

Introduction
Myxovirus direnci (Mx) proteinler, viral patojenlere karşı doğal bağışıklık savunma önemli bir parçasıdır. Bu proteinler tip I ve tip III interferonlar tarafından indüklenen büyük Dynamin benzeri GTPazlar bulunmaktadır. karşılık gelen Mx genleri, bir ya da birden çok kopya halinde hemen hemen tüm omurgalıların içinde bulunmaktadır ve bunların gen ürünleri Ortomiksoviridae (örn., influenza virüsü), Rhabdoviridae (örn., veziküler stomatit virüsü), Bunyaviridae (örneğin, de dahil olmak üzere, virüslerin bir geniş inhibe eder. la crosse virüsü) ve (örneğin, insan bağışıklık eksikliği virüsü-1), 1-4 Retroviridae. Bu proteinler virüslerin böyle geniş bir dizi tanımak nasıl görünürde herhangi paylaşılan birincil dizisi bu virüsler, motiflerin olmadan, belli değil. Potansiyel, onların viral hedefleri ile Mx proteinlerinin etkileşimi analiz diğer ana hücre faktörleri ile yüksek mertebeden kompleksleri içeren, t moleküler mekanizmalarını anlamak için yardımcı olacaktırşapka virüs ve ev sahipleri arasında silahlanma yarışında gelişmiştir.
Memeli Mx proteinleri ve viral hedefler arasındaki etkileşim, insan mxa için en yaygın çalışılmıştır. İnsan MxA ortomiksovirüsler influenza A ve Thogoto virüsü de dahil olmak üzere pek çok virüslerin çoğalmasını inhibe edebilmektedir. MxA böylece enfeksiyon 5 blok sonuçlanır nükleer girişini engelleyen Thogoto virüs ribonükleoprotein kompleksleri (vRNPs) bağlanmaktadır. MxA ve Thogoto virüs vRNPs arasındaki bu etkileşim, eş çökeltme ve ko-immünopresipitasyon deneyleri 6-9 ile gösterilmiştir. Mx proteinleri influenza A virüsleri engel Nasıl daha az açıktır. Önemli bir sorun, bir MX proteini ve bir grip gen ürünü arasındaki bir etkileşimi göstermek için açık değildir olmasıdır. Bir rapor influenza A virüs bulaşmış hücreleri 10 insan mxa ve NP proteini arasında bir etkileşim göstermiştir. Bu etkileşim sadece eş-immunopr tarafından gösterilen olabilirecipitation hücre etkileşimi, geçici ve / veya zayıf olduğunu düşündürmektedir, liziz önce çapraz bağlanma belirteci ditiyobis (sukkinimidil propiyonat) ile muamele edilmiş olsaydı. Daha yeni çalışmalar, farklı grip A suşları ayırıcı Mx duyarlılığı NP proteininin 11,12 kökeni ile tespit edildiğini göstermiştir. Bu doğrultuda, influenza A virüsleri kısmen NP proteini 13 özel kalıntıları mutasyona Mx kontrolünden kaçabilir. Böylece, ana Mx için influenza A virüs temel hedefi, büyük olasılıkla NP vRNP kompleksleri monte NP protein olduğunu göstermektedir. Ancak, bu daha yeni çalışmaların hiçbiri grip NP veya vRNPs ve ya insan mxa veya fare MX1 arasında bir etkileşim göstermiştir.
Yakın zamanda ilk kez gösterdi burada ayrıntılı olarak tarif edilen bir optimize edilmiş ko-immünopresipitasyon protokolü 14 ile grip NP ve fare MX1 proteini arasındaki etkileşim. Genel co-immunoprecipitation, protein-protein etkileşimi araştırmak için en sık kullanılan biyokimyasal yaklaşımlardan biridir. Bu doğal ortamda, protein-protein etkileşimlerini incelemek sağlar çünkü bu teknik, çoğu zaman, alternatif teknikler, örneğin, maya iki hibrid tercih edilir. Ilgi konusu olan proteinlerin karşı antikorların olup olmadığını ko-immünopresipitasyon endojen olarak ifade edilen proteinleri ile gerçekleştirilebilir. Alternatif olarak, ilgi duyulan proteinlerin transfeksiyon ya da enjeksiyon aracılığıyla hücre içinde eksprese edilebilir ve bir afinite etiketi kullanılabilir. Yukarıda belirtilen avantajlara ilave olarak, tarif edilen ko-immünopresipitasyon protokolü zayıf ve / veya geçici protein etkileşimleri saptanmasını sağlar. Bu optimize edilmiş protokolde ana bileşeni hücre parçalama tamponu içinde, N-etilmaleimit (NEM) eklenmesidir. NEM 6,5-7,5 arasında bir pH değerinde, örneğin sistein, mevcut serbest tiyol grupları ile reaksiyona giren bir alkile edici bir madde, sabit bir tiyo-ester oluşturmak üzere(Şekil 1). Yüksek bir pH'de, NEM da amino grupları ile reaksiyona giren ya da hidroliz 15 uğrayabilir. NEM tipik haliyle disülfid bağı oluşumunu önlemek ya da enzimatik aktivitesini inhibe etmek için, serbest tiyol gruplarını bloke