

Abstract
La jonction neuromusculaire (JNM) est le grand, le relais synapse cholinergique par lequel les neurones moteurs de mammifères contrôlent contraction des muscles volontaires. Les changements structurels au NMJ peuvent entraîner l'échec de la neurotransmission, résultant dans la faiblesse, l'atrophie et même la mort de la fibre musculaire. De nombreuses études ont étudié comment les modifications génétiques ou de maladies peuvent modifier la structure de la jonction neuromusculaire de la souris. Malheureusement, il peut être difficile de comparer directement les résultats de ces études parce qu'ils travaillent souvent différents paramètres et les méthodes analytiques. Trois protocoles sont décrits ici. La première utilise la projection d'intensité maximale images confocales pour mesurer la surface des récepteurs de l'acétylcholine (RACh) riche domaines membranaires post-synaptiques à la plaque d'extrémité et la surface de la vésicule synaptique coloration dans la terminaison nerveuse présynaptique sus-jacente. Le deuxième protocole compare les intensités relatives des immunocoloration des protéines synaptiques dans la membrane post-synaptique. La troisième protocole utilise Fluorescence Resonance Energy Transfer (FRET) pour détecter les changements dans l'emballage d'AChR post-synaptiques à la plaque d'extrémité. Les protocoles ont été développé et affiné au cours d'une série d'études. Les facteurs qui influencent la qualité et la cohérence des résultats sont discutés et données normatives sont fournis pour NMJs chez les jeunes souris adultes en bonne santé.
Introduction
La jonction neuromusculaire (JNM) est la synapse de relais essentiel qui médie la communication entre le système nerveux et le muscle squelettique. Il est nécessaire pour tout mouvement volontaire. La microscopie à fluorescence a longtemps été utilisé pour étudier les effets des transgènes sur la souris NMJ 1-3 ou de comparer les effets de l'âge, l'alimentation, l'exercice et la maladie sur NMJs de rongeurs 4-11. Ces études nous ont beaucoup appris sur la physiologie et la physiopathologie de la NMJ, mais les divers paramètres déclarés (par exemple, la zone AChR, zone flasque, longueur du périmètre, indices de fragmentation) font qu'il est souvent difficile de comparer les résultats de ces études. Il ya une attente de plus en plus pour les chercheurs précliniques pour pouvoir démontrer la reproductibilité, en particulier dans les études avec des modèles de rongeurs de la maladie 12. Les protocoles décrits ici ont été affinées par une série d'études qui a enquêté ch développement, physiologique et physiopathologiqueAnges à l'NMJ. Ces études nécessitent la mesure de la zone de spécialisations synaptiques à la plaque motrice de la souris et de la densité relative de l'emballage de protéines synaptiques dans les spécialisations post-synaptiques 13-15.
L'utilité de ces méthodes est illustrée par des études récentes dans un modèle de souris de l'anti-MuSK myasthénie. Injections quotidiennes d'IgG de myasthénie anti-MuSK positif gravis patients dans des souris adultes causés à devenir faible dans les deux semaines 16. Confocale images maximale de projection de sections musculaires qui ont été doublement marquée pour la synaptophysine (en nerveuses terminaux) et post-synaptiques AChR révélé un déclin progressif dans le domaine de AChR coloration que le principal changement. Surtout le taux de déclin était suffisante pour expliquer les baisses comparables dans l'amplitude des potentiels synaptiques, l'échec de la transmission synaptique et une faiblesse musculaire 17,18. Qualitativement résultats similaires ont été rapportés par d'autres groupes de recherche10,19. Les mêmes méthodes de mesure NMJ ont depuis été utilisés pour évaluer l'impact de trois médicaments pour le traitement anti-MuSK myasthénie grave dans ce modèle de souris 20,21.
Vieillissement sédentaire peut conduire à la perte de connexions neuromusculaires. Les protocoles décrits ici ont révélé une baisse liée à l'âge dans le domaine de terminaison nerveuse synaptophysine à plaques motrices que les souris progressent dans la vieillesse. Les mêmes méthodes ont révélé que l'exercice volontaire pourrait largement éviter la réduction de nerf présynaptique zone de la borne 22, en accord avec les travaux antérieurs par d'autres groupes 4. Perte de connexions neuromusculaires se produit également dans le modèle de la souris SOD1G93A de la sclérose latérale amyotrophique 9,23.
Les études mentionnées ci-dessus démontrent que une variété de conditions de santé peut conduire à des réductions dans le domaine de spécialisation soit pré- ou post-synaptiques au NMJ. Cela peut entraîner dans l'amusement synaptique altéréection ou peut annoncer une perte complète de la connexion neuromusculaire. Trois protocoles sont décrits qui permettent la quantification de la superficie et de la densité de spécialisations synaptiques. Le but du premier protocole est de fournir une mesure pratique et reproductible des zones de pré et post-synaptiques spécialisations et leur alignement au NMJs mammifères, en utilisant la microscopie de fluorescence. Images bidimensionnelles projection confocale maximales et analyse d'image avec les NIH ImageJ est utilisé pour détecter des changements dans le domaine de la coloration (synaptophysine des vésicules synaptiques), AChR post-synaptiques et de la zone de chevauchement synaptique. Paramètres d'imagerie confocale (gain et décalage de niveau) sont optimisés pour chaque NMJ de façon à maximiser l'information visuelle utilisée pour discerner le domaine de spécialisation synaptique. Insuffisance neuromusculaire peut également résulter de changements dans la densité post-synaptique du CCRS et / ou d'autres protéines synaptiques. Le deuxième protocole peut être appliqué à détecter les changements dans la densité relative de ces protéines postsynaptiquesque le musc, rapsyne, dystroglycane, phosphorylée kinase Src et phosphorylée AChR 18,21.
Dans la myasthénie grave, une densité réduite de AChR dans la membrane post-synaptique est la cause immédiate de l'insuffisance synaptique et une faiblesse musculaire. Le troisième protocole décrit une méthode Fluorescence Resonance Energy Transfer (FRET) pour évaluer les changements dans la proximité de AChR adjacentes dans des membranes post-synaptiques 14,15. Cette méthode détecte transfert d'énergie entre AChR voisins marqués par fluorescence-α-bungarotoxine (BGT). FRET se produit uniquement lorsque les sondes fluorescentes donneur et accepteur sont à moins de 10 nm de l'autre. Cela peut révéler (microscopiques) des changements dans l'étanchéité de AChR emballage qui peuvent être directement liés à l'amplitude des potentiels synaptiques.
Ces trois protocoles, raffinés au cours de la dernière décennie, fournissent des mesures complémentaires d'intégrité NMJ d'une manière cohérente et reproductible. Utilisation de protocoles normalisés unend paramètres devraient faciliter la comparaison des effets des gènes et des interventions environnementales sur le NMJ mammifères.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
REMARQUE: Conception, réalisation et l'enregistrement de l'expérimentation animale devraient tenir compte des lignes directrices actuelles 24. Ce travail doit être approuvée à l'avance par l'autorité de protection des animaux locale (dans notre cas, le Comité d'éthique animale de l'Université de Sydney).
1. L'euthanasie de l'animal et Muscle Dissection
- Transférer la souris à partir de la salle d'attente dans une salle séparée où il est euthanasié par une injection intrapéritonéale de solution de pentobarbital (30 mg / kg) en utilisant la méthode de manipulation de la souris détaillé par Shimizu 25. Placez la souris de nouveau dans sa cage.
- Une fois que la respiration de la souris se est arrêté pendant plus de 1 min, tester le réflexe pieds retrait en pinçant doucement le pied, et le réflexe de la cornée en effleurant la cornée. Seulement lorsque les réponses réflexes sont absents de la souris peut être préparé pour la dissection.
- Consultez un atlas de l'anatomie rongeurs tels que Chiasson 26 et / ou demander l'aide d'un experienanatomiste ced avant de tenter la dissection du muscle d'intérêt. Dans chaque cas, enlever les poils de la peau recouvrant aide d'un petit rasoir électrique avant d'ouvrir la peau pour exposer le muscle.
NOTE: La dissection sera différent pour chaque muscle anatomiquement distincts. - En utilisant des pinces contondants libérer le muscle de membranes recouvrant et les tissus environnants. Saisir et couper le tendon distal de séparer le muscle de son insertion.
- Taquiner doucement et coupez le muscle libre du tissu environnant de retour à son origine. Placer brièvement le muscle nouvellement disséqué dans 0,1 M de tampon phosphate salin (PBS) ou la solution de Ringer avant tout autre traitement.
2. Préparation de la Muscle pour Cryosectioning
REMARQUE: conservation structurelle optimale peut être obtenue par perfusion de l'animal entier comme précédemment détaillé 27, ou la fixation de l'immersion (par petits muscles) comme décrit à l'étape 2.1 en option. Cependant,4% de paraformaldéhyde fixation peut nuire coloration subséquente avec de nombreuses sondes anticorps et fluorescente-BGT. Le glutaraldéhyde en particulier doit être évitée. Si les muscles ne sont pas à fixer, ils doivent être immédiatement congelés rapidement (passer à 2,3).
