

Summary
Her rapporterer vi dobbelt mærkning af neurale crest celler og blodkar ved hjælp af chickGFP neuralrør intraspecies podning kombineret med intra-vaskulære DiI injektion. Denne eksperimentelle teknik giver os mulighed for samtidig at visualisere og studere udviklingen af det NCC-afledte (enteriske) nervesystem og det vaskulære system under organogenese.
Abstract
Alle udviklende organer skal tilsluttes både nervesystemet (til sensorisk og motorisk kontrol) samt det vaskulære system (til gasudveksling, væske- og næringsstofforsyning). Derfor udvikler både nerve- og vaskulære systemer sig side om side og deler slående ligheder i deres forgrenede arkitektur. Her rapporterer vi embryonale manipulationer, der giver os mulighed for at studere den samtidige udvikling af neuralt crest-afledt nervevæv (i dette tilfælde det enteriske nervesystem) og det vaskulære system. Dette opnås ved at generere kylling kimærer via transplantation af diskrete segmenter af neuralrøret, og tilhørende neurale crest, kombineret med vaskulære DiI injektion i samme embryo. Vores metode bruger transgene chickGFP embryoner til intraspecies podning, hvilket gør transplantationsteknikken mere kraftfuld end den klassiske vagtel-chick interspecies podning protokol, der anvendes med stor effekt siden 1970'erne. ChickGFP-chickintraspecies podning letter billeddannelse af transplanterede celler og deres fremskrivninger i intakt væv, og eliminerer enhver potentiel bias i celleudvikling i forbindelse med artsforskelle. Denne metode udnytter fuldt ud avianembryoets let adgang (sammenlignet med andre hvirveldyrembryoner) til at studere den fælles udvikling af det enteriske nervesystem og det vaskulære system.
Introduction
Kyllingembryoet er en uvurderlig modelorganisme i hvirveldyrudviklingsbiologi, ikke mindst fordi dets udvikling i ovo tillader eksperimentelle manipulationer, der ellers er umulige at udføre i hvirveldyr, der udvikler sig i livmoderen. Denne tilgængelighed og lethed af manipulationer har ført til, at kyllingembryoet spiller nøgleroller i mange skelsættende opdagelser inden for udviklingsbiologi. Blandt de mest kraftfulde teknikker har været brugen af vagtler-chick kimære embryoner til at studere celle skæbne, en metode banebrydende af professor Nicole Le Douarin i 1970'erne1-3. Især har vagtler-chick kimærer været særligt nyttige til genetisk mærke og følge stærkt vandrende neurale crest celle (NCC) befolkninger i den tidlige udvikling. NCC er en multipotent population af migrerende celler, der opstår i ryghvirvelserotallet i udkanten af neuralrøret, der giver anledning til en bred vifte af celletyper i hele hvirveldyrembryoet. Disse omfatter kraniofacial strukturer (brusk, knogler, muskler), neuroner og glia (i de sensoriske og autonome nervesystemer), melanocytter og en delpopulation af celler i det endokrine system2,4,5. En af de vigtigste faktorer, der påvirker NCC skæbne er deres oprindelige placering langs den forreste-bageste akse af neuralrøret. For eksempel opstår enteriske NCC, som giver anledning til neuronerne og glia i det enteriske nervesystem (ENS), fra to diskrete underpopulationer: den første placeret i vagalområdet (kaudale hindbrain) og den anden i neuralrørets sakrale region6-13. Inter- eller intraartstransplantation af de tilsvarende regioner i neuralrøret har været de foretrukne teknikker til permanent at mærke disse celler og efterfølgende tillade sporing fra deres fødsel i udkanten af neuralrøret til deres endelige destinationer inden for fordøjelseskanalen6,7,10.
En anden embryonal manipulation lettere at udføre i kylling, sammenlignet med andre dyremodeller, er den vitale mærkning af det vaskulære system. Faktisk, som kyllingembryoet udvikler sig, lægger det oven på et ekstra embryonalt vaskulært netværk, der cirkulerer ilt og næringsstoffer fra æggeblommen. Dette tilgængelige vaskulære netværk, der er placeret på æggeblommens overflade, kan bruges som en gateway til at mærke embryoets udviklende vaskulære system under organogenese12,14-17. Intravaskulær injektion af forskellige farvestoffer, såsom det lipofile farvestof DiI, gør det muligt at afgrænse/plette alle de lysarmede beholdere i det spirende vaskulære netværk.
Fordi udviklende organer skal tilsluttes både nervesystemet (for sensorisk og motorisk kontrol) samt det vaskulære system (til gasudveksling, væske og næringsstofforsyning), udvikler de to netværk sig sammen med hinanden og deler slående ligheder i deres forgrenede arkitektur18-20. Her rapporterer vi embryonale manipulationer, der giver os mulighed for at studere den samtidige udvikling af den NCC-afledte ENS sammen med det vaskulære system under organogenese. Dette opnås ved at generere kylling kimærer via transplantation af diskrete segmenter af neuralrøret, herunder neurale crest, kombineret med vaskulære DiI injektion. Som et fremskridt fra vagtel-kylling kimærer, vores metode bruger transgene GFP chick embryoner til intraspecies podning, hvilket gør transplantationen teknikken mere kraftfuld, i form af billeddannelse celler og deres fremskrivninger, og fjerne eventuelle bias forbundet med arter forskelle.