etmek için kullanılır. Örneğin, NEM, genellikle sistein proteazlarıdır, ki desumoylating enzimler, bloke edilmesi için kullanılmaktadır. Grip proteinleri sumoylation viral proteinlere 16 arasındaki etkileşimi etkileyebileceği bildirilmiştir çünkü tarif edilen ko-immünopresipitasyon protokolde, NEM ilk liziz tamponuna dahil edildi. Beklenmedik, Milli Ekonomi Modeli'nde eklenmesi eş immunopresipitasyon influenza NP ve fare MX1 arasındaki etkileşimi belgelemek için anahtar olduğunu kanıtladı. Milli Ekonomi Modeli'nde eklenmesi NP-MX1 etkileşimini tespit etmek önemlidir nedeni belli değildir. Muhtemelen etkileşim çok geçici ve / veya zayıf. NEM MX1 belirli bir konformasyon, bir viral protein ya da bilinmeyen bir üçüncü alçı koruyarak, örneğin etkileşimi, stabilize olabilirDaimi. NEM gibi bir stabilize edici etkisi ribonükleotid redüktaz M1 ve inhibitör gemsitabin (F2dC) 17 arasındaki etkileşim için, örneğin, daha önce tespit edilmiştir. MX1 ve NP hem NEM tarafından değiştirilebilir çoklu sistein artıklarını ihtiva etmektedir. Örneğin, Rennie'yle ve ark. Tarafından yapılan bir çalışmada, sapsız MxA varyantı iyodoasetamit ile modifiye edilebilir, üç çözücüye maruz kalan sistein artıklarını ihtiva ettiğini ortaya çıkarmıştır. Serinler, bu kalıntıların mutasyona tâbi mXA enzimatik aktivitesini etkileyebilir fakat disülfitin aracı toplanmasını 18 engelledi değildi. Bu sisteinler MX1 muhafaza edilir, bu MX1 analog sisteinler NEM tarafından, bu etkinin de uyum ve çözünürlük gibi modifiye edilebileceğini de önermektedir. Buna ek olarak, NEM da MX1 anti-influenza aktivitesi için gerekli olan MX1, bir GTPaz aktivitesini etkiler ve böylece MX1 ve NP arasındaki etkileşimi sabitleme olabilir. Ancak, GTPaz acti tarihinde NEM doğrudan etkisiNEM grip NP ve GTPaz MX1 proteininin 14 inaktif mutantlar arasındaki etkileşimi tespit etmek için gerekli olduğu gibi MX1 ve vite, olası değildir. Açıkçası, daha fazla araştırma NP-MX1 etkileşim NEM etkisini çözülmeye ihtiyaç vardır.
Özet olarak, tarif edilen ko-immünopresipitasyon protokolü antiviral MX1 proteini ve bunun viral hedef influenza NP proteininin etkileşimini çalışma sağlar. Bu protokol, aynı zamanda, belirli protein şeklindeki stabilizasyonu bağımlı olan diğer zayıf ya da geçici etkileşimler üzerinde çalışmak için de kullanılabilir. Belirli düzenlemelere bağlı protein-protein etkileşimi gibi kalmodulin 19 kalsiyum-bağlayıcı proteinler, örneğin, daha önce tarif edilmiştir. Son olarak, NEM uygun rolü, aynı zamanda, eş çökeltme tahlilleri gibi protein-protein etkileşimlerini tespit diğer yöntemler de kullanılabilir.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Not: Aşağıdaki transfeksiyon ve ko-immünopresipitasyon protokolü 9 cm Petri kabı biçimi için kurulur. Diğer formatlar protokolü ölçekleme sonra da mümkündür.
1. Tohum insan embriyonik böbrek (HEK) 293T hücreleri
- % 10 fetal dana serumu, 2 mM L-glutamin, 0.4 mM Na-piruvat ile takviye edilmiş Dulbecco tadil edilmiş Eagle ortamı (DMEM) içinde 12 ml 9 cm'lik petri tabağı başına 1.2 x 10 6 hücre transfeksiyondan önce HEK293T hücreleri bir gün Seed, 0.1 mM temel olmayan amino asit, 100 U / ml penisilin ve 0.1 mg / ml streptomisin.
- Hücreler, 37 ° C'de 16 saat ve% 5 CO2 büyütün.
- Görme transfeksiyondan önce ters ışık mikroskobu ile morfoloji ve hücrelerin canlılığını kontrol edin. Hücreler uygun transfeksiyon etkinliği için alt konfluent olması gerekir.
HEK293T Hücreler 2. Kalsiyum-fosfat transfeksiyonu
Not: Kullanım 0.5-1 pCAXL-NP veya 9 cm tabak başına pCAXL-MX1 1-3 mikrogram ile birlikte boş pCAXL plazmid ug. Tüm örneklerde toplam plazmid DNA eşit miktarda kullanmak; Boş plazmid gerekirse ayarlayın.
- Aşağıdaki transfeksiyon tamponlar hazırlanması:
- 1.0 mM Tris-HCI, pH 8.0 ve 0.1 mM EDTA pH 8.0 konsantrasyonda Tris-EDTA (TE) hazırlayın.
- 25 mM HEPES konsantrasyonlarda BS / HEPES hazırlayın (5.96 g / L, 4- (2-hidroksietil) -1-piperazinetansülfonik asit), 274 mM NaCI (16 g / L), 10 mM KCI (0.74 g / L), 1.5 mM NaHPO 4 · 12H H2O (0.5 gr / L) ve 11.1 mM dekstroz (2 g / L). 7,05 pH ayarlayın.
