

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Dit protocol beschrijft de isolatie van satelliet-cellen uit branchiomeric hoofd spieren van een 9 weken oude ratten. De spieren komen uit verschillende kieuwbogen. Vervolgens worden de satelliet cellen gekweekt op een plek bekleding millimeter grootte hun differentiatie te bestuderen. Deze aanpak voorkomt de uitbreiding en de passage van satelliet-cellen.
Introduction
Ongeveer 1: 500-1: 1000 pasgeborenen vertonen gespleten waarbij de lip en / of gehemelte (CLP); waardoor dit de meest voorkomende congenitale misvormingen bij mensen 1. De spieren van het zachte gehemelte cruciaal zijn voor de werking van het zachte gehemelte tijdens spraak, slikken en zuigen. Wanneer een spleet van het zachte gehemelte aanwezig is, zijn deze spieren abnormaal ingebracht in het achterste uiteinde van de palatale bot.
Het zachte gehemelte en neer beweegt tijdens toespraak, het voorkomen van lucht kan ontsnappen door de neus. Kinderen met een spleet in de mond niet deze regelfunctie waardoor depressiviteit en velofaryngale dysfunctie 2,3 hebben. Hoewel de behandeling protocollen zijn variabel, chirurgische reparatie van het zachte gehemelte vindt plaats in de vroege kindertijd (6-36 maanden oud) 4. De abnormaal ingevoegd spieren van het zachte gehemelte kunnen chirurgisch worden gecorrigeerd 5-7 echter velofaryngale disfunctie blijft in 7% tot 30%van de patiënten 2,3,8-10.
Het vermogen van de skeletspieren te regenereren door de werking van satelliet-cellen (SC) is goed ingeburgerd 11,12. Bij spierblessure worden geactiveerd SC en migreren naar de plaats van letsel. Ze vervolgens woekeren, differentiëren en samensmelten tot nieuwe myovezels of reparatie beschadigde 13 vormen. Quiescent SC drukken de transcriptiefactor Pax7 14,15, terwijl hun nageslacht, de woekerende myoblasts, bovendien drukken de myogene bepaling factor 1 (MyoD) 16. Onderscheidende myoblasts beginnen te myogenin uiten (MyoG) 17. De terminale differentiatie van myoblasten wordt gekenmerkt door de vorming van spiervezels en de expressie van spier-specifieke eiwitten zoals myosine zware keten (MyHC) 16,18.
Onlangs hebben verschillende strategieën gebruikt in de regeneratieve geneeskunde om spieren regeneratie van ledematen spieren 19-23 te verbeteren. Specifieke studies opbranchiomeric hoofdspieren ook belangrijk omdat het werd onlangs aangetoond dat ze verschillen van andere spieren in diverse aspecten 24. In tegenstelling spieren van de ledematen, is gesuggereerd dat branchiomeric hoofdspieren bevatten minder SCS 25, regenereren langzamer en meer fibreus bindweefsel wordt gevormd na letsel 26 Bovendien prolifererende SC's van branchiomeric hoofdspieren ook andere transcriptiefactoren tot expressie. Zo wordt Tcf21, een transcriptiefactor voor craniofaciale spiervorming sterk tot expressie in regenererende hoofdspieren maar nauwelijks het regenereren ledemaatspieren 25. De spieren in het zachte gehemelte van CLP patiënten meestal kleiner en minder overzichtelijk vergelijking met normale palatal spieren 27,28. Langzame en snelle vezels zijn beide aanwezig in het zachte gehemelte spieren, maar de trage vezels zijn meer overvloedig. Daarentegen gespleten spieren bevatten een groter aandeel snelle vezels en ook een verminderde aanvoer capillairvergeleken met normale zachte gehemelte spieren 29-31. Snelle vezels zijn meer vatbaar voor krimp veroorzaakte letsel 31-33. De bijbehorende slechte capillaire aanbod kan ook het bevorderen van fibrose 34,35. Al deze aspecten kunnen bijdragen aan de slechte regeneratie van zacht gehemelte spieren na chirurgische gespleten sluiting 36. Met het oog hierop, een protocol voor de isolatie en karakterisering van branchiomeric head spier SC cruciaal. Dit biedt de mogelijkheid om SC biologie van branchiomeric hoofd spieren te bestuderen. Bovendien kunnen nieuwe therapieën gebaseerd op tissue engineering worden ontwikkeld spier regeneratie te bevorderen na operatie bij CLP en andere omstandigheden afbreuk het craniofaciale gebied.
In het algemeen kan SC's worden verkregen na dissociatie van spierweefsel 14. Fijnhakken, enzymatische digestie, en trituratie algemeen moeten SC ontslaan van hun niche. SC's kunnen worden gezuiverd door pre-plating op ongecoat schotels 14,37,38, fractionation op Percoll 39,40, of fluorescent- of magnetische celsortering 41-43. Hier presenteren we een nieuwe economische en snelle protocol voor de isolatie van satelliet-cellen uit branchiomeric hoofd spieren van jonge volwassen ratten. Dit protocol is gebaseerd op een eerdere manuscript 14 en specifiek aangepast aan kleine weefselmonsters. De isolatie van SC uit representatieve spieren afkomstig van de 1 e, 2 e en 4 e branchial bogen worden beschreven. Na isolatie lage aantallen satelliet cellen worden gekweekt op extracellulaire matrix gel spots van millimeter grootte hun differentiatie te bestuderen. Deze aanpak voorkomt dat de eis voor de uitbreiding en de passage van de SC.
Protocol
Alle experimenten hierin beschreven werden door de lokale raad voor Dierproeven van de Radboud Universiteit Nijmegen is goedgekeurd in overeenstemming met de Nederlandse wet- en regelgeving (RU-december 2013-205).
