

Summary
Her præsenterer vi en protokol til at opdage enkelt, SNARE-medieret fusion begivenheder mellem liposomer og understøttede dobbeltlag i mikrofluide kanaler ved hjælp polariseret TIRFM, med enkelt molekyle følsomhed og ~ 15 ms tidsopløsning. Lipid- og opløseligt fragt frigivelse kan detekteres samtidigt. Liposomstørrelse, lipid diffusivitet, og fusion pore egenskaber måles.
Abstract
I den allestedsnærværende proces med membranfusion åbningen af en fusion pore etablerer den første forbindelse mellem to tidligere adskilte rum. Under neurotransmitter eller hormon frigivelse via exocytose, fusion pore kan forbigående åbne og lukke flere gange, regulerer fragt release kinetik. Pore dynamik også bestemme tilstanden af vesikel genbrug; irreversible genforseglingsegenskaber resulterer i forbigående, "kiss-and-run" fusion, mens dilation fører til fuld fusion. For bedre at forstå, hvilke faktorer regulerer pore dynamik, vi udviklet et assay til overvågning membran fusion med polariseret total indre refleksion fluorescens (TIRF) mikroskopi med enkelt molekyle følsomhed og ~ 15 msek tidsopløsning i en biokemisk veldefineret in vitro-system. Fusion af fluorescensmærket små unilamellare vesikler indeholdende v-SNARE-proteiner (v-SUV'er) med en plan dobbeltlag bærende t-snarer, understøttet på en blød polymer pude (t-SBL, t-understøttet dobbeltlag), Overvåges. Assayet anvender mikrofluide strømningskanaler, der sikrer minimal prøve forbrug under tilførsel en konstant densitet på SUV'er. Udnyttelse af den hurtige signal forstærkning efter overførsel af lipid etiketter fra SUV til SBL under fusion, er kinetik overførsel lipid farvestof overvåges. Følsomheden af TIRF mikroskopi tillader sporing enkelt fluorescerende lipid etiketter, hvorfra lipid diffusivitet og SUV størrelse kan udledes for hver fusion begivenhed. Lipid gange farvestof release kan være meget længere end forventet for uhindret passage gennem permanent åbne porer. Ved hjælp af en model, der antager retardering af lipid frigivelse skyldes pore flimrende, en pore "åbenhed", den del af tiden pore forbliver åben under fusion, kan estimeres. Et opløseligt markør kan indkapsles i SUV'er til samtidig overvågning af lipid og opløseligt fragt frigivelse. Sådanne målinger indikerer nogle porer kan tillukkes efter at have tabt en del af den opløselige last.
Introduction
Membrane fusion er en universel biologisk proces kræves for intracellulær handel med lipider og proteiner, sekretion, gødskning, udvikling, og indhyllet virus indrejse i værtsorganismer 1-3. For de fleste intracellulær fusion reaktioner, herunder frigivelse af hormoner og neurotransmittere via exocytose, er energien til at smelte to lipid-dobbeltlag, som dannelsen af en fire-helix bundt mellem beslægtet opløseligt N-ethylmaleimid-følsomme faktor vedhæftet protein receptor (SNARE) proteiner, forankret i vesiklen (v-SNARE) og målmembranen (t-SNARE) 4. Synaptisk vesikel exocytose er den mest stramt reguleret fusionsreaktion og forekommer inden for et millisekund efter ankomsten af et aktionspotentiale 1,4,5. Fusionen pore, den oprindelige forbindelse mellem de to fusing rum, kan flimre åbne og lukkede flere gange, før genlukning eller udvide irreversibelt 5-7. De tidligere resultateri forbigående, "kys & run" fusion, mens sidstnævnte fører til fuld fusion. Faktorer for balancen mellem disse to former for fusion og mekanismer, der regulerer pore flimmer er ikke godt forstået 5,8.
SNARE proteiner er nødvendige for exocytose; synaptisk vesikel fusion afskaffes ved spaltning af snarer ved nervegifte 9. Bulk fusionseksperimenter med små unilamelblærer (SUV'er) viste, at Snares ikke kræves kun, men også tilstrækkelig til at drive membran fusion 10. I denne bulk-assay SUV'er rekonstitueret med v-Snares (v-SUV) blev doteret med fluorescerende phospholipider (N - (7-nitro-2-1,3-benzoxadiazo-4-yl) -phosphoethanolamine (NBD-PE) og ( N -. (lissamin rhodamin B sulfonyl) -phosphoethanolamine (LR-PE) og blandes med umærkede vesikler indeholdende t-snarer (t-SUV) Oprindelig fluorescensen af NBD-PE i V-SUV'er standses ved Forster-resonansenergioverførsel (FRET ) til LR-PE. Som labELED v-SUV'er fuse med umærkede t-SUV'er, er fluoroforen overflade tætheden i den nu kombineret membran reduceret, og den deraf følgende stigning i NBD-PE fluorescens rapporterer omfanget af lipid blanding 10. Da hovedparten assay er nem at sætte op og analysere, har det været almindeligt anvendt til at undersøge mekanismerne for SNARE-medieret fusion 10-14. Det har imidlertid adskillige begrænsninger, såsom lav følsomhed og dårlig tidsopløsning. Vigtigst er det, som et ensemble måling, det gennemsnit resultaterne for alle hændelser gør forskelsbehandling mellem docking og fusion, samt påvisning af hemifusion mellemprodukter vanskelig.
Gennem det seneste årti flere grupper, herunder vores, har udviklet nye analyser for at overvåge fusion begivenheder på enkelt vesikel niveauet 15-27. Ha og kolleger brugte v-SUV'er tøjret på en overflade og overvåget deres fusion med gratis t-SUV'er 18,19. Lipidblanding blev overvåget ved anvendelse FRET mellem et par lipid-bundne fluoroforer emsengs i v- og t-SUV'er henholdsvis hjælp total intern refleksion fluorescens (TIRF) mikroskopi 18. Senere Brunger laboratorium brugt en enkelt lipid-label arter sammen med en indhold markør til samtidig påvisning af lipid og indhold blande 20,28. Både lipid og indholdet markører blev medtaget ved høje, selv-quenching koncentrationer; fusion med umærkede SUV'er resulterede i fluorescens-dequenching 20,28.
Andre har smeltet v-SUV'er til plane dobbeltlag rekonstitueret med t-Snares 15-17,21-27,29. Den plane geometri af målet (t-SNARE holdige) dobbeltlaget bedre efterligner den fysiologiske fusionsprocessen af små, meget krumme vesikler med en flad plasmamembranen. Den Steinem beskæftigede koncernen pore-spænder membraner rekonstitueret med t-Snares, suspenderet over en porøs silicium nitrid substrat og opdaget fusion med individuelle v-SUVs bruger konfokal laser scanning mikroskopi 23. Andre fbrugte v-SUV'er til plane dobbeltlag rekonstitueret med t-Snares, støttet på et glas substrat 15-17,21,22,24-27,29. Den store fordel ved at anvende understøttede dobbeltlag (SBLs) er den TIRF mikroskopi kan anvendes til at detektere docking og fusionsbegivenheder med fremragende signal-til-støj-forhold og uden interferens fra fri V-SUV'er, selv om anvendelse af mikrofluidik også tilvejebringer single-event opløsning ved brug standard fjernfelts epifluorescens mikroskopi 24.
En stor bekymring er, om og hvordan substrat-dobbeltlag-interaktioner påvirker understøttet dobbeltlag kvalitet og fusionsprocessen. Tidlig arbejde gjort brug af plane SBLs, der blev støttet direkte på et glas eller kvarts substrat 15-17. Disse SBLs blev foretaget ved adsorption, fyldt, spredning og fusion af t-SUV membraner på substratet. Det blev snart indså dog, at udelade en vigtig t-SNARE komponent, SNAP25, fra SBLs fremstillet på denne måde resulterede i v-SUV docking og fusion kinetik indistinguisHable fra dem, der opnås ved hjælp af den fuldstændige t-snarer 17. Fordi SNAP25 er absolut nødvendig for fusion in vivo 30,31, blev den fysiologiske relevans af disse tidlige forsøg sat spørgsmålstegn. Tamm gruppe overvandt denne udfordring ved at bruge bedre kontrolleret understøttet dobbeltlag dannelse 21. Det bruges Langmuir-Blodgett deposition for proteinfrit første blad af SBL, efterfulgt af fusion af denne monolag med t-SUV'er 21. Dette resulterede i SNAP25-afhængig fusion.
At undgå potentielle artefakter forbundet med et dobbeltlag direkte understøttet på et glassubstrat uden behov for anvendelse af Langmuir-Blodgett metoder, Karatekin et al. Introducerede en blød, hydratiseret poly (ethylenglycol) (PEG) stødpude mellem dobbeltlaget og substratet 24. Denne ændring resulterede også i SNAP25-afhængig fusion 24. Dobbeltlag polstret på en blød polymerlag havde været kendt til bedre bevare transmembraneprotein mobilitet og funktion 32, og var blevet brugt i fusion studier med virus 33. Desuden PEGylerede dobbeltlag synes at bevare en vis evne til selv at helbrede og er meget robust 34,35. Først en brøkdel af kommercielt tilgængelige, lipid-bundne PEG-kæder med i t-SUV membran. Når disse t-SUV'er briste og danne et plant dobbeltlag på et glassubstrat, en PEG børste dækker både foldere af den plane dobbeltlaget. Fordi plane dobbeltlagsformation drives af adhæsion af PEG-kæderne omgivende t-SUV'er på hydrofile glasoverflade, liposom sprængning og plane dobbeltlagsformation er relativt ufølsomme for lipidsammensætningen anvendt. Men når store mængder af kolesterol er inkluderet, hvilket øger de kohæsive egenskaber af SUV'er, SUV'er kan ikke briste spontant. Hvis dette er tilfældet, kan osmotisk chok eller divalente ioner at medvirke til at plane dobbeltlagsformation 25.
