

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
Questo protocollo descrive l'isolamento delle cellule satelliti da muscoli della testa branchiomeric di un ratto 9 settimane di età. I muscoli provengono da diversi archi branchiali. Successivamente, le cellule satelliti sono coltivate su un rivestimento macchia di dimensioni millimetriche per studiare la loro differenziazione. Questo approccio evita l'espansione e passaging di cellule satelliti.
Introduction
Circa 1: 500-1: 1000 neonati presentano una fenditura che coinvolge il labbro e / o palato (CLP); Così questo è la malformazione congenita più comune negli esseri umani 1. I muscoli del palato molle sono fondamentali per il funzionamento del palato molle durante il discorso, deglutizione e succhiare. Se una fessura del palato molle è presente, questi muscoli sono anormalmente inseriti nell'estremità posteriore dell'osso palatale.
Il palato molle si muove su e giù durante il discorso, impedendo che l'aria di fuggire attraverso il naso. I bambini con una fenditura nel palato non hanno questa funzione di controllo con conseguente fenomeno noto come velopharyngeal disfunzione 2,3. Anche se i protocolli di trattamento sono variabili, la riparazione chirurgica del palato molle si svolge nella prima infanzia (6-36 mesi di età) 4. I muscoli anormalmente inseriti del palato molle possono essere corretti chirurgicamente 5-7, tuttavia, disfunzione velopharyngeal persiste nel 7% al 30%dei pazienti 2,3,8-10.
La capacità del muscolo scheletrico di rigenerare attraverso l'azione di cellule satelliti (SCS) è ben consolidata 11,12. Su infortunio muscolare, SC sono attivati e migrano al sito di lesione. Poi proliferano, si differenziano e si fondono per formare nuove fibre muscolari o riparare danneggiati quelle 13. Quiescenti CS esprimono il fattore di trascrizione Pax7 14,15, mentre la loro progenie, i mioblasti proliferanti, inoltre esprimere il fattore miogenico determinazione 1 (MyoD) 16. Mioblasti differenziazione iniziano a esprimere miogenina (MyoG) 17. La differenziazione terminale dei mioblasti è caratterizzato dalla formazione di miofibre, e l'espressione di proteine muscolo-specifici come catena pesante della miosina (MyHC) 16,18.
Recentemente, diverse strategie sono state utilizzate nella medicina rigenerativa per migliorare la rigenerazione muscolare dei muscoli degli arti 19-23. Studi specifici sumuscoli della testa branchiomeric sono importanti anche perché è stato recentemente dimostrato che si differenziano da altri muscoli in diversi aspetti 24. In contrasto con i muscoli degli arti, è stato suggerito che i muscoli della testa branchiomeric contengono meno SC 25, rigenerano più lento, e il tessuto connettivo fibroso più si forma dopo l'infortunio 26 Inoltre, proliferando SC da muscoli della testa branchiomeric esprimere anche altri fattori di trascrizione. Ad esempio, Tcf21, un fattore di trascrizione per la formazione del muscolo craniofacciale è fortemente espresso in rigenerare muscoli della testa, ma difficilmente a rigenerare i muscoli degli arti 25. I muscoli del palato molle dei pazienti CLP sono solitamente più piccoli e meno ben organizzata rispetto ai normali muscoli palatini 27,28. Fibre lente e veloci sono entrambi presenti nei muscoli del palato molle ma le fibre lente sono più abbondanti. Al contrario, i muscoli leporino contengono una percentuale di fibra veloci e anche una fornitura capillare ridottarispetto ai normali muscoli del palato molle 29-31. Le fibre veloci sono maggiormente soggetti a danni contrazione indotta 31-33. Il povero fornitura capillare di accompagnamento può anche favorire la fibrosi 34,35. Tutti questi aspetti possono contribuire alla scarsa rigenerazione dei muscoli del palato molle dopo la chiusura chirurgica schisi 36. In considerazione di ciò, un protocollo per l'isolamento e la caratterizzazione di muscolo testa branchiomeric SC è cruciale. Questo offre la possibilità di studiare la biologia SC dei muscoli della testa branchiomeric. Inoltre, le nuove terapie a base di ingegneria dei tessuti possono essere sviluppate per promuovere la rigenerazione muscolare dopo l'intervento chirurgico in CLP e le altre condizioni che compromettono la zona cranio-facciale.
In generale, SC possono essere ottenuti dopo la dissociazione del tessuto muscolare 14. Tritare, digestione enzimatica, e triturazione sono generalmente tenuti a rilasciare SC dalla loro nicchia. SC può essere purificato mediante pre-placcatura su piatti non rivestite 14,37,38, fractionation su Percoll 39,40, cellulare o fluorescente-o magnetico di ordinamento 41-43. Qui vi presentiamo un nuovo protocollo economica e rapida per l'isolamento delle cellule satelliti da muscoli della testa branchiomeric di giovani ratti adulti. Questo protocollo è basato su una precedente manoscritto 14 e specificamente idonei per piccoli campioni di tessuto. L'isolamento di SC da muscoli rappresentativi provenienti dal 1 °, 2 °, e 4 ° archi branchiali sono descritti. Dopo l'isolamento, un basso numero di cellule satelliti sono coltivate su extracellulari macchie gel matrice di dimensioni millimetriche per studiare la loro differenziazione. Questo approccio consente di evitare l'obbligo per l'espansione e passaging di SC.
Protocol
Tutti gli esperimenti qui descritti sono stati approvati dal Consiglio locale per esperimenti su animali dalla Radboud University Nijmegen in conformità alle leggi e ai regolamenti (RU-dicembre 2013-205) Olandese.