1. Extracellulaire Matrix Gel Locaties
- Voer de volgende stappen een dag voor de isolatie:
- Ontdooi een hoeveelheid extracellulaire matrix gel (100 ui) bij 4 ° C gedurende ten minste 1,5 uur. Verdun 1:10 in gemodificeerd Eagle's medium Dulbecco's; met 4500 mg / l glucose, 4 mM L-glutamine en 110 mg / ml natriumpyruvaat (DMEM). Houd de extracellulaire matrix gel bij 4 ° C te allen tijde. Let op: abrupte veranderingen in temperatuur zal resulteren in ongelijkmatige coating en kristalvorming.
- Houd de verdunde extracellulaire matrix geloplossing op ijs gedurende 15 min.
- Pre-chill een 20 ul micropipet gedurende 10 min.
- Zet 8-goed kamer dia's in een 100 mm petrischaal en breng het gerecht op een koude ondergrond (bijvoorbeeld een vriezer pack) gedurende 10 min.
- Met de pre-gekoelde micropipet een druppel 10 gl extracellulaire matrix gel in elk putje gebracht. Houd de petrischaal op de koude ondergrond nog minstens 7 min (Figuur 1A).
- Volledig verwijderen van de overblijvende extracellulaire matrix gel (Figuur 1B), en droog de putjes bij 37 ° C overnacht.
2. Dissectie van Head Spieren (kauwspieren, de kauw- en Levator Veli Palatini)
- Vóór versnijding Bereid 50 ml met fosfaat gebufferde zoutoplossing (PBS) aangevuld met 2% penicilline-streptomycine (P / S). Keep on ice.
- Na euthanasie van een jonge volwassene rat (9 weken) met een CO 2 / O 2, onthoofden de kop en verwijder de huid van het hoofd. Breng de kop ijskoude PBS gesupplementeerd met 2% P / S in een 50 ml buis.
- Kauwspieren (afgeleid van de 1e kiemboog)
- Plaats het hoofd met de ene kant op een siliconen pad en bevestig met onderhuidse needles (Figuur 2A).
- Identificeer de parotis en de gezichtszenuw (Figuur 2A). Expose de diepe fascia die de klier. Snijd de fascia en verwijder de klier met dissectie schaar. Identificeer de uitwendige gehoorgang. Trace de gezichtszenuw uit de stylomastoid foramen en het tijdelijke, jukbeen en buccale takken verwijder voorzichtig met een scalpel No. 15.
- Bevrijd de oppervlakkige hoofd van de kauwspieren door het verwijderen van de fascia. Identificeer zowel oppervlakkige en diepe hoofden van de kauwspieren. Trace de oppervlakkige hoofd tot zijn dikke tendineuze aponeurose ingebracht in het jukbeen proces van de bovenkaak.
- Scheid de pees van zijn oorsprong in het jukbeen proces met een rechte tang. Snijd het met een scalpel No. 15 of dissectie schaar en zorgvuldig leven is (Figuur 2B).
- Ontleden de oppervlakkige hoofd van de kauwspieren tot zijn inbrengen in de hoek en inferieure helft van tHij mantelvlak van de ramus van de onderkaak met een scalpel No. 15 (figuur 2C). Nu, volledig verwijderen van de spier.
- Achterste buik van de kauw- spieren (afgeleid van de 2 e kiemboog)
- Plaats het hoofd in een liggende positie op de siliconen pad en bevestig met injectienaalden (figuur 3A).
- Verwijder het onderhuidse vet bovenliggende zowel sublinguale en submandibulaire klieren. Verwijder vervolgens de oppervlakkige fascia en klieren met dissectie schaar. Expose de tweebuikvliezend spier (voorste en achterste buik).
- Houd de voorste pees van het achterste buik met een rechte tang, snijden en ontleden voorzichtig om tot zijn oorsprong in het trommelvlies bulla (Figuur 3B). Doe hetzelfde aan de contralaterale zijde.
- Levator veli palatini spier (afgeleid van het 4e kiemboog)
- Na dissectie van de achterste buik van de kauw- spieren, Lokaliseren de musculus stylohyoideus, trek het zijdelings, en verwijder deze (Figuur 4A) voorzichtig.
- Lokaliseren van de pees van de levator veli palatini dat voegt aan het trommelvlies bulla (Figuur 4A). Ontleden het zorgvuldig en snijd het aan beide kanten.
- Kijk voor de luchtpijp en de slokdarm dat er achter het loopt. Til de slokdarm en de keelholte, strottenhoofd en het zachte gehemelte bloot.
- Lokaliseren op het gebied van het zachte gehemelte waar de levator veli palatini is geplaatst en snijd het los (figuur 4B).
Opmerking: Direct na dissectie, verwijder voorzichtig pezen en bindweefsel van elke spier onder de stereo-microscoop. Dompel alle exemplaren snel in ethanol 70%, en overbrengen naar ijskoude PBS aangevuld met 2% P / S in een 15 ml buis.
3. Isolatie van satelliet-cellen
- Voer de volgende voorbereidende stappen voor SC isolatie van 3 groepen van de spieren:
- Bereid 7,5 ml van 0,1% pronase in DMEM. Filtreer de oplossing door een 0,22 pm filter. Vooraf verwarmen van de oplossing bij 37 ° C in een waterbad gedurende 10 min voor isolatie.
- Bereid 35 ml DMEM aangevuld met 10% paardenserum (HS) en 1% P / S. Ook voorverwarmen bij 37 ° C in een waterbad.
- Bereid 15 ml kweekmedium bestaande uit DMEM gesupplementeerd met 20% foetaal runderserum (FBS), 10% HS, 1% P / S en 1% kippenembryo extract (CEE). Voorverwarmen bij 37 ° C in een waterbad.
- Pre-coat zes plastic pipetten (10 ml) met HS en drogen ten minste 10 minuten voor gebruik.
- In de cultuur kap, breng elke spier in een putje van een 6-wells plaat. De dissectie schaar knippen de spier in kleine stukjes van ongeveer 2 mm. Wees voorzichtig het weefsel niet blad voor de mond te veel.