Som nævnt ovenfor i denne approach en PEG børste dækker begge sider af plane, støttet dobbeltlaget. Børsten overfor mikrofluid strømningskanal med til at forhindre ikke-specifik adhæsion af indkommende v-SUV'er, som også normalt dækket med en PEG lag. Dannelse af V- og t-SNARE komplekser starter fra membranen-distale N-terminaler og fortsætter i etaper mod den membran-proksimale domæner 36. For v-SUV'er til at interagere med T-SBL, den v- og t-SNARE N-terminaler behov for at rage op over PEG børster, hvilket synes at være tilfældet under betingelserne for assayet. Børste højde kan tilpasses til at studere andre end snarer proteiner ved at variere tætheden af PEGylerede lipider og PEG kædelængde 37,38. En anden fordel ved PEG børster dækker de proksimale overflader af fusing dobbeltlag er, at de efterligner de overfyldte miljø i biologiske membraner, som er pakket med 30.000-40.000 integrale membranproteiner per kvadrat mikron 39. Ligesom PEG-kæderne i dette assay repulsive protein lag, der dækker biologiske membraner skal skubbes til side for at give mulighed for kontakt mellem de to phospholipiddobbeltlag til fusion at forekomme.
Mikrofluid strømningskanaler anvendes i dette assay, da de tilbyder unikke fordele. Først mikrofluid flow muliggør mere ensartet afsætning af T-SUV'er til at sprede og sikringen til dannelse af t-SBL. For det andet, den lille kanal volumen (<1 ul) minimerer forbruget prøve. Tredje de små rumfang, der kræves tillade hele forsøget kan udføres under konstant flow. Flow fjerner svagt, formentlig ikke-specifikt, klæbes v-SUV'er fra SBL 16. Det opretholder også en konstant tæthed af v-SUV'er over t-SBL, forenkling kinetisk analyse 17. Endelig er forankret vesikler let skelnes fra frie dem båret af strømmen 25. Fjerde, flere mikrofluidkanaler kan bruges på samme dækglas, hvert sondering en anden tilstand. Dette tillader sammenligning af vilkår during samme eksperimentelle løb. En lignende fremgangsmåde er blevet anvendt af van Oijen gruppe til at studere fusion mellem influenzavirus og polstrede SBLs 33.
I TIRF mikroskopi, den eksponentielle henfald af flygtige felt (med en faldekonstant ~ 100 nm) begrænser fluorescens excitation til de molekyler, der er i meget tæt nærhed af glasset-buffer interface. Dette minimerer bidrag af fluorescerende molekyler, som er længere væk, øger signal-til-støj-forhold, og tillader enkelt molekyle følsomhed med eksponering ramme tider med 10-40 msek. Den kortvarigt felt fører også til et signal stigning på fusion: som den mærkede overførsel lipider fra SUV til SBL, de befinder sig i gennemsnit i en stærkere excitation felt. Denne stigning i fluorescens er stærkere for større liposomer.
Hvis der anvendes polariseret lys til at generere den flygtige felt, yderligere effekter bidrage til ændringer i fluorescens ved T-ransfer etiketter fra SUV til SBL. Nogle lipid farvestoffer har en overgang dipol orienteret med en foretrukken gennemsnitlig vinkel i forhold til dobbeltlaget, hvori de er indlejret. Dette skaber en forskel i mængden af fluorescens, der udsendes af de fluoroforer, når de er i SUV versus SBL, eftersom det polariserede stråle vil excitere farvestoffer i de to membraner forskelligt. For førstnævnte vil excitation stråle interagere med overgangseffekter dipoler orienteret omkring den sfæriske SUV, mens der for sidstnævnte, vil dipol orienteringer begrænses af den flade SBL geometri. For eksempel, når s-polariseret indfaldende lys (polariseret vinkelret på planet af forekomst) anvendes, magnetisering er mere effektiv, når farvestoffet er i SBL end i SUV til et lipid farvestof overgang dipol orienteret parallelt med membranen 29,40 (som den i Dii eller har 41-43). En SUV doteret med sådan en fluorofor vises dim når den kører onto SBL (figur 7, Representat ive Resultater). Som en fusion pore åbner og forbinder SUV og SBL membraner, fluorescerende prober diffundere ind i SBL og bliver mere tilbøjelige til at blive exciteret af s-polariseret kortvarigt felt 25,27,29. Følgelig fluorescenssignalet integreret omkring fusionsstedet øges kraftigt under overførsel farvestof fra SUV ind SBL 27 (figur 3 og figur 7). En yderligere faktor, der bidrager til at signalere ændringer, som ledsager fusion dequenching af fluorescerende mærker, som de er fortyndet, når de overføres til SBL. Bidraget fra dequenching er sædvanligvis mindre sammenlignet med flygtige felt henfald og polarisationseffekter i assayet beskrevet her, fordi kun en lille brøkdel (  ) Af lipiderne er mærket.
) Af lipiderne er mærket.
Signalet stigning ved fusion kan udnyttes til at udlede fusion poreegenskaberne ved at sammenligne den tid,1 "src =" / files / ftp_upload / 54349 / 54349eq2.jpg "/>, der kræves for et lipid at undslippe gennem en pore, der er frit gennemtrængelig for lipider til den aktuelle udgivelse tid,  . Hvis de to tidsskalaer er sammenlignelige, ville det konkluderes, at pore præsenterer lidt modstand til lipid flow. Men hvis den faktiske frigivelse tid er betydeligt længere end den tid, for diffusion begrænset udgivelse, ville dette betyde en proces, såsom pore flimrende, forsinker lipid frigivelse. Udbredelsen-begrænset release tid,
. Hvis de to tidsskalaer er sammenlignelige, ville det konkluderes, at pore præsenterer lidt modstand til lipid flow. Men hvis den faktiske frigivelse tid er betydeligt længere end den tid, for diffusion begrænset udgivelse, ville dette betyde en proces, såsom pore flimrende, forsinker lipid frigivelse. Udbredelsen-begrænset release tid,  , Afhænger af størrelsen af fusing liposom og lipid-diffusivitet; sin vurdering kræver disse to parametre, der skal kvantificeres. Den enkelt molekyle sensitivitet tillader lipid diffusivitet skal måles ved at spore flere enkelt lipid fluoroforer efter deres løsladelse i SBL for hver fusion begivenhed 26. Størrelsen af hver fusing vesikelkan estimeres 27 ved at kombinere (i) intensiteten af en enkelt lipid farvestof, (ii) ændringen i den totale fluorescens omkring en docking websted efter alle fluoroforer overføres til SBL ved fusion, (iii) den kendte mærkning tæthed af SUV lipider, og (iv) det areal pr lipid. For mange fusionshændelser blev de faktiske lipid release gange fundet at være meget langsommere end forventet ved diffusion-controlled release 27, som blev bemærket tidligere antagelse ensartet SUV størrelse 44. Antages retardering af lipid frigivelse skyldes pore flimrende, en kvantitativ model tillader estimering af "pore åbenhed", den del af tiden pore forbliver åben under fusion 27.
, Afhænger af størrelsen af fusing liposom og lipid-diffusivitet; sin vurdering kræver disse to parametre, der skal kvantificeres. Den enkelt molekyle sensitivitet tillader lipid diffusivitet skal måles ved at spore flere enkelt lipid fluoroforer efter deres løsladelse i SBL for hver fusion begivenhed 26. Størrelsen af hver fusing vesikelkan estimeres 27 ved at kombinere (i) intensiteten af en enkelt lipid farvestof, (ii) ændringen i den totale fluorescens omkring en docking websted efter alle fluoroforer overføres til SBL ved fusion, (iii) den kendte mærkning tæthed af SUV lipider, og (iv) det areal pr lipid. For mange fusionshændelser blev de faktiske lipid release gange fundet at være meget langsommere end forventet ved diffusion-controlled release 27, som blev bemærket tidligere antagelse ensartet SUV størrelse 44. Antages retardering af lipid frigivelse skyldes pore flimrende, en kvantitativ model tillader estimering af "pore åbenhed", den del af tiden pore forbliver åben under fusion 27.
Når praktisk, er det vigtigt at teste fusion mekanismer ved hjælp af både lipid og opløselige indhold etiketter. For eksempel kunne lipid frigivelse forsinkes af andre end pore flimrende processer, såsom begrænsning af lipid diffusion af SNARE proteiner, surround than pore. Hvis dette var tilfældet, så slip af indhold vil gå forud frigivelse af lipid etiketter, forudsat pore er stor nok til at tillade passage af opløselige sonder. En mere grundlæggende fejl i tilgangen kunne være i den antagelse, at overførslen af mærkede lipider til SBL sker gennem en smal fusion pore forbinder SBL til en vesikel, der har stort set bevaret sin pre-fusion form. Lipid overførsel til SBL kan også skyldes en hurtig udvidelse af den fusion pore med en samtidig, ekstremt hurtige sammenbrud af SUV i SBL membran, som tidligere foreslået baseret på lipid release data alene 29. Overvågning både lipid og indholdet frigiver samtidigt, blev det konstateret, at mange porer lukkes igen efter udløsning alle deres lipid etiketter, men bevaret nogle af deres opløselige last 27. Dette indikerer, at i det mindste har nogle liposomer ikke skjule i SBL efter fusion, og at lipid farveoverføringsinhiberende ind SBL sker gennem en fusion pore. Derudover lipid og indhold udgivelse forekom samtidigt 27, hvilket gør det usandsynligt, at forsinkelse af lipid frigivelse skyldtes hindring for, lipid diffusion af SNARE proteiner omgiver pore 45.