1. Spot matrice extracellulare Gel
- Effettuare le seguenti operazioni un giorno prima l'isolamento:
- Scongelare una aliquota extracellulare matrice gel (100 microlitri) a 4 ° C per almeno 1,5 ore. Diluire 1:10 in modificata mezzo di Eagle Dulbecco; con 4.500 mg / L di glucosio, 4 mM L-glutammina, e 110 mg / ml di sodio piruvato (DMEM). Mantenere il gel matrice extracellulare a 4 ° C in ogni momento. Nota: sbalzi termici si tradurrà in rivestimento irregolare e formazione di cristalli.
- Conservare la soluzione di gel matrice extracellulare diluito in ghiaccio per 15 min.
- Pre-raffreddare una micropipetta 20 microlitri per 10 min.
- Mettere vetrini camera 8 pozzetti in una piastra di Petri 100 mm e trasferire il piatto su una superficie fredda (ad esempio un pacchetto congelatore) per 10 min.
- Utilizzare la micropipetta pre-raffreddata a mettere una goccia di 10 ml di gel matrice extracellulare in ogni pozzetto. Mantenere il piatto Petri sulla superficie fredda per almeno un altro 7 min (Figura 1A).
- Completamente rimuovere il residuo di gel matrice extracellulare (Figura 1B), e asciugare i pozzetti a 37 ° C durante la notte.
2. La dissezione di testa Muscoli (Massetere, digastrico, e muscolo elevatore del velo palatino)
- Prima di dissezione, preparare 50 ml di tampone fosfato salino (PBS) integrato con il 2% penicillina-streptomicina (P / S). Tenere su ghiaccio.
- Dopo l'eutanasia di un giovane ratto adulto (9 settimane) con CO 2 / O 2, decapitare il capo e togliere la pelle dalla testa. Trasferire la testa di PBS ghiacciato completato con 2% P / S in un tubo da 50 ml.
- Muscolo masticatore (derivato dal 1 ° arco branchiale)
- Mettere la testa con un lato su un tappetino di silicone e fissare con ipodermico needles (Figura 2A).
- Identificare la ghiandola parotide e il nervo facciale (Figura 2A). Esporre la fascia profonda che copre la ghiandola. Tagliare la fascia e rimuovere la ghiandola con le forbici dissezione. Identificare il canale uditivo esterno. Tracciare il nervo facciale dal forame stylomastoid e rimuovere con attenzione i rami temporali, zigomatico, e vestibolari con un bisturi lama n ° 15.
- Liberare il capo superficiale del muscolo massetere rimuovendo la fascia. Identificare teste sia superficiali e profonde del muscolo massetere. Tracciare la testa superficiale fino al suo spessore aponeurosi tendinea inserito nel processo zigomatico del mascellare superiore.
- Separare il tendine dalla sua origine al processo zigomatico con una pinza diritte. Tagliare con una lama di bisturi No. 15 o forbici dissezione e accuratamente la vita (Figura 2B).
- Sezionare il capo superficiale del massetere fino al suo inserimento con l'angolo e la metà inferiore di tha superficie laterale del ramo della mandibola con una lama di bisturi No. 15 (Figura 2C). Ora, rimuovere completamente il muscolo.
- Ventre posteriore del muscolo digastrico (derivato dal arco branchiale 2 °)
- Posizionare la testa in una posizione supina sul pad silicone e fissare con aghi ipodermici (Figura 3A).
- Rimuovere il grasso sottocutaneo sovrastante sia sublinguale e ghiandole sottomandibolari. Quindi, rimuovere la fascia superficiale e ghiandole con le forbici dissezione. Esporre il muscolo digastrico (anteriore e ventre posteriore).
- Tenere il tendine anteriore del ventre posteriore con una pinza diritte, tagliare e sezionare con cura fino alla sua origine nella bolla timpanica (Figura 3B). Fare la stessa cosa sul lato controlaterale.
- Muscolo elevatore del velo palatino muscolare (derivato dal 4 ° arco branchiale)
- Dopo la dissezione del ventre posteriore del muscolo digastrico, Localizzare il muscolo stiloioideo, tirare lateralmente, e rimuovere con cautela (Figura 4A).
- Localizzare il tendine del velo palatino elevatore che inserisce alla bolla timpanica (figura 4A). Sezionare attentamente e tagliare su entrambi i lati.
- Cercare la trachea e l'esofago che corre dietro. Sollevare l'esofago, ed esporre la faringe, laringe e il palato molle.
- Localizzare la zona del palato molle in cui è inserito il palatino elevatore veli e tagliarlo sciolti (Figura 4B).
Nota: Subito dopo la dissezione, rimuovere con attenzione tendine e il tessuto connettivo di ogni muscolo sotto il microscopio stereo. Immergere tutti i campioni rapidamente in etanolo al 70%, e trasferirli PBS ghiacciato integrato 2% P / S in una provetta da 15 ml.
3. Isolamento di cellule satellite
- Effettuare le seguenti operazioni di preparazione per SC isolamento da 3 gruppi di muscoli:
- Preparare 7,5 ml di 0,1% Pronasi in DMEM. Filtrare la soluzione attraverso un filtro di 0,22 micron. Pre-riscaldare la soluzione a 37 ° C a bagnomaria per 10 minuti prima di isolamento.
- Preparare 35 ml di DMEM supplementato con 10% siero di cavallo (HS) e 1% P / S. Pre-calda anche a 37 ° C in un bagno d'acqua.
- Preparare 15 ml terreno di coltura costituito da DMEM integrato con 20% siero bovino fetale (FBS), 10% HS, 1% P / S e 1% di estratto di embrione di pollo (CEE). Pre-calda a 37 ° C in un bagno d'acqua.
- Pre-coat sei pipette di plastica (10 ml) con HS e asciutti per almeno 10 minuti prima dell'uso.
- Nella cappa coltura, trasferire ogni muscolo in un pozzetto di una piastra da 6 pozzetti. Utilizzando le forbici dissezione, tagliare il muscolo in piccoli pezzi di circa 2 mm. Fare attenzione a non sminuzzare il tessuto troppo.