- Voeg voorzichtig 2,5 ml 0,1% pronase oplossing in elk putje en incuberen bij 37 ° C gedurende 60 min. Schud de plaat na 20, 40, en 60 min. Opmerking: De exacte DUURn van de incubatie afhankelijk van factoren zoals leeftijd en stam van de dieren.
- Monitor onder de microscoop. Controleer de spier fragmenten en stoppen van de enzymatische digestie als de vezels bundels ontvang losgemaakte uiterlijk (figuur 5).
- Voeg 2,5 ml DMEM aangevuld met 10% HS en 1% P / S. Transfer naar een 15 ml buis en centrifugeer de buizen bij 400 xg gedurende 5 minuten. Verwijder het supernatant door decanteren.
- Voeg 5 ml DMEM gesupplementeerd met 10% HS en 1% P / S. Pipetteer de oplossing op en neer met een 10 ml plastic pipet (fijnwrijven) minstens 20 keer om het weefsel te homogeniseren.
- Centrifugeer de buizen bij 200 xg gedurende 4 min. Verzamel de bovenstaande vloeistof en breng in een 15 ml buis.
- Voeg 5 ml DMEM gesupplementeerd met 10% HS en 1% P / S. Wederom pipet met een 10 ml plastic pipet totdat de weefselfragmenten passeert gemakkelijk door de pipet.
- Centrifugeer de buizen bij 200 xg gedurende 4 minuten en verzamel de bovenstaande vloeistof in een 15 ml buis.
- Put een cel zeef (40 um) naar een 50 ml buis en breng het supernatant dat de gedissocieerde cellen op het filter. Wassen met 1 ml DMEM voor maximale celherstel.
- Centrifugeer de buisjes bij 1000 xg gedurende 10 min en het supernatant met een pipet ontdoen.
- Resuspendeer de pellet in 300 pl cultuurmedium en tel de cellen in een hemocytometer.
4. Differentiatie van satelliet-cellen op de extracellulaire matrix Gel Spots
- Verdun de celsuspensie tot 1,5 x 10 3 cellen te verkrijgen in 10 pl kweekmedium.
- Bevestig de covers van de kamers dia's met tape en markeer de plekken met een zwarte marker op de onderkant van het object glas.
- Met behulp van een micropipet, een druppel van 10 ui celsuspensie op de extracellulaire matrix gel spot. Controleer onder de microscoop of de druppel celsuspensie correct is geplaatst op de plek. Incubeer gedurende zes uur bij 37 ° C.
- Voorzichtigly voeg 400 ul kweekmedium (DMEM gesupplementeerd met 20% FBS, 10% HS, 1% P / S en 1% CEE) en incubeer gedurende drie dagen bij 37 ° C.
NB: Op dit punt, zijn vers geïsoleerde SC onderworpen aan massale trauma (enzymatische digest en hard wrijven) en die ze nodig hebben om te herstellen. Laat de cellen niet verstoren tijdens de eerste drie dagen 37. Vervolgens kan het kweekmedium worden veranderd afhankelijk van het soort experiment.
De extracellulaire matrix gel spots zijn geënt met een hoge celdichtheid (1,5-2,5 x 10 20/03 ul) tot differentiatie assay. Het kweekmedium (DMEM gesupplementeerd met 20% FBS, 10% HS, 1% P / S en 1% extract van kippenembryo's) kunnen elke derde dag vervangen. - Als alternatief, als uitbreiding en passing gewenst is volg dan de volgende stappen:
- Ontdooi een hoeveelheid extracellulaire matrix gel (500 ui) bij 4 ° C gedurende ten minste 1,5 uur. Verdun 1:10 in DMEM en volg de aanbevelingen in punt 1.1.1.
- Pre-chill een pipet 10 ml gedurende 10 min bij 4 &# 176; C.
- Transfer drie T75 flessen op een koude ondergrond (bijvoorbeeld een vriezer pack) gedurende 10 min.
- Met de pre-gekoelde pipet 1 ml extracellulaire matrix gel gebracht in elke kolf. Controleer dat het oppervlak volledig bedekt is. Houd de kolven op het koude oppervlak nog tenminste 7 min (Figuur 1A).
- Volledig verwijderen van de overblijvende extracellulaire matrix gel met een pipet 10 ml, en droog de putjes bij 37 ° C gedurende 1 uur.
- Na het tellen, resuspendeer de vers geïsoleerde SC in 10 ml kweekmedium (DMEM gesupplementeerd met 20% FBS, 10% HS, 1% P / S en 1% kippenembryo extract) en zaad in de pre-coated T75 flessen.
- Na drie dagen, wijzigt het medium (en elke derde dag) tot 80% confluentie bereikt. Voor passage, was de T75 kolven driemaal met PBS. Voeg vervolgens 1 ml 0,25% trypsine-oplossing en incubeer gedurende drie minuten bij 37 ° C. Resuspendeer in 9 ml kweekmedium (DMEM gesupplementeerd met 20% FBS, 10% HS, 1% P / Sen 1% kippenembryo extract) en centrifugeer bij 200 xg gedurende 5 minuten. Verwijder het supernatant. Na het tellen, mengen 1 x 10 6 cellen in 1000 ul kweekmedium en het bevriezen van de cellen.
Representative Results
Met behulp van dit protocol, de kauwspieren (een zijde) levert 0,8-1 x 10 6 cellen, de kauw- spieren (posterior buik) levert 1,5-2 x 10 5 cellen en levator veli palatini spier opbrengsten 1-1,5 x 10 5 cellen. Cell rendement afhankelijk van het type spier, stam en leeftijd van het dier. Bij vergelijking van de drie spiergroepen werden vers geïsoleerde SC uitgezaaid met dezelfde celdichtheid (1,5 x 10 3/10 gl). Direct na isolatie, meer dan 90% van de vers geïsoleerde cellen brengen Pax7 (figuur 6).
