות ישמשו לendplates פרט לשקופיות ספציפיות.
- עובר לערוץ הקרינה אנטי-synaptophysin וחזור על צעדים 5.1-5.5, אבל לערוץ FITC (איור 3 ב ', ד' ו-ו '). המטרה היא להתאים את הסף, כך שהוא יוצר תמונה בינארי ש, ככל האפשר, תואם את הגבולות של מכתים כפי שהיא נתפסת על ידי העין. רשום את ערך הסף.
- למדוד את השטח של חפיפה על ידי יישום הצעדים הבאים: פתח את הקובץ המקורי מכיל שתי תמונות הערוץ ולפצל אותה לשתי תמונות נפרדות על ידי בחירה: תמונה> סטאקס> מחסנית לתמונות.
- שימוש בתוסף colocalization (הוריד והתקין מדף אינטרנט ImageJ) בחר: Pluggin> colocalization וקלט ערכי הסף נרשמו בעבר לערוצי AChR ועצב לb שאילתא הערוץ המתאיםשור. זו תניב תמונת חפיפה בפיקסלים לבנים (איור 3H וצילומי מסך ImageJ קשור).
- להמיר את תמונת החפיפה החדשה שנוצרה לפורמט בגווני אפור ולהחיל סף לערך המקסימאלי. הסף המקסימאלי רק יבחר פיקסלים הלבנים, מתאים לאזור החפיפה של שני הערוצים הקודמים. שיא בגיליון האלקטרוני וכתוצאה מכך ערך שטח של 'colocalization', המייצג את אזור חפיפה בפיקסלים.
- הכן גיליון אלקטרוני של אמצעי מדגם נתונים, לחשב ולתכנן סטיות תקן וסטיות התקן כהיסטוגרמות או scatterplots 20,22. שים לב שהערך של n בדרך כלל מייצג את מספר העכברים לכל קבוצת מדגם למטרות סטטיסטיות.
- endplate עלילת אזורי AChR כscatterplots או היסטוגרמות תדירות כדי לקבוע אם הנתונים מתפלגים נורמלי לפני בדיקה סטטיסטית (איור 6).
6. מכתים יחסיתעוצמות לעומת שימוש בסעיפים אופטיים רוחביים
הערה: לתהליך זה פרוטוקול יחד את כל דגימות השריר ותמונה במושב confocal יחיד. בתכנון ניסוי מאפשר עד 30 דקות זמן הדמיה לדגימה שריר.
- לחתוך 15 מיקרומטר cryosections הרוחבי לציר של סיבי השריר הארוך ולאסוף על גבי שקופיות כפי שמתואר בשלב 3.1.
- לבצע צביעת הקרינה כמתואר בשלבים 3.2-3.13.
- קוד שקופיות המוכתמות כך שההדמיה והניתוח מתבצעים עם עיוור מפעיל לקבוצת טיפול, כפי שתוארו בשלב 4.1.
- באמצעות מטרת 40X הקרינה (NA 0.75) בקצרה תסקור חלק מכל שקופית כדי לקבוע רווח בודד וקיזוז הגדרת רמה לAChR שתהיה מתאים לכל endplates בכל שקופיות המדגם. Endplate הבהירה אז צריכה להיות ממש מתחת 256 אפורה בקנה המידה. אופטימיזציה זה צריך להיעשות בנפרד עבור fluorescenc השניערוץ E (שנאסף ברציפות). רשום את הרווח הקבוע ולקזז את הגדרות רמה ואינו משנה אותם לאורך כל פגישת ההדמיה.
- לאסוף תמונות של שקופיות הקרינה סטנדרטית (לדוגמא, חרוזי ניאון הלא הלבנה), תוך שימוש באותם הפרמטרים, בתחילת וסוף פגישת confocal כדי לאתר את כל תנודות אפשריות בעוצמת לייזר.
- השתמש בערוץ AChR כדי לסרוק את השקופיות בהדרגה לאתר endplates.
- להתמקד כדי למצוא את מטוס הסעיף האופטי היחיד בכל תחום מיקרוסקופ המכיל את המספר הגדול ביותר של endplates הצבעוני AChR.
- סריקת סעיף אופטי יחיד זה פעמיים ולשמור את התמונה בממוצע (איור 4G).
- שמירה על אותו מתג מוקדי מטוס לערוץ הקרינה השני (חלבון של עניין) ולאסוף את התמונה כבשלב 6.8. שמור את קובץ התמונה, ובכלל זה בשם הקובץ: מועד פגישת הדמיה, קוד לדוגמא, מספר תמונה וסמל כדי לציין את ערוץ הניאון.
- הזז את הבמה לשדה הבא שיכלול את endplates אחד או יותר וחזור על שלב 6.8-6.9. חזור על פעולה זו עד סך של 60 endplates הם צילמו.
- בסופו של העברת פגישת הדמיה את כל הקבצים למחשב אחר ולגבות אותם.
- לפתוח כל קובץ תמונה מקורי ותוך כדי צפייה בערוץ AChR, בחר: תמונה> סטאקס> מחסנית לתמונות, לפיצול ערוצים.
- בחר: תמונה> סוג> 8bit להמיר לפורמט בגווני אפור 8 סיביות על המסך. לעשות את זה בשני ערוצי הקרינה.
- בחר: תמונה> תמונות סטאקס> למחסנית. פתח ערימה חדשה משתי תמונות של 8 סיביות הופרדו בעבר. אפשר לאחר מכן לעבור בנוחות בין שני ערוצי הקרינה בתוך החלון אחד.
- השתמש בכלי המצולע לצייר linדואר בחוזקה סביב הגבול של הכתמת AChR (איור 4I).
- בחר: ניתוח> למדוד למדוד את עוצמת פיקסל הממוצעת לAChR בתוך השטח המגודר (שים לב לחשיבותו של ציור הקו בחוזקה). העתק ערך זה לגיליון אלקטרוני שכותרתו.