- Aggiungere con cautela 2,5 ml di soluzione pronase 0,1% in ciascun pozzetto ed incubare a 37 ° C per 60 min. Agitare delicatamente la piastra dopo 20, 40, e 60 min. Nota: Il duratio esatton dell'incubazione dipende da fattori quali l'età e la tensione degli animali.
- Monitorare al microscopio. Controllare i frammenti muscolari e fermare la digestione enzimatica quando le fibre fasci ottenere un aspetto allentata (Figura 5).
- Aggiungere 2,5 ml di DMEM supplementato con 10% HS e 1% P / S. Trasferire in un tubo da 15 ml e centrifugare le provette a 400 xg per 5 min. Eliminare il surnatante tramite decantazione.
- Aggiungere 5 ml DMEM supplementato con 10% HS e 1% P / S. Pipettare la soluzione su e giù con una pipetta 10 ml di plastica (triturazione) per almeno 20 volte per omogeneizzare il tessuto.
- Centrifugare le provette a 200 xg per 4 minuti. Raccogliere il surnatante e trasferire in una provetta da 15 ml.
- Aggiungere 5 ml DMEM supplementato con 10% HS e 1% P / S. Pipetta nuovo con una pipetta da 10 ml fino i frammenti di tessuto passa facilmente attraverso la pipetta.
- Centrifugare le provette a 200 xg per 4 minuti e raccogliere il surnatante in una provetta da 15 ml.
- Put un filtro cella (40 micron) su un tubo da 50 ml e trasferire il surnatante contenente le cellule dissociate sul filtro. Lavare con 1 ml DMEM per il recupero delle cellule massima.
- Centrifugare le provette a 1000 xg per 10 minuti e scartare il surnatante con una pipetta.
- Risospendere il pellet in 300 microlitri terreno di coltura e contare le cellule in un emocitometro.
4. La differenziazione delle cellule satelliti su Spot matrice extracellulare Gel
- Diluire la sospensione cellulare per ottenere 1,5 x 10 3 cellule in 10 ml di terreno di coltura.
- Fissare le coperture delle camere diapositive con nastro adesivo e segnare i punti con un pennarello nero sul lato inferiore del vetro dell'oggetto.
- Usando una micropipetta, mettere una goccia di sospensione cellulare 10 microlitri sul punto di gel matrice extracellulare. Controllare al microscopio se la goccia di sospensione cellulare è stato posizionato correttamente sul posto. Incubare per sei ore a 37 ° C.
- Attentoly aggiungere 400 ml di terreno di coltura (DMEM supplementato con 20% FBS, 10% HS, 1% P / S e 1% CEE) e incubare per tre giorni a 37 ° C.
Nota: A questo punto, appena isolato SC sono sottoposti a massicci traumi (digerito enzimatico e aspra triturazione), e hanno bisogno di recuperare. Non disturbare le cellule durante i primi tre giorni 37. Successivamente, il terreno di coltura può essere cambiato a seconda del tipo di esperimento.
Le macchie gel matrice extracellulare possono essere seminati con una densità cellulare di alta (1,5-2,5 x 10 3/20 ml) per il dosaggio differenziazione. Il terreno di coltura (DMEM integrato con 20% FBS, 10% HS, 1% P / S e 1% di estratto embrionale pollo) può essere sostituito ogni tre giorni. - In alternativa, se si desidera la crescita e il passaggio seguire i seguenti passi:
- Scongelare una aliquota extracellulare matrice gel (500 microlitri) a 4 ° C per almeno 1,5 ore. Diluire 1:10 in DMEM e seguire le raccomandazioni di cui al punto 1.1.1.
- Pre-raffreddare una pipetta 10 ml per 10 minuti a 4 &# 176; C.
- Trasferire tre fiasche T75 su una superficie fredda (ad esempio un pacchetto congelatore) per 10 min.
- Usare la pipetta pre-raffreddata a mettere 1 ml di gel matrice extracellulare in ogni pallone. Verificare che la superficie è coperta completamente. Mantenere i flaconi su superficie fredda per almeno altri 7 minuti (Figura 1A).
- Completamente rimuovere il gel matrice extracellulare rimanente con una pipetta 10 ml, e asciugare i pozzetti a 37 ° C per 1 ora.
- Dopo il conteggio, risospendere le SCs appena isolate in 10 ml di terreno di coltura (DMEM supplementato con 20% FBS, 10% HS, 1% P / S e 1% di estratto embrionale pollo) e seme nelle fiasche T75 pre-rivestite.
- Dopo tre giorni, cambiare il supporto (ed ogni terzo giorno) fino all'80% di confluenza è raggiunto. Per passaging, lavare i fiaschi T75 tre volte con PBS. Successivamente aggiungere 1 ml di 0,25% soluzione tripsina e incubare per tre minuti a 37 ° C. Risospendere in 9 ml di terreno di coltura (DMEM integrato con 20% FBS, 10% HS, 1% P / Se 1% di estratto embrionale pollo) e centrifugare a 200 xg per 5 min. Scartare il surnatante. Dopo aver contato, risospendere 1 x 10 6 cellule in 1000 ml di terreno di coltura e di congelare le cellule.
Representative Results
Usando questo protocollo, il muscolo massetere (un lato) produce 0,8-1 x 10 6 cellule, il muscolo digastrico (posteriore pancia) produce 1,5-2 x 10 5 cellule, e le rese palatini muscolo elevatore veli 1-1,5 x 10 5 cellule. I rendimenti cellulari dipendono dal tipo di muscolo, affaticamento, e l'età dell'animale. Per confronto tra i tre gruppi muscolari, SC appena isolate sono state seminate alla stessa densità di cellule (1,5 x 10 3/10 ml). Direttamente dopo l'isolamento, più del 90% delle cellule appena isolate esprimere Pax7 (Figura 6).