Dag 4, 7 en 10 kweken werden gekleurd met antilichamen tegen Pax7, MyoD, MyoG en MyHC immunokleuring. Vijf willekeurige velden werden geteld per cultuur met een 20X objectief. Op dag 4 en 7 Pax Myo D is uitgedrukt in alle spiergroepen (figuren 6 en 7 en 8), maar het nageslacht van SatCs van de kauwspieren en de kauwspieren start expreszingen myogenin eerder dan de levator veli palatini spier (figuur 9). Op dag 10 wordt de expressie van MyoG sterk verlaagde in alle groepen (Figuur 9). Enkele dagen na het zaaien op de extracellulaire matrix gel spots, de prolifererende cellen beginnen te smelten en vormen multi-kernhoudende myotuben, die myosine zware keten tot expressie. Kleine myotubes zijn duidelijk zichtbaar op dag 7 (Figuur 10). Op dag 10, kan trillen van de myotubes opgemerkt (Video 1).

Figuur 1:. Extracellulaire matrix gel spots in een kamer schuif (A) voor gemakkelijke manipulatie, zet de 8-well kamer dia in een 100 mm petrischaal. Pipet 10 ul extracellulaire matrix gel in elke kamer en zet het op een koude ondergrond (7 min). (B) Kamer dia na de overtollige extracellulaire matrix gel wordt verwijderd.

Figuur 2:. Dissectie van de kauwspieren (A) hoofd van het dier in een zijaanzicht. Oor (E), parotis (P) en gezicht zenuw (VII). (B) tendineuze aponeurose (Te) van de oppervlakkige hoofd van de kauwspieren (mevrouw) en temporele spieren (T). Scheid de pees van de insertie met een pincet. (C) te ontleden de spier voorzichtig totdat het inbrengen in de ramus van de onderkaak. E: oor, P: oorspeekselklier, VII: gezichtszenuw, T: Temporal spier, mevrouw: oppervlakkige hoofd van de kauwspieren, Te: pees, Mp: diepe hoofd van de kauwspieren.

Figuur 3: Dissectie van de achterste buik van de kauw- spieren (.A) kop van het dier in een liggende positie. Lokaliseer de submandibulaire klier (Sg), kauwspieren (M), gezichtszenuw (VII) en sternocleidomastoideus (SCM). Verwijder de submandibulaire klier. (B) Lokaliseer de kauw- spieren anterior (AD) en posterior buik (PD). Met een rechte tang, neem de voorste pees van de achterste buik, snijd het ontleden en het zorgvuldig tot zijn oorsprong in het trommelvlies bulla (ty). E: oor, Sg: submandibulaire klier, VII: gezichtszenuw, M: kauwspieren, SMC: sternocleidomastoideus, AD: voorste buik tweebuikvliezend spier, PD: posterior buik tweebuikvliezend spier, Ty: Tympanic bulla.

Figuur 4:. Dissection van de levator veli palatini spier (A) De algemene opvatting na dissectie van de kauw- spieren (posterior buik). Musculus stylohyoideus (St) en de pees van de levatorveli palatini kunnen worden gelokaliseerd. Let op de trachea (T) en de slokdarm (Es) die achter. (B) Na het optillen van de luchtpijp en de slokdarm de keelholte (P) wordt blootgesteld. Het inbrengen van de levator veli palatini in het zachte gehemelte is nu zichtbaar. De pijl geeft de ontleed zachte gehemelte met de levator veli palatini spieren aan beide zijden. E: oor, St: musculus stylohyoideus, VII: gezichtszenuw, M: kauwspieren, AD: voorste buik tweebuikvliezend spier, PD: posterior buik tweebuikvliezend spier, T: luchtpijp, Es: slokdarm, P: keelholte, * levator veli palatini spier .

Figuur 5: Weergave van het spierweefsel (A) vóór en (B) na enzymatische digestie met pronase. Merk op dat spierbundels lijken losgemaakt na enzymatische digestie.
Figuur 6: Pax 7 immunokleuring Vers geïsoleerde SC, toegepast op extracellulaire matrix gel eind isolatie (ongeveer 6 uur na de eerste tissue digestie).. Vijf willekeurige velden werden geteld onder toepassing van een 10X objectief met een gemiddelde van 210 cellen per veld. Ongeveer 90% van de cellen positief Pax 7. DAPI: blauw, Pax7: rood. Schaal bar, 100 micrometer.

Figuur 7:. Pax 7, MyoD immunokleuring Dag 4, 7 en 10 kweken werden gekleurd met antilichamen tegen Pax7 en MyoD immunokleuring. (A - C) en (D - F) Representatieve microfoto's van dag 4 en 7 culturen uit de kauwspieren. (G en H + en MyoD + kernen per microscopische veld werd geteld en uitgedrukt als een percentage van het totale aantal kernen (DAPI). DAPI: blauw, Pax7: rood en MyoD: groen. Schalen bar, 100 micrometer. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 8:. Verdeling Pax7 ± / ± MyoD in kweken van mononucleaire cellen in kweken van kauwspieren, kauw- en levator veli palatine spier (A - C) Dag 4, 7 en 10 kweken werden gekleurd met antilichamen tegen Pax7 en MyoD immunokleuring. Het totaal aantal cellen op basis van het totale aantal kernen (DAPI). (D) Gegevens kwantificering van Pax7 ± / MyoD ± CELls. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 9:. Myogenin immunokleuring dag 4, 7 en 10 kweken werden gekleurd met antilichamen tegen myogenin. (A - D) Vertegenwoordiger microfoto van dag 4 en 7 culturen uit de levator veli palatine spier. (E) Het aantal MyoG + nuclei per microscopische gebied werd geteld en uitgedrukt als percentage van het totale aantal kernen (DAPI). (F) gegevens kwantificering van MyoG + cellen. DAPI: blauw, myogenin: groen. Schalen bar, 100 micrometer. Klik hier om een grotere versie van deze figur bekijkene.