En SUV-SBL fusion protokol, der ikke overvåger opløselige indhold udgivelse er tidligere blevet udgivet af Karatekin og Rothman 25. Her er mere den seneste udvikling inkluderet, nemlig samtidig overvågning af lipid og indhold frigivelse og estimering af SUV, lipid, og fusion pore egenskaber 27. Protokollen begynder med instruktioner til at forberede mikrofluide celler, lavet af limning en poly (dimethylsiloxan) (PDMS) elastomer blok indeholder riller med et dækglas 25. Dernæst udarbejdelse af v-SUV'er med både lipid og indhold markører forklaret. Afdeling 4 og 5 give anvisninger til at samle de mikrofluide celler, der danner SBLs in situ og kontrol for fejl og flydende, indførelse af vSUVS ind flowcellerne og detektion af fusionsbegivenheder. Afsnit 6 indeholder instruktioner til dataanalyse.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Udarbejdelse af en PDMS blok at danne Mikrofluid Channel

Figur 1. mikrofabrikation af flowcelle skabelon og PDMS blok forberedelse. (A) Design af en fire-kanal flowcelle, der passer på en 24 x 60 mm dækglas (nederst). Seks identiske designs er indrettet til at passe på en 10 cm siliciumskive (øverst). (B) Skær blok på ca. 5-8 mm tykke PDMS på et hul puncher. (C) Indsættelse af slangen i den udstansede hul med en pincet. Klik her for at se en større version af dette tal.
- Opnå en flow-celle skabelon som den i figur 1A. Typiske kanaler er 0,3-1 mm brede, 70-100 um høj, og 1-2 cm lange. Fabrikere en skabelon osing standard fotolitografiske teknikker i et renrum facilitet 25 eller ordre, hvis renrum adgang er ikke tilgængelig. Alternativt maskine en skabelon med større dimensioner fra et egnet materiale.
Bemærk: renrummet personale kan træne og vejlede uerfarne brugere (adgang til et renrum anlægget er altid begrænset til brugere med en ordentlig uddannelse) i design og bestilling af en maske, wafer rengøring, og fotolitografi. Når en skabelon er opnået, kan det bruges uden renrummet gentagne gange, forudsat det holdes i et lukket fad og omhu for at udelukke støv. - Klargør en blanding af ~ 100 ml PDMS (silikone elastomer base) og ~ 10 ml cross-linker (hærder) i en engangs plastik kop. Bræk spidsen af en engangs pipette til at håndtere de viskøse PDMS lettere. Afveje PDMS i plastik kop som pipettering er ikke korrekt.
- Omrør blandingen godt. Fjern de mange luftbobler ved afgasning i vakuum (ca. 20min). Som bægeret anbragt under vakuum, vil boblerne oprindeligt vokse i størrelse, øge mængden af blandingen betydeligt. Styr vakuum, der finder anvendelse, og sørg koppen er dyb nok til at undgå et udslip.
- Hæld en stor dråbe den afgassede PDMS blandingen i et glas petriskål (150 mm x 20 mm), og tryk skiven onto PDMS med templaten opad. Derved undgås bobler bliver fanget under skiven. Trapped bobler vil ekspandere og kan vippe skiven når skålen anbringes i ovnen. Nu hældes afgassede PDMS onto skabelonen oven på skiven, indtil det er omfattet af ca. 5-8 mm af PMDS. Hvis en luftboble sker forsigtigt fjerne det med en pipettespids.
- Bag PMDS i en ovn ved 60 ° C i 3 timer. Sørg for, at fadet er niveauet.
- Brug en ny skalpel til at skære en PDMS blok indeholder de støbte kanal strukturer. Den skåret ud blok skal passe på en dækglas (se trin 4.1.9).
- Skræl den skåret ud blok fra fadet og place det på et stykke rent aluminiumsfolie.
Bemærk: De tværbundne PMDS blokke kan opbevares i et par måneder. På en enkelt wafer, der er 6 sæt strømningskanaler (figur 1A), så 6 PDMS blokke kan fremstilles i en enkelt PDMS støbning. Efter at skære ud alle 6 af de PDMS blokke, rene løse stykker af PDMS, men ellers ikke fjerne de resterende PDMS før hælde og krydsbinding den næste batch, som vil kræve kun omkring halvdelen af de PDMS anvendes til det første parti. En god skabelon kan vare et par år. - Brug et hul Puncher at bore gennem PDMS blok i en lige bevægelse (figur 1B). Start på den side af kanalen riller. Sørg for at fjerne det udstansede stykke PDMS. Gentag dette for alle otte huller af fire kanals design.
- Placer PDMS blok kanalsideinformation ned på en ny og rynkefri stykke aluminiumsfolie. Opbevar blokken i op til et par måneder i en tør kasse, ideelt i en ekssikkator.
- Skub slangen (0,25 mm ID, 0,76 mm OD) omkring en tredjedel i det udstansede hul med en pincet (figur 1C). Skær slangen skrå lettere indføring. Efterlad rør lang nok til at nå SUV reservoiret og sprøjtepumpen hhv. Efter placering af samlet chip på mikroskopet, skære rørene igen, hvis de er for lange.
- For at forbinde til sprøjterne pumpens, skære et kort stykke større silikoneslanger (0,51 mm id, 2,1 mm OD) og sæt tyndere slange i den ene side. Denne klar-til-brug blok af PDMS kan holdes i et par måneder.
2. Coverslip Rengøring
- Rene dækglas ifølge protokollen beskrevet i Karatekin og Rothman 25 under anvendelse af en stærk oxiderende blanding af svovlsyre (H 2 SO 4) og hydrogenperoxid (H 2 O 2). Udfør denne procedure i en renrum, og observere ordentlige sikkerhedsforanstaltninger. Alternativt kan du bruge et renrum rengjort coverslip der er kommercielt tilgængelige (se Materialer List).
3. Udarbejdelse af v-SUV'er indeholdende både lipid og indhold etiketter

Figur 2. Skematisk af SUV præparat. Lipider blandes i et glasrør (1), og opløsningsmidlet afdampes til dannelse af en lipidfilm ved at dreje røret i et vandbad (2). Resterende spor af opløsningsmiddel fjernes under højvakuum (3). Lipidfilmen hydratiseres i rekonstitution buffer indeholdende detergent og protein mens hvirvelbehandlet (4). Hvis indholdet farvestof skal indkapsles, er det inkluderet i dette trin samt i fortyndingsluften trin (5). Fortynding af detergentkoncentrationen under dets kritiske micellekoncentration fører liposomdannelsen. Vaskemiddel dialyseres væk natten over (6). For t-SUV'er til SBL dannelse herunder NBD-PE (grøn), er vesikler flød i en densitetsgradient og indsamlet pågrænsefladen mellem to lag (7a). For at adskille v-SUV'er med indkapslet indhold markør fra fri farvestof prøven køres over en søjle størrelse-udelukkelse og opsamles i 0,5 ml fraktioner (7b). Klik her for at se en større version af dette tal.
- Forbered 4 l rekonstituering puffer, indeholdende 25 mM HEPES-KOH, 140 mM KCI, 100 uM EGTA og 1 mM DTT, pH 7,4. Bruge de fleste af bufferen til dialyse og 100 ml for andre trin.
Bemærk: Andre puffere kan anvendes, men det er vigtigt at holde bufferen osmolaritet konsekvent gennem hele eksperimentet og vælge tilstande, hvor proteinerne er funktionelle. For at forberede v-SUV'er kun indeholder lipid etiketter og t-SUV'er, følg Karatekin og Rothman 25. - Som lipid lagre opbevares ved -20 ° C i chloroform eller chloroform: methanol (2: 1, vol / vol), lad hætteglas stuetemperatur før åbning tilundgå kondens.
- Skyl et glasrør med chloroform for at fjerne spormængder af detergent og blandes lipiderne ved det ønskede forhold med en endelig mængde på 1 pmol i en blanding af chloroform og methanol (2: 1, vol / vol). Brug kun glas sprøjter / rør til at håndtere organiske opløsningsmidler / opløste lipider.
Bemærk: De anvendte lipider her er POPC: SAPE: DOPS: Cholesterol: PEG2000-DOPE: gjorde ved et molforhold på 57,4: 15: 12: 10: 4,6: 1 til v-SNARE indeholdende vesikler og POPC: SAPE: DOPS: Cholesterol : hjerne PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5) i t-SNARE SUV'er. Se Materiale for detaljer. Andre lipidsammensætninger kan anvendes. - Opløsningsmidlet afdampes under en svag strøm af nitrogen, eller i en rotationsfordamper. Fordybe spidsen af røret i et vandbad (37 ° C) opvarmes over den højeste smeltetemperatur af lipiderne til opnåelse af en homogen lipidfilm og for at undgå store ændringer i temperatur, mens afdampning af opløsningsmidlet. Start vakuum ved omkring300 mbar indtil lipidfilmen er dannet, derefter fortsætte til ~ 2 minutter ved højeste mulige vakuum i rotationsfordamper.
- Fjern glasrøret og pak det i alufolie for at undgå udsættelse for lys og fjerne de resterende spor af opløsningsmiddel ved at placere røret i en ekssikkator under højt vakuum i mindst 2 timer. Dette trin kan også køre natten over.
- At rehydrere den tørrede lipidfilm fremstilles en blanding af detergent (n-octyl-β-D-glucopyranosid, OG), protein (v-snarer), og SRB i rekonstituering puffer (500 pi slutvolumen). Hold den endelige detergentkoncentration ~ 2 gange over den kritiske micellekoncentration (CMC, 20-25 mM for OG) og det endelige rumfang justeres sådan, at detergenten og lipid forholdet er> 10. Sulforhodamin B (SRB) Pulveret opløses i den proteinholdige detergentopløsning til opnåelse af en slutkoncentration på 10-50 mM SRB ved forsigtigt at ryste opløsningen. Juster osmolaritet rekonstitutionsbuffer når der tilføjes høje koncentrationer af SRB ved at reducere concentration af kaliumchlorid i overensstemmelse hermed.