- 1.25 M CaCI2 · 2H 2 O (183.8 g / L) ve 125 mM HEPES (29,79 g / L) arasında dağılım gösterebilir ve CaCl2 / HEPES hazırlayın. NaOH ile 7,05 pH ayarlayın.
- Kullanımdan önce 37 ° C 'de transfeksiyon tamponlar ısıtın.
- TE 600 ul plazmid DNA seyrelterek plazmid örnekleri hazırlanır.6 gözlü bir levhanın gözleri içine bu karışımları hazırlayın.
- Plazmid örnekleri damla damla şekilde CaCl2 / HEPES 150 ul ekleyin ve yukarı ve aşağı 3 kez pipetleme karıştırın.
- (750 ul TE + DNA + CaCl2 / HEPES) taze 6-çukurlu plaka içindeki Resim BS / HEPES tamponu 750 ul damla plazmid çözeltisi eklenerek damla transfeksiyon çözelti hazırlayın. Tam da içeren BS / HEPES tamponu üzerinde eşit plazmid çözümü dağıtın.
- 1000 RPM'de 90 sn için, bir plaka karıştırıcısı üzerinde transfeksiyon solüsyonu çalkalayın.
- Oda sıcaklığında 5 dakika süreyle inkübe karışımı.
- Transfeksiyon solüsyonu (1.5 mi) hücrelere damla damla ekleyin. Hücreler üzerinde transfeksiyon solüsyonu damla P1000 mikropipet kullanın. Tam 9 cm Petri kabı üzerine karışımı dağıtılır ve çok yavaşça plaka sallayın.
- 6 st için CO 2, 37 ° C'de kuluçkaya bırakılır ve% 5. Sonra aspirasyon orta kaldırmak ve hemen wi yerine12 ml taze, önceden ısıtılmış orta inci. Yavaşça hücre ayrışması önlenmiş, hücrelere taze ortam ilave edin. Bunun için, kuyunun kenarına pipet ucu tutun ve yavaşça orta dışarı itmek.
- 37 ° C'de ilave bir 16 saat daha kuluçkaya bırakılır ve% 5 CO2.
3. ko-immünopresipitasyon
Not: transfeksiyondan sonra eş-immunoprecipitation 24 saat gerçekleştirin.
- Düşük tuz liziz tamponuyla yıkandı ve yüksek tuzlu yıkama tamponu hazırlanması.
- NEM miktarının tartılması ve mutlak etanol içinde çözülmesiyle 2 M, N-etilmaleimit (NEM) bir stok çözeltisi hazırlandı. Kullanmadan önce taze NEM stok çözeltisi hazırlayın.
DİKKAT: NEM, çok toksik hazırlamak ve davlumbaz, bu stok solüsyonu kullanın. - 50 mM Tris-HCI, pH 8, 150 mM NaCl, 5 mM etilendiamintetraasetik asit (EDTA),% 1 NP-40 ve bir proteaz inhibitör kokteyli konsantrasyonlarda düşük tuz liziz tamponu (1 tablo çözülür50 ml lisiz tampon maddesi içinde) ihtiva eder. 25 mM (örneğin, 1:80 seyreltilir), bir son konsantrasyon elde edecek NEM ekleyin. Proteaz inhibitörleri ve NEM ekledikten sonra buz üzerinde tutun.
Not: Kullanmadan önce her zaman taze proteaz inhibitörleri ve NEM ekleyin. - 50 mM Tris-HCI, pH 8, 500 mM NaCI, 5 mM EDTA ve% 1 NP40 konsantrasyonlarda yüksek tuzlu yıkama tamponu hazırlayın. Yüksek tuz yıkama tamponu NEM içermediğini unutmayın.
- NEM miktarının tartılması ve mutlak etanol içinde çözülmesiyle 2 M, N-etilmaleimit (NEM) bir stok çözeltisi hazırlandı. Kullanmadan önce taze NEM stok çözeltisi hazırlayın.
- Hücre lizatlarının Hazırlanması.
- Ortamı çıkarın ve buz soğuğu fosfat tamponlu tuzlu su, 2 ml (PBS) ile hücreleri yıkayın. HEK293T hücreleri kolayca ayırmak gibi çok nazikçe, yıkama tamponu ekleyin.
- PBS çıkarın ve 9 cm Petri başına buz soğuk düşük tuz lizis tamponu 600 ul ekleyin.
- Buz üzerinde 20 dakika süreyle inkübe edin. Plakalar yatay tutulur lizis tamponu ile plaka yüzeyinin tam kapsama sağlamak için, emin olun. Yavaşça her 5 dk plakaları sallayın.
- 1.5 ml mikrosantrifüj hücre lizat toplayın4 ° C'de 3 dakika 16,000 x g'de için boru ve santrifüj çözünmeyen fraksiyonu pelet haline getirmek üzere.
- Yeni bir 1.5 mL mikrosantrifüj tüpüne hücre lizatı, yani çözünür fraksiyonu aktarın ve buz üzerinde tutun. Hemen etkileşim proteinlerin ayrışma önlemek için, ko-immünopresipitasyon protokolü ile devam edin. Lizatlarında proteolitik etkinliğini sınırlamak için buz üzerinde mümkün olduğu kadar ya da 4 ° C 'de, aşağıdaki adımları.
- Bağışıklık-komplekslerinin Nesil.