Giorno 4, 7 e 10 colture sono state colorate con anticorpi contro Pax7, MyoD, MyoG e MyHC immunocolorazione. Cinque campi arbitrari sono stati contati per la cultura con un obiettivo 20X. Al giorno 4 Pax 7 e Myo D si esprime in tutti i gruppi muscolari (figure 6 e 7 e 8), ma la progenie di SatCs dal massetere e muscoli digastrici iniziano exprescantare miogenina prima del muscolo elevatore del velo palatino muscolare (Figura 9). Al giorno 10, l'espressione di MyoG è fortemente ridotta in tutti i gruppi (Figura 9). Pochi giorni dopo la semina sui punti gel matrice extracellulare, le cellule proliferanti cominciano a fondersi e formare miotubi multi-nucleate, che esprimono la catena miosina pesante. Piccoli miotubi sono chiaramente visibili al giorno 7 (Figura 10). Al giorno 10, spasmi dei miotubi può osservare (Video 1).

Figura 1:. Extracellulari macchie gel matrice in una diapositiva camera (A) per una facile manipolazione, posizionare la slitta camera 8-bene in un piatto da 100 millimetri di Petri. Dispensare 10 ml di gel matrice extracellulare in ciascuna camera e metterlo su una superficie fredda (7 min). (B) Camera slitta dopo il superamento della matrice extracellulare gel viene rimosso.

Figura 2:. Dissezione del muscolo massetere (A) testa dell'animale in una vista laterale. Ear (E), ghiandola parotide (P) e del nervo facciale (VII). (B) tendinee aponeurosi (Te) della testa superficiale del muscolo massetere (Ms) e muscolo temporale (T). Separare il tendine dalla sua inserzione con una pinza. (C) sezionare con cautela il muscolo fino al suo inserimento al ramo della mandibola. E: orecchio, P: ghiandola parotide, VII: nervo facciale, T: muscolo temporale, Ms: capo superficiale del muscolo massetere, Te: tendine, Mp: profondo capo del muscolo massetere.

Figura 3: La dissezione del ventre posteriore del muscolo digastrico (.A) Testa dell'animale in posizione supina. Localizzare ghiandola sottomandibolare (Sg), massetere (M), nervo facciale (VII) e muscolo sternocleidomastoideo (SCM). Rimuovere la ghiandola sottomandibolare. (B) Localizzare anteriore digastrico muscolo (AD) e ventre posteriore (PD). Con una pinza dritto, prendere il tendine anteriore del ventre posteriore, tagliare e sezionare con cura fino alla sua origine nella bolla timpanica (ty). E: orecchio, Sg: ghiandola sottomandibolare, VII: nervo facciale, M: massetere muscolare, SMC: muscolo sternocleidomastoideo, AD: anteriore ventre muscolare digastrico, PD: posteriore ventre muscolare digastrico, Ty: bolla timpanica.

Figura 4:. Dissezione del muscolo elevatore del velo palatino (A) Veduta generale dopo la dissezione del muscolo digastrico (ventre posteriore). Muscolo stiloioideo (St) e tendine del muscolo elevatorevelo palatino può essere localizzato. Nota la trachea (T) e dell'esofago (Es) esecuzione dietro. (B) Dopo aver sollevato la trachea e l'esofago faringe (P) è esposto. L'inserimento del palatini elevatore veli nel palato molle è ora visibile. La freccia indica il palato molle sezionato con i muscoli Palatini elevatore veli su entrambi i lati. E: orecchio, St: muscolo stiloioideo, VII: nervo facciale, M: massetere muscolare, AD: anteriore ventre muscolare digastrico, PD: posteriore ventre muscolare digastrico, T: trachea, Es: esofago, P: Faringe, * muscolo elevatore del velo palatino muscolare .

Figura 5: Aspetto del tessuto muscolare (A) prima e (B) dopo la digestione enzimatica con Pronasi. Si noti che fasci muscolari sembrano essere allentato dopo la digestione enzimatica.
Figura 6: Pax 7 immunostaining SCs appena isolate, applicati al gel matrice extracellulare alla fine di isolamento (circa 6 ore dopo la digestione del tessuto iniziale).. Cinque campi arbitrari sono stati contati utilizzando un obiettivo 10X con una media di 210 cellule per campo. Circa il 90% delle cellule sono Pax 7 positive. DAPI: blu, Pax7: rosso. Barra Scala, 100 micron.

Figura 7:. Pax 7, MyoD immunostaining Giorno 4, 7 e 10 colture sono state colorate con anticorpi contro Pax7 e MyoD immunocolorazione. (A - C) e (D - F) microfotografie rappresentative della giornata 4 e 7 le culture del muscolo massetere. (G e H + e MyoD + nuclei per campo microscopico è stato contato e espresso come percentuale del numero totale di nuclei (DAPI). DAPI: blu, Pax7: rosso, e MyoD: verde. Bilancia bar, 100 micron. Cliccate qui per vedere una versione più grande di questa figura.

Figura 8:. Distribuzione di Pax7 ± / MyoD ± in colture di cellule mononucleate in colture da massetere, digastrico e elevatore veli muscolare palatina (A - C) Giorno 4, 7 e 10 colture sono state colorate con anticorpi contro Pax7 e MyoD immunocolorazione. Il numero totale di cellule si basa sul numero totale di nuclei (DAPI). (D) la quantificazione dei dati Pax7 ± / MyoD ± cells. Clicca qui per vedere una versione più grande di questa figura.

Figura 9:. Immunostaining Myogenin Giorno 4, 7 e 10 colture sono state colorate con anticorpi contro Myogenin. (A - D) microfotografie rappresentativi di giorno 4 e 7 culture dal Palatino muscolo elevatore veli. (E) Il numero di MyoG + nuclei per campo microscopico stato contato ed espresso come percentuale del numero totale di nuclei (DAPI). Quantificazione (F) I dati delle cellule MyoG +. DAPI: blu, Myogenin: verde. Bilancia bar, 100 micron. Cliccate qui per vedere una versione più grande di questo figure.