Protocol
1. Forberedelse af mikro-skalpel til neuralrørsblationer
- Form en mikro-skalpel fra en kommercielt tilgængelig stålsypind.
- Flad først nålen på begge sider ved hjælp af et slibehjul monteret på en drevet bænkkværn.
- Begynd at forme skalpellen, først på en grov klasse Arkansas sten ved hjælp af en kontrolleret cirkulær bevægelse, i alternative retninger, på begge sider af nålen.
- Fortsæt de samme skarphedsbevægelser på en ekstra fin kvalitet Arkansas sten for at forme en ultrafin mikro-skalpel, med en veldefineret skærkant(Figur 1A, B).
BEMÆRK: Alternativer til mikro-skalpel kan være elektrolitically skærpet nåle, kommercielt tilgængelige wolfram nåle, eller trukket glas nåle.
2. Inkuber vilde type og GFP æg til den ønskede fase
- Opbevar befrugtede kyllingæg og transgene GFP kyllingæg i en afkølet inkubator ved 14 - 15 °C før inkubation, da udviklingen standses ved denne temperatur. Opbevar æg i et par dage, op til en uge.
- For at påbegynde udviklingen skal du placere vilde type- og GFP-æg på en bakke vandret og inkubere samtidigt i en befugtet (58 - 60%) inkubator ved 37,5 ºC, således at embryoner er på matchende stadier for neuralrørstransplantation.
- At opnå embryoner på 10 - 12 somite udviklingsstadiet til udførelse af vagus neuralrørstransplantation, inkubere æg i 1,5 dage (33 - 38 timer) og scenefostre i henhold til udviklingstabellerne i Hamburger og Hamilton21.
3. Forbered æg til vindues- og podning
- Flyt et æg ad gangen til en specialfremstillet æggeholder til vinduesvinduer. Lav et lille hul i æggeskallen ved gentagne gange at trykke med lige saks på den øverste overflade af den spidse ende af ægget.
- 2- 3 ml albumin fjernes fra ægget med en 181/2 G hypodermisk nål og 5 ml sprøjte. Fjernelse af albumin sænker æggeblommen i ægget og letter efterfølgende vindue uden at forårsage nogen skade på embryoet.
- Kassér albumin. Forsegl hullet med en lille strimmel af klar tape skåret i størrelse med fin saks.
- Brug buet saks, tryk på et andet hul i den øverste overflade af æggeskallen. Sæt spidsen af saksen i hullet, og hold saksen parallelt med bænken, arbejd i en cirkulær bevægelse for at skære et ~ 2 cm diameter vindue oven på skallen.
- Hold saksen i stationær position og roter ægget. Kassér den fjernede disk af æggeskal. På E1.5 er embryoet genkendeligt som en mørkere gul disk oven på æggeblommen.
- Fjern eventuelle skalrester, der er faldet i ægget ved hjælp af pincet. Kassér eventuelle ubefrugtede æg (identificeret ved en lille hvid plet på toppen på den ellers lysegule æggeblomme).
4. Forbered host embryoet til at modtage podet væv
- Juster stereomikroskopet til øjenhøjde, og optimer orienteringen af gåsehudlyskilden for at belyse embryoet tilstrækkeligt uden at forårsage refleksioner.
- For at visualisere embryoet korrekt, injicere en lille mængde indisk blæk under midten af den mørkere gule disk, ved hjælp af en mund rør og en trukket glas mikro-pipette(Figur 1C, Oii).
- Blækket fremstilles 50:50 med PBS, der indeholder Penicillin/Streptomycin ved 100 μg/ml endelig koncentration. Sæt mikropipetten gennem æggeblommemembranen uden for blastodermens omkreds, og vinkles derefter forsigtigt spidsen direkte under embryoet.
- Levere blæk under embryoet ved at blæse på mundrøret. Hvis mundpipetter ikke er tilladt, skal du i stedet bruge en 1 ml sprøjte. Pas på ikke at indføre luftbobler under embryoet, hvilket kan føre til forurening, og fjern derefter forsigtigt glasmikropipetten. Dette er et ømtåleligt skridt, der kan føre til embryoets død, hvis det ikke gøres med præcision.
- Stage embryoet med henvisning til Hamburger og Hamilton21 og registrere scenen i et laboratorium bog.
- Brug en specialfremstillet mikro-skalpel (eller en fin wolfram nål) monteret på en nål holder, lave en meget lille flænge i vitelline membranen, ved siden af det område, hvor mikro-kirurgi vil blive udført.
- Påfør forsigtigt 2 - 3 dråber PBS over membranen (ved hjælp af en glasmikropipette og mundrøret) for at skabe plads mellem embryoet og membranen. Skær et større vindue i membranen for at afsløre hele regionen, hvor mikrokirurgi vil finde sted.
- Fjern neuralrørsområdet af interesse ved hjælp af mikro-skalpel, begyndende med rostrale og kaudale tværgående snit på tværs af hele det dorsale neuralrør (på niveauet af somite 1 til 7 i videoen).
- Skær bilateralt mellem neuralrøret og somitterne for at adskille neuralrøret fra det omgivende væv uden at beskadige somitterne.