- Facultatif fixation d'immersion: Pin le muscle à la cire dans un boîte de Petri à longueur de repos. Couvrir le muscle avec 2% p / v de paraformaldehyde (fraîchement dissous dans du PBS) pendant 2 heures à température ambiante. Laver avec trois changements de PBS plus de 30 min (3 x 10 min), puis remplacer le PBS avec 30% p / v de saccharose dans du PBS et incuber O / N à 4 ° C.
- Fabriquer des moules ('bateaux') à l'avance par pliage 2 cm x 1,5 cm morceaux de feuille d'aluminium, comme illustré à la figure 1. Placer un morceau de membrane de nitrocellulose dans le fond du bateau. Verser délicatement cryostat intégration matrice (de table Matériaux) dans le bateau à une profondeur de 2 mm, en prenant soin d'éviter les bulles d'air. Placez le muscle dans le bateau, l'alignant sur les lignes de stylo à bille surla nitrocellulose. Ajouter plus matrice intégrant de façon à couvrir complètement le muscle (Figure 1).
- Pré-étiquette en polypropylène tubes avec un marqueur indélébile. Placez une goutte d'eau dans chaque tube et le tube refroidir dans de l'azote liquide.
REMARQUE: La chute de l'eau gelée maintient la pression de vapeur et empêche la dessiccation pendant prolongé stockage -80 ° C - L'utilisation d'un masque de protection, gants de protection épais et une grande paire de pince émoussée, abaisser partiellement un petit gobelet métallique (3 cm de diamètre, 8 cm de profondeur) contenant 2 cm de profondeur d'isopentane dans un récipient d'azote liquide pendant 30 sec. Retirer le bécher et le placer sur la paillasse. En utilisant une petite paire de pinces émoussés placez le moule contenant le muscle et l'incorporation dans la matrice isopentane refroidi. Prenez soin d'éviter le mélange d'azote liquide avec l'isopentane.
- Autoriser 2 min pour le bloc de geler complètement avant d'utiliser le forceps émoussé pour soulever le bloc congelé sur et le sceller dans le bon pre-étiquetés et tube pré-réfrigéré (étape 2.3).
- Stocker temporairement les tubes dans de l'azote liquide avant le transfert à -80 ° C. Connectez-vous tous les échantillons dans un tableur du contenu des congélateurs.
3. Cryosectioning et coloration fluorescente pour en Face Images de NMJs
- Décoller le moule en aluminium. Au sein de la chambre à -20 ° C cryostat fixer le bloc congelé de la pince de cryostat de façon à découper 20 um cryosections parallèles à l'axe long des fibres musculaires (figure 1). Ramassez les sections sur les poly-L-lysine ou des lames de microscope de gélatine enduit.
- REMARQUE: omettre cette étape si le tissu est fixé avant la congélation. Après avoir laissé 30 min pour les sections de sécher sur les lames, les fixer en plaçant une baisse de 2% de paraformaldehyde dans PBS sur chaque section pendant 15 min à température ambiante.
- Laver les lames 3 x 10 min dans du PBS dans une jarre de Coplin, puis plonger les lames dans du PBS contenant 0,1 M de glycine pendant 30 min pour bloquer des groupes aldéhyde résiduels.
- Laver les lames pendant 10 min dans du PBS, puis plonger dans le méthanol (refroidi à -20 ° C) pendant 7 min. Cette étape de perméabilisation est une partie de la routine d'un double marquage fluorescent avec BGT et anti-synaptophysine mais il peut nuire immunocoloration pour d'autres protéines.
- Laver les lames 2 x 10 min dans du PBS, puis placer chaque diapositive dans une chambre humidifiée stable et nivelé. Couvrir immédiatement chaque section avec 20 ul de solution de blocage (0,2% de Triton X-100, 2% d'albumine de sérum bovin (BSA) dans du PBS) pendant 1 heure à température ambiante. Les articles ne doivent pas être autorisés à sécher à toute étape du processus d'immunocoloration.
- Effectuer la première incubation: Prendre une diapositive à la fois retirer soigneusement la solution de blocage en excès de plus de chaque section et le remplacer par 20 pi de lapin anti-synaptophysine (dilué à 1: 200 dans la solution de blocage).
- Inclure une lame témoin négatif qui sera incubé avec une solution de blocage seulement. Cette «non-contrôle de l'anticorps primaire'Est essentiel dans chaque course immunocoloration.
- Prendre soin que l'anticorps primaire reste en place sur chaque section, fermer la chambre humidifié et incuber pendant 1-2 jours à 4 ºC.
- Inspectez chaque section pour confirmer que l'anticorps primaire reste en place. Utiliser une pipette Pasteur pour rincer délicatement chaque diapositive avec du PBS et le placer dans une jarre de Coplin. Lavez toutes les diapositives 3 x 10 min dans du PBS.
- Effectuer incubation secondaire. Prenant une diapositive à la fois, retirez soigneusement excès de PBS, posez-le dans la chambre humide et couvrir chaque section avec 20 pi d'un mélange contenant FITC conjugué IgG d'âne anti-lapin et BGT conjugué à tétraméthyl rhodamine ou d'une autre fluorophore rouge (TRITC- / redBGT; 5 g / ml) dilué dans une solution de blocage. Incuber à température ambiante pendant 2 h.
- Laver les lames 3 x 10 min dans du PBS dans des bocaux de Coplin.
- Prenant une diapositive à la fois, retirez soigneusement excès de PBS et monter avec une lamelle en utilisant un volume minimal de, glycérobase l, fade-résistant milieu de montage. Sceller les bords des lamelles avec vernis à ongles transparent. Laisser sécher dur.
- Conserver les diapositives dans l'obscurité à 4 ° C pendant jusqu'à une semaine, ou à -20 ° C pour des périodes de stockage plus longues (jusqu'à plusieurs mois).
4. impartiale échantillonnage et d'En Face Imaging Motor Plaques de fond
- Aveugler les diapositives par marquage de chaque lame avec un numéro de code aléatoire qui reste connu que d'un second chercheur (non impliqué dans l'analyse). En conséquence, l'opérateur reste aveugle pour les groupes de traitement jusqu'à ce que la quantification des paramètres NMJ est terminée pour tous les échantillons.
- Placez la lame sur la platine du microscope et voir sous large éclairage sur le terrain avec l'ensemble de filtre TRITC (63X huile 1,3 objectif NA). Déplacez progressivement (champ par champ) de gauche à droite et en arrière jusqu'à une plaque d'extrémité apparaît dans le champ (figure 2A).
critère d'échantillonnage: NOTA: Toute structure AChR teinté qui est relativementplat et fait face à l'objectif (ce est à dire, se étend <15 m dans la dimension z) est considéré comme une plaque d'extrémité et est imagée pour l'analyse (croissants de AChR coloration représentent des sections transversales à travers plateaux et sont donc exclus). - Avec l'ensemble sténopé confocal à 1,0 unité d'Airy et la puissance laser de faible optimiser le gain et le décalage de niveaux TRITC / rouge-BGT (laser de 532 nm) à la plaque d'extrémité qui est à imager. Suivant optimiser FITC / fluorescence synaptophysine en utilisant le laser de 488 nm. Recueillir un z-pile de la plaque d'extrémité avec un intervalle de 0,7 um entre chaque tranche optique. Enregistrez les images avec un nom de fichier qui comprend la date de la session d'imagerie, le nom de code de la diapositive et le numéro de la plaque d'extrémité.
REMARQUE: Les balayages utilisant le 488 nm et 532 nm lasers (FITC et TRITC) doivent être prélevés de manière séquentielle (pas simultanément) pour éviter la contamination du canal FITC par fluorescence du fluorophore rouge et vice-versa (transpercement). - Répétez l'échantillonnage d'une imagerie d'étapes 4.2 à 4.3 jusqu'à 20 plateaux sont collectées de la diapositive / échantillon.
- Changer à la diapositive codé suivant et répétez 04/02 au 04/04. Répétez cette opération pour chacune des lames codées.
- Recueillir quelques images de plateaux de la lame de contrôle (sans primaire contrôle de l'anticorps) en utilisant les paramètres de confocale qui ont été trouvés optimale pour les diapositives expérimentales (le canal de fluorescence FITC devrait apparaître sombre).
- À la fin du transfert de session confocale les fichiers d'image à un autre ordinateur et de sauvegarder les fichiers originaux sur un disque dur externe ou un serveur.
5. Mesurer la zone synaptiques Spécialisations en en Face Images
- Utilisez NIH ImageJ freeware (http://imagej.nih.gov/ij/) pour préparer la projection maximale (MIP) des images de chaque z-stack. Enregistrez-les sous forme de fichiers TIFF (Figure 2A et B). Les noms de fichiers doivent inclure la date de la session de l'image, des exemples de code, le numéro de plaque d'extrémité et le canal fluorescent (par exemple, 060414_5723_7_FITC.tiff).