Figura 10:. Miosina pesante catena immunostaining Day 4, 7 e 10 colture sono state colorate con anticorpi contro catena pesante della miosina (MyHC). Microfotografie rappresentativi di giorno 4, 7 e 10 culture dal digastrico (DIG) muscolare. Al giorno 7, piccoli miotubi sono presenti, mentre a giorni 10 miotubi lunghe e ben organizzati sono evidenti. Bilancia bar, 200 micron. Cliccate qui per vedere una versione più grande di questa figura.
Video 1:.. Myotube spasmi Esempi di due campi rappresentativi con miotubi contrazioni sono indicati per il giorno 10 culture di muscolo digastrico Cliccate qui per vedere il video.
Discussion
SC da diversi muscoli della testa branchiomeric sono stati isolati da una 9 settimane di età ratto Wistar e coltivate direttamente sulla matrice extracellulare spot gel senza previa espansione e passaging. Dopo l'isolamento, le cellule sono state contate e seminate alla stessa densità cellulare. Per l'isolamento parallelo di tre diversi muscoli, questo metodo richiede circa 4 ore. Per evitare la contaminazione cultura, un passaggio critico è il rapido lavaggio in alcool 70% dopo la dissezione dei muscoli.
Durante isolamento SC è importante tagliare il tessuto muscolare in piccoli pezzi (circa 2 mm), ma evitare troppa macinazione come questo si tradurrà in una piccola quantità di cellule a causa di danno cellulare. Inoltre, la durata della digestione enzimatica deve essere controllata attentamente al microscopio per evitare ulteriori danni. Lo scopo della digestione è dissociare le miofibre. Poiché più del 90% delle cellule isolate esprimono Pax7, senza ulteriore purificazione è necessaria (figure 6-8).Questo evita fasi di purificazione supplementare altri metodi come la pre-placcatura sui piatti non patinati 14,37,38, frazionamento su Percoll 39,40, cellulare o fluorescente-o magnetico di ordinamento 41,43. Per triturazione è essenziale per indurre taglio tra i frammenti di tessuto e l'apertura della punta della pipetta come questo permette lo sblocco meccanico del SC. Se la triturazione con una pipetta 10 ml (all'interno Punta di diametro 1 mm) è difficile, a 5 ml (all'interno punta Diametro: 2 mm) pipetta possono essere utilizzati prima. In alternativa, pipette Pasteur di vetro possono essere tagliate a diametro desiderato e possono essere usate. Questo metodo è semplice, efficace e consente l'isolamento simultaneo di SC da campioni di muscolo differenti.
Le piastre di coltura per SCs possono essere rivestiti con gelatina o collagene, ma i nostri studi precedenti mostrano che gel matrice extracellulare è molto meglio per il mantenimento del potenziale miogenico di collagene 38. Le macchie di gel matrice extracellularedimensioni millimetriche (10 microlitri / Ø 2 mm o 20 microlitri / Ø 4 mm) consente lo studio della proliferazione e differenziazione delle CS con un numero limitato di cellule. Per il saggio differenziazione circa 8 a 20 volte meno cellule sono obbligatori confrontati ad una piastra da 24 pozzetti (Ø 15,6 millimetri), e circa 80 a 200 volte meno rispetto a 35 mm di piastre Petri (Ø 35 mm) 14,38.
Dal momento che il gel matrice extracellulare è costoso, questo metodo è anche più conveniente. Inoltre, i vetrini camera possono essere sostituiti da copertura di plastica scivola per ridurre ulteriormente i costi. Per la preparazione del gel matrice extracellulare visto essiccazione overnight delle slitte camera è essenziale. Poiché i punti di gel matrice extracellulare sono trasparenti, è necessario segnare i punti sul lato inferiore con illuminazione posteriore. Le Camere vetrini vengono fissati in una capsula di Petri per una facile manipolazione. Ulteriore espansione coltura cellulare non è necessario, che offre la possibilità di studiare il SC di goccioliler muscoli o campioni di muscolo di piccole dimensioni. In alternativa, ad esempio per la PCR o muscolo costruisce se sono necessari più celle, le SCs appena isolate possono prima essere ampliato in fiasche T75 come indicato sopra.
SC isolato utilizzando questo protocollo non sono suscettibili di depurazione mediante citometria a flusso subito dopo l'isolamento. La digestione con pronasi provoca una digestione notevole della superficie antigeni 14. Siero di cavallo e siero fetale bovino che vengono utilizzati per la coltura delle cellule devono prima essere adeguatamente caratterizzati prima di isolamento, come diversi numeri di lotto interessati differenziale mioblasti proliferazione e differenziazione.
Negli ultimi anni, vi è un crescente interesse nei muscoli derivanti dagli archi branchiali e mesoderma testa (ad esempio, i muscoli estrinseci) 24. È stato chiaramente dimostrato che i muscoli della testa e degli arti possiedono proprietà altamente differenti. Muscolo masticatore da vecchi animali sembra ritain loro capacità rigenerativa in confronto con i muscoli degli arti 25,26. SC dai muscoli estrinseci in possesso di una solida capacità di proliferazione e differenziazione paragonabili a SC da muscoli della testa, e mostrano un potenziale attecchimento più grande di arto muscolare SC 24.