Bemærk: Lipid-til-protein-forhold (LP) for t-snarer er høj (LP ~ 20.000, ~ 70 t-snarer per sq mikron. 25). Markant højere t-SNARE tætheder i SBL kan føre til inaktive proteinaggregater med væsentligt reduceret de fusion satser 17,24. LP for v-snarer er meget lavere (LP ~ 200, ~ 7.000 v-snarer pr um 2), tæt på de v-SNARE densiteter findes på synaptiske vesikler 46,47. At sikre proteinfunktion efter rekombinant ekspression og oprensning af v- og t-snarer er det nyttigt at udføre en simpel bulk-fusion assay 48 før de går gennem alle trin af den indre vesikelfusion assay. - Ryst forsigtigt forvarmet glasrør indeholdende den tørrede lipidfilm under tilsætning af protein-detergent-SRB-opløsning fra trin 3.6. Prøv at undgå at skabe bobler. Fortsæt med at ryste i 15 minutter ved 37 ° C.
- Fortynd detergent femdobbelte ved tilsætning rekonstitutionbuffer indeholdende SRB (tilsættes 2 ml, 2,5 ml endelig volumen), mens hurtigt vortex at undgå koncentrationsgradienter. Fortsæt med at ryste i 15 minutter til 1 time ved 37 ° C.
Bemærk: Længere inkubation stigning protein rekonstituering effektivitet. Den hurtige fortynding formindsker detergentkoncentrationen under CMC og fører til dannelse af små liposomer. - Dialysere det vehikelsuspension først mod ~ 1 I rekonstitution puffer i 1-2 timer ved stuetemperatur og derefter mod 3 liter rekonstitutionsbuffer natten over ved 4 ° C med 4 g polystyren adsorbent, ved anvendelse af en 20.000 MWCO dialyserør eller kassette. Brug forskellige bægre til dialyse af vesikler med og uden SRB at undgå krydskontaminering.
- Ækvilibrer en gelfiltreringssøjle med rekonstitution puffer. Kør vehikelsuspension gennem søjlen for at separere fri SRB fra v-SUV'er med indkapslet SRB. Brug rekonstitutionsbuffer uden SRB som eluent når hele prøven harindtastet kolonnen. Saml v-SUV'er i 0,5 ml fraktioner.
- Bekræft vellykket SRB indkapsling ved selvstændige slukket koncentrationer ved at måle SRB fluorescens før og efter tilsætning af vaskemiddel til en portion af vesikler 16. Ved hjælp af en fluorescens spektrometer, ophidse prøven ved 550 nm og scanne SRB emission mellem 570 nm og 630 nm. Forvente en 4-8 gange stigning i SRB fluorescens afhængigt af den indkapslede mængde på detergent tilsætning som membranen er solubiliseret og det frigivne SRB fortyndes.
- Karakterisere lipid og protein recovery anvendelse af fluorescens spektroskopi 24 og SDS-PAGE-gelelektroforese, hhv. Brug en følsom farvningsmetode, især for de høje LP t-SUV prøver (se Materialer List). Typisk omkring 50% af både lipid og protein input er tabt under udarbejdelsen hvilket resulterer i en LP tæt på den nominelle værdi.
- Karakterisere SUV størrelser ved hjælp af dynamisk lysspredning 24 eller elektron microscopy 48. Store SRB indeholdende V-SUV'er ved 4 ° C op til ~ 3-4 dage. Må ikke fryses, da nedfrysning og optøning bryder membranen og frigiver indkapslet SRB.
4. SUV-SBL Fusion analysen til overvågning Lipid Slip Kun
- Dannelse af den bundne understøttede dobbeltlaget inden for de mikrofluide strømningskanaler
- Placer PDMS blok (trin 1.11) under højvakuum i mindst 20 min før forsøget for at fjerne opløste gasser. Dette reducerer risikoen for luftbobler i mikrofluidkanaler under fusionsprocessen eksperimentet.
- Tænd mikroskopet opsætning og opvarme den fase og prøveholderen (figur 3 og 4) til den ønskede temperatur.
- Foretage rekonstitutionsbuffer anvendes til fortynding vesiklerne gennem et filter med 0,45 um eller mindre porestørrelse.
- Fortynd ~ 30 pi af NBD-PE-mærket t-SUV'er eller protein-fri (PF-SUV'er) kontrol liposomer (lageropløsning 0,5-1 mM lipid) med ~ 60 pi puffer. Slutkoncentrationen er ikke kritisk her.
- Afgasse denne blanding under anvendelse af en 3 ml sprøjte. Tryk ud det meste af luften over prøven, mens du holder sprøjten lodret. Forsegl (nål gratis) tip ved hjælp paraffin film og skabe et vakuum ved at trække stemplet ned. Tap sprøjtecylinderen at fremskynde afgasning af opløsningen. Gentag denne proces et par gange, indtil der ikke opstår flere bobler når vakuum påføres.
- Brug en injektionsnål med en diameter lidt større end slangen, der er fastgjort til PDMS blok at punch et hul i hatten på et mikrocentrifugerør. Sørg for, at udstansede stykke plastik er ikke inde i røret, da det kan tilstoppe slangen forbundet til mikrofluid kanal indløb senere.
- Fyld afgasset SUV opløsningen i mikrocentrifugerør har et hul i hætten og placere den i holderen på mikroskopet scenen i ligevægt til den indstillede temperatur.
- place en tidligere renset dækglas (afsnit 2) i et plasma renere og køre luft plasma til omkring 5 min. Sætte plasmaet behandlede dækglas (behandlet opad) oven på et par fnugfri væv tjener som en pude.
- Fjern aluminiumfolie fra den afgassede PDMS blok og placere blokken på toppen af dækglasset. Når genbrug PDMS blok sætte på og afmontere et stykke tape til kanalen side at rense den. Tryk ned PDMS blok på dækglasset ved hjælp af en pincet til at gøre det stick, men ikke trykke for hårdt, da glasset kan bryde.
- Placer den samlede flow celle på mikroskopet scenen og tilslut slangen til SUV reservoiret og sprøjten pumpe, hhv. Tape dækglasset på scenen (figur 3B).
- Start udsugning af SUV'er på 3 pl / min, indtil opløsningen fylder kanalerne fuldstændigt (~ 2,25 mm / sek for 75 um x 300 um kanaltværsnit). Når løsninger til alle kanaler begynder at flytte op the slange på udgangssiden, reducere strømning til 0,5 pl / min og inkuberes i 30-45 min.
Bemærk: Den tid mellem plasma behandle objektglas og strømmer SUV'er ind i kanalen bør ikke overstige 10-20 min som effekten af plasmaet behandling er forbigående. - Tjek kanalerne til eventuelle udslip. Brug en 10-20x luft målsætning at observere NBD-PE fluorescens fra NBD-PE indeholdende T- eller pf-SUV'er. Excite NBD fluorophorer hjælp af 488 nm laser. Utætheder er vanskelige at opdage ved hjælp af lys belysning felt.
- Sæt et rør med afgasset rekonstituering buffer ind i holderen og lad temperaturen ligevægt så hurtige ændringer i temperatur kan forårsage fejl i SBL. Stop flowet og vente ~ 1 min for at sikre strømmen er standset helt, og luftbobler vil blive suget ind i røret, før du skifter indløbs- rør til den afgassede buffer.
- Skyl alle kanaler med afgasset buffer til at vaske væk ubundne SUV'er.
- Skift til en higher forstørrelse TIRF mål (60X, olie, NA 1,45-1,49), og kontrollere, at dobbeltlaget ser homogene og er uden nogen tydelige store defekter såsom mørke pletter eller lipid tubuli strækker sig ud fra SBL.
- Tjek fluiditet dobbeltlaget
- Hvis en dedikeret FRAP enhed ikke er tilgængelig, eller hvis et FRAP sekvens ikke kan programmeres, så kvalitativt test membran fluiditet som følger.
- Luk feltblændet til en lille størrelse (~ 40 um diameter) og juster 488 nm excitation lysintensitet via softwaren (20-80 pW, eller 15-60 nW / um 2) at blege NBD-PE-fluorescens i den udsatte område betydeligt, men ikke fuldstændigt. For en fluid dobbeltlag, ved steady state, bør fluorescensintensiteten i midten af det udsatte område være lavere end ved kanterne, som intakte NBD-PE-molekyler ind i det eksponerede område og diffundere en vis afstand før blegning. Derimod hvis overfladen-klæbet SUVs undlod at briste eller anden grund det understøttede dobbeltlag er ikke flydende, bør alle fluoroforer i det udsatte område blege.
Bemærk: Laser intensitetsværdier i denne og senere trin er givet som et groft udgangspunkt, og bør optimeres for et givet sæt betingelser. - Stop belysning og starte det igen et par minutter senere for at bekræfte resultaterne af steady state målinger
- Luk feltblændet til en lille størrelse (~ 40 um diameter) og juster 488 nm excitation lysintensitet via softwaren (20-80 pW, eller 15-60 nW / um 2) at blege NBD-PE-fluorescens i den udsatte område betydeligt, men ikke fuldstændigt. For en fluid dobbeltlag, ved steady state, bør fluorescensintensiteten i midten af det udsatte område være lavere end ved kanterne, som intakte NBD-PE-molekyler ind i det eksponerede område og diffundere en vis afstand før blegning. Derimod hvis overfladen-klæbet SUVs undlod at briste eller anden grund det understøttede dobbeltlag er ikke flydende, bør alle fluoroforer i det udsatte område blege.
- Programmere en FRAP sekvens for en mere kvantitativ måling, hvis det er muligt. Se supplerende filer og den tilsvarende legende for detaljer.
Bemærk: Nogle gange SUV'er vil klæbe på dækglas, men undlader at briste og danne en fluid dobbeltlaget. Hvis dette sker, skylles kanalerne med afgasset rekonstitution indeholdende 10 mM Mg2 + for at hjælpe understøttet dobbeltlagsformation. Brug fluorescens blegning at vurdere dobbeltlag flydende som i 4.2. Når et fluid dobbeltlag er dannet, skyl med Mg2 + -fri Reconstitution puffer.
- Hvis en dedikeret FRAP enhed ikke er tilgængelig, eller hvis et FRAP sekvens ikke kan programmeres, så kvalitativt test membran fluiditet som følger.