- Ouvrez l'image z-projection dans ImageJ. Sélectionnez l'image canal de récepteur de l'acétylcholine (figure 3A) et sélectionnez: Image> Type> 8-bit pour convertir l'image RVB 24 bits couleur en trois huit bits images en niveaux de gris sur l'écran.
- Utilisation de l'outil polygone ImageJ dessiner une esquisse autour de la plaque d'extrémité d'intérêt dans le redBGT teinté (CADH) canal de manière à inclure toutes les régions colorées apparents de la plaque d'extrémité individu en particulier, tout en excluant tout coloration qui ne proviennent pas de la plaque d'extrémité d'intérêt ( Figure 3C).
- Appliquer un seuil minimal d'intensité de l'image en sélectionnant: Image> Réglages> Seuil (Figure 3E et captures d'écran de ImageJ associés).
- Ajustez le niveau de seuil de façon à isoler les parties AChR colorés tout en excluant entourant signal de fond en tant que sous-seuil (Figure 3E). Ouvrez un second souffleow avec l'image originale (ton continu) immédiatement à côté de la fenêtre de comparaison, afin de faciliter la décision quant à la valeur de seuil. Notez la valeur de seuil pour une utilisation ultérieure dans colocalisation analyse.
- Conserver le polygone contour autour de la plaque d'extrémité sélectionnez: Analyse> Analyser particules. Dans le menu pop-up spécifier la gamme de tailles que: 50 à l'infini pixels (ce qui élimine les artefacts minuscules dus au bruit électrique dans le photomultiplicateur).
- Analyser commande particules génère une fenêtre avec une liste de domaines supra-seuil discrètes et leurs valeurs d'intensité de fluorescence numérotée comme ils apparaissent dans l'image binaire (Figure 3G et associé ImageJ capture d'écran). Copiez ces données dans un tableur marqué.
- Mesurer la surface de flasque total (de la zone dans le polygone) en sélectionnant: Analyse> Mesure. Cela donne la zone de flasque totale. Copiez et collez les données pour les régions et l'intensité dans AChRune feuille de calcul en veillant à étiqueter correctement colonnes, rangées sera utilisé pour plateaux individuels pour des diapositives spécifiques.
- Commuter sur le canal de fluorescence anti-synaptophysine et répétez les étapes 5.1 à 5.5, mais pour le canal FITC (figure 3B, D et F). Le but est d'ajuster le seuil afin qu'il crée une image binaire qui, aussi étroitement que possible, correspondant aux limites de coloration telle que perçue par l'œil. Notez la valeur de seuil.
- Mesurer la zone de chevauchement en appliquant les étapes suivantes: Ouvrez le fichier original contenant les deux images canal et le diviser en deux images séparées en sélectionnant: Image> Piles> Empiler aux images.
- Utiliser le plugin de colocalisation (téléchargé et installé à partir de la page Web ImageJ) Sélectionner: Pluggin> colocalisation et d'entrée les valeurs de seuil précédemment enregistrés pour les canaux RACh et nerveuses dans le canal respective requête bbœuf. Cela donnera une image de chevauchement en pixels blancs (Figure 3H et captures d'écran de ImageJ associés).
- Convertir l'image de chevauchement nouvellement créé dans un format en niveaux de gris et d'appliquer un seuil à la valeur maximale. Le seuil maximum ne sélectionner les pixels blancs correspondant à la zone de recouvrement des deux voies précédentes. La fiche dans la feuille de calcul de la valeur de la surface résultant de la «co-localisation», qui représente la zone de chevauchement en pixels.
- Préparer une feuille de calcul des moyennes des échantillons de données, calculer et tracer les écarts-types et les erreurs standard que les histogrammes ou des diagrammes de dispersion 20,22. A noter que la valeur de n représente généralement le nombre de souris par groupe d'échantillons à des fins statistiques.
- Terrain plateaux vertébraux zones AChR que des nuages de points ou histogrammes de fréquence pour déterminer si les données sont normalement distribué avant le test statistique (Figure 6).
6. coloration RelativeIntensités comparées en utilisant des sections optiques transversales
REMARQUE: Pour que ce processus tous les échantillons musculaires ensemble de protocole et de l'image en une seule séance confocale. En planifiant une expérience permettre jusqu'à 30 min Temps d'imagerie par échantillon musculaire.
- Couper 15 um cryosections transversal à l'axe longitudinal des fibres musculaires et de recueillir sur des lames comme décrit à l'étape 3.1.
- Effectuer la coloration fluorescente comme décrit dans les étapes 3.2 à 3.13.
- Code, les Les lames colorées sorte que l'imagerie et analyses sont menées avec les aveugles de l'opérateur pour le groupe de traitement, comme décrit dans l'étape 4.1.
- En utilisant un objectif de fluorescence 40X (NA 0,75) brièvement en revue une section de chaque diapositive pour déterminer un seul gain et offset réglage de niveau pour AChR qui sera adapté pour tous les plateaux à travers toutes les diapositives de l'échantillon. La plus brillante flasque devrait alors être juste en dessous de 256 sur l'échelle de gris. Cette optimisation devrait être effectuée séparément pour la deuxième fluorescence canal (recueillies successivement). Enregistrez le gain fixe et le décalage réglages de niveau et ne les modifie pas pendant toute la session d'imagerie.
- Recueillir des images d'une lame standard de fluorescence (par exemple, non-blanchiment billes fluorescentes), en utilisant les mêmes paramètres, au début et à la fin de la session confocale pour détecter toute variation possible de l'intensité du laser.
- Utilisez le canal AChR pour numériser la diapositive progressivement pour localiser plateaux.
- Concentrer pour trouver le plan de coupe optique unique dans chaque champ microscopique qui contient le plus grand nombre de plateaux AChR colorées.
- Scannez cette section optique unique deux fois et enregistrer l'image moyenne (figure 4G).
- Garder la même focale plan de commutation au second canal de fluorescence (protéine d'intérêt) et de recueillir l'image comme à l'étape 6.8. Enregistrez le fichier d'image, y compris dans le nom de fichier: date de la séance d'imagerie, des exemples de code, numéro de l'image et un symbole pour indiquer le canal fluorescent.
- Déplacez la scène pour le champ suivant qui contient un ou plusieurs plateaux et répétez l'étape 6.8 à 6.9. Répétez jusqu'à ce que un total de 60 plateaux sont imagés.
- A la fin du transfert de la session d'imagerie tous les fichiers vers un autre ordinateur et de les sauvegarder.
- Ouvrez chaque fichier image d'origine et tout en regardant le canal AChR, sélectionnez: Image> Piles> Empiler aux images, de diviser les canaux.
- Sélectionner: Image> Type de> 8bit pour convertir à 8 bits en niveaux de gris le format sur l'écran. Faites cela pour deux canaux de fluorescence.
- Sélectionner: Image> Piles> les images à pile. Ouvrez une nouvelle pile de deux précédemment séparés images 8-bits. On peut ensuite passer facilement entre les deux canaux de fluorescence dans le guichet unique.
- Utilisez l'outil de polygone pour dessiner un line étroitement autour de la limite de la coloration AChR (figure 4E).
- Sélectionnez: Analyse> Mesure pour mesurer l'intensité de pixel moyenne pour AChR dans la zone délimitée (noter l'importance de tirer la ligne serrée). Copiez cette valeur dans un tableur marqué.
- En conservant le même contour du polygone (pour définir la zone à mesurer), passer sur la deuxième chaîne fluorescent (par exemple, la figure 4B, D, F) et sélectionnez: Analyse> mesure. Cela donnera l'intensité de coloration moyenne pour la protéine d'intérêt à l'intérieur de la zone définie par synaptique AChR coloration.
- Choisissez une zone éloignée de coloration de plaque d'extrémité visible puis sélectionnez: Analyse> Mesure pour mesurer l'intensité moyenne de fluorescence de fond. Répétez cette opération pour l'autre canal de fluorescence / s et copier les valeurs de fond dans le chiffrier de valeurs de fluorescence.
- Soustracter les valeurs de fond moyennes à partir des valeurs des plateaux vertébraux pour obtenir les intensités corrigées pour AChR et la protéine d'intérêt à chaque flasque.
- Divisez les valeurs d'intensité des plateaux vertébraux corrigées pour la protéine d'intérêt par l'intensité de fluorescence de BGT corrigée pour donner les ratios d'intensité de fluorescence 14,21
7. comparant la densité post-synaptique Membrane AChR utilisant FRET
NOTE: Ce protocole évalue la mesure dans laquelle AChR sont étroitement emballés (<10 nm espacement) dans la membrane post-synaptique. La combinaison de donneur et accepteur fluorophore précise est critique à cette analyse FRET. Les noms et détails des fluorophores sont donnés dans le tableau des matériaux. Leurs propriétés spectrales, par rapport à FRET, sont discutées dans nos articles précédents 14,15.
- Préparer cryosections transversales fixes comme décrit dans la section 6.1. Tous les groupes d'échantillons doivent être traités ensemble et de l'imaged dans la même session confocale.