Not: Bu adımda, ilgi konusu bir protein, uygun antikor ile bağlıdır. NP MX1 etkileşimini incelemek için bir fare anti-NP monoklonal antikor kullanılır.- Her numune için, bir anti-NP monoklonal antikorun 2 ul ve düşük tuz lisiz tamponu (250 ul toplam hacim) 113 ul lizat 135 ul karıştırın. , Örneğin batı beneklemesi gibi daha fazla analiz için -20 ° C'de, kalan lisat saklayın edebilmiş Intera ifade seviyelerini belgeTransfekte edilmiş hücrelerde ölü m işbirliği yapmaktadır.
Not: Alternatif olarak, her bir lizattan için, örneğin, 400 ug (Bradford reaktifi ile, örneğin) lizat proteini konsantrasyonunu ölçmek ve toplam proteinin sabit bir miktar kullanılır. - 4 ° C'de bir döner tekerlek üzerinde 3 saat süre ile, antikor lizat karışımı inkübe edin. Bu aşama, bir gece boyunca inkübasyon için genişletilebilir.
- Her numune için, bir anti-NP monoklonal antikorun 2 ul ve düşük tuz lisiz tamponu (250 ul toplam hacim) 113 ul lizat 135 ul karıştırın. , Örneğin batı beneklemesi gibi daha fazla analiz için -20 ° C'de, kalan lisat saklayın edebilmiş Intera ifade seviyelerini belgeTransfekte edilmiş hücrelerde ölü m işbirliği yapmaktadır.
- Protein G, tanelerin hazırlanması.
Not: Protein G, boncuk sevk ve korunması için% 20 etanol içinde saklanır. Boncuk bulamaç normal olarak% 50 tanecikleri içerir ve immün komplekslerin immünopresipitasyon için kullanılmadan önce, bu boncuklar yıkandı gerekmektedir.- Her bir örnek için, örneğin, boncuklar, 50 ul, boncuk çamurunun 100 ul kullanın. Bir tüp içinde ko-immünopresipitasyon deneyinde bütün örnekler için gerekli olan tanelerin miktarının yıkayın. Boncuk-bulamaç pipetlenmesini kolaylaştırmak için 1 ml pipet ucu kesin.
- 8.000 x G proteini boncuk bulamaç santrifüjg ve 30 saniye süreyle 4 ° C'de. Etanol çözeltisi çıkarın ve düşük tuz lisis tampon maddesinin eşit hacmi ekleyin. Santrifüj protein G 8000 xg'de boncuk bulamaç ve 30 saniye süreyle 4 ° C'de yavaşça süpernatant kaldırmak. Bu yıkama adımı 3 kez tekrarlayın.
Not: proteaz inhibitörleri ya da NEM içermesi gerekmez boncuk yıkamak için kullanılan düşük tuz lizis tamponu. - Protein G, tanelerin hacim tahmin ve düşük tuz lisis tampon maddesinin eşit hacmi düşük tuz liziz tamponu içinde yeni bir% 50 boncuk bulamaç ekleyin.
- Her numune için, yeni bir 1.5 ml mikrosantrifüj tüpü içinde, boncuk çamurunun 100 ul transfer edilir ve kullanıma kadar buz üzerinde muhafaza edin. Bu boncuklar hızla tüpün dibine sediment gibi, bölünmeden önce boncuk bulamaç tekrar süspansiyon dikkatli olun.
- Protein G, boncuk ve bunların elüsyon ile immün komplekslerin immünopresipitasyonu.
- Santrifüj tüm boruları 30 sn 8,000 xg'de ve 4 ° C ve immunoprecipitation protein G boncuklar kullanmadan önceTüm örneklerde mevcut olan tanelerin eşit miktarda olduğu görsel inceleme ile kontrol edin. Gerekirse yeniden örneklerin ve santrifüj bazı tanelerin miktarını ayarlar. Süpernatantlar atın. Peletlendi protein G boncuk rahatsız etmemek için dikkatli olun.
- Kısaca immün kompleksleri santrifüj (örn., Bir antikor, 250 ul ile lizatları) 8000 x g, 30 saniye ve 4 ° C'de borunun alt kısmında tamamen örnek toplamak için. Protein G, boncuklar (50 ul) ile immün kompleksleri aktarın.
- 4 ° C'de bir döner tekerlek üzerinde 60 dakika kuluçkalayın. G proteini tanelerine proteinlerin spesifik olmayan bağlanmayı azaltmak için, bu immün kompleksleri boncuklar ile daha uzun bir süre 75 dakika inkübe etmeyin.
- 8,000 x g'de 30 saniye için (bağlı immün kompleksleri ile), protein G boncuk santrifüj ve 4 ° C ve süpernatantlar çıkarın. Peletlendi protein G boncuk rahatsız etmemek için dikkatli olun. İsteğe bağlı: veya -20, daha sonra analiz için C, 4 ° C 'de bu süpernatantlar depolamakÖrneğin, bağlanmamış protein miktarını tahmin etmek.
- Yüksek tuzlu liziz tamponu içinde 900 ul, yaklaşık 5 dakika boyunca, protein G boncuk yıkayın. Boncuklar bütünüyle optimal yıkama için, yıkama tampon maddesi içinde yeniden süspansiyon haline getirilmiş olduğundan emin olun. 8,000 x g'de 30 saniye için, protein G boncuk santrifüj ve 4 ° C ve süpernatantlar atın. Bu yıkama adım 4 kez tekrarlayın. Çökeltilir malzeme kaybını önlemek için topaklandı protein G boncuk rahatsız etmemek için dikkatli olun.