- Meget forsigtigt adskille neuralrør fra den underliggende notochord, som bør forblive intakt. Bemærk, at vellykket neuralrør excision vil efterlade alle omkringliggende væv helt intakt (Figur 2).
- Fjern det fjernede neuralrør ved at suge det ind i en glasmikropipette, og kassér derefter.
- Optag niveauet af neuralrørsablation i en laboratoriebog. Værtsfostret er nu klar til at modtage donor neuralrøret.
5. Forbered donortransplantationsvævet
- Vælg et fasematchet GFP-foster med vinduer ved at se under et fluorescerende stereomikroskop med FITC-filter. GFP fluorescens gør det meget nemt at visualisere somitterne og iscenesætte embryoet.
- Når et fasematchet foster er identificeret, skal du fjerne embryoet fra ægget ved at lave 4 snit, med Pascheff-Wolff-forårssaks(Figur 1C, l) i rektangelform omkring embryoet og derefter forsigtigt samle det op med en embryoske.
- Placer embryoet i et firkantet urglas med en sylgard polymerbase. Ryst forsigtigt embryoet med Dumont #5 pincet for at fjerne enhver vedhæftet æggeblomme. Fjern vitellinemembranen, og fastgør embryoet på polymerbasen ved hjælp af rustfrie minutienstifter (Figur 1C).
- Brug springsaksen til at lave 4 snit i rektangulær form omkring neuralrøret og de omkringliggende somites i samme region, der er blevet fjernet fra værtsembryoet.
- Ved hjælp af en plastoverførsel pipette, overføre neuralrør og somite væv fra donor GFP embryo i et ur glas, der indeholder 0,2% pancreatin i Pen / Strep PBS.
- Lad enzymatisk fordøjelse fortsætte i 10 minutter på RT for at hjælpe med at adskille vævene. Efter inkubation i enzym, skal du bruge rustfrit minutien stifter monteret på et håndtag til manuelt at adskille neuralrør fra alle tilstødende væv.
- Ved hjælp af en mikropipette af glas overføres det dissociated neuralrør til et andet urglas, der indeholder DMEM + 10% serum(f.eks.ged, hest eller fosterkalv) på is, for at skylle den overskydende pancreatin og stoppe enzymatisk fordøjelse. Efter 5 min. er det dissekerede neuralrør klar til at blive podet ortopisk ind i kyllingeværten (Figur 2 og S1).
6. Graft vævet
- Ved hjælp af en glasmikropipette skal du forsigtigt overføre det dissekerede neuralrør fra urglasset til værtsembryoet. Placer neuralrøret i den korrekte forreste-bageste orientering og skub forsigtigt explanten ved siden af kyllingeværtens punkterede region ved hjælp af mikro-skalpel. Efterlad et lille fragment af ectoderm fastgjort til, eller ved at skære et lille nick i, den dorsale overflade for at identificere retningen af neuralrøret.
- Hvis det er nødvendigt, skal du bruge mikro-skalpel til at trimme explant til den nøjagtige størrelse af det punkterede område.
- Før forsigtigt neuralrøret ind i det ablerede område og placer det på en sådan måde, at den dorsale side er korrekt orienteret. Brug en glasmikropipette, monteret på et mundrør, til at fjerne PBS og/eller væske omkring graften. Dette hjælper donor og vært væv til at overholde og graft at blive etableret.
- Luk hele vinduet med 24 mm bredt klart tape for at forhindre dehydrering og forurening.
- Mærk det kimære embryo ved at markere med en blyant på æggeskallen og registrere dens nummer i laboratoriebogen. Returner ægget til inkubatoren for yderligere udvikling.
7. Injicere diI i blodkarrene af Host Embryo
- På det ønskede eksperimentelle tidspunkt (her, 3 - 10 dage senere), skal du hente det kimære embryo fra inkubatoren og fjerne det klare bånd ved hjælp af lige saks for at få adgang til embryoet i ægget.
- Hvis det er nødvendigt, forstørre vinduet i skallen ved hjælp af saksen. Pas på ikke at beskadige chorioallantoic membranen, hvis den er fastgjort til skallen, hvilket ville resultere i blødning og bringe blodkarmærkningen i fare.
- Vælg en tilgængelig vene på æggeblommen, og sørg for, at blodgennemstrømningen er rettet mod embryoet. Vælg et forgreningspunkt for en af vitellin venerne (Figur 3B, C).
BEMÆRK: På E6.5 - E7.5, chorioallantoic membranen kan være nødvendigt at forsigtigt flyttes til side med pincet for at få adgang til æggeblomme vener. Efter E8.5 er den eneste mulighed at injicere i en af de chorioallantoic membran vener, da chorioallantoic membranen på dette tidspunkt fuldt ud dækker embryoet. - Fjern vitellinmembranen over det valgte injektionspunkt ved hjælp af to Dumont-#5 pincet ved at rive i modsatte retninger.
- Bryd en trukket glasnål ved hjælp af en Dumont-#5 og juster dens diameter til den omtrentlige størrelse af venen før pålæsning med CellTracker CM-DiI. DiI-stamopløsningen fremstilles ved 40 μg/μl i DMSO og opbevares ved -20 °C. Arbejdsopløsningen fremstilles i 0,3 M saccharose/PBS i en koncentration på 4 μg/μl.