- שמירה על אותו המתווה המצולע (כדי להגדיר את האזור כדי להימדד), עובר לערוץ השני הניאון (למשל, איור 4 ב ', ד', ו ') ובחר: ניתוח> מדד. זו תניב את עוצמת כתמים הממוצעת לחלבון של עניין בתחום הסינפטי שהוגדר על ידי צביעת AChR.
- בחר אזור מהכתמת endplate גלויה ולאחר מכן בחר: ניתוח> למדוד למדוד את עוצמת הקרינה רקע הממוצעת. חזור על פעולה זו עבור ערוץ הקרינה / s האחרות ולהעתיק את ערכי רקע לגיליון האלקטרוני של ערכי הקרינה.
- תתמערכת ערכי רקע הממוצע מערכי endplate להשיג עוצמות המתוקנות לAChR והחלבון של עניין בכל endplate.
- מחלקים את ערכי עצמת endplate המתוקנות לחלבון של עניין על ידי עוצמת הקרינה BGT תיקנה להניב יחסי עוצמת הקרינה 14,21
7. השוואת צפיפות Postsynaptic ממברנה AChR שימוש סריג
הערה: פרוטוקול זה מעריך בקרום postsynaptic המידה שבה הם ארוזים באופן הדוק AChRs (<10 ננומטר מרווח). שילוב תורם fluorophore acceptor המדויק הוא קריטי לassay סריג זה. מקבלים שמות ופרטים של fluorophores בטבלת חומרים. התכונות ספקטרליות שלהם, ביחס לסריג, הם דנו במאמרים הקודמים שלנו 14,15.
- הכן cryosections הרוחבי קבוע כמפורט בסעיף 6.1. כל קבוצות המדגם חייבים להיות מעובד ביחד ותמונהד באותה ההפעלה confocal.
- לערבב ביסודיות אדום BGT (תורם סריג) 2.5 גר '/ מיליליטר עם 10 גר' / מיליליטר כה אדומה-BGT (סריג acceptor) עם פתרון חסימה בצינור פלסטיק קטן על ידי pipetting למעלה ולמטה 12 פעמים. זה 1: תערובת טוחנת 4 ממקסם את היעילות של 14 סריג.
- מניחים כל שקופית בתא humidified, לכסות בזהירות כל חלק עם ירידה (μl 12) של התערובת מעל ודגירה של 1.5 שעות ב RT.
- סעיפי שליטה: לכסות מספר קטן של חלקים עם 2.5 גר '/ מיליליטר אדום-BGT (תורם בלבד; בקרות C1 שכותרתו), וגם כמה קטעים עם 10 גר' / כה-אדום-BGT מיליליטר (acceptor בלבד; בקרות C2 שכותרתו). דגירה בקרות אלה כבשלב 7.3.
- לשטוף שקופיות 3 x 10 דקות ב PBS ו הר הבמבוסס על גליצרול, הרכבה בינונית (ראה שלב 3.12) העמידה לדהייה.
- לבצע דגימה של endplates כמו בשלב 6.7. הקרינה מתורם acceptor צריכה להיות מושלם שיתוף מקומי-בendplates בשל אקראי המחייב של מולקולות ניאון-BGT.
- תמונות בקרה: שימוש במטרה 40X ורווח redBGT נמוך לייזר לייעל כוח ולקזז את הגדרות רמה לendplates מC1 שקופיות שליטה. לייעל רווח מרחיק redBGT ולקזז רמות לendplates מC2 שקופיות שליטה. לאשר את היעדר לדמם דרך הקרינה.
- מבלי לשנות את כוח הלייזר, להרוויח או הגדרות ברמת קיזוז לעבור לשקופיות הניסיוניות ולאסוף תמונות (לפני photobleach) לשני ערוצי הקרינה.
- סלקטיבי photobleach מרחיק האדומה-BGT על חלק של endplate אחת על ידי התקרבות באזור הסריקה ולאחר מכן סריקת 10 פעמים עם לייזר 633 ננומטר ב 100% כוח. הקרינה באזור שנסרק צריכה להיות עמומה.
- איפוס כוח הלייזר וזום ולאסוף תמונות לאחר אקונומיקה על שני ערוצי הניאון באמצעות הגדרות confocal הוקמו על 7.7.
- לחשב את יעילות סריג (E) מאחוז העלייה בתורם הקרינה (אדום-BGT) הבא photobleach של acceptor (מרחיק האדום-BGT) על פי הנוסחא * הבאה:

* לכל המצבים שבם הקרינה של התורם מגבירה לאחר photobleaching acceptor.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
מדידה של שטח Synaptic בNMJ
כל הערכה של האזור מסתמכת על הציור של גבול כדי להגדיר את היקף התמחויות הסינפטי. בשרירים צעירים מבוגרים בריאים תמונות NMJ צריכים להציג גבולות מוגדרים היטב עבור שני AChR וצביעת synaptophysin (איור 2 א 'וב'). עוצמת הקרינה לשני AChR וsynaptophysin עולה בחדות בגבול בין חלק פרי הסינפטי והסינפטי של endplate מנוע (איור 5 א 'ו-ב' '). לתמונות כאלה סף מינימאלי (רק מעל הקרינה רקע extrasynaptic) יהיה קושי לבודד את אזור AChR העשיר או synaptophysin העשיר של endplate (קווים מקווקווים אופקיים ב( איור 5 א 'ו-ב' ')). בעכברים בגיל ובחלק מכתמי endplate מצבי מחלה לAChR עשוי להיות פחות אינטנסיבי, קצות אשכול AChR עשויים להופיע מטושטשים ומחדש עשוי להיות רמות גבוהות יותר של autofluorescence extrasynaptic (איור 5D; 17,22). צביעת הקרינה עם גבולות ברורים יכולה להציג את השגיאה להערכות של אזור הסינפטי. בכל מצבי המטרה היא לבחור סף שמניב תמונה בינארי דומה בצורה ובגודל לAChR- או אזורי synaptophysin העשיר כפי שהם מופיעים לעין בתמונה המקורית, רציף-הטון. ביצוע הניתוח עיוור לקבוצת הטיפול צריך להפחית את הסיכון של הטיה סובייקטיבית בצעד thresholding (שלב 4.1). לפעמים תמונות endplate קלושות או מטושטשות יכולות להיגרם כתוצאה מעיבוד תת-אופטימלי. איור 2C ו- D מראה דוגמא של תמונה באיכות נמוכה endplate מעכבר ישן 2 חודשים בריאים. קצוות מטושטשים וצביעת synaptophysin קלושה אולי נובעים במקרה זה מהפשרה וrefreezing חלקיות של השריר לפני cryosectioning. חלקים מסוימים של שריר צעיר ובריא (ביקורת חיובית) צריכים להיות מחולקיםnd מעובד במקביל עם דגימות ניסוי על מנת להבטיח כי כל ירידת הערך בא לידי ביטוי בתמונות NMJ הוא לא בגלל בעיות עם immunostaining. קבוצות של תמונות נפגעו על ידי עיבוד תת-אופטימלי צריכים להיות מחוץ ניתוחים.