Figuur 10:. Myosine heavy chain-immunokleuring dag 4, 7 en 10 kweken werden gekleurd met antilichamen tegen myosine zware keten (MyHC). Representatieve microfoto's van dag 4, 7 en 10 kweken van de kauw- (DIG) spier. Op dag 7, kleine myotubes zijn aanwezig, terwijl op dag 10 lang en goed georganiseerde myotubes zijn evident. Schalen bar, 200 pm. Klik hier om een grotere versie van deze figuur te bekijken.
Video 1:.. Myotube spiertrekkingen Voorbeelden van twee representatieve velden met spiertrekkingen myotubes worden getoond voor dag 10 culturen uit tweebuikvliezend spier Klik hier om deze video te bekijken.
Discussion
SC's uit verschillende branchiomeric hoofdspieren werden geïsoleerd uit een 9-weken oude Wistar rat en gekweekt direct aan extracellulaire matrix gel spots zonder expansie en passage. Na isolatie werden de cellen geteld en geënt bij dezelfde celdichtheid. Voor de parallelle isolatie van drie verschillende spieren, neemt deze methode ongeveer 4 uur. Cultuur verontreiniging te voorkomen, een cruciale stap is de snelle wassen in alcohol 70% na dissectie van de spieren.
Tijdens SC isolatie is het belangrijk om het spierweefsel in kleine stukjes (ongeveer 2 mm) gesneden maar vermijd teveel fijnhakkende omdat dit zal resulteren in een kleine celopbrengst door celbeschadiging. Ook de duur van de enzymatische digestie zorgvuldig worden onder de microscoop gecontroleerd om verdere schade te voorkomen. Doel van de spijsvertering met de myovezels dissociëren. Omdat meer dan 90% van de geïsoleerde cellen brengen Pax7, wordt geen verdere zuivering vereist (figuren 6-8).Dit voorkomt extra zuiveringsstappen in andere werkwijzen zoals pre-plating op ongecoat schotels 14,37,38, fractionering op Percoll 39,40 of fluorescent- of magnetische celsortering 41,43. Want tritureren is het essentieel om afschuiving tussen het weefsel fragmenten en de opening van de pipetpunt induceren omdat hiermee de mechanische ontgrendeling van de SC. Als het wrijven met een pipet 10 ml (binnendiameter tip: 1 mm) is moeilijk, een 5 ml (binnendiameter tip: 2 mm) kan de pipet eerst worden gebruikt. Alternatief kan glas Pasteur pipetten op de gewenste diameter worden gesneden en gebruikt. Deze werkwijze is eenvoudig, efficiënt en maakt de gelijktijdige isolatie van SC uit verschillende spieren samples.
De kweekplaten van SC's kunnen ook worden bekleed met gelatine of collageen, maar onze eerdere studies tonen aan dat extracellulaire matrix gel is beter voor het onderhoud van het myogene potentiaal dan 38 collageen. De extracellulaire matrix gel spots vanmillimeter size (10 gl / Ø 2 mm of 20 gl / Ø 4 mm) maakt het onderzoek mogelijk van de proliferatie en differentiatie van SCS met een beperkt aantal cellen. Voor de differentiatie assay ongeveer 8 tot 20 maal minder cellen worden vereist vergeleken met een 24-well plaat (Ø 15,6 mm) en ongeveer 80 tot 200 keer minder in vergelijking met 35 mm petrischalen (Ø 35 mm) 14,38.
Omdat extracellulaire matrix gel is duur, deze methode ook kosteneffectiever. Bovendien kan de kamer objectglaasjes worden vervangen door plastic dekglaasjes in de kosten verder te verlagen. Voor de bereiding van de extracellulaire matrix gel vlekken nadrogen van de kamer objectglaasjes noodzakelijk. Aangezien de extracellulaire matrix gel spots transparant, is het noodzakelijk om de vlekken aan de onderzijde markeren met achterverlichting. De kamers dia's worden vastgesteld in een petrischaal voor eenvoudige manipulatie. Verdere celkweek uitbreiding niet noodzakelijk, wat de mogelijkheid van de SC smal studie biedtLER spieren of kleine spier monsters. Alternatief, bijvoorbeeld voor PCR of spier constructen als meer cellen nodig, kan de vers geïsoleerde SC's eerst worden geëxpandeerd in T75 flessen zoals hierboven aangegeven.
SC's die met dit protocol zijn niet geschikt voor verdere zuivering met flowcytometrie onmiddellijk na isolatie. De digestie met pronase veroorzaakt uitgebreide digestie van het 14 oppervlak antigenen. Paardenserum en foetaal runderserum die gebruikt worden voor celkweek eerst goed worden gekarakteriseerd voor isolatie, aangezien verschillende lotnummers verschillend beïnvloed myoblasten proliferatie en differentiatie.
De laatste jaren is er een groeiende belangstelling voor de spieren afkomstig van de kieuwbogen en de kop mesoderm (bv oculaire spieren) 24. Het is duidelijk aangetoond dat het hoofd en de ledematen spieren hebben zeer verschillende eigenschappen. Kauwspieren van oude dieren lijkt te reTain hun regeneratieve capaciteit in vergelijking met de ledematen spieren 25,26. SC van de oculaire spieren hebben een robuuste proliferatie en differentiatie capaciteit vergelijkbaar met SC's van het hoofd spieren, en tonen een grotere aanslaan potentieel dan ledemaat spier SCS 24.