La composizione di distribuzione e miosina tipo di fibra varia tra i gruppi muscolari e anche tra le specie. Muscoli provenienti dal primo arco branchiale nell'uomo contengono fibre sia lente e veloci (sottotipi IIA e IIX), miosine neonatali e myosins tipici per lo sviluppo del muscolo cardiaco. Nei roditori questi muscoli contengono circa il 95% fibre veloci miosina IIA e IIB) 44-46. Gli studi sui muscoli aviaria mostrano che SC di tipi di fibre muscolari differenti variano nella capacità di differenziazione. SC da fibre veloci differenziano solo in fibre muscolari veloci, mentre SC da fibre lente possono differenziarsi in entrambi i tipi di fibra 47. Inoltre, la percentuale di SC nel muscolo velocefibre è inferiore a quello delle fibre muscolari lente 48,49. Ciò indica che la distribuzione tipo fibra deve essere preso in considerazione per studi sui muscoli nella zona craniofacciale. Simile ai muscoli palatoschisi, la LVP nei roditori contiene quasi esclusivamente fibre veloci 50. Per questo motivo, SC del LVP sono adatti per gli studi pre-clinici nel campo della palatoschisi.
Questo protocollo offre nuove possibilità per studiare SCs derivate dai muscoli della testa branchiomeric o altri piccoli muscoli o muscoli campioni più piccoli. Ciò faciliterà lo sviluppo di nuove terapie per migliorare la rigenerazione dei muscoli nella zona maxillofacciale in condizioni come palatoschisi ma anche in altre condizioni che interessano muscoli più piccoli.
Materials
| Name | Company | Catalog Number | Comments |
| Hypodermic Needle 25 G 0.5 x 25 m | BD Microlance | 300400 | |
| Dissecting scissors | Braun | BC154R | |
| Micro forceps straight | Braun | BD330R | |
| Surgical Scalpel Blade No. 15 | Swann-Morton | 0205 | |
| Alcohol 70% | Denteck | 2,010,005 | |
| Permanox Slide, 8 Chamber | Thermo Scientific | 177445 | |
| 6 well cell culture plate | Greiner bio-one | 657160 | |
| Cell Culture Dishes (100 x 20 mm) | Greiner bio-one | 664160 | |
| 15 ml sterile conical centrifuge tube | BD Biosciences | 352097 | |
| 50 ml sterile conical centrifuge tube | BD Biosciences | 352098 | |
| Cell strainer (40 μm) | Gibco | 431750 | |
| 10 ml serological pipette | Greiner bio-one | 607180 | |
| 20 µl FT20 | Greiner bio-one | 774288 | |
| Matrigel, Phenol-Red Free | BD Biosciences | 356237 | 10 ml |
| Pronase | Calbiochem | 53702 | 10 KU |
| Phosphate Buffered Saline | Gibco | 14190-144 | 500 ml |
| Dulbecco's Modified Eagle Medium, high glucose, GlutaMAX Supplement, pyruvate | Gibco | 10569-010 | 500 ml |
| Fetal Bovine Serum | Fisher Scientific | 3600511 | 500 ml |
| Horse Serum | Gibco | 26050088 | 500 ml |
| Penicillin-Streptomycin (10,000 U/ml) | Gibco | 15140-122 | 100 ml |
| Chicken Embryo Extract | MP Biomedicals | 2850145 | 20 ml |
References
- Gritli-Linde, A. Molecular control of secondary palate development. Developmental Biology. 301, 309-326 (2007).
- Marrinan, E. M., LaBrie, R. A., Mulliken, J. B. Velopharyngeal function in nonsyndromic cleft palate: relevance of surgical technique, age at repair, and cleft type. The Cleft Palate-Craniofacial Journal. 35, 95-100 (1998).
- Morris, H. L. Velopharyngeal competence and primary cleft palate surgery, 1960-1971: a critical review. The Cleft Palate Journal. 10, 62-71 (1973).
- Mossey, P. A., Little, J., Munger, R. G., Dixon, M. J., Shaw, W. C. Cleft lip and palate. Lancet. 374, 1773-1785 (2009).
- Boorman, J. G., Sommerlad, B. C. Musculus uvulae and levator palati: their anatomical and functional relationship in velopharyngeal closure. British Journal of Plastic Surgery. 38, 333-338 (1985).
- Bae, Y. C., Kim, J. H., Lee, J., Hwang, S. M., Kim, S. S. Comparative study of the extent of palatal lengthening by different methods. Annals of Plastic Surgery. 48, 359-362 (2002).
- Braithwaite, F., Maurice, D. G. The importance of the levator palati muscle in cleft palate closure. British Journal of Plastic Surgery. 21, 60-62 (1968).
- Inman, D. S., Thomas, P., Hodgkinson, P. D., Reid, C. A. Oro-nasal fistula development and velopharyngeal insufficiency following primary cleft palate surgery--an audit of 148 children born between 1985 and 1997. British Journal of Plastic Surgery. 58, 1051-1054 (2005).
- Phua, Y. S., de Chalain, T. Incidence of oronasal fistulae and velopharyngeal insufficiency after cleft palate repair: an audit of 211 children born between 1990 and 2004. The Cleft Palate-Craniofacial Journal. 45, 172-178 (1990).
- Kirschner, R. E., et al. Cleft-palate repair by modified Furlow double-opposing Z-plasty: the Children's Hospital of Philadelphia experience. Plastic and Reconstructive Surgery. 104, 1998-2010 (1999).
- Mauro, A. Satellite cell of skeletal muscle fibers. The Journal of Biophysical and Biochemical Cytology. 9, 493-495 (1961).
- Yablonka-Reuveni, Z. The skeletal muscle satellite cell: still young and fascinating at 50. The Journal of Histochemistry and Cytochemistry. 59, 1041-1059 (2011).
- Ten Broek, R. W., Grefte, S., Von den Hoff, J. W. Regulatory factors and cell populations involved in skeletal muscle regeneration. Journal of Cellular Physiology. 224, 7-16 (2010).
- Danoviz, M. E., Yablonka-Reuveni, Z. Skeletal muscle satellite cells: background and methods for isolation and analysis in a primary culture system. Methods in Molecular Biology. 798, 21-52 (2012).
- Seale, P., et al. Pax7 is required for the specification of myogenic satellite cells. Cell. 102, 777-786 (2000).