לen Z- מחסנית תמונות פנים, 15-20 endplates הוא גודל מדגם סביר להערכת אזורים הסינפטי. גיוון רחב בצורות וגדלים של NMJs נמצא בתוך כל שריר נתון. Scatterplots לחשוף טווח ניכר באזור AChR העשיר בין endplates של השריר הקדמי tibialis של כל עכבר בודד (איור 6 א). עם זאת, השטח הממוצע AChR (המבוסס על 15-20 en תמונות הפנים endplate) היה דומה בין שבעה עכברי מדגם (~ 200 מ '2; איור 6 א). האזור של מכתים synaptophysin endplate גם שונה בצורה משמעותית בין endplates משריר נתון. שוב, באמצעות גודל מדגם של 15-20 endplates האזור הממוצע של synaptophysin endplate היה סימיlar בקרב העכברים 7 למד (~ 170m 2; איור 6). היסטוגרמות תדר של נתונים ויקוו חשפה הפצות בערך נורמליות לאזור של AChR endplate וsynaptophysin (איור 6 א 'ו-ב' '). עם זאת התפלגות נורמלית של אזורים הסינפטי לא ניתן להניח במצבי מחלה כגון מיאסטניה גרביס 16,20. זה עשוי להשפיע על הבחירה של המבחן הסטטיסטי.
שולחן אני מפרט תחומי התמחות של לפני ואחרי סינפטי לNMJs לC57Bl / 6J נקבה בריא בת 2 חודשים (למבוגרים צעירים) ממחקרים קודמים. תחומי ההתמחות הן לפני והפוסט-סינפטי ירדו עם ישיבת הזדקנות 22. אזור AChR גם מופחת במידה ניכרת בעכברים שהוזרקו עם IgG ממיאסטניה גרביס אנטי-מושק חולים 17,21. עכברי Myasthenic טופלו בתרופה מעכבת cholinesterase, פירידוסטיגמין, מוצגים הפחתה משמעותית נוספת בendplate אזור AChR 20 </ Sup>.
עוצמה היחסית של תיוג endplate הקרינה
העצמה היחסית של תיוג immunofluorescence יכולה לגלות שינויים בצפיפות של עניין חלבון-of-סינפטי עם גיל, גנוטיפ ו / או מצב מחלה. הקרינה AChR (אדומה-BGT או רחוק-אדומה-BGT) משמשת ראשונה להגדיר את מיקומו של NMJ. הבהירות של הקרינה בתחום AChR העשיר 8 סיביות לאחר מכן נעשה שימוש כדי להעריך שינויים בריכוז של החלבון של עניין, ביחס לשליטה בעלי חיים. בסעיפים רוחביים AChRs endplate מופיע בדרך כלל כצורה חצי סהר, אבל צורה זו היא לעתים קרובות לא סדירה (איור 4 א, C, E, H). הקרינה רקע בעצימות נמוכות בדרך כלל מגלה אם תיקון של מכתים AChR מייצג endplate אחת, או שני endplates הנפרד הממוקם בסיבי שריר סמוכים. חלבונים רבים הסינפטי (כגון rapsyn, מושק וSRC) הםcolocalized עם AChR בendplate (איור 4 א - ד). גם מכתים immunofluorescence עם נוגדנים ספציפיים phospho יכול לשמש כדי להשוות את ההשפעה של התערבויות ניסיוניות על מצב זרחון של חלבונים בממברנה פוסט-סינפטי בפרט 21.
האמינות והשחזור של מדידת עוצמת הקרינה תלויה במידה רבה על שלמות השריר הקפוא והאיכות של immunostaining. צריכים להיות גזורים שרירים וsnap-קפוא באופן מיידי או (בתוך דקות של מותו של בעל החיים)-קבוע paraformaldehyde כדי למנוע שינויים ניווניים בNMJ. Immunostaining תלוי מאוד על האיכות של חומרים כימיים ואופטימיזציה של הפרוטוקול מכתים לנוגדנים ספציפיים. לכל קבוצה חדשה של ניסויי immunostaining טייס נוגדן ראשוני נחוצים. cryosections טרי חתוך של שרירים צעירים ובריאים מודגרת עם דילולים פי 2 סידוריים של הנוגדן הראשוני.נוגדנים משני אמין ידועים משמש והתוצאות השוואה. אם החלבון של עניין ידוע להיות מוגבל לNMJ אז ריכוז הנוגדן הטוב ביותר הוא זו שמניבה את היחס הגבוה ביותר של עוצמת הקרינה NMJ ביחס לזה שנמצא בחלקי extrasynaptic של השריר (הקרינה רקע). עוצמת extrasynaptic (כנראה שאינה ספציפית) הקרינה לא צריכה בדרך כלל תעלה על 15% מעוצמת הקרינה endplate. באופן דומה "אין שליטת נוגדן ראשונית חתכים (מודגרות רק עם נוגדנים משני) אמורה להופיע כהה, המאשר כי הנוגדנים משני לא להיקשר שאינו ספציפי. האיכות של קבוצות שונות של נוגדנים (polyclonal) משניים יכולים להשתנות קבוצות נוגדנים משני במידה ניכרת כל כך חלופיות יש להשוות לפני הקמת פרוטוקול סטנדרטי. המבחן האידיאלי עבור הסגוליות של immunofluorescence כרוך השוואה ישירה של חלקים מעכברי wild-type וחתכים שלילית-שליטה מt העכבריםכובע חסר (נוקאאוט הגן) החלבון של הריבית. אזכור הבא מתאר quantitation של מכתים הקרינה endplate למושק, rapsyn, dystroglycan, src וAChR 13,14,18,21.