Het type vezel distributie en myosine samenstelling varieert tussen spiergroepen en ook tussen de soorten. Spieren afkomstig van de eerste kiemboog bij de mens bevatten zowel langzame en snelle vezels (subtypes IIA en IIX), neonatale myosins en myosins typisch voor het ontwikkelen van de hartspier. In knaagdieren deze spieren bevatten ongeveer 95% snelle vezels myosine IIA en IIb) 44-46. Studies over aviaire spieren aangetoond dat SC's uit verschillende vezelsoorten spier variëren differentiatievermogen. SC van snelle vezels alleen differentiëren in snelle spiervezels, terwijl SC uit langzame vezels kunnen differentiëren in zowel vezels types 47. Bovendien is het percentage SC's in snelle spierenvezels is lager dan in langzame spiervezels 48,49. Dit geeft aan dat het type vezelverdeling te houden bij studies moet worden genomen spieren in het craniofaciale gebied. Vergelijkbaar met gespleten gehemelte spieren, de LVP bij knaagdieren bevat bijna uitsluitend snelle vezels 50. Daarom SC van de LVP geschikt voor preklinische studies op het gebied van gespleten gehemelte.
Dit protocol biedt nieuwe mogelijkheden om SC afgeleid van branchiomeric hoofd spieren of andere kleinere spieren of kleinere spieren monsters te bestuderen. Dit zal de ontwikkeling van nieuwe therapieën vergemakkelijken de regeneratie van spieren in de maxillofaciale gebied te verbeteren zoals een gespleten gehemelte, maar ook in andere aandoeningen van kleine spieren.
Materials
| Name | Company | Catalog Number | Comments |
| Hypodermic Needle 25 G 0.5 x 25 m | BD Microlance | 300400 | |
| Dissecting scissors | Braun | BC154R | |
| Micro forceps straight | Braun | BD330R | |
| Surgical Scalpel Blade No. 15 | Swann-Morton | 0205 | |
| Alcohol 70% | Denteck | 2,010,005 | |
| Permanox Slide, 8 Chamber | Thermo Scientific | 177445 | |
| 6 well cell culture plate | Greiner bio-one | 657160 | |
| Cell Culture Dishes (100 x 20 mm) | Greiner bio-one | 664160 | |
| 15 ml sterile conical centrifuge tube | BD Biosciences | 352097 | |
| 50 ml sterile conical centrifuge tube | BD Biosciences | 352098 | |
| Cell strainer (40 μm) | Gibco | 431750 | |
| 10 ml serological pipette | Greiner bio-one | 607180 | |
| 20 µl FT20 | Greiner bio-one | 774288 | |
| Matrigel, Phenol-Red Free | BD Biosciences | 356237 | 10 ml |
| Pronase | Calbiochem | 53702 | 10 KU |
| Phosphate Buffered Saline | Gibco | 14190-144 | 500 ml |
| Dulbecco's Modified Eagle Medium, high glucose, GlutaMAX Supplement, pyruvate | Gibco | 10569-010 | 500 ml |
| Fetal Bovine Serum | Fisher Scientific | 3600511 | 500 ml |
| Horse Serum | Gibco | 26050088 | 500 ml |
| Penicillin-Streptomycin (10,000 U/ml) | Gibco | 15140-122 | 100 ml |
| Chicken Embryo Extract | MP Biomedicals | 2850145 | 20 ml |
References
- Gritli-Linde, A. Molecular control of secondary palate development. Developmental Biology. 301, 309-326 (2007).
- Marrinan, E. M., LaBrie, R. A., Mulliken, J. B. Velopharyngeal function in nonsyndromic cleft palate: relevance of surgical technique, age at repair, and cleft type. The Cleft Palate-Craniofacial Journal. 35, 95-100 (1998).
- Morris, H. L. Velopharyngeal competence and primary cleft palate surgery, 1960-1971: a critical review. The Cleft Palate Journal. 10, 62-71 (1973).
- Mossey, P. A., Little, J., Munger, R. G., Dixon, M. J., Shaw, W. C. Cleft lip and palate. Lancet. 374, 1773-1785 (2009).
- Boorman, J. G., Sommerlad, B. C. Musculus uvulae and levator palati: their anatomical and functional relationship in velopharyngeal closure. British Journal of Plastic Surgery. 38, 333-338 (1985).
- Bae, Y. C., Kim, J. H., Lee, J., Hwang, S. M., Kim, S. S. Comparative study of the extent of palatal lengthening by different methods. Annals of Plastic Surgery. 48, 359-362 (2002).
- Braithwaite, F., Maurice, D. G. The importance of the levator palati muscle in cleft palate closure. British Journal of Plastic Surgery. 21, 60-62 (1968).
- Inman, D. S., Thomas, P., Hodgkinson, P. D., Reid, C. A. Oro-nasal fistula development and velopharyngeal insufficiency following primary cleft palate surgery--an audit of 148 children born between 1985 and 1997. British Journal of Plastic Surgery. 58, 1051-1054 (2005).
- Phua, Y. S., de Chalain, T. Incidence of oronasal fistulae and velopharyngeal insufficiency after cleft palate repair: an audit of 211 children born between 1990 and 2004. The Cleft Palate-Craniofacial Journal. 45, 172-178 (1990).
- Kirschner, R. E., et al. Cleft-palate repair by modified Furlow double-opposing Z-plasty: the Children's Hospital of Philadelphia experience. Plastic and Reconstructive Surgery. 104, 1998-2010 (1999).
- Mauro, A. Satellite cell of skeletal muscle fibers. The Journal of Biophysical and Biochemical Cytology. 9, 493-495 (1961).
- Yablonka-Reuveni, Z. The skeletal muscle satellite cell: still young and fascinating at 50. The Journal of Histochemistry and Cytochemistry. 59, 1041-1059 (2011).
- Ten Broek, R. W., Grefte, S., Von den Hoff, J. W. Regulatory factors and cell populations involved in skeletal muscle regeneration. Journal of Cellular Physiology. 224, 7-16 (2010).
- Danoviz, M. E., Yablonka-Reuveni, Z. Skeletal muscle satellite cells: background and methods for isolation and analysis in a primary culture system. Methods in Molecular Biology. 798, 21-52 (2012).