- Introduktion v-SUV'er i mikrofluide strømningskanaler
- Degas rekonstitutionsbuffer og bruge den til at fortynde v-SUV stamopløsning med en faktor på ca. 10 3 til 10 5 afhængig af v-SUV stamkoncentration. Målet for en fortynding, der resulterer i omkring 10-100 fusionsbegivenheder inden 60 sekunder i synsfeltet.
Bemærk: Alt for mange fusioner øger baggrund fluorescens (da hver event indskud LR-PE eller gjorde lipid etiketter i SBL) og gøre detektion og analyse af fusion begivenheder vanskelig. I modsætning hertil en fusion sats, der er for lave resultater i dårlig statistik eller kræver erhvervelse af mange flere film. For en v-SUV koncentration på 0,1 mM lipid, starte ved at fortynde 5 pi SUV lager i 995 pi rekonstituering buffer og derefter fortynde 5-50 pi af dette i 950-995 pi rekonstituering buffer. - Lad temperatur ligevægt før standse strømmen og inserTing indløbsrøret ind i fortyndede v-SUV-opløsning.
- Juster TIRF vinkel og polarisering som følger.
- Efter indstilling af den ønskede polarisation ved at dreje excitationsstrålen, justere hældningen af spejlet kommanderende positionen af strålen via en stepmotor gennem softwaren. Flyt langsomt position af laserstrålen fra centrum af målet på bagsiden brændplanet til den ene side excentrisk. Observere lyset af formålet ved forsiden dukke med en stigende vinkel i forhold til målet akse som position bevæges yderligere excentrisk.
- Bemærk motorens position, når den udstrømmende stråle først forsvinder ind i målet, dvs. når TIR først opnås.
- holde langsomt bevæger strålen position yderligere off-center under overvågning af fluorescensen fra overfladen. Bemærk motorens position, når overfladen fluorescens forsvinder, når strålen er flyttet for langt off-center.
- Vælg en stråle positio n mellem de to grænser fastlagt ovenfor. For bedste signal-støj-forhold og mere signal enhancement på fusion, vælge en lavvandet indtrængningsdybde (stråle position tættere på kanten af målet), som stadig giver en ensartet belysning af Viewfield.
Bemærk: Det er bedst at holde den samme TIR stråle position (samme indtrængningsdybde) for alle eksperimenter efter indstillinger optimeres. Sørg dreje polarisering ikke resulterer i ændringer i strålen position.
- Flow v-SUV'er i kanalen ved en strømningshastighed på 2 gl / min, svarende til en gennemsnitlig lineær strømningshastighed på ~ 1,5 mm / sek til et tværsnit på 75 um x 300 um. Skift til indstillingerne for excitation / emission til overvågning af lipid blanding (LR eller har kun).
- Degas rekonstitutionsbuffer og bruge den til at fortynde v-SUV stamopløsning med en faktor på ca. 10 3 til 10 5 afhængig af v-SUV stamkoncentration. Målet for en fortynding, der resulterer i omkring 10-100 fusionsbegivenheder inden 60 sekunder i synsfeltet.
- Observation fusion mellem enkelt v-SUV'er og SBL
.jpg "/>
Figur 3. Den eksperimentelle pTIRF opsætning. (A) Skematisk repræsentation af en v-SUV og en t-SBL på et glassubstrat. Falske farver TIRFM billeder af enkelt SUV-SBL fusion begivenhed viser lipid docking (1) og frigivelse af lipid farvestof i SBL (2) efterfulgt af blegning og fald i fluorescensintensiteten (3). Den totale fluorescensintensitet (summen af pixelværdier i 5,3 um x 5,3 um box) signal vises. (B) Den dækglas bundet til PDMS blok er tapede på den opvarmede fase. Indløbsrør for mikrofluidkanaler udtage prøver af rør i metal prøveholder (højre) opsuget af sprøjtepumpen (venstre). Under pumpen er den dobbelte emission enhed. Klik her for at se en større version af dette tal.
OAD / 54349 / 54349fig4.jpg "/>
Figur 4. Skematisk af forsøgsopstillingen. Den kortvarige bølge er skabt på glas-buffer-interface i mikrofluid kanal. SBL er dannet på glasset og v-SUV opsuges fra indehaveren af metal prøve (øverst til højre) gennem kanalen ind i sprøjten (øverst til venstre). M, spejl; DM, dikroisk spejl; L, linse; F, filter; P, polarisator. Klik her for at se en større version af dette tal.
- Kontinuerligt excite og overvåge v-SUV fluorescens under anvendelse af en 561 nm (for LR-PE) eller 638 nm (for gjorde) laser afhængigt af fluoroforen inkorporeret i V-SUV'er. Som v-SUV'er nå strømningskanalen og dock på og sikring med SBL, baggrunden fluorescenssignalet begynder at akkumulere.
- Juster excitation laser intensitet via softwaren til løbende blege baggrundenfluorescens, således at i steady state nye docking og fusion begivenheder let kan observeres.
Bemærk: Hvis blegning er for langsom, vil baggrundsfluorescens være for høj. Hvis blegning er for hurtig, vil signalerne fra forankret SUV'er og fra fluorophorer der frigives til SBL fade hurtigt, afkortning af vinduet i hvilken der kan detekteres fusion af forankret SUV'er eller en fluorofor kan spores. For LR-PE anslået ved 561 nm, 2,5-7,5 mW effekt belysning af en cirkel 190 um diameter (100-250 nW / um 2) er en rimelig værdi til at starte. For 638 nm excitation af gjorde, 0,8-1,6 mW magt over en cirkel 190 um diameter (30-60 nW / um 2) kan bruges til indledende tests. - Anskaf flere film på forskellige positioner i en given kanal. Check NBD-PE-fluorescens at kontrollere, at SBL ikke har defekter på disse positioner. Anskaf fuld frame film ved den maksimale hastighed (~ 50 billeder / sek) eller en beskåret område af interesse ved højere billedhastighed (op til~ 100 Hz) i 60 sek.
- Flyt til en anden mikrofluid kanal og gentag optagelserne til andre betingelser. Omfatte negative kontroller såsom et protein-frit SBL eller SUV'er, tilføje det opløselige cytoplasmatiske domæne af v-SNARE VAMP2 (CDV) som en inhibitor eller behandle v-SUV med tetanusneurotoksin (30 min ved 37 ° C) i en eller flere af kanaler på samme dækglas.
- Ryd op og PDMS genbrug
- For at genbruge den PDMS blok, skylles de mikrofluidkanaler med ~ 200 pi af 70% ethanol ved en strømningshastighed på 5 ul / min. Endelig aspireres luft gennem slangen og kanaler.
- Frigør de PDMS blokere forsigtigt fra dækglasset ved at klemme det lidt. Placer den på en ren stykke alufolie. Opbevar i vakuum. Det kan genbruges flere gange.
- For en mere grundig rengøring skylles kanalerne med et detergent eller natriumhydroxid før 70% ethanolopløsning. Alternativt, fjerne alle rør fro m PDMS og sonikeres blokken i 30 minutter i isopropanol, før tørring og indsættelse af ny slange.
5. SUV-SBL Fusion analysen til overvågning Lipid og indhold Slip Samtidigt
- For dobbelt farve overvågning af samtidige lipid og opløselige indhold frigive, følge de samme trin som i § 4, bortset fra brug liposomer mærket med både en lipid (gjorde) og en opløselige indhold (SRB) etiket, som forklaret i afsnit 3. SRB er indkapslet ved 10 mM og er oprindeligt meget selvstændig standset.
- Brug af konfiguration vist skematisk i figur 4, excite SRB og gjorde fluorescens samtidigt ved anvendelse 561 nm og 638 nm lasere hhv. En dikroisk spejl (640 nm) opdeler emissionen i to stråler, der kører gennem en kort (595/50 nm) og en lang (700/75 nm) bølgelængde filter til at detektere SRB og gjorde emissioner hhv. De to emissioner bjælker projiceres side om side på EM-CCD-chip.
- Analyse af FRAP data
- Brug MATLAB programmet leveres i de supplerende oplysninger at estimere lipid diffusionskoefficienten, D. Programmet læser en liste over OME-TIFF-filer 49, detekterer bleget område, plotter gennemsnitlige pixelværdier i bleget område som en funktion af tid, og passer den resulterende recovery kurve til en model af Soumpasis 50 at ekstrahere restitutionstid,
 , Hvor w er radius af den blegede cirkel.
, Hvor w er radius af den blegede cirkel.
Bemærk: En MATLAB program blev fastsat tidligere 25 for analyse af FRAP film ved hjælp Nikon ND2 filer. Det nuværende program læser OME-TIFF filer 49, fordi de fleste filformater let kan konverteres til OME-TIFF. En kvantitativ analyse af FRAP data nemmeste og mest præcise, når blegning er øjeblikkelig, det bleached region er en cirkel, og blegning under udlæsning er ubetydelig. Selv om disse kriterier ikke er strengt opfyldt i de enkle FRAP målinger beskrevet her, kan der opnås et rimeligt skøn over diffusionskoefficienten. For en mere præcis vurdering, bruge sporing af enkelte lipid farvestoffer (afsnit 6.3).
- Brug MATLAB programmet leveres i de supplerende oplysninger at estimere lipid diffusionskoefficienten, D. Programmet læser en liste over OME-TIFF-filer 49, detekterer bleget område, plotter gennemsnitlige pixelværdier i bleget område som en funktion af tid, og passer den resulterende recovery kurve til en model af Soumpasis 50 at ekstrahere restitutionstid,
- Analyse af docking sats, fusion sats og docking-til-fusion forsinkelsestider
- Brug ImageJ til at åbne en film til at analysere og justere lysstyrke og kontrast til klart at identificere vesikel docking og fusing begivenheder. Start SpeckleTrackerJ 26 plugin. Se dokumentationen til SpeckleTrackerJ Smith et al. 26 og online for instruktioner til brug.
- Identificer alle nyligt forankret SUV'er i SpeckleTrackerJ. For at skelne SUV'er som dock stramt fra dem, hoppe ud SBL, pålægge en docking varighed på et par frames minimum. Gem sporene, og gentag for alle film.