גודל מדגם קטן להציג את השגיאה להערכות של עוצמת הקרינה היחסית. endplates הבודד השתנה באופן ניכר בעוצמת הקרינה. יש להניח ששונות זו בין endplates בתוך שריר נתון משקפת את הגיוון של הבדלי מבנה וסיכוי NMJ בסעיף האופטי שנדגמו. עם זאת, הגדלת מספר endplates נדגמו תוצאות בהערכה יציבה יותר של עוצמת הקרינה ממוצעת (איור 7). כדי להעריך את עוצמת הקרינה הממוצעת endplate 40-60 endplates מכל מדגם שריר צריך להיות בממוצע.
AChR-AChR סריג
כל AChR הוא pentamer עם שני אתרי קישור לBGT (הממוקמים אחד על כל אלפא-מקטע). עקידתאדום-BGT ומרחיקה אדומה-BGT לשני אתרים אלה יניבו הפרדת acceptor התורם של כ 28-30 9nm. כך ניתן לאתר יעילות נמוכה סריג עוד לפני AChRs להרכיב לאשכולות 14. עם זאת, היעילות של סריג בendplates העכברי בערך הוכפלה לאחר הלידה, עולה בקנה אחד עם השילוב יעיל יותר של AChRs לסריג קרום postsynaptic צפוף 14. שימוש באדום-BGT ומרחיקה אדומה-BGT כתורם סריג וacceptor (בהתאמה), endplates של עכברים ישנים 1-2 חודש מיוצר יעילות סריג ממוצעת נעה 20-37% (טבלה 2). יעילות סריג של 20% או יותר חשבו לייצג אריזה הדוקה של 14 AChRs. יעילות endplate סריג הייתה מעט מופחתת הבאה denervation 14, ומופחתת במידה ניכרת אחרי העכברים הוזרקו IgG ממיאסטניה אנטי-מושק חיובית גרביס חולים 18. אלה הם תנאים שבהם AChRs הבודד עמוס פחות בחוזקה לpostsyפיגום קרום naptic על ידי המערכת המושק / rapsyn postsynaptic 31.
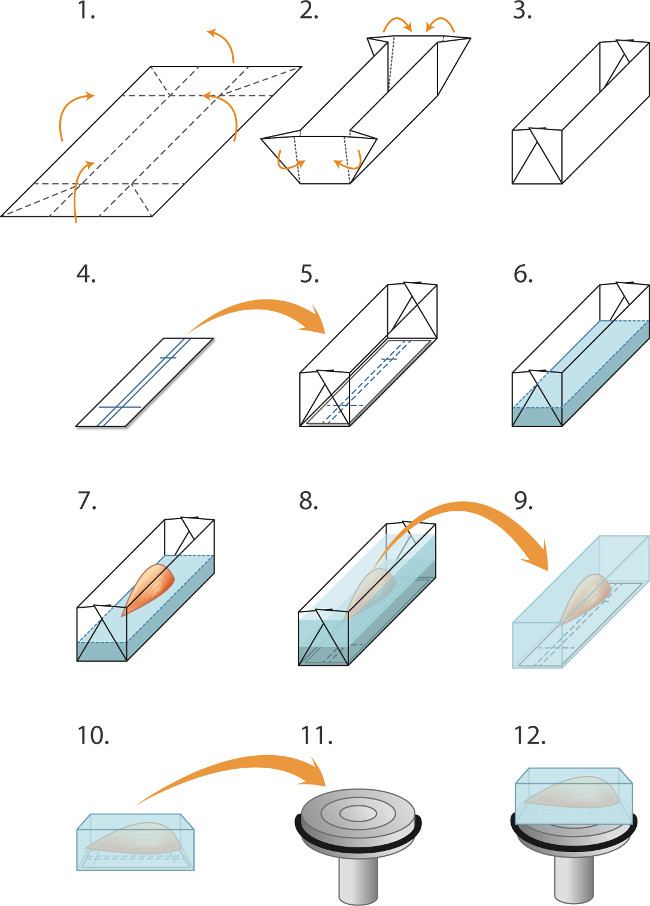
איור 1. הטבעה ושרירים הקפאה לcryosectioning (1-5) הכנת תבניות ("סירות") לפני ההקפאה אצווה של שרירים:. (1) רדיד אלומיניום הוא לחתוך למלבנים (2.0 x 3.0 סנטימטר) ו- (2). מקופל כנגדו כתב אישום ליצירת עובש / סירה (3). (4) מלבנים של נייר immunoblotting nitrocellulose נחתכים כדי להתאים בעובש ועט כדורי משמש לשלוט קווים לorientating השריר. (5) מלבן nitrocellulose ממוקם בעובש. (6-8) Embedding והקפאת השרירים: (6) cryostat נוזל מטריצת הטבעה הוא שפך בעדינות לתוך התבנית (על גבי nitrocellulose) עד לעומק של 2 מ"מ. (7) מלקחיים גלם המשמשים להורדת השריר, גידו על ידי לתוך מטריצת ההטבעה, בתיאום עם nitrocelluלאבד את פסקי דין, (8) מטריצת הטבעה נוספות היא שפך בעדינות לתוך התבנית כדי לכסות את השריר, לטפל כדי למנוע יצירת בועות. השריר המשובץ לאחר מכן הצמיד את קפוא, חתום בצינורות ומאוחסנים ב -80 ° C כפי שמתוארת בטקסט. (9-12) הכנה לcryosectioning: (9) המלקחיים פיין משמשים לקלף עובש האלומיניום והגוש הקפוא ממוקם אז ב-20 ° C תא cryostat, (10) הסימונים בnitrocellulose משמשים כדי ליישר במקביל לשרירי הפנים של צ'אק (11) לחתך אורכי (12). ירידה של מדיום ההטבעה-cryo נוזל משמשת לצרף את הגוש לצ'אק המצונן. מלקחיים בוטים משמשים כדי לתפעל את בלוק השריר הקפוא. להשיג חלקים רוחביים, הבלוק הוא רכוב במקום, כך שהשרירים הוא בניצב למשטח של צ'אק (לא מוצג).