- Bien mélanger 2,5 g / ml rouge-BGT (donneur de FRET) avec 10 g / ml rouge lointain-BGT (FRET accepteur) avec une solution de blocage dans un petit tube en plastique par pipetage de haut en bas 12 fois. Ce mélange 1: 4 molaire maximise l'efficacité de FRET 14.
- Placer chaque lame dans une chambre humidifiée, couvrir soigneusement chaque section avec une goutte (12 ul) du mélange ci-dessus et on incube pendant 1,5 heure à température ambiante.
- sections de contrôle: couvrent un petit nombre de sections avec 2,5 g / ml rouge-BGT (donneur seulement; contrôles C1 marqués), et aussi quelques sections avec 10 g / ml rouge lointain-BGT (accepteur seulement; contrôles C2 marqués). Incuber ces contrôles comme à l'étape 7.3.
- Laver les lames 3 x 10 min dans du PBS et monter en base de glycérol, fade résistant milieu de montage (voir l'étape 3.12).
- Effectuer l'échantillonnage des plateaux comme dans l'étape 6.7. Fluorescence du donneur et accepteur doit être parfaitement co-localisée à plateaux en raison de la liaison aléatoire des molécules fluorescentes-BGT.
- images de contrôle: Utilisation de l'objectif 40X et de faible puissance optimiser le gain de redBGT laser et offset réglages de niveau de plateaux d'un tiroir de commande C1. Optimiser le gain extrême redBGT et de compenser les niveaux pour les plateaux de lame de contrôle C2. Confirmer l'absence de toute fluorescence purge travers.
- Sans changer la puissance du laser, gagnent ou réglages de niveau offset se déplacent vers les diapositives expérimentales et de recueillir des images (pré-photoblanchiment) pour les deux canaux de fluorescence.
- Photobleach sélectivement l'extrême-rouge-BGT sur une partie d'un seul flasque en zoomant la zone de numérisation numérisation puis 10 fois avec le laser de 633 nm à 100% de la puissance. La fluorescence dans la zone balayée devrait devenir dim.
- Réinitialiser la puissance du laser et de zoom et de recueillir des images post-blanchiment sur les deux canaux fluorescents en utilisant les paramètres confocale établies à 7,7.
- Calculer l'efficacité de FRET (E) de l'augmentation en pourcentage de donateurs (rouge-BGT) fluorescence suivante photoblanchiment de l'accepteur (à l'extrême-rouge-BGT) selon la formule suivante *:

* Pour toutes les situations où la fluorescence du donneur augmente après photoblanchiment de l'accepteur.

Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Mesure de Synaptic zone au NMJ
Toute estimation de la zone repose sur le dessin d'une frontière à définir l'étendue de spécialisations synaptiques. Chez les jeunes adultes en bonne santé muscles images NMJ devraient afficher limites bien définies à la fois pour AChR et synaptophysine coloration (figure 2A et B). L'intensité de fluorescence à la fois pour la synaptophysine AChR et augmente brusquement à la limite entre la partie péri-synaptique et synaptique de la plaque motrice (figure 5A 'et B'). Pour ces images, un seuil minimum (juste au-dessus extrasynaptique fluorescence de fond) sera facilement isoler la zone AChR-riche ou synaptophysine riche de la plaque d'extrémité (lignes horizontale en pointillés (figure 5A 'et B')). Chez les souris âgées et, dans certains états pathologiques coloration de plaque d'extrémité pour AChR peut être moins intense, AChR bords de cluster peuvent sembler floue et lare peuvent être des niveaux plus élevés de autofluorescence extrasynaptique (Figure 5D; 17,22). La coloration par fluorescence avec des frontières indistinctes peut introduire des erreurs dans les estimations de la zone synaptique. Dans tous les cas, le but est de choisir un seuil qui donne une image binaire semblable dans la forme et la taille à l'AChR- ou zones de synaptophysine riches tels qu'ils apparaissent à l'œil dans l'image originale, en tons continus. Conduite de l'analyse en aveugle au groupe de traitement devrait réduire le risque de partialité subjective à l'étape de seuillage (étape 4.1). Parfois, des images claires ou floues plateaux vertébraux peuvent résulter d'une transformation sous-optimale. Figure 2C et D montre un exemple d'une image basse de plaque d'extrémité de la qualité d'un 2 mois vieille souris en bonne santé. Contours flous et faible coloration de synaptophysine peuvent ont surgi dans ce cas de la fonte partielle et de recongeler le muscle avant cryosectioning. Certaines sections de jeunes en bonne santé (contrôle positif) musculaire devraient être sectionnés une traitée en parallèle avec les échantillons expérimentaux pour se assurer que toute atteinte évidente dans les images NMJ ne est pas en raison de problèmes avec immunomarquage. Lots d'images compromis par le traitement sous-optimal devraient être exclus des analyses.
Pour en Face images z-stack, 15-20 plateaux est un échantillon de taille raisonnable pour estimer les zones synaptiques. Une grande diversité dans les formes et tailles de NMJs se trouvent dans ne importe quel muscle donné. Les diagrammes de dispersion révèlent une gamme considérable dans le domaine AChR riche parmi les plateaux du muscle tibial antérieur de toute souris individuelle (figure 6A). Néanmoins, la zone AChR moyenne (sur la base de 15 à 20 en images le visage des plateaux vertébraux) était similaire chez les souris sept échantillons (~ 200 m 2; Figure 6a). Le domaine de la plaque d'extrémité synaptophysine coloration varie aussi considérablement entre plateaux d'un muscle donné. Une fois de plus, en utilisant une taille de 15 à 20 de l'échantillon plateaux de la zone moyenne de flasque synaptophysine était SimiLAR chez les souris 7 étudié (~ 170m 2; Figure 6B). histogrammes de fréquence des données regroupées ont révélé distributions normales pour à peu près le domaine de la plaque d'extrémité AChR et synaptophysine (figure 6A 'et B'). Cependant une distribution normale des zones synaptiques ne peut pas supposer dans des états pathologiques tels que la myasthénie grave 16,20. Cela peut influer sur le choix du test statistique.
Tableau I énumère les domaines de spécialisations pré- et post-synaptiques pour NMJs pour saine vieille 2 mois (jeune adulte) femelle C57BL / 6J des études précédentes. Les domaines de spécialisation à la fois avant et post-synaptiques diminué avec le vieillissement sédentaire 22. AChR zone a également été nettement réduite chez les souris injectées avec des IgG anti-MuSK de myasthénie patients 17,21. Myasthéniques souris traitées avec le médicament inhibiteur de la cholinestérase, pyridostigmine, affichent une nouvelle réduction significative de la plaque d'extrémité zone AChR 20 </ Sup>.
Intensité relative des plateaux vertébraux marquage fluorescent
L'intensité relative du marquage en immunofluorescence peut révéler des changements dans la densité d'une protéine d'intérêt-de-synaptique avec l'âge, le génotype et / ou de l'état de la maladie. AChR fluorescence (rouge-BGT ou de loin-rouge-BGT) sert d'abord à définir l'emplacement de la jonction neuromusculaire. La luminosité de fluorescence dans le domaine de 8 bits AChR riche est ensuite utilisé pour évaluer les changements dans la concentration de la protéine d'intérêt, par rapport aux animaux témoins. Dans les sections transversales les AChR des plateaux vertébraux apparaissent généralement comme une forme de croissant, mais cette forme est souvent irrégulière (figure 4A, C, E, H). Intensité faible fluorescence de fond révèle généralement si une tache de coloration AChR représente une simple plaque d'extrémité, ou deux plateaux séparés situés sur les fibres musculaires adjacentes. Beaucoup de protéines synaptiques (comme rapsyne, de musc et CRS)colocalisés avec AChR à la plaque d'extrémité (figure 4A - D). La coloration par immunofluorescence avec des anticorps phospho-spécifiques peut également être utilisé pour comparer l'effet des interventions expérimentales sur l'état de phosphorylation de certaines protéines de la membrane 21 postsynaptiques.
La fiabilité et la reproductibilité de la mesure de l'intensité de fluorescence dépend fortement de l'intégrité du muscle congelé et la qualité de l'immunocoloration. Muscles doivent être disséqués et congélation rapide immédiatement ou paraformaldéhyde-fixe (à quelques minutes de la mort de l'animal) pour éviter des changements dégénératifs au NMJ. L'immunocoloration dépend fortement de la qualité des réactifs et d'optimiser le protocole de coloration pour des anticorps spécifiques. Pour tout nouveau lot d'anticorps primaire expériences pilotes immunocoloration sont nécessaires. Cryosections fraîchement coupées de jeunes muscles sains sont incubées avec des dilutions en série de 2 fois de l'anticorps primaire. Unanticorps secondaire fiables connue est utilisée et les résultats sont comparés. Si la protéine d'intérêt est connue pour être limitée à la JNM alors la meilleure concentration de l'anticorps est celle qui donne le ratio le plus élevé de l'intensité de fluorescence NMJ par rapport à celle trouvée dans les parties extrasynaptiques du muscle (fluorescence de fond). Extrasynaptique (probablement non spécifique) intensité de la fluorescence devrait normalement pas dépasser 15% de l'intensité de fluorescence de plaque d'extrémité. De même «pas de contrôle de l'anticorps primaire» des sections (incubées uniquement avec anticorps secondaire) devrait apparaître sombre, confirmant que l'anticorps secondaire ne se lie pas de manière non spécifique. La qualité de différents lots d'anticorps secondaires (polyclonaux) peuvent varier de façon marquée d'autres lots d'anticorps secondaires doit être comparé avant d'établir un protocole standard. Le test idéal pour la spécificité de immunofluorescence implique une comparaison directe de sections de souris de type sauvage et sections négative de contrôle de souris tchapeau ne ont pas la (gène KO) protéines d'intérêt. Les références suivantes décrivent la quantification de la plaque d'extrémité coloration fluorescente de musc, rapsyne, dystroglycane, src et AChR 13,14,18,21.