Bemærk: dockonge sats, der betyder noget er at identificere, når en SUV kuperet, så kun det første billede, hvor SUV'er kuperet behov, der skal registreres i disse spor. Resten af numrene vil ikke blive taget i betragtning i analysen. Imidlertid auto-tracking hver SUV indtil det bleger eller sikringer hjælper mark SUV'er, der allerede er sporet. - Identificer alle fusing vesikler. For disse bør sporene omfatter alle rammer fra den første ramme en SUV forankret indtil den første ramme, hvori fusion er tydeligt ved en pludselig stigning i fluorescensen af den sporede stedet. Varigheden af disse spor anvendes til at beregne docking-til-fusion forsinkelser. Gem alle spor. Gentag for alle film.
- For batch analyse af docking eller fusion af data, udarbejde en liste over bane filer fra de relevante film og køre MATLAB programmer, med Karatekin og Rothman 25. Følg vejledningen, der leveres sammen med programmerne.
Bemærk: Programmerne plotte cumulative docking og fusion begivenheder som en funktion af tid baseret på oplysninger fra de bane filer. Docking og fusion satser er estimeret fra skråningerne af disse grunde. For fusion data, er docking-til-fusion forsinkelser også beregnes og deres fordeling afbildet som en overlevelse plot, dvs. sandsynligheden for, at fusion endnu ikke har fundet sted ved en given forsinkelse efter docking.
- Lipid diffusivitet
- For hver fusion begivenhed, spore enkelt fluorescerende lipider som de bliver mærkbar, når de har diffunderet tilstrækkeligt væk fra fusionen (figur 6). Brug SpeckleTrackerJ til sporing, og gemme numre til yderligere analyse ved hjælp af MATLAB. Afhængig af størrelsen af fusing vesikel, typisk 3-30 enkelt fluoroforer kan spores. Fordi længere spor er ønskelige til beregning af
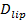 Prøv at spore enkelte molekyler som langsom muligt, ved hjælp af manuel korrektion af baner, hvis det er nødvendigt.
Prøv at spore enkelte molekyler som langsom muligt, ved hjælp af manuel korrektion af baner, hvis det er nødvendigt. - Beregn den gennemsnitlige kvadrerede forskydning (MSD) for enlige lipid markør baner, der holder> 40-50 frames (ca. ~ 1,5 sek). Brug MSD at beregne lipid diffusionskoefficienten, . Se Smith et al. 26 og Stratton et al. 27 for yderligere oplysninger.
- For hver fusion begivenhed, spore enkelt fluorescerende lipider som de bliver mærkbar, når de har diffunderet tilstrækkeligt væk fra fusionen (figur 6). Brug SpeckleTrackerJ til sporing, og gemme numre til yderligere analyse ved hjælp af MATLAB. Afhængig af størrelsen af fusing vesikel, typisk 3-30 enkelt fluoroforer kan spores. Fordi længere spor er ønskelige til beregning af
- Enkelt lipid farvestof intensitet, SUV-SBL reduktion intensitet faktor, og vesikel størrelse
- Måle summen af pixelværdier for et lipid markør i en 3 x 3 pixel (0,8 um x 0,8 um) området omkring markør for ~ 15 billeder før og efter markøren blegemidler i et enkelt trin. Fratræk baggrunden intensitet som gennemsnittet for de post-blegemiddel rammer fra den før-blegemiddel intensitet gennemsnit før blegning at opnå intensiteten af den sporede lipid markør. Gentag målingen for så mange markører som praktisk for en given film.
- Plot fordelingen af enkelt lipid label intensiteter,
 Og passe en gaussisk at estimere middelværdien.
Og passe en gaussisk at estimere middelværdien. - Beregn forsinkelse mellem når fusion fandt sted, og når den bane af en enkelt lipid markør udgivet i så fald endte i et enkelt trin blegning. Anskaf blegning tid til en fluorofor i SBL,
 , Ved at plotte den overlevende funktion af forsinkelserne og montering til en eksponentiel.
, Ved at plotte den overlevende funktion af forsinkelserne og montering til en eksponentiel.
Bemærk: Da de polariserede excitation felt par mere svagt til fluoroforer på en SUV, blegning tid en SUV er normalt meget langsommere 27. - Skøn
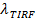 , Reduktion intensitet faktor for et lipid farvestof når i SUV forhold til når det er i SBL, efter Stratton 27 (tp_upload / 54349 / 54349eq8.jpg "/> er intensiteten af en enkelt farvestof i SUV).
, Reduktion intensitet faktor for et lipid farvestof når i SUV forhold til når det er i SBL, efter Stratton 27 (tp_upload / 54349 / 54349eq8.jpg "/> er intensiteten af en enkelt farvestof i SUV).
Bemærk: Hvis blegning var ubetydelig, ville være lig med kuperet intensitet  (punkt (1) i figur 3A, højre panel), divideret med det totale intensitet nås efter alle fluoroforer er deponeret i SBL ved fusion (stiplet linie mærket
(punkt (1) i figur 3A, højre panel), divideret med det totale intensitet nås efter alle fluoroforer er deponeret i SBL ved fusion (stiplet linie mærket  i figur 3). Men på grund af hurtig blegning i SBL, typisk ikke nået, og et nøjagtigt skøn over kræver montering frigivelseskinetikken til et udtryk 27 at incl udes både (Se 6.4.3) og . Separate udtryk findes i Stratton et al 27 for de tilfælde af pore-begrænset og diffusion begrænset release kinetik.; den bedste pasform tilfælde varierer fra arrangement til arrangement (se 6.5.1 i tilfælde af pore-begrænset kinetik).
i figur 3). Men på grund af hurtig blegning i SBL, typisk ikke nået, og et nøjagtigt skøn over kræver montering frigivelseskinetikken til et udtryk 27 at incl udes både (Se 6.4.3) og . Separate udtryk findes i Stratton et al 27 for de tilfælde af pore-begrænset og diffusion begrænset release kinetik.; den bedste pasform tilfælde varierer fra arrangement til arrangement (se 6.5.1 i tilfælde af pore-begrænset kinetik). - Beregn vesikel område for enkelte begivenheder fra 27
 , hvor er forankret SUV intensitet, er den gennemsnitlige intensitet af et enkelt lipid farvestof i SBL, er reduktionen intensitet faktor for et lipid farvestof, når den er i SUV forhold til når det er i SBL, ogation 1 "src =" / files / ftp_upload / 54349 / 54349eq12.jpg "/> er det kendte areal tæthed af lipid farvestoffer.
, hvor er forankret SUV intensitet, er den gennemsnitlige intensitet af et enkelt lipid farvestof i SBL, er reduktionen intensitet faktor for et lipid farvestof, når den er i SUV forhold til når det er i SBL, ogation 1 "src =" / files / ftp_upload / 54349 / 54349eq12.jpg "/> er det kendte areal tæthed af lipid farvestoffer.
- Fusion poreegenskaberne
- Hvis man antager, udgivelse er pore-begrænset, passer kinetikken lipid label release til 27
 , hvor
, hvor  er forankret SUV intensitet lige før fusion og de andre parametre er som defineret tidligere. Bruge værdien af opnået i 6.4.3 som en fast parameter, og udtrække de bedst tilpassede estimater for og
er forankret SUV intensitet lige før fusion og de andre parametre er som defineret tidligere. Bruge værdien af opnået i 6.4.3 som en fast parameter, og udtrække de bedst tilpassede estimater for og  .
. - Vurdere den del af tiden pore er åben, P 0, antager forsinkelse af lipid frigivelse skyldes pore flimren 27:60;
 =
=  , Hvor A ves er vesikel område (afsnit 6.4.5), b er pore højde (typisk for at være ~ 15 nm), r p ≈ 3 nm er den effektive pore radius som set af de spredende lipid etiketter og omfatter halvdelen dobbeltlaget tykkelse (~ 2 nm), er lipid diffusivitet (beregnet i 6.3), og er det tid til de lipider at blive frigivet fra SUV til SBL (fra 6.5.1).
, Hvor A ves er vesikel område (afsnit 6.4.5), b er pore højde (typisk for at være ~ 15 nm), r p ≈ 3 nm er den effektive pore radius som set af de spredende lipid etiketter og omfatter halvdelen dobbeltlaget tykkelse (~ 2 nm), er lipid diffusivitet (beregnet i 6.3), og er det tid til de lipider at blive frigivet fra SUV til SBL (fra 6.5.1). - For at bekræfte, at en nominel P 0> 1 indikerer en fuldt åben pore, P 0 = 1, så det passer til intensiteten tidsforløbet til ligning 4 i Stratton et al. 27, de forudsagte kinetik for en permanent åben pore. Denne pasform skal være bedre end fitting udtrykket i 6.5.1 for en permanent åben pore.
- Hvis man antager, udgivelse er pore-begrænset, passer kinetikken lipid label release til 27
 , Hvor w er radius af den blegede cirkel.
, Hvor w er radius af den blegede cirkel.  , Ved at plotte den overlevende funktion af forsinkelserne og montering til en eksponentiel.
, Ved at plotte den overlevende funktion af forsinkelserne og montering til en eksponentiel.  (punkt (1) i figur 3A, højre panel), divideret med det totale intensitet nås efter alle fluoroforer er deponeret i SBL ved fusion (stiplet linie mærket
(punkt (1) i figur 3A, højre panel), divideret med det totale intensitet nås efter alle fluoroforer er deponeret i SBL ved fusion (stiplet linie mærket  i figur 3). Men på grund af hurtig blegning i SBL,
i figur 3). Men på grund af hurtig blegning i SBL,  , hvor
, hvor  , hvor
, hvor  er forankret SUV intensitet lige før fusion og de andre parametre er som defineret tidligere. Bruge værdien af
er forankret SUV intensitet lige før fusion og de andre parametre er som defineret tidligere. Bruge værdien af  =
=  , Hvor A ves er vesikel område (afsnit 6.4.5), b er pore højde (typisk for at være ~ 15 nm), r p ≈ 3 nm er den effektive pore radius som set af de spredende lipid etiketter og omfatter halvdelen dobbeltlaget tykkelse (~ 2 nm),
, Hvor A ves er vesikel område (afsnit 6.4.5), b er pore højde (typisk for at være ~ 15 nm), r p ≈ 3 nm er den effektive pore radius som set af de spredende lipid etiketter og omfatter halvdelen dobbeltlaget tykkelse (~ 2 nm), Subscription Required. Please recommend JoVE to your librarian.