Figure 2. תמונות פנים En של NMJs מהשריר הקדמי tibialis של עכברי C57Bl6J 2 חודשים ישנים נשיים. השריר הטרי היה הצמד קפוא והחלקים קבועים בשקופית כמתואר בשלב 3.2. Synaptophysin תמונות הקרנת עוצמה מקסימלית של Z- ערימות התקבלו כמתואר בפרוטוקול הראשון. מכתים () האדומה-BGT מגלה endplate מנוע בודדת מורכבת משני סטים של מרזבי AChR עשירים עיקריים postjunctional (en פנים נוף). (ב) צביעה עם נוגדנים משני FITC מצומדות- חושפת את מסוף עצב presynaptic, כובש את המרזבים הסינפטי העיקריים. חלק מהאקסון מראש המסוף הוא גם נראה לעין בחלקו העליון של הלוח. (C & D) דוגמא של תמונה באיכות ירודה מNMJ עכבר צעיר ובריא. הצביעה היא חלשה ואת הגבולות של ההתמחויות של לפני ואחרי הסינפטי הם מטושטשים. זה יוחס לליקויים בtהוא העיבוד של הרקמות ו / או הזמן לקוי אפשר לדגירת הנוגדן הראשונית. בר סולם בפנל D מייצג 10 מיקרומטר. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

איור 3. שלבים בעיבוד של תמונות en NMJ פנים (כפי שמתואר בפרוטוקול 1). צביעת AChR לאחר המרה לתמונה בגוונים אפורים 8 סיביות ושימוש בכלי המצולע להתוות endplate (A & B) תמונות MIP הצליל מתמשכות מקוריות להראות הקרינה אדומה-BGT חושפת AChR וimmunofluorescence הירוק לsynaptophysin בהתאמה. (C) (קו צהוב דק). קו הגבול (D) endplate יעבור לתמונת synaptophysin הירוקה. (E) Application של פקודת סף עוצמה מינימאלית כדי ליצור תמונה בינארי שמבודדת הקרינה suprathreshold האדומה-BGT (AChR). הרצף של צילומי מסך של ממשק משתמש ImageJ מוצג בצד השמאל של התמונות. תמונת synaptophysin ינארי לאחר היישום של סף נפרד (F). זיהוי (G) של תחומים AChR העשירים suprathreshold הבדיד בתוך endplate על ידי יישום של פיקוד החלקיקים לנתח לתמונה האדומה-BGT בינארי. ImageJ המקביל (משמאל של פנל (G) מציג את נתוני הקלט נדרש. הגודל המינימאלי של אזורי פיקסל suprathreshold הנדרשים יש להזין. (H) זיהוי של אזורי חפיפה של synaptophysin בינארי ותמונות AChR. החפיפה מיוצגת על ידי פיקסלים לבנים . ImageJ צילומי מסך ממשק משתמש (להלן H פנל) להראות את הצעדים כדי להגיע לתמונת החפיפה בינארי. ערכי סף עוצמה המינימליים שנבחרו עבור כל c הקרינהhannel יש להזין בתוך החלון "colocalization '. בר סולם מייצג 10 מיקרומטר. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

איור 4. דוגמאות לסעיפים אופטיים רוחביים משמשים להשוואת עוצמות ניאון היחסית (פרוטוקול 2). השריר היה צמד קפוא והחלקים קבועים בשקופית כמתואר בשלב 3.2. (A & B) endplate אחת פעמיים שכותרתו עם הרבה -אדומות-BGT (AChR; מוצגת כאן בpseudocolor הכחול) וFITC-אנטי-rapsyn ממחישות את שיתוף הלוקליזציה של שני חלבוני אינטראקציה אלה בקרום postsynaptic (C & D) שני endplates על סיבי שריר סמוכים תצוגת שיתוף מקומי AC. משאבי אנוש ומושק. (E & F) endplate כפולה שכותרתו עבור AChR ו-dystroglycan (-DG). -DG משתרע תקין מסביב סיבי שריר אבל הוא מועשר בendplate (סרגל קנה מידה בF, לAF לוחות: 25 מיקרומטר) (G - I) בידוד endplate למדידת עוצמת (G) שדה מיקרוסקופ טיפוסי,.. המכיל שלושה endplates המוכתם רחוק-אדומה-BGT (סרגל קנה מידה בG פנל: 40 מיקרומטר (H) תמונה מוגדלת של endplate בארגזים (I) אותו endplate מומרת לתמונה בגווני אפור 8 סיביות והתוותה באמצעות המצולע.. . כלי של ImageJ (קו צהוב דק) עוצמת הקרינה ממוצעת נמדדת בתוך קו זה גבול. (סרגל קנה מידה עבור H & I: 10 מיקרומטר) אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