Petite taille des échantillons introduisent des erreurs dans les estimations de l'intensité relative de fluorescence. Plaques d'extrémité individuelles variaient considérablement l'intensité de fluorescence. On peut supposer que cette variabilité entre les plateaux dans un muscle donné reflète la diversité de la structure et de la chance NMJ différences dans la section optique échantillonnée. Cependant, l'augmentation du nombre de plaques d'extrémité de l'échantillon se traduit par une estimation plus stable de l'intensité de fluorescence moyenne (figure 7). Pour estimer la plaque d'extrémité intensité de fluorescence moyenne de 40 à 60 plaques d'extrémité de chaque échantillon de muscle doivent être moyennées.
AChR-AChR FRET
Chaque AChR est un pentamère avec deux sites de liaison pour BGT (situés l'un sur chaque sous-unité alpha). La liaison derouge-BGT et rouge lointain-BGT à ces deux sites donneraient une séparation donneur-accepteur d'environ 9 nm 28-30. Ainsi faible efficacité FRET peut être détecté avant même AChR assemblent en grappes 14. Cependant, l'efficacité de FRET à plateaux murins à peu près doublé après la naissance, conformément à l'incorporation plus efficace des AChR dans un treillis de la membrane postsynaptique 14 serrés. En utilisant le rouge-BGT et rouge lointain-BGT comme donneur FRET et l'accepteur (respectivement), les plateaux de souris âgées de 1-2 mois produites efficacité de FRET moyennes allant de 20 à 37% (tableau 2). FRET efficacité de 20% ou plus sont considérés comme représentant l'emballage serré de AChR 14. L'efficacité de FRET plaque d'extrémité a été légèrement réduite après dénervation 14, et nettement réduite après les souris ont été injectées avec de l'IgG anti-MuSK myasthénie positif gravis 18 patients. Ce sont des conditions dans lesquelles AChR individuels sont moins serrés dans le postsynaptic échafaudage de la membrane par le système MuSK / rapsyne postsynaptique 31.
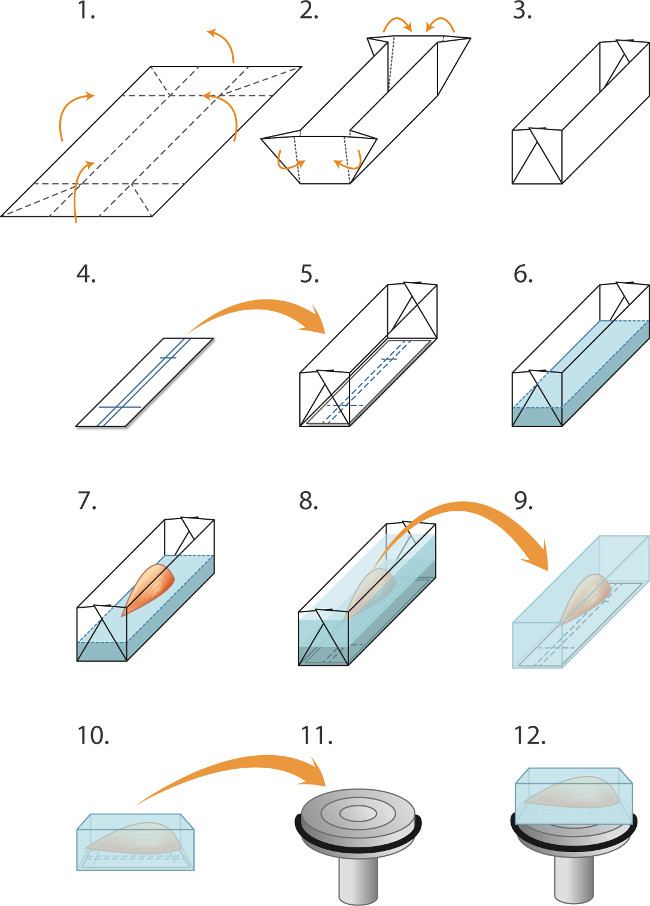
Figure 1. Embedding et les muscles de congélation pour cryosectioning (1-5) Préparation des moules («bateaux») avant la congélation d'un lot de muscles:. (1) Le papier d'aluminium est coupé en rectangles (2,0 x 3,0 cm) et (2). Plié comme inculpé pour créer un moule / bateau (3). (4) Les rectangles de papier immunoblotting de nitrocellulose sont coupés pour se adapter dans le moule et un stylo à bille est utilisé pour exclure les lignes pour orienter le muscle. (5) Le rectangle de nitrocellulose est placée dans le moule. (8.6) Incorporation et le gel des muscles: (6) Cryostat enrobage liquide de matrice est doucement versé dans le moule (au-dessus de la nitrocellulose) à une profondeur de 2 mm. (7) une pince fine sont utilisés pour abaisser le muscle, tendon par son incorporation dans la matrice, en alignement avec l'nitrocelluperdre décisions, (8) la matrice d'intégration supplémentaires est doucement versé dans le moule pour couvrir le muscle, en prenant soin d'éviter de créer des bulles. Le muscle est ensuite incorporé congelé instantanément, dans des tubes scellés et conservés à -80 ° C comme décrit dans le texte. (9-12) Préparation pour cryosectioning: (9) pinces fines sont utilisées pour décoller le moule en aluminium et le bloc congelé est ensuite placé dans la chambre de -20 ° C de cryostat, (10) Le repérage dans la nitrocellulose sont utilisées pour aligner le muscle parallèlement à la face du mandrin (11) de sectionnement longitudinal (12). Une goutte de milieu de cryo-enrobage liquide est utilisé pour fixer le bloc au mandrin refroidi. Forceps Blunt sont utilisés pour manipuler le bloc de muscle congelé. Pour obtenir des sections transversales, le bloc est monté à la place de telle sorte que le muscle est perpendiculaire à la surface du mandrin (non représenté).

Faigure 2. FR images de visage de NMJs du jambier antérieur des souris âgées de C57BL6J femmes 2 mois. Le muscle frais était une congélation instantanée et les sections fixés sur la glissière comme décrit à l'étape 3.2. Maximum images de projection d'intensité de z-piles ont été obtenus comme décrit dans le premier protocole. (A) rouge-BGT coloration révèle une seule plaque motrice constitué de deux ensembles de AChR riches gouttières postjonctionnelle primaires (en vue de face). (B) Synaptophysin coloration avec un anticorps secondaire conjugué à FITC révèle la terminaison nerveuse présynaptique, occupant les gouttières synaptiques primaires. Une partie de l'axone pré-terminale est également visible dans la partie supérieure du panneau. (C & D) Un exemple d'une mauvaise image de NMJ de la qualité d'un jeune souris en bonne santé. La coloration est faible et les limites des spécialisations pré- et post-synaptiques sont floues. Cela a été attribué à des carences en til traitement du tissu et / ou le temps insuffisant autorisé pour l'incubation de l'anticorps primaire. La barre d'échelle dans le panneau D représente 10 um. Se il vous plaît cliquer ici pour voir une version plus grande de cette figure.

Figure 3. Les étapes du traitement des images en Face NMJ (tel que décrit dans le protocole 1). (A & B) images ton MIP continues originales montrent fluorescence rouge-BGT révélant AChR et immunofluorescence vert pour la synaptophysine respectivement. (C) de la CADH coloration après conversion en une image en niveaux de gris de 8 bits et l'utilisation de l'outil polygone pour délimiter le plateau vertébral (mince ligne jaune). (D) ligne de démarcation de plateau vertébral reporté à l'image verte de la synaptophysine. (E) Application d'une commande minimum de seuil d'intensité pour créer une image binaire qui isole supraliminaire fluorescence rouge-BGT (CCRS). La séquence de ImageJ screenshots de l'interface utilisateur se affiche à gauche des images. Image binaire de synaptophysine après l'application d'un seuil séparé (F). (G) Identification de supraliminaires discrète domaines AChR-riches dans le plateau vertébral par l'application de la commande particules Analyser à l'image rouge-BGT binaire. Le ImageJ correspondant (à gauche du panneau (G) montre les données d'entrée nécessaires. La taille minimale des zones de pixels supraliminaires requises devrait être conclu. (H) identification des régions de chevauchement de la synaptophysine binaire et images RACh. Chevauchement est représenté par des pixels blancs . ImageJ screenshots de l'interface utilisateur (ci-dessous groupe H) montrer les étapes pour arriver à l'image de chevauchement binaire. Les valeurs de seuil d'intensité minimum choisis pour chaque fluorescence cHannel doit être saisi dans la fenêtre «colocalisation». La barre d'échelle représente 10 um. Se il vous plaît cliquer ici pour voir une version plus grande de cette figure.