Representative Results
SBL Kvalitet
Det er afgørende at kontrollere kvaliteten og flydende SBL forud for fusion eksperiment. Fluorescensen i bunden, glas side af en mikrofluid kanal bør være ensartet, uden nogen åbenlyse mangler. Hvis en luftboble passerer selvom kanalen, det normalt efterlader synlige ar på SBL. Hvis der er så store skala ar / defekter, skal du ikke bruge denne kanal. Sommetider SUV'er kan klæbe på substratet, men undlader at briste og danne en kontinuerlig SBL. Hvis dette er tilfældet, kan fluorescensen stadig forekommer meget ensartet, men vil ikke inddrive efter blegning en lille region. Introduktion 10 mM Mg 2+ hjælper ofte at løse dette problem. Andre midler kan indføres redskaber bursting SUV'er, såsom andre divalente kationer 51, en polymeropløsning 52, eller osmotisk chok 25.
Temperaturændringer resultere i ændringer af SBL område, fordi det gennemsnitlige areal besat af en lipid afhænger af temperaturen. Forøgelse af temperaturen eller aftagende den ved 5-10 ° C eller mere kan resultere i overskydende område går i rør ekstruderes fra overfladen, eller mørke, SBL-free pletter optræder på overfladen, henholdsvis 53. Det er derfor vigtigt at holde temperaturvariationer på et minimum.
figur 5 for et SBL der var relativt fejlfri og væske.

Figur 5. FRAP at undersøge SBL flydende. En sekvens af billeder, der viser et belyst område afgrænset af den lukkede membran feltet (diameter 49 um) før (første ramme), og efter blegning med en stærk lys puls. Området er afbildet ved lav eksponering lys til at følge fluorescens inddrivelse ved diffusion af NBD mærkede lipider. Grafen viser den normaliserede fluorescens som en funktion af tid for flere kanaler fra forskellige dækglas angiver det høje metodens reproducerbarhed. Lipidsammensætningen af t-SBL var POPC: SAPE: DOPS: Cholesterol: hjerne PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5 mol% ) og LP = 20.000. Den blege området var 984.203 μm 3. Opsvinget tid og diffusionskoefficienten beregnet ud fra et anfald (se 6.1) var τ = 67,3 sek og D = 1,99 um 2 / sek, henholdsvis. Klik her for at se en større version af dette tal.
Single-farve Påvisning af Docking og Fusion Events Ved hjælp af en Lipid Label
Manuel optælling af docking og fusion begivenheder er en kedelig opgave, som der er behov for 100-400 hændelser pr betingelse for god statistik. Da SUV'er kan bevæge sig på SBL, inden den brændes, er der behov for en tracking software. SpeckleTrackerJ 26 plugin til ImageJ har funktioner, der specifikt er udviklet til at spore SUV'er til analyse af docking og fusion begivenheder. Ofte kun en brøkdel af forankret vesikler ender fusing;at beregne docking rate mange flere SUV'er skal spores end de, som fusionerer eller kun en del af filmen skal analyseres, indtil der detekteres et tilstrækkeligt antal hændelser for en pålidelig estimering. Hvis det primære fokus er den fusion sats og / eller docking-til-fusion forsinkelser, kun fusing SUV'er skal identificeres og spores.
Efter at sikre kvaliteten af SBL, er SUV'er indføres i kanalerne og deres fluorescens kontinuerligt spændt under TIR hjælp s-polariseret lys. For de indledende forsøg kan være nødvendigt at justere at tune indtrængningsdybden af den resulterende kortvarig bølge indfaldsvinklen af TIR exciteringsstrålen. Et repræsentativt enkelt SUV docking og fusion begivenhed er vist i figur 6A. Den vesikel forankret under rammen markeret D og fusion startede i ramme F. Om 1,3 sek senere (frame S) enkelt lipider blev mærkbar, da de spredes væk fra fusion siddee. Den totale fluorescensintensitet (summen af pixelværdier) i hvert felt er afbildet som en funktion af tid i figur 6B, med rammerne, hvor docking og fusion forekom angivet. Den kumulative antal fusionsbegivenheder som funktion af tiden afsættes i figur 6C, i fem forskellige film ved anvendelse af t-SNARE rekonstituerede SBLs (t-SBL, LP = 20.000, ca. 70 t-snarer per sq. Micron). Fusionshastigheden fra hver film blev beregnet ud fra en lineær tilpasning til hældning. Denne gennemsnitlige fusion sats ± SEM er vist som et søjlediagram i panel D. docking-til-fusion forsinkelse for den i figur 6A, B, eksempel var 0,07 sek. Fordelingen af forsinkelser er beregnet på lignende måde for i alt 158 hændelser er vist i figur 6D som en overlevende plot, dvs. sandsynligheden for, at en forankret vesikel endnu ikke har fusioneret med en given forsinkelse. Omkring 1/2 af disse vesikler fusioneret inden for 100 ms. Med stigende kolesteroltal, den distribution af docking-til-fusion forsinkelser afkorter, selvom lipid diffusion bremser 27.
Det er vigtigt at køre kontrolforsøg parallelt. På samme dækglas, hvor dataene i figur 6B, D blev erhvervet, blev kontrolbetingelser testet i de omkringliggende kanaler. I en kanal, har SBL ikke indeholder nogen t-snarer. Ud af 5 film varede 60 sek hver, blev kun 7 fusionsbegivenheder registreres i forhold til 158, når de t-snarer blev inkluderet (figur 6C). I en anden kanal, blev det opløselige cytoplasmatiske domæne af v-SNARE VAMP2 (CDV) medfølger. CDV binder t-snarer på SBL, forhindrer binding af fuld-længde VAMP2 der er på SUV'er. I fem 60 sec film, blev der ikke fusionsbegivenheder detekteret i nærvær af CDV (figur 6C).

Figur 6. SUV-SBL docking og fusion begivenheder. (A) Sekvens af billeder af en v-SUV fusion begivenhed med t-SBL afbildet af pTIRF mikroskopi efter spredning af LR-PE. Billeder er taget hver 18 ms, kun hver anden ramme er vist. D og F markerer starten af docking og fusion hhv. Individuelle lipid etiketter blevet genkendeligt som de diffunderer væk fra fusionsstedet og hinanden (S). Frame dimensioner: 16,7 um x 16,7 um. (B) Fluorescens intensitet som funktion af tid for fusion begivenhed vist i (A). (C) Akkumuleret antal fusionsbegivenheder som funktion af tid for v-SUV'er mærket med DiD fra et andet sæt af eksperimenter. (D) den normaliserede fusion rate (gennemsnit ± SEM, sx um 2 x pM SUV) -1, idet det antages 2 x 10 4 lipider pr SUV og (E) overlevelsessandsynligheden for en given tid efter docking af vesiklen. Sammensætningen til alle SBLs var POPC: SAPE: DOPS: Cholesterol: hjerne PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5 mol%) og LP = 20.000. v-SUV'er blev sammensat af POPC: SAPE: DOPS: Kolesterol: PEG2000-DOPE: LR-PE (57,4: 15: 12: 10: 4,6: 1 mol%) og LP = 200 for A og B. For CE, v -SUV sammensætning var ens, bortset gjorde, var i stedet brugt af LR-PE. klik her for at se en større version af dette tal.