- Yablonka-Reuveni, Z., et al. The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Developmental Biology. 210, 440-455 (1999).
- Zammit, P. S., Partridge, T. A., Yablonka-Reuveni, Z. The skeletal muscle satellite cell: the stem cell that came in from the cold. The Journal of Histochemistry And Cytochemistry. 54, 1177-1191 (2006).
- Andres, V., Walsh, K. Myogenin expression, cell cycle withdrawal, and phenotypic differentiation are temporally separable events that precede cell fusion upon myogenesis. The Journal of Cell Biology. 132, 657-666 (1996).
- Fukushima, K., et al. The use of an antifibrosis agent to improve muscle recovery after laceration. The American Journal of Sports Medicine. 29, 394-402 (2001).
- Grefte, S., Kuijpers-Jagtman, A. M., Torensma, R., Von den Hoff, J. W. Skeletal muscle fibrosis: the effect of stromal-derived factor-1α-loaded collagen scaffolds. Regenerative Medicine. 5, 737-747 (2010).
- Jackson, W. M., Nesti, L. J., Tuan, R. S. Potential therapeutic applications of muscle-derived mesenchymal stem and progenitor cells. Expert Opinion on Biological Therapy. 10, 505-517 (2010).
- Sato, K., et al. Improvement of muscle healing through enhancement of muscle regeneration and prevention of fibrosis. Muscle, & Nerve. 28, 365-372 (2003).
- Tatsumi, R., Anderson, J. E., Nevoret, C. J., Halevy, O., Allen, R. E. HGF/SF is present in normal adult skeletal muscle and is capable of activating satellite cells. Developmental Biology. 194, 114-128 (1998).
- Stuelsatz, P., et al. Extraocular muscle satellite cells are high performance myo-engines retaining efficient regenerative capacity in dystrophin deficiency. Developmental biology. , (2014).
- Ono, Y., Boldrin, L., Knopp, P., Morgan, J. E., Zammit, P. S. Muscle satellite cells are a functionally heterogeneous population in both somite-derived and branchiomeric muscles. Developmental Biology. 337, 29-41 (2010).
- Pavlath, G. K., et al. Heterogeneity among muscle precursor cells in adult skeletal muscles with differing regenerative capacities. Developmental Dynamics. 212, 495-508 (1998).
- Koo, S. H., Cunningham, M. C., Arabshahi, B., Gruss, J. S., Grant, J. H. 3rd The transforming growth factor-beta 3 knock-out mouse: an animal model for cleft palate. Plastic and Reconstructive Surgery. 108, 938-948 (2001).
- Fara, M., Brousilova, M. Experiences with early closure of velum and later closure of hard palate. Plastic and Reconstructive Surgery. 44, 134-141 (1969).
- Lindman, R., Paulin, G., Stal, P. S. Morphological characterization of the levator veli palatini muscle in children born with cleft palates. The Cleft Palate-Craniofacial Journal. 38, 438-448 (2001).
- Hanes, M. C., et al. Contractile properties of single permeabilized muscle fibers from congenital cleft palates and normal palates of Spanish goats. Plastic and Reconstructive Surgery. 119, 1685-1694 (2007).
- Rader, E. P., et al. Contraction-induced injury to single permeabilized muscle fibers from normal and congenitally-clefted goat palates. The Cleft Palate-Craniofacial Journal. 44, 216-222 (2007).
- Rader, E. P., et al. Effect of cleft palate repair on the susceptibility to contraction-induced injury of single permeabilized muscle fibers from congenitally-clefted goat palates. The Cleft Palate-Craniofacial Journal. 45, 113-120 (2008).
- Macpherson, P. C., Dennis, R. G., Faulkner, J. A. Sarcomere dynamics and contraction-induced injury to maximally activated single muscle fibres from soleus muscles of rats. The Journal of Physiology. 500 (Pt 2), 523-533 (1997).
- Koch, K. H., Grzonka, M. A., Koch, J. The pathology of the velopharyngeal musculature in cleft palates). Annals of Anatomy. 181, 123-126 (1999).
- Fara, M., Dvorak, J. Abnormal anatomy of the muscles of palatopharyngeal closure in cleft palates: anatomical and surgical considerations based on the autopsies of 18 unoperated cleft palates. Plastic and Reconstructive Surgery. 46, 488-497 (1970).
- Carvajal Monroy, P. L., Grefte, S., Kuijpers-Jagtman, A. M., Wagener, F. A., Von den Hoff, J. W. Strategies to Improve Regeneration of the Soft Palate Muscles After Cleft Palate Repair. Tissue Engineering. Part B, Reviews. , (2012).
- Grefte, S., Kuijpers, M. A., Kuijpers-Jagtman, A. M., Torensma, R., Von den Hoff, J. W. Myogenic capacity of muscle progenitor cells from head and limb muscles. European Journal of Oral Sciences. 120, 38-45 (2012).
- Grefte, S., Vullinghs, S., Kuijpers-Jagtman, A. M., Torensma, R., Von den Hoff, J. W. Matrigel but not collagen I, maintains the differentiation capacity of muscle derived cells in vitro. Biomedical Materials. 7, 055004 (2012).
- Kastner, S., Elias, M. C., Rivera, A. J., Yablonka-Reuveni, Z. Gene expression patterns of the fibroblast growth factors and their receptors during myogenesis of rat satellite cells. The Journal of Histochemistry and Cytochemistry. 48, 1079-1096 (2000).
- Yablonka-Reuveni, Z., Quinn, L. S., Nameroff, M. Isolation and clonal analysis of satellite cells from chicken pectoralis muscle. Developmental Biology. 119, 252-259 (1987).
- Sherwood, R. I., et al. Isolation of adult mouse myogenic progenitors: functional heterogeneity of cells within and engrafting skeletal muscle. Cell. 119, 543-554 (2004).