- Aspirat mellem 5 - 10 μl diI i 0,3 M saccharose/PBS i nålen ved hjælp af sugning med et mundrør. Ældre embryoner kan kræve op til 25 μl eller mere. Fra E8.5 har embryoner større, mere muskuløse vener, som muligvis skal holdes på plads med en Dumont-#5, før de stikkes med den DiI-belastede glasnål.
- Sæt hurtigt nålen i venen og blæs støt med mundrøret for at gøre det muligt for DiI at slutte sig langsomt til blodgennemstrømningen uden at danne en blodprop. Alternativt kan du bruge en trykinjektor til diI-levering.
8. Høst embryoner til sektionsopdejning eller fuldkornsundersøgelse
- For at bevare så meget DII i embryoet som muligt, høste embryoet umiddelbart efter injektionen ved at scooping det på en perforeret ske og skære blodkarrene og bindevæv med en par lige saks, for at befri embryoet fra æggeblommen.
- Fjern eventuelle løse membraner og dissekere de organer af interesse(dvs.lungerne og fordøjelseskanalen i denne tutorial), idet stor omhu for ikke at komprimere vævet, hvilket skaber diffusion af DiI. Straks fastsætte væv ved nedsænkning i 4% PFA for 1 - 2 timer på RT.
- Vævet skylles i 5 minutter i PBS og derefter 15 min i PBS, der indeholder 5 μg/ml DAPI. Monter prøverne på en bro mikroskop dias for hele mount undersøgelse eller integrere dem til kryo-sektion.
Representative Results
Figur 1 viser typiske instrumenter, der er nødvendige for at udføre mikrokirurgisk isolering og transplantation af neuralrøret. Figur 2 viser transplantationsproceduren. Efter transplantation embryoner screenes for transplantation succes. Dette indebærer undersøgelse af embryoet under et stereofluorescensmikroskop, typisk morgenen efter mikrokirurgi, for tilstedeværelsen af graft-afledt (GFP+) NCC. Hvis transplantation har været en succes, så GFP + NCC kan observeres i nærheden af neuralrøret og i tidlig migration veje, der fører mod forgrunden. Hvis proceduren ikke har været vellykket, vil GFP + NCC ikke blive observeret uden for neuralrøret, eller hvis de er til stede i værten, kan de være i mindre antal. Disse mislykkede embryoner kasseres. Typisk udføres 5-8 neuralrørstransplantationer på en dag, og af disse er 80% vellykkede. Årsagerne til mislykket neuralrørstransplantation omfatter fosterets død på grund af vævsskader, der er opstået under mikrokirurgi, eller svigt af neuralrøret til at integrere sig i værtsembryoet. Sidstnævnte kan skyldes dårlig placering af neuralrøret i værten eller fra et neuralrør af dårlig kvalitet på grund af dårlig dissektionsteknik eller fra overdreven eksponering for dissociationsenzym. Det indledende screeningstrin samt lignende senere undersøgelser for GFP+-celler er nyttigt, da det betyder, at tid og ressourcer ikke spildes ved at udføre forsøg på embryoner, der ikke har GFP-mærket NCC i tarmen.
Figur 3 viser proceduren for diI-injektion af blodkarrene. DiI-injektionsteknikkens effektivitet/succes afhænger af: For det første at skære injektionsnålen til den optimale diameter for den målrettede vene, for det andet en præcis bevægelse, når du indsætter nålen i venen (for ikke at gennembore gennem den anden side), og for det tredje undgå, at nålen bliver tilsluttet under injektionen ved at blæse i konstant hastighed. Hvis nogen af disse tre parametre gøres forkert, vil embryoet bløde ud eller vil have brug for flere timer til at komme sig, før et andet forsøg er gjort som blødning gør det næsten umuligt at re-injicere straks. Vellykkede embryoner bør udvælges straks ved at se under et stereofluorescensmikroskop og skal dissekeres hurtigt. I vellykkede embryoner er diI-mærkede blodkar til stede i hele embryoet (figur 3C,D), herunder kapillære senge (figur 3D).
Ved høst af embryoner og undersøgelse af vævssektioner eller fuldkornsgastrointestinale skrifter typiske resultater afslører GFP+ NCC inden for den primitive ENS og den fine struktur af de DiI-mærkede tarmblodbeholdernetværk (Figur 4) Fuldkornspræparater kan undersøges ved hjælp af konfokal mikroskopi, hvor billedstakke producerer tredimensionelle (3D) rekonstruktioner, der viser sammenhængen mellem de fine fremskrivninger af GFP+ ENS-celler og Det DiI-farvede vaskulære system(figur 4 A-C; G-I; Videoer 1 og 2).

Figur 1. Anbefalede mikrokirurgiinstrumenter. (A) mikro-skalpel formet af en synål. (B) fine Arkansas sten til at forme en mikro-skalpel. c) en lige saks, b) buet saks, c) 5 ml sprøjte med 181/2 G hypodermisk nål, d) plastpipette, e) specialfremstillet ægholder, f) sort blæk g) firkantet urglas, h) firkantet urglas med sort sylgardbase, i) mikroskalel på nåleholder, j) minutienstifter, k) minutien eller wolframnål på nåleholder, l) Pascheff-Wolff-springsaks, m) Dumont #5 pincet, n) perforeret ske, oi) kort ildtrøjet overførselsnål, oii) lang brandhærget inking nål, p) mundrør. Klik her for at se en større version af dette tal.