- Son yıkama aşamasından sonra, boncuklara 2x Laemmli numune tampon maddesi 50 ul ve (ko) imüno proteinleri elüte etmek için 95 ° C'de 10 dakika boyunca bir süspansiyon ısıtın.
- Sodyum dodesil sülfat, 1 g, gliserol ve 3.5 ml 1 M Tris-HCI pH 6.8 ve β-merkaptoetanol ve 420 ul 3.5 ml 6x Laemmli tampon maddesi 10 ml hazırlayın. Damıtılmış su ilave edilerek, 10 ml toplam hacim için ayarlayın. 2x Laemmli tamponu elde etmek üzere distile su içinde 3 kez seyreltilir.
DİKKAT: &# 946; mersaptoetanol toksik, hazırlamak ve davlumbaz Laemmli tampon kullanabilirsiniz.
- Sodyum dodesil sülfat, 1 g, gliserol ve 3.5 ml 1 M Tris-HCI pH 6.8 ve β-merkaptoetanol ve 420 ul 3.5 ml 6x Laemmli tampon maddesi 10 ml hazırlayın. Damıtılmış su ilave edilerek, 10 ml toplam hacim için ayarlayın. 2x Laemmli tamponu elde etmek üzere distile su içinde 3 kez seyreltilir.
- Isıtıldıktan sonra, 8.000 xg'de 30 saniye protein G boncuk santrifüj ve 4 ° C (kısa vadeli) ya da -20 ° C (uzun vadede) ° örnekleri saklamak.
4. immunopresipite Proteinler (co-) analiz
- SDS-PAGE 20 ve 21,22 lekeleme batı hücre lizatı ve ko-immunoprecipitation eluatını mevcut proteinleri gözünüzde canlandırın. Tipik jel üzerinde Laemmli elüatın yarısını yükleyin. Jel yükleme için numune alırken topaklandı protein G boncuk rahatsız etmemek için dikkatli olun. Mx1 ve NP ifade, anti-MX1 ve anti-NP antikoru, sırasıyla 14, bir ile ortaya çıkarılmıştır. bantlar HRP-tabanlı chemiluminescence ve bir X-ışını filmi geliştirici ile tespit edilmiştir.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
N-etilmaleimit sistein proteazlarını geri dönülmez (Şekil 1) önlenmesi için, örneğin, serbest tiyol gruplarını düzenlemek için kullanılabilecek bir organik bileşiktir.
Antiviral MX1 proteini influenza virüs nükleoprotein ile etkileşerek bir virüs çoğalmasını engeller. Burada anlatılan optimize ko-immünopresipitasyon protokolü bu NP-MX1 etkileşim çalışma sağlar. HEK293T hücreleri grip NP proteininin varlığı veya yokluğu antiviral MX1 bir protein için ifade vektörleri ile transfekte edilmiştir. Daha sonra, NP proteininin NP-spesifik monoklonal antikor ile toplam hücre lizatları çekilmiş aşağı oldu. 2 MX1 proteini sadece birlikte imüno-çökeltilmiştir birlikte ifade NP mevcudiyetinde olduğunu göstermektedir. NP yokluğunda MX1 olası spesifik olmayan ko-immünopresipitasyon anti-NP antikoru ile ya da MX1 spesifik olmayan bağlanma ile MX1 proteininin spesifik olmayan pull ya kaynaklanırProtein G, boncuk. Bu nedenle, her zaman bu, spesifik olmayan ko-immünopresipitasyon değerlendirmek için negatif bir kontrol içerir. 3 NP MX1 etkileşimi sadece NEM varlığında tespit edilebilir göstermektedir. Bu deneyde, protein G boncuklara MX1 spesifik olmayan bağlanma anti-NP antikorunun yokluğunda kontrol ko-immünopresipitasyon reaksiyonu ile değerlendirildi.
Bu protokol, aynı zamanda MX1 ve enfekte hücrelerden veya virionlar izole Grip NP protein arasındaki etkileşimi incelemek için kullanılabilir. Bu uygulama için yukarıdaki protokol biraz MX1 ko-immünopresipitasyon protokolü başlamadan önce viral NP protein içeren lizatları ile ifade eden hücrelerin lizatları birleştirilerek uyarlanmıştır. Şekil 4 NP MX1 ko-immünopresipitasyon transfekte edilmiş hücrelerin, enfekte olmuş hücrelerden izole gösterir veya virionlar.
Sonuç olarak, bu sonuçlar, bu ko-immünopresipitasyon korum olduğunu göstermektedirocol bir antiviral protein ve bunun viral hedefleri arasındaki etkileşimlerin incelenmesi için de kullanılabilir.