השפעת איור 5. של איכות תמונה בהערכת שטח הסינפטי. (A & B) באיכות גבוהה en תמונות פנים של NMJ בריא מעכבר ישן 2 חודשים נצפה באדומה-BGT וערוצי הקרינה אנטי-synaptophysin. ('& B ') עוצמת הקרינה פרופילים מתאים לקו המשורטט בendplate לA ו- B בהתאמה. הקו האדום מקווקו האופקי מציין את הסף המינימאלי המשמש ליצירת התמונה בינארי. Endplate (C & D) מעכבר קשישים. צביעת synaptophysin endplate בדרך כלל היא פחות אינטנסיבית. (ג '& D ") פרופילי עוצמה מראים רמה גבוהה של extrasynaptic תנודות (בסיס) הקרינה בsynaptophysin הערוץ (FITC) (בחזרהקרקע) המשפיעה על קביעת סף מתאים. חלק גדול מזה הוא autofluorescence רקמות ספקטרום רחב. ברים סולם מייצגים 10 מיקרומטר. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

השתנות איור 6. אזורים הסינפטי בין NMJs בתוך שריר ובין עכברים. () Scatterplots להציג את אזור AChR העשיר הכולל של endplates מהשריר הקדמי tibialis של שבע / 6J נאיבי ישן 2 חודשים נקבת C57Bl מתקבל על ידי מחבר NT. כל סמל מייצג endplate. כל עמודה מייצגת הממוצעת ± SD לendplates נדגמו מעכבר אחד. Scatterplots (B) מראה את אזור synaptophysin העשיר לאותו endplates. (א ') (ב ') חלוקת תדר לאזור synaptophysin העשיר של endplates (נתונים ויקוו). אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.

איור 7. השפעת גודל מדגם על אומדנים של עוצמת הקרינה endplate. חלקים אופטיים רוחביים שמשו למדידת עוצמת הקרינה (ביחידות שרירותיות) בשעה 40 60 endplates מהשריר הקדמי tibialis של עכבר ישן 2 חודשים בריאים. ממוצעים מצטברים הם זממו לעומת מספר endplates נכלל בממוצע. עוצמת הקרינה endplate () האדומה-BGT מתקבלת על ידי AV מחבר. (B) Anti-synaptophysin עוצמת immunofluorescence עבור אותו endplates כמו בפנל. עוצמת endplate (C) מרחיקות האדומה-BGT הקרינה המתקבלת ממדגם שריר שני על ידי מחבר NG. (ד) עצמת immunofluorescence Anti-rapsyn מאותו endplates כמו בC פנל.
| מחקר | שריר | מספר העכברים | אזור AChR | אזור synaptophysin | אזור חפיפה |
| ממוצע ± SD (μm2) | ממוצע ± SD (μm2) | ממוצע ± SD (μm2) | |||
| Morsch et al. (2012) 1 | הגסטרוקנמיוס | 3 | 181 ± 7 | 163 ± 24 | |
| סרעפת | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch et al. (2013) 1 | tibialis קדמי | 3 | 166 ± 26 | 117 ± 21 | nd |
| Cheng et al. (2013) 2 | tibialis קדמי | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| Tse 2 (לא פורסם) | tibialis קדמי | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| 1 צילם על מיקרוסקופ Meta Zeiss LSM 510 אבל עם רווח קבוע ולקזז רמות. מדידות ערכי סף ואזורים הסינפטי משמשות softwar Metamorphדואר על ידי MM. | |||||
| 2 צילמו על מיקרוסקופ Leica DM IRE2 ונותחו בעקבות פרוטוקול נוכחי על ידי המחבר הראשון מצויינים, עיוור לקבוצת טיפול. | |||||
| nd לא נקבע. | |||||
אזורי טבלה 1. Synaptic לNMJs של נקבה בת 2 חודשי C57Bl / 6J (ביקורת בריאה)
| מחקר | שריר | סריג יעילות (%) * | טווח (%) |
| (ממוצע ± SEM) | |||
| אל Brockhausen et. (2008) | tibialis קדמי | 24 ± 1 | na |
| קול et al. 2,010 | tibialis קדמי | 26 ± 1 | 22-30 |
| Morsch (לא פורסם) | סרעפת | 37 ± 1 | 24-47 |
| Ghazanfari (לא פורסם) | tibialis קדמי | 30 ± 1 | 20 - 45 |
| * סריג בין (הרדיוס האדום-BGT ומרחיקה אדומה-BGT פורסטר לסריג זוג = 51a (Life Technologies). | |||
| נתונים na אינו זמינים | |||
טבלה 2. ייעול לAChR סריג מעכברי עכברים הבוגרים הצעירים C57Bl6J
Subscription Required. Please recommend JoVE to your librarian.
Discussion
הפרוטוקולים שתוארו כאן, אפשרו לנו למדוד באופן מהימן ולכמת שינויים במאפיינים של NMJ על פני מגוון מצבים, כולל מדינות הזדקנות ומחלות נורמליות. השיטות שתוארו עבור en פנים תמונות NMJ תאפשר לחוקרים להשוות את השטח של התמחויות טרום postsynaptic ואזור חפיפה / יישור הסינפטי. כדי להשוות את עוצמתם היחסית של חלבונים טרום postsynaptic הפרוטוקול השני, המשתמש בחלקים אופטיים רוחביים, הוא מועדף. הפרוטוקול השלישי בוחן באופן ספציפי לשינויים בקרבת האריזה של AChRs בקרום postsynaptic.