Figur 2. Intraspecies neuralrørstransplantation. Chick embryo / GFP neuralrør billeder er blevet ændret fra Delalande et al. 12. Vascularisering er ikke nødvendig for tarm kolonisering af enteriske neurale crest celler. Klik her for at se en større version af dette tal.

Figur 3. Intravenøs DiI-injektion. (A) Anbefalede instrumenter: a) CellTracker CM-DiI drop på parafilm, b) trukket glas injektion nål, c) mund rør. (B) Skematisk diagram over intravenøs DiI injektion i E4 kimære chick embryo. (C) i ovo DiI intravenøs injektion med fin glasnål indeholdende DiI indsat i venen (pil). (D) E4 kimære embryo post DiI injektion (rød) med GFP + neuralrør (pil). (E) DiI farvede fine blodkar netværk i et levende foster, 24 timer efter injektion. Br: hjerne; H: hjerte; LB: knop i lemmerne; A: allantois. Billeder i (C) og (D) er blevet ændret fra Delalande et al. 12 år Vaskulærisering er ikke nødvendig for tarm kolonisering af enteriske neurale crest celler. Klik her for at se en større version af dette tal.

Figur 4: Repræsentative resultater i maven og caecum af en E5.5 chick embryo. (A-C) 3 dimensionel (3D) rekonstruktion af en konfokal billedstak i maven, der viser (D) GFP+ enteriske neurale kamceller (ENCC) (E) Det DiI farvede vaskulære system og (F) et fusioneret billede af begge netværk D-F Histologiske sektioner på maveniveau, der viser (G) GFP+ ENCC (H) det DiI farvede vaskulære system og (I) et fusioneret billede af begge netværk. Nuclei er plettet med DAPI (cyan). (G-H) 3D-rekonstruktion af en konfokal billedstak i caecum-regionen, der viser (A) GFP+ ENCC-migreringsfronten i grønt, (B) det DiI-farvede vaskulære system i rødt og (C) et fusioneret billede af begge netværk. Billeder (A-F) er blevet ændret fra Delalande et al. 12 år Vaskulærisering er ikke nødvendig for tarm kolonisering af enteriske neurale crest celler. Klik her for at se en større version af dette tal.

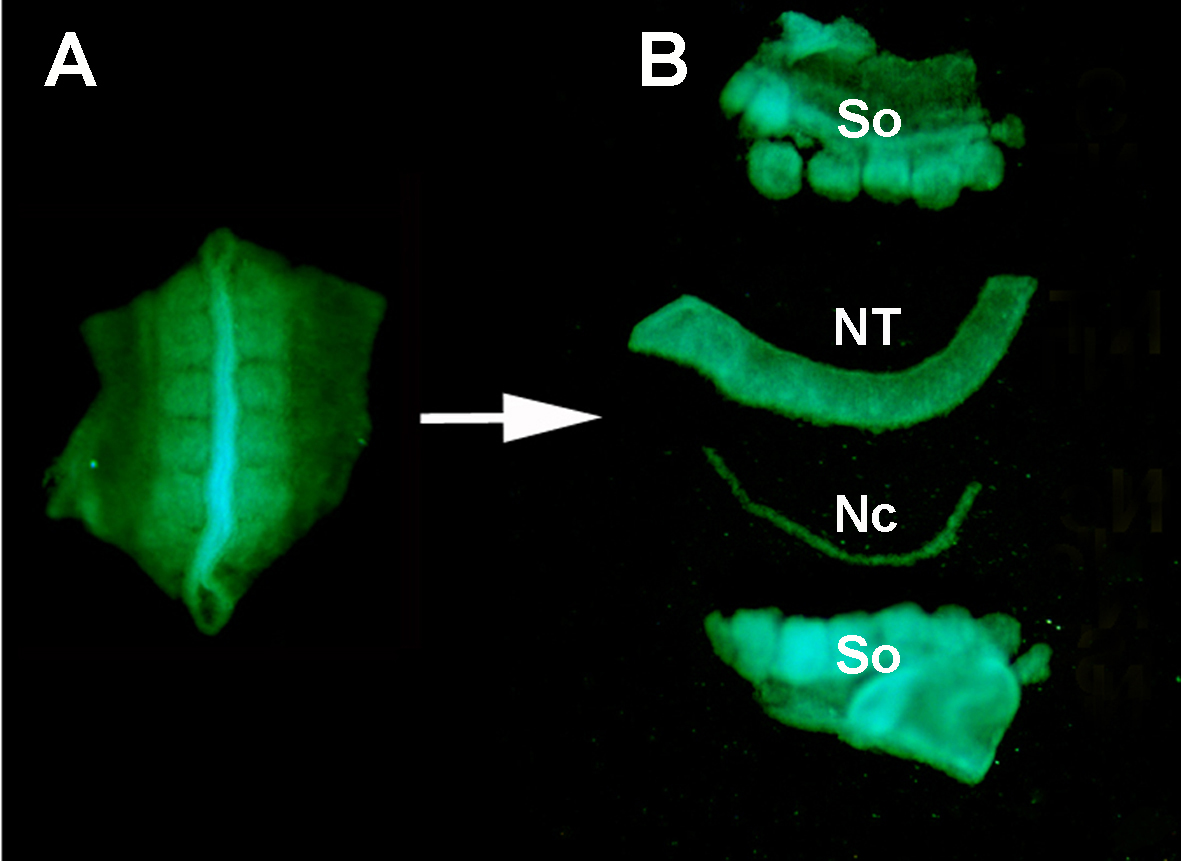
Figur S1. Isolering af en donor GFP + neuralrør fra de omkringliggende væv ved enzymatisk fordøjelse og mikro-dissektion. (A) GFP+ neuralrør og tilstødende somites dissekeret fra donorembryoet. (B) Isoleret neuralrør efter pancreatin fordøjelsen og mikro-dissektion ved hjælp af rustfrit minutien stifter. Så: somites; NT1 neuralrør; Nc: Notochord.