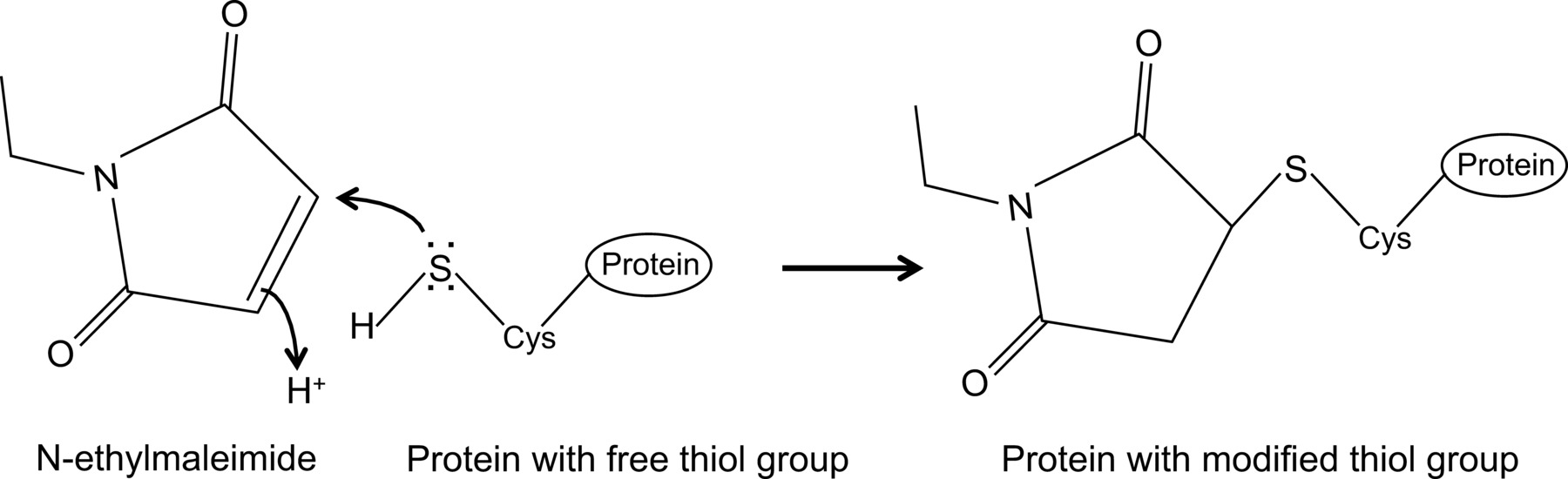
Şekil 1:. N-etilmaleimit serbest tiyol gruplarının Geri dönüşümsüz değişiklik bu rakamın daha büyük bir versiyonunu görmek için burayı tıklayınız.

MX1 ve NP her ikisi de mevcut olan bir olan ve NP proteininin mevcut olduğu, bir kontrol kurulum: Şekil 2:. Iki numune ile birlikte bir ortak immüno-deney MX1 NP ile etkileşime girer, Western blot analizi. NP, anti-NP ve NP ve MX1 ile immüno-çökeltildi batı lekeleme ile görselleştirilmiştir. Bu rakam f modifiye edilmişrom 14.

. Şekil 3: - MX1 etkileşim, N-etilmaleimit varlığında veya yokluğunda gerçekleştirilebilir, bir ko-immünopresipitasyon deneyi Western blot analizi, N-etilmaleimit önemlidir NP tespit etmek. NP, anti-NP ve MX1 ve NP ile immüno-çökeltildi batı lekeleme ile görselleştirilmiştir.

Şekil 4:., Influenza A virionlar izole enfekte hücreleri veya vRNPs gelen vRNPs, vRNPs transfekte edilmiş bir kontrol lizat (hiçbir vRNPs): MX1 ihtiva eden NP enfekte olmuş hücreler veya izole edilmiş virionlar ile MX1 etkileşime Lisatlar, farklı kaynaklardan gelen NP içeren lizatları ile bir araya getirilmiş . Lizatları, karıştırma eş immunopre sonraAnti-NP ile yağmurlama yapıldı ve MX1 ve NP batı lekeleme ile görselleştirilmiştir.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Antiviral proteinler ve bunların viral hedeflere arasındaki etkileşimi incelemek bu proteinlerin antiviral mekanizmasının ayrıntılarını anlaşılması çok önemlidir. Bu yeni antiviral stratejilerin geliştirilmesi için temel virüsler ve onların ana-evrimleşmiş eş nasıl yeni anlayışlar vermek ve olabilir. Burada anlatılan optimize ko-immünopresipitasyon protokol fare MX1 protein ve viral hedef, grip NP proteini arasındaki etkileşimi incelemek için izin verir. NP-MX1 etkileşimi NEM (Şekil 3) yokluğunda saptanamaz olduğu gibi bu protokolün en önemli yönü, lizis tamponunda NEM eklenmesidir. Bugüne kadar, bu NEM varlığı, bu etkileşimi saptamak için temel neden olarak bilinmemektedir. Ancak, bu protokol sisteinler dahil, özellikle, özel protein konformasyonlarının istikrar bağlıdır diğer zayıf ve / veya geçici etkileşimleri incelemek için yararlı olabilir.