Figure 4. Exemples de sections transversales optiques utilisés pour comparer les intensités de fluorescence relative (protocole 2). Le muscle a été congelé instantanément et les sections fixes sur la diapositive comme décrit à l'étape 3.2. (A & B) Une plaque d'extrémité simple double-étiquetés avec beaucoup -rouge-BGT (AChR; montré ici dans pseudo-bleu) et FITC-anti-rapsyne illustre la co-localisation de ces deux protéines qui interagissent dans la membrane post-synaptique (C & D) Deux plateaux sur les fibres musculaires adjacentes affichage co-localisé AC. RH et de musc. (E & F) Une plaque d'extrémité double-étiquetés pour AChR et -dystroglycan (-DG). Le -DG se étend tout autour du périmètre de la fibre musculaire, mais est enrichi au flasque (barre d'échelle dans F, pour les panneaux AF: 25 pm) (G - I) Isoler une plaque d'extrémité pour la mesure de l'intensité (G) Un champ microscopique typique.. contenant trois plateaux rouge lointain-BGT-colorés (barre d'échelle dans le panneau G: 40 pm (H) Une image agrandie de la plaque d'extrémité coffret (I) La même plaque d'extrémité converti en une image en niveaux de gris de 8 bits et délimité en utilisant le polygone.. . outil ImageJ (mince ligne jaune) intensité de fluorescence moyenne est mesurée à l'intérieur de cette limite. (barre d'échelle pour H & I: 10 pm) Se il vous plaît cliquer ici pour voir une version plus grande de cette figure.

Figure 5. Influence de la qualité d'image dans l'évaluation de la zone synaptique. (A & B) de haute qualité en images de visage d'un NMJ saine d'un 2 mois vieille souris vues sur les canaux de fluorescence anti-synaptophysine rouge-BGT et. (A 'et B ') L'intensité de fluorescence profils correspondants à la ligne tracée en travers de la plaque d'extrémité A et B respectivement. La ligne pointillée rouge horizontale indique le seuil minimal utilisé pour créer l'image binaire. (C & D) à partir d'un plateau vertébral souris âgées. Flasque coloration de la synaptophysine est généralement moins intense. (C '& D ») les profils d'intensité montrent un niveau élevé de extrasynaptique (de base) de fluctuation de fluorescence dans le (FITC) canal de la synaptophysine (retoursol) qui affecte la détermination d'un seuil approprié. Beaucoup de ceci est large spectre autofluorescence de tissu. Barres d'échelle représentent 10 um. Se il vous plaît cliquer ici pour voir une version plus grande de cette figure.

Figure 6. Variabilité des zones synaptiques entre NMJs dans un muscle et entre les souris. (A) Les diagrammes de dispersion montrent la CADH riche superficie totale de plateaux du jambier antérieur de sept anciens naïfs deux mois femelles C57BL / 6J obtenus par auteur NT. Chaque symbole représente une plaque d'extrémité. Chaque barre représente la moyenne ± SD pour plateaux échantillonnés d'une souris. (B) Les diagrammes de dispersion montrant la zone de synaptophysine riches pour les mêmes plateaux. (A ') (B ') de distribution de fréquence pour la région de la synaptophysine riche de plateaux (données regroupées). Se il vous plaît cliquer ici pour voir une version plus grande de cette figure.

Figure 7. Effet de la taille de l'échantillon sur des estimations de l'intensité de fluorescence de plaque d'extrémité. Sections optiques transversales ont été utilisés pour mesurer l'intensité de fluorescence (en unités arbitraires) à 40 60 plateaux du jambier antérieur d'un 2 mois vieille souris en bonne santé. Moyennes cumulatives sont tracées en fonction du nombre de plateaux inclus dans la moyenne. (A) plateau vertébral rouge-BGT intensité de fluorescence obtenue par auteur AV. (B) Anti-synaptophysin intensité d'immunofluorescence pour les mêmes plateaux que dans le panneau A. (C) plateau vertébral rouge lointain-BGT intensité de fluorescence obtenue à partir d'un second échantillon de muscle par auteur NG. (D) Anti-rapsyne intensité d'immunofluorescence des mêmes plateaux que dans le panneau C.
| Étude | Muscle | nombre de souris | Zone AChR | zone de Synaptophysin | Zone de recouvrement |
| moyenne ± SD (um2) | moyenne ± SD (um2) | moyenne ± SD (um2) | |||
| Morsch et al. (2012) 1 | jumeau | 3 | 181 ± 7 | 163 ± 24 | |
| diaphragme | 3 | 130 ± 41 | 102 ± 21 | 76 ± 26 | |
| Morsch et al. (2013) 1 | jambier antérieur | 3 | 166 ± 26 | 117 ± 21 | Dakota du Nord |
| Cheng et al. (2013) 2 | jambier antérieur | 4 | 226 ± 18 | 150 ± 44 | 110 ± 34 |
| Tse (non publié) 2 | jambier antérieur | 7 | 213 ± 27 | 157 ± 23 | 111 ± 11 |
| Une image sur un microscope Zeiss LSM Meta 510, mais avec un gain fixe et décalage niveaux. Seuillage et les zones synaptiques mesures utilisées Metamorph software par MM. | |||||
| 2 imagé sur un microscope Leica DM IRE2 et analysés suivant le protocole actuel par le premier auteur est indiqué, aveugle à groupe de traitement. | |||||
| nd non déterminé. | |||||
Tableau 1. zones synaptiques pour NMJs de deux mois vieille femelle C57BL / 6J (témoins sains)
| Étude | Muscle | FRET efficacité (%) * | Gamme (%) |
| (Moyenne ± SEM) | |||
| Brockhausen et al. (2008) | jambier antérieur | 24 ± 1 | n / a |
| Cole et al., 2010 | jambier antérieur | 26 ± 1 | 22 - 30 |
| Morsch (non publié) | diaphragme | 37 ± 1 | 24-47 |
| Ghazanfari (non publié) | jambier antérieur | 30 ± 1 | 20 - 45 |
| * FRET entre (rayon rouge-BGT et rouge lointain-BGT Förster pour paire de FRET = 51A (Life Technologies). | |||
| données na pas disponibles | |||
Tableau 2. Efficacité pour AChR FRET à partir des souris de jeunes souris adultes
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Les protocoles décrits ici nous ont permis de mesurer de manière fiable et quantifier les changements dans les propriétés de la NMJ toute une gamme de conditions, notamment le vieillissement et les maladies états normaux. Les méthodes décrites pour en Face images NMJ permettront aux chercheurs de comparer le domaine des spécialisations pré- et post-synaptiques et la région synaptique de chevauchement / alignement. Pour comparer l'intensité relative des protéines pré- et post-synaptiques le deuxième protocole, qui utilise des sections optiques transversales, est préféré. Le troisième protocole teste spécifiquement pour les changements dans la proximité de l'emballage de AChR dans la membrane post-synaptique.
contrôles de spécificité sont essentielles en microscopie d'immunofluorescence. Lors de l'utilisation de tout anticorps primaire pour l'immunofluorescence indirecte, il faut d'abord veiller à ce qu'il se lie spécifiquement à la protéine cible dans les sections de muscle. Différents types de traitement des tissus et la fixation peuvent différentielle modifier la spécificité desdes anticorps. Il est important de confirmer que immunofluorescence (pour dire rapsyne) est vraiment concentré avec AChR à la plaque motrice. Sections témoins négatifs doivent également être inspectés pour se assurer que la liaison de l'anticorps est spécifique. Par exemple, le meilleur contrôle négatif pour rapsyne immunofluorescence serait sections de rapsyne souris - / -. Ceux-ci devraient montrer aucune coloration de plaque d'extrémité anti-rapsyne. Fluorescence non spécifique peut également provenir de produits chimiques fluorescents endogènes dans le tissu (autofluorescence) ou de la liaison non spécifique par le conjugué d'anticorps secondaire fluorescent. Cette fluorescence est souvent aggravée par l'aldéhyde fixation. En outre, TRITC-BGT coloration de plaques d'extrémité peut parfois être détecté dans le canal de fluorescence FITC et purge à travers ce fluorescent pourrait être confondue avec FITC immunofluorescence spécifique. Pour se prémunir contre les trois dernières formes de fluorescence non spécifique, chaque lot de diapositives qui sont colorées devrait inclure une 'n«sections de commande de o-anticorps primaire (étapes 3.7 et 4.6). Images de plateaux de ces sections de contrôle doivent être comparés à ceux des diapositives expérimentales pour se assurer que l'immunofluorescence indirecte de NMJs reflète vraiment la liaison de l'anticorps primaire.