- Seale, P., et al. Pax7 is required for the specification of myogenic satellite cells. Cell. 102, 777-786 (2000).
- Yablonka-Reuveni, Z., et al. The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Developmental Biology. 210, 440-455 (1999).
- Zammit, P. S., Partridge, T. A., Yablonka-Reuveni, Z. The skeletal muscle satellite cell: the stem cell that came in from the cold. The Journal of Histochemistry And Cytochemistry. 54, 1177-1191 (2006).
- Andres, V., Walsh, K. Myogenin expression, cell cycle withdrawal, and phenotypic differentiation are temporally separable events that precede cell fusion upon myogenesis. The Journal of Cell Biology. 132, 657-666 (1996).
- Fukushima, K., et al. The use of an antifibrosis agent to improve muscle recovery after laceration. The American Journal of Sports Medicine. 29, 394-402 (2001).
- Grefte, S., Kuijpers-Jagtman, A. M., Torensma, R., Von den Hoff, J. W. Skeletal muscle fibrosis: the effect of stromal-derived factor-1α-loaded collagen scaffolds. Regenerative Medicine. 5, 737-747 (2010).
- Jackson, W. M., Nesti, L. J., Tuan, R. S. Potential therapeutic applications of muscle-derived mesenchymal stem and progenitor cells. Expert Opinion on Biological Therapy. 10, 505-517 (2010).
- Sato, K., et al. Improvement of muscle healing through enhancement of muscle regeneration and prevention of fibrosis. Muscle, & Nerve. 28, 365-372 (2003).
- Tatsumi, R., Anderson, J. E., Nevoret, C. J., Halevy, O., Allen, R. E. HGF/SF is present in normal adult skeletal muscle and is capable of activating satellite cells. Developmental Biology. 194, 114-128 (1998).
- Stuelsatz, P., et al. Extraocular muscle satellite cells are high performance myo-engines retaining efficient regenerative capacity in dystrophin deficiency. Developmental biology. , (2014).
- Ono, Y., Boldrin, L., Knopp, P., Morgan, J. E., Zammit, P. S. Muscle satellite cells are a functionally heterogeneous population in both somite-derived and branchiomeric muscles. Developmental Biology. 337, 29-41 (2010).
- Pavlath, G. K., et al. Heterogeneity among muscle precursor cells in adult skeletal muscles with differing regenerative capacities. Developmental Dynamics. 212, 495-508 (1998).
- Koo, S. H., Cunningham, M. C., Arabshahi, B., Gruss, J. S., Grant, J. H. 3rd The transforming growth factor-beta 3 knock-out mouse: an animal model for cleft palate. Plastic and Reconstructive Surgery. 108, 938-948 (2001).
- Fara, M., Brousilova, M. Experiences with early closure of velum and later closure of hard palate. Plastic and Reconstructive Surgery. 44, 134-141 (1969).
- Lindman, R., Paulin, G., Stal, P. S. Morphological characterization of the levator veli palatini muscle in children born with cleft palates. The Cleft Palate-Craniofacial Journal. 38, 438-448 (2001).
- Hanes, M. C., et al. Contractile properties of single permeabilized muscle fibers from congenital cleft palates and normal palates of Spanish goats. Plastic and Reconstructive Surgery. 119, 1685-1694 (2007).
- Rader, E. P., et al. Contraction-induced injury to single permeabilized muscle fibers from normal and congenitally-clefted goat palates. The Cleft Palate-Craniofacial Journal. 44, 216-222 (2007).
- Rader, E. P., et al. Effect of cleft palate repair on the susceptibility to contraction-induced injury of single permeabilized muscle fibers from congenitally-clefted goat palates. The Cleft Palate-Craniofacial Journal. 45, 113-120 (2008).
- Macpherson, P. C., Dennis, R. G., Faulkner, J. A. Sarcomere dynamics and contraction-induced injury to maximally activated single muscle fibres from soleus muscles of rats. The Journal of Physiology. 500 (Pt 2), 523-533 (1997).
- Koch, K. H., Grzonka, M. A., Koch, J. The pathology of the velopharyngeal musculature in cleft palates). Annals of Anatomy. 181, 123-126 (1999).
- Fara, M., Dvorak, J. Abnormal anatomy of the muscles of palatopharyngeal closure in cleft palates: anatomical and surgical considerations based on the autopsies of 18 unoperated cleft palates. Plastic and Reconstructive Surgery. 46, 488-497 (1970).
- Carvajal Monroy, P. L., Grefte, S., Kuijpers-Jagtman, A. M., Wagener, F. A., Von den Hoff, J. W. Strategies to Improve Regeneration of the Soft Palate Muscles After Cleft Palate Repair. Tissue Engineering. Part B, Reviews. , (2012).
- Grefte, S., Kuijpers, M. A., Kuijpers-Jagtman, A. M., Torensma, R., Von den Hoff, J. W. Myogenic capacity of muscle progenitor cells from head and limb muscles. European Journal of Oral Sciences. 120, 38-45 (2012).
- Grefte, S., Vullinghs, S., Kuijpers-Jagtman, A. M., Torensma, R., Von den Hoff, J. W. Matrigel but not collagen I, maintains the differentiation capacity of muscle derived cells in vitro. Biomedical Materials. 7, 055004 (2012).
- Kastner, S., Elias, M. C., Rivera, A. J., Yablonka-Reuveni, Z. Gene expression patterns of the fibroblast growth factors and their receptors during myogenesis of rat satellite cells. The Journal of Histochemistry and Cytochemistry. 48, 1079-1096 (2000).
- Yablonka-Reuveni, Z., Quinn, L. S., Nameroff, M. Isolation and clonal analysis of satellite cells from chicken pectoralis muscle. Developmental Biology. 119, 252-259 (1987).