BirGenel eş-İmmünopresipitasyon önemli kısıtlılığı yüksek afinite ile etkileşim ortaklarından biri de yapısal epitopları tanıyan özel ve kaliteli antikorların bulunmasıdır. NP-MX1 etkileşim evde anti-MX1 poliklonal antiserum üretilen bizim ile ortaya edilemedi. Bu antiserum, aynı zamanda da MX1 yokluğunda, influenza NP protein İmmünoçökeltiler. Ayrıca, kullanılan anti-NP monoklonal antikor, A / Porto Riko / 8/34 influenza suşu NP proteinini tanır, ama ne yazık ki kuş gribi virüsü suşları NP-aşağı çekmek için uygun değildir. Bu ko-immünopresipitasyon protokolünün bir diğer kısıtlılığı protein G boncuk MX1 nonspesifik bağlanma olduğunu. ikinci yıkama tamponu içinde tuz konsantrasyonunun arttırılması ve lizat ve protein G, boncuk arasındaki temas süresini azaltarak aşılabilir bağlanma. Buna ek olarak, tanelerin miktarının azaltılması her immüno-çökeltme için kullanılan25 ul, 50 ul reaksiyon, bundan başka, bu boncuklara MX1 spesifik olmayan azaltabilir. Genel olarak, protein G boncuklarına non-spesifik birleşmesinin sebep olduğu, kirletici proteinlerin istenmeyen açılan, diğer stratejiler ile azaltılabilir. Örneğin, bu proteinlerin, bu proteinlerin antikorun yokluğunda protein G boncuklarla lisat inkübe edilerek uzaklaştırılmış olan bir ön açık adım esnasında çıkarılabilir. kirletici proteinler daha sonra boncuklar ile birlikte çıkarılır ve önceden temizlenmiş lizat birlikte immüno-çökeltme için kullanılmaktadır. Kirletici proteinlerin çalışılan protein (ler) 'den farklı olan, bu strateji çok avantajlıdır. Seçenek olarak ise, protein ile spesifik olmayan bağlanma yerleri, G boncuklar BSA ile bloke edilebilir. Boncuklar birinci antikor ile kaplanmış ise, BSA da (örneğin, indirgenmiş antikor kaplı boncuk bağlama) immün komplekslerinin çökelmesini azaltır Ancak, bu strateji sadece tavsiye edilir. Takenbirlikte, protein G, boncuk ya da kullanılan antikorlara spesifik olmayan ilgi konusu olan proteinlerin bağlanmasının hariç tutmak için uygun kontroller dahil etmek çok önemlidir.
tarif edilen ko-immünopresipitasyon protokolü enfekte olmuş hücrelerde ya da saflaştırılmış vRNPs farklı ortamda mevcut NP proteininin, örneğin, ile birlikte MX1 etkileşimini incelemek için modifiye edilebilir. MX1 grip virüsü enfeksiyonu 23 boyunca, NP dahil olmak üzere viral proteinlerin ekspresyonunu inhibe eder. Bu nedenle, daha sonra enfekte edilir MX1 transfekte edilmiş hücrelerin NP MX1 etkileşimi incelemek için teknik olarak son derece zordur. Yine de, bu ko-immünopresipitasyon protokolü de MX1 NP MX1 etkileşim başarılı tespitine olanak sağlayan, (Şekil 4) hücreleri ve enfekte hücreleri ifade lizatları birleştirildikten sonra gerçekleştirilebilir. NEM önemli hedef bilinmemektedir gibi NEM her iki hücre popülasyonlarının lizizi sırasında ilave edildi. Arzu edildiği takdirde, pHliziz tamponu değiştirilebilir. Burada tarif edilen deneyler (Turan ve diğ. 10'daki gibi), pH 8 ile gerçekleştirilir fakat NP MX1 ko-immünopresipitasyon da başarılı bir şekilde, pH 7.2 liziz tamponu ile gerçekleştirildi. Bu pH değeri NEM serbest tiyol grupları ile özel olarak reaksiyona giren ve bu, pH, aynı zamanda MX1 proteininin ekstraksiyon verimi artırır de çünkü aslında 7.2 arasında bir pH tercih edilmektedir. Buna ek olarak, protein G boncuklar kullanılan antikorlar türetildiği ana-türe bağlı olarak, protein A boncuk ile ikame edilmiş olabilir. Bununla birlikte, daha temiz sonuçlar, protein G boncuklar ile elde edilir. Son olarak, bu protokol, aynı zamanda, örneğin MX1 ve grip proteinleri, PB2 14 arasındaki etkileşimi incelemek için kullanılabilir. Prensip olarak ticari olarak temin edilebilen bir anti-V5 agaroz afinite jeli ile kombine edilebilir bir V5 epitop PB2 kullanılan etiketli Bu durumda,.
Gelecekteki deneylerde, bu protokol MX1 bölgeleri tanımlamak için değerli olabilir veNP MX1 etkileşimi için önemli olan NP. Bu döngü mXA ve Thogoto virüsü NP 8,9 arasındaki etkileşim için kritik olduğu gösterilmiştir olarak MX1 bu tür bir bölge, halka L4 olabilir. Uygun NP-spesifik antikorlar kullanılabilir hale varsa, bu protokol de MX1 için kuş gribi A suşları artan duyarlılık Mx duyarlılık mekanizmasını anlamak için yardımcı olabilir güçlü bir NP-MX1 etkileşim ya da değil, korelasyon olup olmadığını belirlemek için izin verecektir. Buna ek olarak, MX1 ve Ortomiksoviridae dışındaki ailelere mensup virüsler bileşenleri arasındaki etkileşim, aynı zamanda bu uyarlanmış ko-immünopresipitasyon protokolü tarafından ele alınabilir. Son olarak, daha fazla çalışmaya bu özel etkileşimini anlamak için, çok değerli olacaktır NP MX1 ile NEM etkisini çözmek için değil, aynı zamanda, hücre lizizi sağlandı ve ko-immünopresipitasyon deneyler sırasında bu bileşiğin eklenmesi daha geniş uygulamalarda anlamak için.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarlar hiçbir rakip mali çıkarlarını olduğunu beyan ederim.