Transverse sections confocales sont particulièrement utiles pour évaluer les différences de l'intensité relative des immunocoloration à la synapse. Dans les sections transversales confocaux il est plus facile de juger précis co-localisation des protéines synaptiques. Le profil de plaque d'extrémité en forme de croissant représente juste un échantillon découpe à travers le NMJ en question. Cependant, le fond (extrasynaptique) fluorescence est généralement plus faible par rapport à visage en images z-projection. Ainsi, il peut être plus facile de discriminer «réel» immunocoloration (spécifique) et d'établir un gain confocale fixe et les valeurs de décalage en utilisant des sections transversales optiques 13-15,18. Par exemple, dans un modèle de souris de la myasthénie grave (where flasque AChR coloration est nettement réduite) plateaux ont été clairement définis dans les sections transversales optiques 18,21. Les différences dans l'intensité moyenne de fluorescence à la jonction neuromusculaire sont susceptibles de refléter la densité modifiée de la protéine cible à l'intérieur de la spécialisation synaptique. Une mise en garde est que, dans certaines situations, un changement structurel dans la protéine ou une occlusion de la liaison par protéines voisins anticorps cible pourrait expliquer l'intensité de coloration altérée.
La conception d'expériences nécessite une certaine considération. Dans de nombreux cas l'expérience viserait à tester l'impact d'un transgène, knockdown de gènes ou de l'état de la maladie de la taille de la NMJ. Le groupe échantillon expérimental pourrait alors être comparé aux jeunes (de type sauvage) des souris saines du même sexe et le patrimoine génétique. Les valeurs de référence pour le domaine de la plaque d'extrémité synaptophysine, AChR et le chevauchement synaptique pour plusieurs muscles sont donnés dans le tableau 1. Taille de l'échantillon dépendra du degré d'animauxvariation -À-animale au sein des groupes de traitement et de la taille d'effet (différence des moyennes pour la expérimentale contre des groupes de contrôle par l'écart-type). Lorsque l'analyse est limitée à des images de bonne qualité un certain degré de cohérence a été trouvé dans l'échantillon signifie pour les zones des plateaux vertébraux entre deux mois des souris âgées de C57BL6J sains de sexe féminin (figure 6A et B). Ainsi, il était possible de démontrer importantes 30-40% de réduction dans la zone synaptique chez la souris injectées avec des IgG anti-MuSK myasthénie positif gravis patients, par rapport à des contrôles avec une taille de trois souris 17,20,32 de l'échantillon. Souris âgées affichent une plus grande variation d'animal à animal dans les paramètres des plateaux vertébraux que les jeunes souris 22. Par conséquent les expériences impliquant des souris âgées peuvent nécessiter des échantillons plus importants.
Si la principale préoccupation est de mesurer la taille de la plaque d'extrémité de face alors le gain et le décalage de niveau devrait être optimisé pour chaque individu NMJ. IndivNMJs idual peuvent varier considérablement dans la luminosité de AChR et synaptophysine coloration, en particulier lorsque états pathologiques sont examinés. En outre, l'intensité de la fluorescence extra-synaptiques (non spécifique) est souvent plus élevée et plus variable dans les muscles des animaux âgés, par rapport à ceux de jeunes animaux en bonne santé (Figure 5C et D). Le 1-256 gris devrait être pleinement exploité afin de maximiser l'information tonale qui sera retenue dans les images finales. Il se agira de régler le gain et l'offset niveaux pour chaque NMJ pour lequel un z-pile doit être collecté. Figure 5D montre un exemple d'une image NMJ où l'information tonale pourrait être critique dans la définition des limites de la zone de pré et post- spécialisations synaptiques.
Les mesures des zones synaptiques peuvent être appliqués à différentes préparations musculaires et des expériences. La plupart de nos mesures de zones synaptiques ont employé cryosections longitudinales de pression muscles congelés. La congélation du muscle avant la fixation maintient l'antigénicité d'une large gamme de protéines. Lorsque compatible avec l'antigène, la fixation de paraformaldéhyde et l'infiltration de saccharose avant cryosectioning (étape 2.1) peut fournir une meilleure préservation de la structure NMJ. Conservation structurelle optimale peut être obtenue par perfusion avec du paraformaldehyde cardiaque. Artefacts de congélation et de sectionnement peuvent être complètement évités par plateaux d'étiquetage sur la surface du muscle et d'imagerie NMJs intacts sur fascicules taquiné du muscle fixe 21. Indépendamment de la préparation, les procédures d'échantillonnage, de l'imagerie et de la zone quantification restent inchangés (protocole étapes 4-5). L'application uniforme des protocoles d'échantillonnage, imagerie et d'analyse aveugles (en utilisant différents opérateurs, différents échantillons de souris et des moments différents), peut entraîner des valeurs moyennes assez reproductibles (comparer Cheng et al. Et Tse résultats du tableau 1).
"> Plaques de fond ont été décrits comme devenant« fragmentée »dans une variété d'états pathologiques. Par exemple, dans le vieillissement des muscles de souris, la dégénérescence sporadiques d'une fibre musculaire (suivies par sa régénération) entraîné dans le remodelage de la bretzel comme flasque AChR plaque pour former plusieurs petits amas RACh 6. Dans les souris injectées avec de l'IgG anti-MuSK de myasthenia gravis patients, la fragmentation de la plaque d'extrémité est un peu différente. La plaque d'extrémité AChR bretzel largement dispersé, laissant derrière une constellation de petits (<4 m 2) microagrégats RACh ' '20,21. Ces deux exemples soulignent la nécessité de comparer les distributions de taille pour les clusters RACh à plateaux de contrôle par rapport à des animaux de laboratoire 21.D'autres procédés ont été signalés pour évaluer région synaptique ou à l'intensité de coloration NMJ. plaques motrices peuvent parfois être plié de sorte que les images z-projection à deux dimensions utilisées ici pourraient sous-estimer synAPTIC domaines. Reconstructions confocale en trois dimensions pourraient fournir des mesures plus précises si la zone synaptique absolue doit être défini 33. Un avantage clé du protocole z-projection décrit ici, cependant, est sa relative simplicité, ce qui a permis un grand nombre de plateaux à mesurer à partir de plusieurs groupes de traitement et une identification fiable des changements potentiels. Le protocole pour comparer les intensités de coloration plateaux vertébraux peut être adapté pour étudier les changements dans les niveaux de nombreuses protéines synaptiques différentes. La méthode est limitée, cependant, par l'exigence que tous les échantillons sont traitées pour immunofluorescence puis imagé au cours de la même session confocale. Une étude récente de Yampolsky et al. 5 décrit une méthode pour mesurer la densité flasque AChR qui pourraient aider à surmonter cette limitation. Dans cette étude, les mêmes champs de plateaux ont été imagées à plusieurs réglages de puissance laser différents. La pente de la relation entre la puissance laser et rhodamine-BGT intensité de fluorescence a été utilisé pour évaluer les changements relatifs à la densité AChR à plateaux chez la souris différente 5. Cette méthode pourrait être utile pour comparer l'intensité AChR dans des échantillons imagés à différents moments au cours d'une étude prolongée.
AChR-AChR FRET fournit des informations spécifiques et complémentaires sur l'organisation de AChR des plateaux vertébraux. Electron autoradiographie microscopique en utilisant 125. I-α-BGT a montré AChR à emballer hermétiquement avec une densité plane de 10 4 m -2 immédiatement sous chaque site présynaptique de la libération de transmetteur, tandis adjacentes replis de la membrane contiennent des densités beaucoup plus faibles de 34 AChR. AChR-AChR FRET rend relativement facile à détecter (sous-microscopiques) des changements dans AChR emballage. Une réduction de l'efficacité de FRET reflète une redistribution de la sous-microscopique de AChR dans la membrane post-synaptique qui pourraient ne pas être détectées par un changement de l'intensité moyenne de fluorescence de BGT. Multiple facteurs peuvent provoquer une modification de l'efficacité de FRET. Il se agit notamment de l'espacement donneur-accepteur et orientations relatives, ainsi que l'environnement moléculaire 35,36. Une réduction de l'efficacité de FRET plaque d'extrémité pourrait éventuellement résulter d'une modification de la géométrie du réseau AChR. Cependant, le plus probable, il serait due à une réduction du pourcentage de AChR qui sont emballés dans la nano-échelle moléculaire en treillis 14 postsynaptique.