בקרות ספציפיות חיוניות במיקרוסקופ immunofluorescence. בעת השימוש בכל נוגדן ראשוני לimmunofluorescence העקיף יש צורך ראשון להבטיח שהוא נקשר באופן ספציפי לחלבון מטרתה בסעיפי השריר. סוגים שונים של עיבוד וקיבוע רקמה יכולים דיפרנציאלי לשנות את הספציפיות שלנוגדנים. חשוב לוודא שמכתים immunofluorescence (לrapsyn למשל) באמת מרוכזת עם AChR בendplate המנוע. סעיפי ביקורת שליליים גם חייבים להיבדק כדי לוודא שהנוגדן המחייב הוא ספציפי. לדוגמא, הביקורת השלילית הטובה ביותר עבור immunofluorescence rapsyn תהיה חלקים מrapsyn - / - עכברים. אלה צריכים להראות לא מכתים endplate עם אנטי rapsyn. הקרינה שאינה ספציפית יכולה לנבוע גם מכימיקלים ניאון אנדוגני ברקמות (autofluorescence) או מכריכה שאינה ספציפי על ידי המצומד נוגדנים משני הניאון. הקרינה מסוג זה החמיר לעתים קרובות על ידי קיבוע אלדהיד. בנוסף, מכתים TRITC-BGT של endplates ניתן לפעמים למצוא בערוץ הקרינה FITC ולדמם דרך ניאון זה עלול לבלבל אותם עם immunofluorescence FITC הספציפי. כדי להתגונן מפני שלוש הצורות האחרונות של הקרינה שאינה ספציפית, כל אצווה של שקופיות שהם מוכתמים צריכה לכלול 'n כמהסעיפי "שליטת o-יסודי נוגדן (שלבים 3.7 ו -4.6). תמונות של endplates מסעיפי שליטה אלה צריכים להיות בהשוואה לאלו משקופיות הניסוי כדי להבטיח שמכתים immunofluorescent העקיף של NMJs באמת משקף את מחייב של הנוגדן הראשוני.
רוחבי סעיפי confocal הם שימושיים במיוחד להערכת הבדלים בעוצמתם היחסית של immunostaining בסינפסה. בסעיפי confocal רוחביים קל יותר לשפוט שיתוף לוקליזציה מדויקת של חלבונים סינפטיים. פרופיל endplate בצורה חצי סהר מייצג רק לחתוך דרך NMJ מדגם בשאלה. עם זאת, על רקע הקרינה (extrasynaptic) היא בדרך כלל נמוך יותר בהשוואה לen תמונות Z- הקרנת פנים. כך, יכול להיות קל יותר להבחין immunostaining 'האמיתי' (ספציפי) ולהקים רווח confocal קבוע וקיזוז ערכים באמצעות סעיפים אופטיים רוחביים 13-15,18. לדוגמא, במודל של עכברים של מיאסטניה גרביס (wherendplates צביעת AChR endplate הדואר מופחת במידה ניכרת) הותווה בבירור בסעיפים אופטיים רוחביים 18,21. הבדלים בעוצמת הממוצעת של הקרינה בNMJ עשויים לשקף צפיפות שינה של חלבון המטרה בתוך ההתמחות הסינפטי. אזהרה היא ש, במצבים מסוימים, שינוי מבני בחלבון המטרה או החסימה של נוגדן מחייב על ידי חלבונים שכנים עשוי להסביר עוצמת צביעה שונה.
העיצוב של ניסויים דורש קצת התחשבות. במקרים רבים הניסוי היה מטרה לבדוק את ההשפעה של transgene, מציאה גן או מצב מחלה בגודל של NMJ. קבוצת המדגם הניסיונית עשויה אז להיות בהשוואה לעכברים צעירים ובריאים (wild-type) מאותו המין והרקע גנטי. ערכי בסיס לאזור של synaptophysin endplate, AChR וחפיפה הסינפטי לכמה שרירים מוצגים בטבלה 1. גודל מדגם יהיה תלוי במידה של בעלי החייםוריאציה -כדי-החיה בתוך קבוצות טיפול וגודל האפקט (הבדל באמצעים לניסוי לעומת קבוצת ביקורת לסטיית התקן). כאשר הניתוח מוגבל לתמונות באיכות טובות מידת העקביות הוגנת נמצאה במדגם המשמעות לאזורי endplate בקרב עכברים בריאים 2 חודשים ישנים נקבת C57Bl6J (איור 6 א 'וב'). לפיכך, ניתן היה להוכיח הפחתת 30-40% משמעותיים באזור הסינפטי בעכברים שהוזרקו עם IgG ממיאסטניה אנטי-מושק חיובית גרביס חולים, בהשוואה לקבוצת ביקורת עם גודל מדגם של שלושה עכברים 17,20,32. עכברי קשישים מוצג גדולה יותר וריאציה של בעלי חיים לבעלי החיים בפרמטרי endplate מעכברים צעירים 22. כתוצאה מכך עשויים ניסויים מעורבים עכברים בגילים דורשים גודל מדגם גדול יותר.
אם הדאגה העיקרית היא למדוד את גודל en הפנים הגדרות רמה לקזז endplate אז הרווח וצריך להיות מותאם לכל NMJ הבודד. אישי קלאסיNMJs idual יכול להשתנות במידה ניכרת בבהירות של AChR וצביעת synaptophysin, במיוחד כאשר מדינות מחלה נבחנות. יתר על כן עוצמת הקרינה נוספת-סינפטי (לא ספציפית) היא לעתים קרובות יותר ויותר משתנה בשרירים של בעלי חיים מזדקנים, בהשוואה לאלו של בעלי חיים צעירים ובריאים (איור 5 ג ו- D). 1-256 אפורה בקנה המידה צריך להיות מנוצל במלואו על מנת למקסם את מידע הגוונים שיישמרו בתמונות הסופיות. זה יהיה כרוך התאמת הרווח לקזז רמות לכל NMJ שZ- מחסנית היא שייגבה. איור 5D מראה דוגמא של תמונת NMJ שבו מידע גוונים יכול להיות קריטי בהגדרת הגבולות של שטח של לפני ואחרי התמחויות הסינפטי.