Acknowledgments
Bu çalışma FWO-Vlaanderen, IOF projesi IOF10 / StarTT / 027 ve Gent Üniversitesi Özel Araştırma Hibe BOF12 / GOA / 014 tarafından desteklenmiştir.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM high glucose | Gibco | 52100-047 | |
| N-Ethylmaleimide | Sigma | E-3876 | Toxic |
| Igepal CA-630 | Sigma | I-30212 | also known as NP40 |
| Protease inhibitor cocktail | Roche | 11 873 580 001 | |
| anti-NP monoclonal antibody | NIH Biodefense and Emerging Infections Research Resources Repository | NR-4282 | ascites blend of clones A1 and A3 |
| anti-RNP polyclonal serum | NIH Biodefense and Emerging Infections Research Resources Repository | NR-3133 | directed against A/Scotland/840/74 (H3N2) |
| Protein G Sepharose 4FF | GE Healthcare | 17-0618-01 | |
| Hyperfilm ECL 18 x 24 cm | GE Healthcare | 28-9068-36 | |
| ECL western blotting substrate | Pierce | 32106 |
References
- Verhelst, J., Hulpiau, P., Saelens, X. Mx proteins: antiviral gatekeepers that restrain the uninvited. Microbiol Mol Biol Rev. 77 (4), 551-566 (2013).
- Goujon, C., et al. Human MX2 is an interferon-induced post-entry inhibitor of HIV-1 infection. Nature. 502 (7472), 559-562 (2013).
- Kane, M., et al. MX2 is an interferon-induced inhibitor of HIV-1 infection. Nature. 502 (7472), 563-566 (2013).
- Liu, Z., et al. The interferon-inducible MxB protein inhibits HIV-1 infection. Cell Host Microbe. 14 (4), 398-410 (2013).
- Kochs, G., Haller, O. Interferon-induced human MxA GTPase blocks nuclear import of Thogoto virus nucleocapsids. Proc Natl Acad Sci U S A. 96 (5), 2082-2086 (1999).
- Flohr, F., Schneider-Schaulies, S., Haller, O., Kochs, G. The central interactive region of human MxA GTPase is involved in GTPase activation and interaction with viral target structures. FEBS Lett. 463 (1-2), 24-28 (1999).
- Kochs, G., Haller, O. GTP-bound human MxA protein interacts with the nucleocapsids of Thogoto virus (Orthomyxoviridae). J Biol Chem. 274 (7), 4370-4376 (1999).
- Mitchell, P. S., et al. Evolution-guided identification of antiviral specificity determinants in the broadly acting interferon-induced innate immunity factor MxA. Cell Host Microbe. 12 (4), 598-604 (2012).
- Patzina, C., Haller, O., Kochs, G. Structural requirements for the antiviral activity of the human MxA protein against Thogoto and influenza A virus. J Biol Chem. 289 (9), 6020-6027 (2014).
- Turan, K., et al. Nuclear MxA proteins form a complex with influenza virus NP and inhibit the transcription of the engineered influenza virus genome. Nucleic Acids Res. 32 (2), 643-652 (2004).
- Dittmann, J., et al. Influenza A virus strains differ in sensitivity to the antiviral action of Mx-GTPase. J Virol. 82 (7), 3624-3631 (2008).
- Zimmermann, P., Manz, B., Haller, O., Schwemmle, M., Kochs, G. The viral nucleoprotein determines Mx sensitivity of influenza A viruses. J Virol. 85 (16), 8133-8140 (2011).
- Manz, B., et al. Pandemic influenza A viruses escape from restriction by human MxA through adaptive mutations in the nucleoprotein. PLoS Pathog. 9 (3), e1003279 (2013).
- Verhelst, J., Parthoens, E., Schepens, B., Fiers, W., Saelens, X. Interferon-inducible protein Mx1 inhibits influenza virus by interfering with functional viral ribonucleoprotein complex assembly. J Virol. 86 (24), 13445-13455 (2012).
- Brewer, C. F., Riehm, J. P. Evidence for possible nonspecific reactions between N-ethylmaleimide and proteins. Anal Biochem. 18 (2), 248-255 (1967).
- Wu, C. Y., Jeng, K. S., Lai, M. M. The SUMOylation of matrix protein M1 modulates the assembly and morphogenesis of influenza A virus. J Virol. 85 (13), 6618-6628 (2011).
- Chen, Z., Zhou, J., Zhang, Y., Bepler, G. Modulation of the ribonucleotide reductase M1-gemcitabine interaction in vivo by N-ethylmaleimide. Biochem Biophys Res Commun. 413 (2), 383-388 (2011).
- Rennie, M. L., McKelvie, S. A., Bulloch, E. M., Kingston, R. L. Transient dimerization of human MxA promotes GTP hydrolysis, resulting in a mechanical power stroke. Structure. 22 (10), 1433-1445 (2014).
- Gifford, J. L., Walsh, M. P., Vogel, H. J. Structures and metal-ion-binding properties of the Ca2+-binding helix-loop-helix EF-hand motifs. Biochem J. 405 (2), 199-221 (2007).
- Separating Protein with SDS-PAGE. , JoVE. Available from: http://www.jove.com/science-education/5058/separating-protein-with-sds-page (2014).
- Gallagher, S., Chakavarti, D. Immunoblot analysis. J Vis Exp. (16), (2008).
- Eslami, A., Lujan, J. Western blotting: sample preparation to detection. J Vis Exp. (44), (2010).
- Pavlovic, J., Haller, O., Staeheli, P. Human and mouse Mx proteins inhibit different steps of the influenza virus multiplication cycle. J Virol. 66 (4), 2564-2569 (1992).