- Gilbert, P. M., et al. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science. 329, 1078-1081 (2010).
- Motohashi, N., Asakura, Y., Asakura, A. Isolation culture, and transplantation of muscle satellite cells. Journal of Visualized Experiments. , (2014).
- Sciote, J. J., Horton, M. J., Rowlerson, A. M., Link, J. Specialized cranial muscles: how different are they from limb and abdominal muscles. Cells, Tissues, Organs. 174, 73-86 (2003).
- Rowlerson, A., Mascarello, F., Veggetti, A., Carpene, E. The fibre-type composition of the first branchial arch muscles in Carnivora and Primates. Journal of Muscle Research and Cell Motility. 4, 443-472 (1983).
- Muller, J., et al. Comparative evolution of muscular dystrophy in diaphragm, gastrocnemius and masseter muscles from old male mdx mice. Journal of Muscle Research and Cell Motility. 22, 133-139 (2001).
- Feldman, J. L., Stockdale, F. E. Skeletal muscle satellite cell diversity: satellite cells form fibers of different types in cell culture. Developmental Biology. 143, 320-334 (1991).
- Schmalbruch, H., Hellhammer, U. The number of nuclei in adult rat muscles with special reference to satellite cells. The Anatomical Record. 189, 169-175 (1977).
- Gibson, M. C., Schultz, E. The distribution of satellite cells and their relationship to specific fiber types in soleus and extensor digitorum longus muscles. The Anatomical Record. 202, 329-337 (1982).
- Carvajal Monroy, P. L., et al. A rat model for muscle regeneration in the soft palate. PloS One. 8, e59193 (2013).
Tags
Biologia dello Sviluppo Numero 101 muscoli Head muscolo elevatore del velo palatino muscolare muscolo digastrico massetere cellule satelliti cellule primarie di isolamento palatoschisi la medicina rigenerativa l'ingegneria dei tessuti cellule staminali differenziazione myofibersErratum
Formal Correction: Erratum: Isolation and Characterization of Satellite Cells from Rat Head Branchiomeric Muscles
Posted by JoVE Editors on 10/01/2015.
Citeable Link.
An erratum was issue for Isolation and Characterization of Satellite Cells from Rat Head Branchiomeric Muscles. The fourth figure was updated to explain the isolation of the LVP better.
Steps 2.5.3 and 2.5.4 were updated from:
2.5.3. Look for the trachea and the esophagus that runs behind it. Lift the esophagus, and expose the pharynx, larynx and the soft palate.
2.5.4. Localize the area of the soft palate where the levator veli palatini is inserted and cut it loose (Figure 4B).
to
2.5.3. Look for the trachea and the esophagus that runs behind it. Lift the esophagus, and expose the pharynx and the larynx.
2.5.4 Localize and dissect the area of the superior pharyngeal constrictor muscle. Identify the levator veli palatini and cut it at both sides (Figure 4B).
Figure 4 and its legend were updated from:
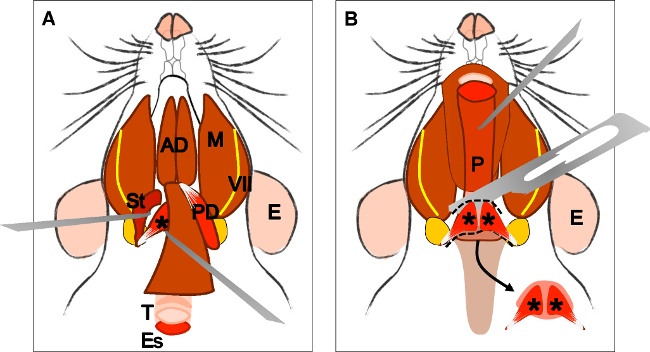
Figure 4: Dissection of the levator veli palatini muscle. (A) General view after dissection of the digastric muscle (posterior belly). Stylohyoid muscle (St) and tendon of the levator veli palatini can be localized. Note the trachea (T) and esophagus (Es) running behind it. (B) After lifting the trachea and the esophagus the pharynx (P) is exposed. The insertion of the levator veli palatini into the soft palate is now visible. The arrow indicates the dissected soft palate with the levator veli palatini muscles at both sides. E: ear, St: stylohyoid muscle, VII: facial nerve, M: masseter muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, T: trachea, Es: esophagus, P: Pharynx, *levator veli palatini muscle.
to
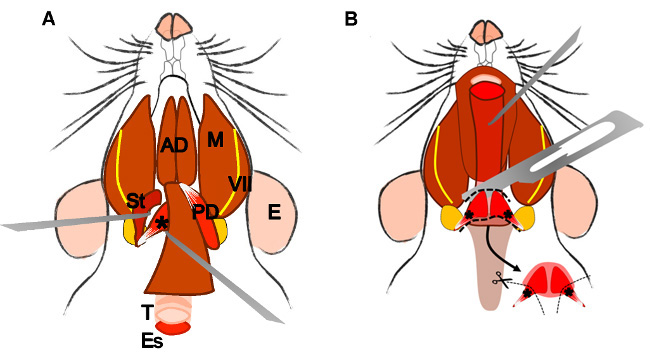
Figure 4: Dissection of the levator veli palatini muscle. (A) General view after dissection of the digastric muscle (posterior belly). Stylohyoid muscle (St) and tendon of the levator veli palatini can be localized. Note the trachea (T) and esophagus (Es) running behind it. (B) After lifting the trachea and the esophagus the pharynx (P) is exposed. The levator veli palatini that runs laterally towards the soft palate is now visible. The arrow indicates the dissected superior pharyngeal constrictor muscle; note the levator veli palatini muscles at both sides. E: ear, St: stylohyoid muscle, VII: facial nerve, M: masseter muscle, AD: anterior belly digastric muscle, PD: posterior belly digastric muscle, T: trachea, Es: esophagus, P: Pharynx, *levator veli palatini muscle.