Video 1. 3-dimensionel 360° rotation af billedet i figur 4C, der viser det vaskulære system og ENCC i maven ved E5.5 (HH27-28). Klik her for at se denne video.

Video 2. 3-dimensionel 360° rotation af billedet i figur 4I, der viser det vaskulære system og ENCC-migreringsfronten i caecumområdet ved E5.5 (HH27-28). Klik her for at se denne video.
Discussion
Metoden til intraspecies neuralrørstransplantation kombineret med blodkarmærkning beskrevet her, drager fuld fordel af avianembrynets let adgang til ægget (sammenlignet med andre hvirveldyrembryoner) for at studere samudviklingen af et element i det autonome nervesystem (ENS) og det vaskulære system.
Til mærkning af NCC-derivater har kyllingenGFP-chick intraspecies podningsmetode, vi beskriver, en række fordele i forhold til den klassiske vagtel-chick chimera-metode, der blev etableret for over 40 år siden1-3. For det første, under FITC lys, GFP fluorescens er meget lyse, i det omfang, at GFP + celler er let mærkbar i levende kimære embryoner. Dette gør det muligt at kontrollere graftens succes i ovo, mens vagtel-chick podning kræver, at embryoet aflives, behandles og immunostained ved hjælp af QCPN, før graftens succes kan fastslås2. For det andet er GFP-udtrykket i den transgene kyllingGFP cytoplasmisk, derfor er det ikke kun etiketter cellelegemer, men tillader også, at fremskrivningerne af de transplanterede celler visualiseres22. Dette gør det muligt at observere indviklede neuronale netværk ved høj opløsning (bemærk, at fine fremskrivninger bedst visualiseres, når prøven immunostained med anti-GFP antistof). Da QCPN-mærkning er begrænset til vagtelcellekernen, afsløres sådanne netværk ikke ved hjælp af vagtel-chick kimærer. For det tredje eliminerer intraspecies podning eventuelle artsforskelle mellem celler i det kimære embryo. Da vagtele embryoner har en kortere inkubationstid end kylling (19 dage versus 21 dage) er det blevet foreslået, at vagtler celler har en højere spredning end chick celler, som potentielt kan påvirke udviklingen af kimære væv23. Interessant nok har det også vist sig i planter, at interspecies podning kan producere omfattende ændringer i DNA-methyleringsmønstre i vært 24. For det fjerde, chickGFP letter back-transplantation eksperimenter for at løse emner som NCC skæbne og celle engagement25. For det femte er den transgene kyllingGFP også nyttig til mange andre teknikker, herunder FACS sortering af GFP + celle delpopulationer, organotypisk kultur af organer, der indeholder GFP + celler, genetisk manipulation af GFP + podet væv via elektroporation af udtrykket plasmids26, og andre billeddannelse teknologier såsom optisk projektion tomografi27.
Den neurale rørtransplantation tilgang kan ændres ved mikrokirurgisk erstatte kortere mængder af neuralrør. Ved at bruge mindre segmenter af neuralrør mikrokirurgi er potentielt mindre skadeligt for embryoet og overlevelse kan forbedres. Ulempen ved at transplantere mindre neuralrør er imidlertid, at antallet af GFP + NCC i værten vil blive reduceret. Brugere kunne forsøge at opnå en balance mellem mængden af neuralrør transplanteret for at give optimal overlevelse af embryoner, og antallet af GFP + NCC i vært tarmen tilstrækkelig til at give informative resultater.
Til skibsmaling har DiI den fordel, at dets fluorescens er meget lyst og robust. Det har også kapacitet til at diffuse under fiksering forsikre farvning af de fineste åbne kapillærer. Da det er et vitalt farvestof, kan embryoner overleve injektionsproceduren og fortsætte med at udvikle sig med et farvet vaskulært system (op til 24 timer i vores hænder, selvom farvningen bliver mere punktlig over tid, se figur 3E). Kombinationen af chickGFP podning med DiI vaskulært maleri er derfor kompatibel med levende billeddannelse. Ud over alle disse fordele er det vigtigt at bemærke, at vaskulær injektion kun mærker lysarmerede beholdere og derfor ikke identificerer uåbnede kapillærer, endotelspidsceller eller isolerede endotelceller. Yderligere fremskridt inden for fugletransgenese kan imidlertid give nye måder at omgå sådanne problemer på, som eksemplificeret ved forsøg med Tg(tie1:H2B-eYFP) vagteulære embryoner til at studere vaskulær morfogenese28. En anden begrænsning af denne teknik er, at for effektiv mærkning af beholdere i embryoner ved E7.5 og derefter skal der injiceres større mængder farvestof, hvilket kan gøre eksperimenter dyre. En ændring af teknikken kan dog omfatte billige blodkar mærkningbrug highlighter blæk14, selv om denne fremgangsmåde ikke er blevet prøvet i vores hænder.