- Sherwood, R. I., et al. Isolation of adult mouse myogenic progenitors: functional heterogeneity of cells within and engrafting skeletal muscle. Cell. 119, 543-554 (2004).
- Gilbert, P. M., et al. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science. 329, 1078-1081 (2010).
- Motohashi, N., Asakura, Y., Asakura, A. Isolation culture, and transplantation of muscle satellite cells. Journal of Visualized Experiments. , (2014).
- Sciote, J. J., Horton, M. J., Rowlerson, A. M., Link, J. Specialized cranial muscles: how different are they from limb and abdominal muscles. Cells, Tissues, Organs. 174, 73-86 (2003).
- Rowlerson, A., Mascarello, F., Veggetti, A., Carpene, E. The fibre-type composition of the first branchial arch muscles in Carnivora and Primates. Journal of Muscle Research and Cell Motility. 4, 443-472 (1983).
- Muller, J., et al. Comparative evolution of muscular dystrophy in diaphragm, gastrocnemius and masseter muscles from old male mdx mice. Journal of Muscle Research and Cell Motility. 22, 133-139 (2001).
- Feldman, J. L., Stockdale, F. E. Skeletal muscle satellite cell diversity: satellite cells form fibers of different types in cell culture. Developmental Biology. 143, 320-334 (1991).
- Schmalbruch, H., Hellhammer, U. The number of nuclei in adult rat muscles with special reference to satellite cells. The Anatomical Record. 189, 169-175 (1977).
- Gibson, M. C., Schultz, E. The distribution of satellite cells and their relationship to specific fiber types in soleus and extensor digitorum longus muscles. The Anatomical Record. 202, 329-337 (1982).
- Carvajal Monroy, P. L., et al. A rat model for muscle regeneration in the soft palate. PloS One. 8, e59193 (2013).
Tags
Developmental Biology hoofd spieren levator veli palatini spier kauw- spieren kauwspieren satelliet-cellen isolatie primaire cellen gespleten gehemelte regeneratieve geneeskunde tissue engineering stamcellen differentiatie myofibersErratum
Formal Correction: Erratum: Isolation and Characterization of Satellite Cells from Rat Head Branchiomeric Muscles
Posted by JoVE Editors on 10/01/2015.
Citeable Link.
An erratum was issue for Isolation and Characterization of Satellite Cells from Rat Head Branchiomeric Muscles. The fourth figure was updated to explain the isolation of the LVP better.
Steps 2.5.3 and 2.5.4 were updated from:
2.5.3. Look for the trachea and the esophagus that runs behind it. Lift the esophagus, and expose the pharynx, larynx and the soft palate.
2.5.4. Localize the area of the soft palate where the levator veli palatini is inserted and cut it loose (Figure 4B).
to
2.5.3. Look for the trachea and the esophagus that runs behind it. Lift the esophagus, and expose the pharynx and the larynx.
2.5.4 Localize and dissect the area of the superior pharyngeal constrictor muscle. Identify the levator veli palatini and cut it at both sides (Figure 4B).
Figure 4 and its legend were updated from:
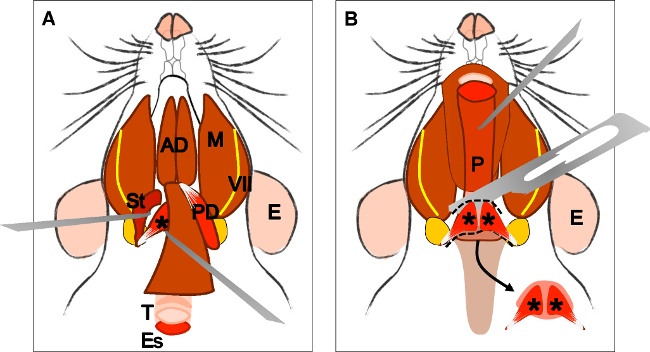
Figure 4: Dissection of the levator veli palatini muscle. (A) General view after dissection of the digastric muscle (posterior belly). Stylohyoid muscle (St) and tendon of the levator veli palatini can be localized. Note the trachea (T) and esophagus (Es) running behind it. (B) After lifting the trachea and the esophagus the pharynx (P) is exposed. The insertion of the levator veli palatini into the soft palate is now visible. The arrow indicates the dissected soft palate with the levator veli palatini muscles at both sides. E: ear, St: stylohyoid muscle, VII: facial nerve, M: masseter muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, T: trachea, Es: esophagus, P: Pharynx, *levator veli palatini muscle.
to
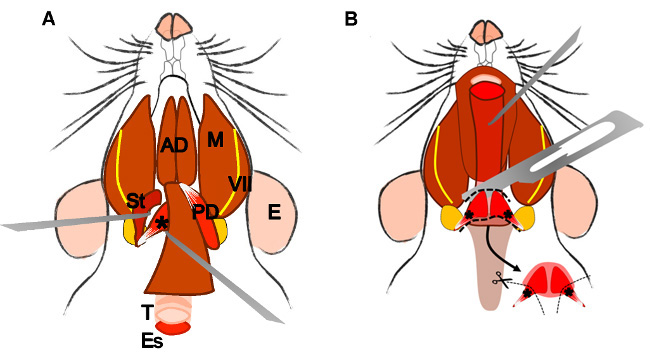
Figure 4: Dissection of the levator veli palatini muscle. (A) General view after dissection of the digastric muscle (posterior belly). Stylohyoid muscle (St) and tendon of the levator veli palatini can be localized. Note the trachea (T) and esophagus (Es) running behind it. (B) After lifting the trachea and the esophagus the pharynx (P) is exposed. The levator veli palatini that runs laterally towards the soft palate is now visible. The arrow indicates the dissected superior pharyngeal constrictor muscle; note the levator veli palatini muscles at both sides. E: ear, St: stylohyoid muscle, VII: facial nerve, M: masseter muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, T: trachea, Es: esophagus, P: Pharynx, *levator veli palatini muscle.