La perte de la connexion entre les neurones moteurs et les fibres musculaires semble être la cause immédiate de la faiblesse musculaire dans la maladie du neurone moteur et dans le vieillissement sédentaire 9,22,23. Méthodes et paramètres partagés pour NMJs mesure devraient rendre plus facile pour différents groupes de recherche de comparer et de contraste conclusions publié. Le partage des protocoles détaillés (et améliorations futures sur eux) peut aider à accélérer les progrès dans la compréhension des mécanismes de l'entretien NMJ et comment il peut être affecté dans les états pathologiques.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| Scanning confocal microscope | Leica | DM 2000 with TCS SP2 system | Most scanning confocal microscopes should be suitable. |
| Zeiss | LSM 510 Meta | ||
| Leica | SPE-II | ||
| Alexa555-a-bungarotoxin (red-BGT) | Life technologies | B35451 | Used for labelling AChRs |
| Alexa647-α-bungarotoxin (far-red-BGT) | Life technologies | B35450 | Far red fluorescence: barely visible through the eyepiece |
| rabbit anti-synaptophysin | Life technologies | 18-0130 | Different batches of primary antibody differ in effective working dilution |
| FITC-anti-rapsyn mab1234 | Milipore | FCMAB134F | Monoclonal antibody conjugated to FITC |
| FITC-donkey anti-rabbit IgG | Jackson | 711-095-152 | Polyclonal secondary antibodies can vary in quality according to source and batch |
| Optimal Cutting Temperature compound (O.T.C.) | ProSciTech | IA018 | Cryostat embedding matrix for freezing muscles |
| DABCO | Sigma | 10981 | Mounting medium that slows photobleaching of fluorophores |
References
- Schmidt, N., et al. Neuregulin/ErbB regulate neuromuscular junction development by phosphorylation of α-dystrobrevin. J Cell Biol. 195, 1171-1184 (2011).
- Amenta, A. R., et al. Biglycan is an extracellular MuSK binding protein important for synapse stability. J Neurosci. 32, 2324-2334 (2012).
- Samuel, M. A., Valdez, G., Tapia, J. C., Lichtman, J. W., Sanes, J. R. Agrin and Synaptic Laminin Are Required to Maintain Adult Neuromuscular Junctions. PLOS ONE. 7, e46663 (2012).
- Valdez, G., et al. Attenuation of age-related changes in mouse neuromuscular synapses by caloric restriction and exercise. Proc Natl Acad Sci (USA). 107, 14863-14868 (2010).
- Yampolsky, P., Pacifici, P. G., Witzemann, V. Differential muscle-driven synaptic remodeling in the neuromuscular junction after denervation). Eur J Neurosci. 31, 646-658 (2010).
- Li, Y., Lee, Y., Thompson, W. J. Changes in Aging Mouse Neuromuscular Junctions Are Explained by Degeneration and Regeneration of Muscle Fiber Segments at the Synapse. J Neurosci. 31, 14910-14919 (2011).
- Zhu, H., Bhattacharyya, B. J., Lin, H., Gomez, C. M. Skeletal muscle IP3R1 receptors amplify physiological and pathological synaptic calcium signals. J Neurosci. 31, 15269-15283 (2011).
- Valdez, G., Tapia, J. C., Lichtman, J. W., Fox, M. A., Sanes, J. R. Shared resistance to aging and ALS in neuromuscular junctions of specific muscles. PLoS ONE. 7, e34640 (2012).
- Perez-Garcia, M. J., Burden, S. J. Increasing MuSK Activity Delays Denervation and Improves Motor Function in ALS Mice. Cell reports. 2, 1-6 (2012).
- Klooster, R., et al. Muscle-specific kinase myasthenia gravis IgG4 autoantibodies cause severe neuromuscular junction dysfunction in mice. Brain. 135, 1081-1101 (2012).
- Pratt, S. J., Shah, S. B., Ward, C. W., Inacio, M. P., Stains, J. P., Lovering, R. M. Effects of in vivo injury on the neuromuscular junction in healthy and dystrophic muscles. J Physiol. 591, 559-570 (2013).
- Landis, S. C., et al. A call for transparent reporting to optimize the predictive value of preclinical research. Nature. 490, 187-191 (2012).
- Gervásio, O. L., Phillips, W. D. Increased ratio of rapsyn to ACh receptor stabilizes postsynaptic receptors at the mouse neuromuscular synapse. J Physiol. 562, 673-685 (2005).
- Gervásio, O. L., Armson, P. F., Phillips, W. D. Developmental increase in the amount of rapsyn per acetylcholine receptor promotes postsynaptic receptor packing and stability. Dev Biol. 305, 262-275 (2007).
- Brockhausen, J., Cole, R. N., Gervásio, O. L., Ngo, S. T., Noakes, P. G., Phillips, W. D. Neural agrin increases postsynaptic ACh receptor packing by elevating rapsyn protein at the mouse neuromuscular synapse. Dev Neurobiol. 68, 1153-1169 (2008).
- Cole, R. N., Reddel, S. W., Gervásio, O. L., Phillips, W. D. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol. 63, 782-789 (2008).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Muscle Specific Kinase autoantibodies cause synaptic failure through progressive wastage of postsynaptic acetylcholine receptors. Exp Neurol. 237, 237-286 (2012).
- Cole, R. N., Ghazanfari, N., Ngo, S. T., Gervasio, O. L., Reddel, S. W., Phillips, W. D. Patient autoantibodies deplete postsynaptic Muscle Specific Kinase leading to disassembly of the ACh receptor scaffold and myasthenia gravis in mice. J Physiol. 588, 3217-3229 (2010).
- Viegas, S., et al. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp Neurol. 234, 506-512 (2012).
- Morsch, M., Reddel, S. W., Ghazanfari, N., Toyka, K. V., Phillips, W. D. Pyridostigmine but not 3,4-diaminopyridine exacerbates ACh receptor loss and myasthenia induced in mice by Muscle Specific Kinase autoantibody. J Physiol. 591, 2747-2762 (2013).
- Ghazanfari, N., Morsch, M., Reddel, S. W., Liang, S. X., Phillips, W. D. Muscle Specific Kinase autoantibodies suppress the MuSK pathway and ACh receptor retention at the mouse neuromuscular junction. J Physiol. 592, 2881-2897 (2014).
- Cheng, A., Morsch, M., Murata, Y., Ghazanfari, N., Reddel, S. W., Phillips, W. D. Sequence of age-associated changes to the mouse neuromuscular junction and the protective effects of voluntary exercise. PLoS One. 8, e67970 (2013).
- Schaefer, A. M., Sanes, J. R., Lichtman, J. W. A compensatory subpopulation of motor neurons in a mouse model of amyotrophic lateral sclerosis. J Comp Neurol. 490, 209-219 (2005).
- Kilkenny, C., Browne, W. J., Cuthill, I. C., Emerson, M., Altman, D. G. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLos Biol. 8, e1000412 (2010).
- Shimizu, S. Routes of Administration. The Laboratory Mouse. Hedrich, H. J., Bullock, G. , Elsevier. (2004).
- Chiasson, R. B. Laboratory anatomy of the white rat. , Brown. Dubuque, Iowa. (1988).
- Gage, G. J., Kipke, D. R., Shain, W. Whole Animal Perfusion Fixation for Rodents. J. Vis. Exp. (65), e3564 (2012).
- Mitra, A. K., Stroud McCarthy, M. P., M, R. Three-dimensional structure of the nicotinic acetylcholine receptor and location of the major associated 43-kD cytoskeletal protein, determined at 22A by low dose electron microscopy and x-ray diffraction to 12.5A. J Cell Biol. 109, 755-774 (1989).
- Paas, Y., et al. Electron microscopic evidence for nucleation and growth of 3D acetylcholine receptor microcrystals in structured lipid-detergent matrices. Proc. Natl Acad. Sci. (USA). 100, 11309-11314 (2003).
- Samson, A. O., Scherf, T., Eisenstein, M., Chill, J. H., Anglister, J. The mechanism for acetylhcoline receptor inhibition by α-neurotoxins and species-specific resistance to α-bungarotoxin revealed by NMR). Neuron. 35, 319-332 (2002).
- Ghazanfari, N., et al. Muscle Specific Kinase: Organiser of synaptic membrane domains. Int J Biochem Cell Biol. 43, 295-298 (2011).
- Ghazanfari, N., Morsch, M., Tse, N., Reddel, S. W., Phillips, W. D. Effects of the β2-adrenoceptor agonist, albuterol, in a mouse model of anti-MuSK myasthenia gravis. PLoS ONE. 9, e87840 (2014).
- Prakash, Y. S., Miller, S. M., Huang, M., Sieck, G. C. Morphology of diaphragm neuromuscular junctions on different fibre types. J Neurocytol. 25, 88-100 (1996).
- Salpeter, M. M., Harris, R. Distribution and turnover rate of acetylcholine receptors throughout the junction folds at a vertebrate neuromuscular junction. J Cell Biol. 96, 1781-1785 (1983).
- Soper, S. A., Nutter, H. L., Keller, R. A., Davis, L. M., Shera, E. B. The photophysical constants of several fluorescent dyes pertaining to ultrasensitive fluorescence spectroscopy. Photochem Photobiol. 57, 972-977 (1993).
- Panchuk-Voloshina, N., et al. Alexa dyes, a series of new fluorescent dyes that yield exceptionally bright, photostable conjugates. J Histochem Cytochem. 47, 1179-1188 (1999).