Kritiske trin i procedurerne omfatter processen med at visualisere embryoet ved at injicere blæk under blastodisc. Hvis membranen, der dækker æggeblommen, er revet af den blækfyldte nål på dette stadium, bliver embryoets overlevelse alvorligt kompromitteret. Det er også vigtigt, når man forbereder et donor neuralrør, at vævet ikke efterlades i alt for lang tid i bugspytkirtlen (overvej ca. 10 minutter som et maksimum). Langvarig udsættelse for pancreatin skader væv og neuralrøret er derefter vanskeligt at håndtere, og det vil ikke indarbejde godt ind i værten. Det er vigtigt at få erfaring med diI-injektionsteknikken på embryoner af vild type, før der injiceres kimære embryoner, da kun ét forsøg på injektion generelt er muligt for hvert foster. DiI volumen og nål diameter er kritiske parametre for hvert foster og bør vurderes på vilde type, fase matchede kontrol.
Afslutningsvis kan vores dobbelte mærkningsmetode til neuralrørstransplantation og DiI-karmaling i levende kyllingembryoner bruges til at undersøge de indbyrdes relationer mellem NCC og blodkarnetværk under organogenese. I betragtning af de mekanismer, der er ansvarlige for at etablere korrekt mål innervation og vaskularisering under organudvikling er stadig stort set ukendt, denne metode rummer potentiale for fremtidige opdagelser på dette område.
Disclosures
Forfatterne erklærer, at de ikke har nogen konkurrerende finansielle interesser.
Acknowledgments
Befrugtede GFP kyllingæg blev leveret af prof. Helen Sang, The Roslin Institute, og University of Edinburgh, UK. Roslin Transgenic Chicken Facility er finansieret af Wellcome Trust og af Biotechnology and Biological Sciences Research Council (BBSRC). Arbejdet blev delvist finansieret, og NT støttet af Great Ormond Street Hospital Children's Charity, London, UK. Forfatterne takker Ben Jevans, UCL Institute of Child Health, for hjælp til at forberede embryoner til podning.
Materials
| Name | Company | Catalog Number | Comments |
| Fertilised chick eggs | Henry Stewart and Co, Louth, UK | ||
| Fertilised GFP chick eggs | The Transgenic Chicken Facility, The Roslin Institute, The University of Edinburgh | ||
| Egg incubator (Profi-H Hatcher) | Lyon Technologies, CA, USA | 910-033 | |
| 14C Incubator | Precision Cooled Incubator, Leec Ltd., Nottingham, UK | Model LT2 | |
| Stereo-microscope | LEICA | Model MZ 12.5 | |
| Digital Camera | LEICA | DC500 | |
| Image acquisition software | LEICA | IM50 | |
| Goose neck halogen cold light source | Advanced Imaging Concepts, Inc | KL 1500 LCD | |
| 181⁄2 G hypodermic needle | SIGMA - ALDRICH | HSWNH181 | |
| Pancreatin | SIGMA - ALDRICH | P3292 | |
| DMEM | SIGMA - ALDRICH | D5030 | |
| Goat serum | SIGMA - ALDRICH | G6767 | |
| 5 ml syringe | SIGMA - ALDRICH | Z248010 | |
| Mouth tube | SIGMA - ALDRICH | A5177 | |
| Sigma Pasteur pipettes non-plugged, L 5 3/4 in. | SIGMA - ALDRICH | S6018 | |
| Transfer pipettes, polyethylene | SIGMA - ALDRICH | Z350796 | |
| Borosillicate glass capillaries, thin wall without filament | Harvard apparatus | PY8 30-0035 | |
| Iris Scissors - ToughCut | Fine Science Tools | 14058-09 | |
| Curved Iris Scissors - ToughCut | Fine Science Tools | 14059-09 | |
| Needle holders (Nickel-plated pin holder) | Fine Science Tools | 26018-17 | |
| Pascheff-Wolff Spring Scissors | Fine Science Tools | 15371-92 | |
| Dumont #5 forceps | Fine Science Tools | 11251-30 | |
| Minutien pins | Fine Science Tools | 26002-15 | |
| Dumont AA forceps, Inox Epoxy- coated | Fine Science Tools | 11210-10 | |
| Perforated spoon | Fine Science Tools | 10370-18 | |
| Tungsten needles (0.125mm diameter) | Fine Science Tools | 10130-05 | |
| Sellotape (clear, 24 mm width) | Any Supplier | ||
| Pen/Strep (Penicillin, Streptomycin) Solution | VWR international | 101447-068 | |
| Sylgard 184 silicone elastomer kit | Dow Corning | S09 512 516 | |
| Pelikan black ink | Pelikan | 211-169 | |
| CellTracker CM-DiI | Molecular Probes | C-7001 | |
| DAPI (4',6-Diamidino-2-Phenylindole, Dihydrochloride) | Molecular Probes | D1306 | |
| Settings for glass needle puller | Sutter Instruments | Flaming/Brown micropipette puller model P-86 | |
| Heat 950; Pull 150; Velocity 100; Time 200; Pressure 500 |
References
- Le Douarin, N. A biological cell labeling technique and its use in expermental embryology. Developmental Biology. 30, 217-222 (1973).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 461, 337-350 (2008).