מדידות של אזורים הסינפטי יכולות להיות מיושמות על הכנות שרירים שונות וניסויים. רוב המדידות של אזורים הסינפטי שלנו מועסקים cryosections האורך מצמד שרירים קפוא. הקפאת השרירים לפני הקיבוע שומרת antigenicity של מגוון רחב של חלבונים. כאשר תואם עם אנטיגן, קיבעון paraformaldehyde וחדירת סוכרוז לפני cryosectioning (שלב 2.1) יכול לספק שימור טוב יותר של מבנה NMJ. שימור מבני אופטימלי עשוי להיות מושגת על ידי זלוף לב עם paraformaldehyde. חפצים של הקפאה וחתך ניתן להימנע לחלוטין על ידי endplates תיוג על פני השטח של NMJs השריר וההדמיה שלם על fascicles הקניט מהשרירים קבועים 21. ללא קשר להכנה, הנהלים לדגימה, הדמיה וquantitation האזור יישארו ללא שינוי (פרוטוקול השלבים 4-5). יישום עקבי של פרוטוקולי דגימה, הדמיה וניתוח עיוורים (באמצעות מפעילים שונים, מדגמים שונים של עכברים וזמנים שונים), יכול לגרום לערכים ממוצע למדי לשחזור (השווה Cheng et al. וTse תוצאות בטבלה 1).
"> Endplates תוארו כהופך" מקוטע "במגוון רחב של מצבי מחלה. לדוגמא, בהזדקנות שרירי עכבר, ניוון סדיר של סיבי שריר (ואחריו התחדשותה) הביא לשיפוץ של שלט AChR endplate כמו כעך ל יצירת צבירי AChR מרובים קטנים יותר 6. בעכברים שהוזרקו עם IgG ממיאסטניה גרביס אנטי-מושק חולים, הפיצול של endplate היה שונה למדי. כעך AChR endplate במידה רבה התפזר, והותיר אחריו קבוצת כוכבים של (<4 מ '2) microaggregates' AChR הזעיר '20,21. שתי דוגמאות אלו מדגישות את הצורך להשוות את הפצות גודל לאשכולות AChR בendplates שליטה לעומת חיות ניסוי 21.שיטות אחרות לא דווחו להערכת שטח הסינפטי או מכתים עוצמה בNMJ. לפעמים יכול להיות מקופל endplates מנוע כך שתמונות Z- ההקרנה דו-ממדיות המשמשות כאן עלולות לזלזל synאזורי aptic. שחזורי confocal תלת-ממדיים עשויים לספק אמצעים מדויקים יותר אם האזור הסינפטי מוחלט חייב להיות מוגדרים 33. יתרון עיקרי של פרוטוקול Z- ההקרנה המתואר כאן, לעומת זאת, הוא הפשטות היחסית שלה, שאפשרה למספר גדול של endplates להימדד מקבוצות טיפול מרובות וזיהוי אמין של שינויים אפשריים. הפרוטוקול להשוואת עוצמות צביעת endplate יכול להיות מותאם ללימוד שינויים ברמות של חלבונים סינפטיים רבים ושונים. השיטה היא מוגבלת, עם זאת, על ידי הדרישה שכל הדגימות מעובדות לimmunofluorescence אז צילם באותה הפגישה confocal. מחקר שנערך לאחרונה על ידי ימפולסקי et al. 5 תאר שיטה למדידת צפיפות endplate AChR שעשויה לעזור להתגבר על מגבלה זו. במחקר זה, באותו תחום endplates היו צילם במספר הגדרות כוח הלייזר שונה. השיפוע של היחסים בין כוח הלייזר וrhodamiעוצמת הקרינה ne-BGT שימשה להערכת שינויים יחסי בצפיפות AChR בendplates בעכברים שונים 5. שיטה זו עשויה להיות שימושית להשוואת עוצמת AChR בדגימות צילמו במועדים שונים במהלך מחקר ממושך.
AChR-AChR סריג מספק מידע ספציפי ומשלים על הארגון של AChRs endplate. autoradiography המיקרוסקופי אלקטרונים באמצעות 125. I-α-BGT הראתה AChRs להיות צפוף עם צפיפות מישוריים של 10 מ '4 -2 מייד תחת כל אתר presynaptic של שחרור משדר, ואילו infoldings קרום הסמוך להכיל צפיפויות נמוכות בהרבה של AChR 34. AChR-AChR סריג עושה את זה קל יחסית כדי לזהות שינויים (תת-מיקרוסקופיים) באריזת AChR. ירידה ביעילות סריג משקפת חלוקה מחדש תת-מיקרוסקופי של AChRs בקרום postsynaptic שעשוי להיות לא זוהה על ידי שינוי בעוצמת הקרינה BGT ממוצעת. f מרובהשחקנים עשויים לגרום לשינוי ביעילות של סריג. אלה כוללים את מרווח acceptor התורם ואורינטציות יחסי, כמו גם את הסביבה המולקולרית 35,36. ירידה ביעילות סריג endplate עשויה אולי לנבוע משינוי בגיאומטריה של סריג AChR. עם זאת, סביר להניח שזה יהיה בשל הפחתה בשיעור של AChRs שהם ארוזים בתוך בקנה מידה ננו סריג המולקולרי postsynaptic 14.
נראה אובדן הקשר בין הנוירונים מוטוריים וסיבי שריר להיות הגורם המיידי לחולשת שרירים במחלת הנוירון המוטורי ובהזדקנות פעילה 9,22,23. שיטות משותפות ופרמטרים למדידת NMJs צריכים לעשות את זה קל יותר עבור קבוצות מחקר שונות כדי להשוות וממצאי ניגוד פורסם. השיתוף של פרוטוקולים מפורטים (ושיפורים עתידיים עליהם) עשוי לעזור להאיץ את ההתקדמות בהבנת המנגנונים של תחזוקת NMJ ואיך זה יכול להיות מושפע במצבי מחלה.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