- Teillet, M. A., Ziller, C., Le Douarin, N. M. Quail-chick chimeras. Methods in Molecular Biology. 97, 305-318 (1999).
- Garcia-Castro, M., Bronner-Fraser, M. Induction and differentiation of the neural crest. Current Opinion In. Cell Biology. 11, 695-698 (1999).
- Bhatt, S., Diaz, R., Trainor, P. A. Signals and switches in Mammalian neural crest cell differentiation. Cold Spring Harbor Perspectives In Biology. 5, (2013).
- Burns, A. J., Douarin, N. M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: spatiotemporal analysis of the development of the enteric nervous system. Development. 125, 4335-4347 (1998).
- Burns, A. J., Le Douarin, N. M. Enteric nervous system development: analysis of the selective developmental potentialities of vagal and sacral neural crest cells using quail-chick chimeras. The Anatomical Record. 262, 16-28 (2001).
- Burns, A. J., Delalande, J. M., Le Douarin, N. M. In ovo transplantation of enteric nervous system precursors from vagal to sacral neural crest results in extensive hindgut colonisation. Development. 129, 2785-2796 (2002).
- Burns, A. J., Champeval, D., Le Douarin, N. M. Sacral neural crest cells colonise aganglionic hindgut in vivo but fail to compensate for lack of enteric ganglia. Developmental Biology. 219, 30-43 (1006).
- Wang, X., Chan, A. K., Sham, M. H., Burns, A. J., Chan, W. Y. Analysis of the sacral neural crest cell contribution to the hindgut enteric nervous system in the mouse embryo. Gastroenterology. 141, 992-1002 (2011).
- Goldstein, A. M., Hofstra, R. M., Burns, A. J. Building a brain in the gut: development of the enteric nervous system. Clinical Genetics. 83, 307-316 (1111).
- Delalande, J. M., et al. Vascularisation is not necessary for gut colonisation by enteric neural crest cells. Developmental Biology. 385, 220-229 (2014).
- Anderson, R. B., Stewart, A. L., Young, H. M. Phenotypes of neural-crest-derived cells in vagal and sacral pathways. Cell And Tissue Research. 323, 11-25 (2006).
- Takase, Y., Tadokoro, R., Takahashi, Y. Low cost labeling with highlighter ink efficiently visualizes developing blood vessels in avian and mouse embryos. Development, Growth & Differentiation. 55, 792-801 (2013).
- Bates, D., Taylor, G. I., Newgreen, D. F. The pattern of neurovascular development in the forelimb of the quail embryo. Developmental Biology. 249, 300-320 (2002).
- Mayes, P., Dicker, D., Liu, Y., El-Deiry, W. Noninvasive vascular imaging in fluorescent tumors using multispectral unmixing. BioTechniques. 45, 459-460 (2008).
- Li, Y., et al. Direct labeling and visualization of blood vessels with lipophilic carbocyanine dye DiI. Nature Protocols. 3, 1703-1708 (2008).
- Eichmann, A., Thomas, J. L. Molecular parallels between neural and vascular development. Cold Spring Harbor Perspectives In Medicine. 3, a006551 (2013).
- Weinstein, B. M. Vessels and nerves: marching to the same tune. Cell. 120, 299-302 (2005).
- Carmeliet, P., Tessier-Lavigne, M. Common mechanisms of nerve and blood vessel wiring. Nature. 436, 193-200 (2005).
- Hamburger, V., Hamilton, H. L. A series of normal stages in the development of the chick embryo. Journal of Morphology. 88, 49-92 (1951).
- Barraud, P., et al. Neural crest origin of olfactory ensheathing glia. Proceedings of the National Academy of Sciences of the United States of America. 107, 21040-21045 (2010).
- Senut, M. C., Alvarado-Mallart, R. M. Cytodifferentiation of quail tectal primordium transplanted homotopically into the chick embryo. Brain Research. 429, 187-205 (1987).
- Wu, R., et al. Inter-species grafting caused extensive and heritable alterations of DNA methylation in Solanaceae plants. PLoS One. 8, e61995 (2013).
- Freem, L. J., Delalande, J. M., Campbell, A. M., Thapar, N., Burns, A. J. Lack of organ specific commitment of vagal neural crest cell derivatives as shown by back-transplantation of GFP chicken tissues. The International Journal Of Developmental Biology. 56, 245-254 (2012).
- Delalande, J. M., et al. The receptor tyrosine kinase RET regulates hindgut colonization by sacral neural crest cells. Developmental Biology. 313, 279-292 (2008).
- Freem, L. J., et al. The intrinsic innervation of the lung is derived from neural crest cells as shown by optical projection tomography in Wnt1-Cre;YFP reporter mice. Journal of Anatomy. 217, 651-664 (2010).
- Sato, Y., et al. Dynamic analysis of vascular morphogenesis using transgenic quail embryos. PloS One. 5, e12674 (2010).

{kind=link}