Lipid Diffusivitet, vesikelstørrelse, og Fusion Pore Properties
Den fælles-molekyle følsomheden af analysen tillader udvinding af flere parametre ud over de docking og fusion satser og docking-til-fusion forsinkelse distributioner. Enkelt, fluorescensmærkede lipider bliver mærkbar, når de diffunderer væk fra fusionen (figur 6A). Sporing these direkte giver lipid diffusivitet 26, 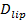 Og en digital måling af blegning sats for fluoroforer i SBL 27. Som nævnt i indledningen, fluorescensemissionen af etiketterne afviger i SUV versus i SBL, skyldes tre faktorer, der er generelt vanskeligt at udrede: (i) dequenching af etiketter ved fusion, på grund af fortynding, når de overføres til SBL , (ii) en højere excitation intensitet i SBL grund af henfald af det flygtige felt genereret af TIR, og (iii) orienteringen af fluoroforen overgangen dipoler i forhold til polarisationen af excitationsstrålen. Størrelsen af faktor (ii) afhænger af de relative størrelser af det flygtige felt henfald længde og vesikel størrelse og er forskelligt for hver fusion begivenhed grund dispersitet i SUV størrelser. Faktor (iii) afhænger af fluoroforen og polarisering anvendes. Snarere end at forsøge at di sentangle disse effekter, kan man måle størrelsen af fluorescensen ændringen direkte ved at sammenligne de totale pixelværdier i et område, der omgiver en SUV før fusion og efter at alle fluoroforer er deponeret i SBL efter fusion (figur 3A (1) vs. ( 2)). I modsætning til tidligere arbejde 29,44, bør regionen vælges tilstrækkeligt stor, så der ikke fluoroforer vil have forladt det via diffusion for de tider, der anses efter fusion. At vælge en 30 x 30 pixel region (8 um x 8 um) til analyse op til 1,6 sek efter fusion er normalt tilstrækkelig 27. Ignorerer blegning for øjeblikket, er forholdet mellem
Og en digital måling af blegning sats for fluoroforer i SBL 27. Som nævnt i indledningen, fluorescensemissionen af etiketterne afviger i SUV versus i SBL, skyldes tre faktorer, der er generelt vanskeligt at udrede: (i) dequenching af etiketter ved fusion, på grund af fortynding, når de overføres til SBL , (ii) en højere excitation intensitet i SBL grund af henfald af det flygtige felt genereret af TIR, og (iii) orienteringen af fluoroforen overgangen dipoler i forhold til polarisationen af excitationsstrålen. Størrelsen af faktor (ii) afhænger af de relative størrelser af det flygtige felt henfald længde og vesikel størrelse og er forskelligt for hver fusion begivenhed grund dispersitet i SUV størrelser. Faktor (iii) afhænger af fluoroforen og polarisering anvendes. Snarere end at forsøge at di sentangle disse effekter, kan man måle størrelsen af fluorescensen ændringen direkte ved at sammenligne de totale pixelværdier i et område, der omgiver en SUV før fusion og efter at alle fluoroforer er deponeret i SBL efter fusion (figur 3A (1) vs. ( 2)). I modsætning til tidligere arbejde 29,44, bør regionen vælges tilstrækkeligt stor, så der ikke fluoroforer vil have forladt det via diffusion for de tider, der anses efter fusion. At vælge en 30 x 30 pixel region (8 um x 8 um) til analyse op til 1,6 sek efter fusion er normalt tilstrækkelig 27. Ignorerer blegning for øjeblikket, er forholdet mellem  før og efter fusion giver reduktionen fluorescensintensitet faktor,
før og efter fusion giver reduktionen fluorescensintensitet faktor, 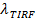 . Vesiklen størrelse kan estimeres gives fluorescensintensiteten af en enkelt etiket i SBL (Iles / ftp_upload / 54349 / 54349eq6.jpg "/>), den faktor, som dens intensitet vil blive reduceret, hvis det var i en SUV ( ), Den kendte densitet af labels i SUV, og på arealet et lipid. I praksis blegning er normalt hurtig nok, at når alle markører er deponeret i SBL, har nogle allerede bleget. Af denne grund, for en nøjagtig estimering, reduktion fluorescensintensitet faktor, Og lipid release tid,
. Vesiklen størrelse kan estimeres gives fluorescensintensiteten af en enkelt etiket i SBL (Iles / ftp_upload / 54349 / 54349eq6.jpg "/>), den faktor, som dens intensitet vil blive reduceret, hvis det var i en SUV ( ), Den kendte densitet af labels i SUV, og på arealet et lipid. I praksis blegning er normalt hurtig nok, at når alle markører er deponeret i SBL, har nogle allerede bleget. Af denne grund, for en nøjagtig estimering, reduktion fluorescensintensitet faktor, Og lipid release tid,  Er bedst ekstraheret fra en pasform til tidsforløbet af de samlede pixelværdier, der tager højde blegning 27. Den blegning på etiketter i SBL uafhængigt estimeret ud fra enkelt lipid farvestof sporing, eller ved at montere en eksponentiel til den samlede intensitet profil,
Er bedst ekstraheret fra en pasform til tidsforløbet af de samlede pixelværdier, der tager højde blegning 27. Den blegning på etiketter i SBL uafhængigt estimeret ud fra enkelt lipid farvestof sporing, eller ved at montere en eksponentiel til den samlede intensitet profil,  , Ved lange tider (f.eks regime (3) i figur 3). Kendskab til størrelsen af en fusing vesikel og lipid diffusivitet muliggør en sammenligning af den faktiske lipid release tid, , Til diffusionstiden af et lipid omkring vesiklen
, Ved lange tider (f.eks regime (3) i figur 3). Kendskab til størrelsen af en fusing vesikel og lipid diffusivitet muliggør en sammenligning af den faktiske lipid release tid, , Til diffusionstiden af et lipid omkring vesiklen  , Hvor A VES er vesiklen område og er lipid diffusivitet. Hvis de to tidsskalaer er sammenlignelige, pore præsenterer lidt modstand til lipid release, og frigivelse er diffusion-begrænset. Hvis derimod
, Hvor A VES er vesiklen område og er lipid diffusivitet. Hvis de to tidsskalaer er sammenlignelige, pore præsenterer lidt modstand til lipid release, og frigivelse er diffusion-begrænset. Hvis derimod  Så pore forsinker lipid flux betydeligt. En kvantitativ måling af retardering er "pore åbenhed", P 0, lig med forholdet
Så pore forsinker lipid flux betydeligt. En kvantitativ måling af retardering er "pore åbenhed", P 0, lig med forholdet  gange en faktor of rækkefølge enhed relateret til poregeometri 27. For en to-stats-, åben / lukket pore, P 0 er den del af tiden i åben tilstand. For en flimrende pore, hvis størrelse varierer kontinuerligt, P 0 repræsenterer tidsgennemsnitlig radius i forhold til den helt åbne radius.
gange en faktor of rækkefølge enhed relateret til poregeometri 27. For en to-stats-, åben / lukket pore, P 0 er den del af tiden i åben tilstand. For en flimrende pore, hvis størrelse varierer kontinuerligt, P 0 repræsenterer tidsgennemsnitlig radius i forhold til den helt åbne radius.
Signal-støj-forholdet skal være tilstrækkeligt godt til sporing enkelt lipid fluoroforer. Dette er lettere at opnå, i ensfarvede eksperimenter, anvendelse af en poleret markør, såsom LR-PE.
Samtidig detektion af lipid og opløseligt Cargo Frigivelse
En repræsentativ begivenhed er vist i figur 7. Ved de høje koncentrationer, der anvendes til indkapsling, SRB fluorescens indledningsvis selvstændig standset. Derfor er de fleste kuperet v-SUV'er er kun påvises ved deres havde tilkendegivet 27. En kraftig stigningi DiD fluorescens markerer lipid blanding, som i ensfarvede lipid-label forsøg. Starten af lipid udgivelse faldt sammen med starten på en stigning i SRB fluorescens. Denne stigning var mest sandsynligt på grund af SRB dequenching som SRB molekyler undslap SUV og den resterende SRB blev fortyndet til under selvstændige quenching koncentrationer. Interessant SRB fluorescens nåede et stabilt plateau. Hvis fusion pore forblev åbne, ville man forvente SRB signal at stige til et maksimum på grund af dequenching, efterfulgt af et fald til baggrund som den resterende SRB forlod SUV, eller hvis SUV kollapsede i SBL. Den stabile plateau sandsynligvis indikerer pore genforseglet efter frigivelse alle lipid etiketter og en del af den opløselige last.
I nogle tilfælde blev signal lipid frigivelse ikke ledsaget af nogen påviselige indhold frigive signaler. Dette kunne være enten fordi porerne var for små til at tillade passage af SRB, eller noglehvordan SUV allerede havde mistet sin opløselige last. For at skelne mellem disse to muligheder, kan analyseres blegning af dokkede SUV'er. Blegning af lipid etiket ikke reducerer dets fluorescenssignal inden for få sekunder, mens blegning af de oprindeligt selvstændige slukket SRB resulterer i en forøgelse af SRB signal som en brøkdel af SRB er foto-konverteret til andre produkter og selv-quenching er lettet. Mens de fleste forankret SUV'er (48 ud af 58 analyserede hændelser), der vises den forventede stigning i SRB fluorescens i længere tid ad belysning, gjorde en lille fraktion (10/58) ikke viser nogen SRB signaler overhovedet. Således kan en lille del af de SUV'er mister deres opløselige indhold i perioden mellem deres fremstilling og anvendelse i eksperimenterne.
I mere end 80% af begivenheder (74 ud af 91), hvor både SRB og gjorde signaler kunne følges, SRB fluorescens steget og forblev på et stabilt plateau 27. I ~ 20% af tisse tilfælde den resterende SRB fluorescens blev pludselig frigivet op til et par sekunder senere. Dette kan enten skyldes en genåbning af pore muliggør en hurtig, fuld frigørelse indenfor 1-2 rammer (18-36 ms) eller en sprængning af SUV grundet akkumuleret foto-skade 16. Uanset deres oprindelse, denne anden, hurtig frigivelse af SRB udelukker den mulighed, at den tilbageværende SRB signal efter den oprindelige udgivelse kan skyldes SRB forbliver indesluttet i mellemrummet mellem SBL og glassubstratet efter fuld frigivelse. En sådan ordning blev foreslået i en tidligere undersøgelse, hvor SBL blev støttet direkte på glas, med lidt plads mellem de to 16. Derimod bør de PEGylerede lipider, der anvendes i dette assay giver 4-5 nm af plads mellem SBL og glasset 25, således at SRB at diffundere væk fra fusionen site.

Figur 7. Simultan EOU lipid og indhold udgivelse. Fluorescens signaler fra lipid (blå) og indhold (rød) markør for en SUV-SBL fusion begivenhed (for detaljer se tekst). Snapshots fra hver kanal er vist nederst (4,0 um bred). Tegnefilm (i midten) viser de forskellige stadier. (1) SUV dokker, har fluorescens er lav. (2) Fusion tillader overførsel DID ind SBL og gjorde fluorescens stiger. SRB signal øger også som selvstændige quenching er lettet ved udslip af SRB gennem poren. (3) SRB signalet forbliver stabile, mens DID molekyler diffunderer i SBL indikerer en mulig genlukning af pore. (4) SRB signaler falde meget hurtigt, på grund af hurtig fuld fusion eller liposom sprængning. Sammensætningen af v-SUV'er var POPC: SAPE: DOPS: PEG2000-DOPE: gjorde (67: 15: 12: 5: 1 mol%), LP 200, mens t-SBLs blev sammensat af POPC: SAPE: DOPS: hjerne PI (4,5) P2: PEG2000-DOPE: NBD-PE (64,5: 15: 12: 3: 5: 0,5 mol%) og LP = 20.000. = "_ Blank"> Klik her for at se en større version af dette tal.
FRAP_instructions.pdf -. Vejledning til opsætning af en FRAP sekvens Klik her for at downloade denne fil.
ReadMe_FRAP.txt -. Tekstfil, der beskriver proceduren for at analysere FRAP recovery data ved hjælp af MatLab kode, der leveres Klik her for at downloade denne fil.
EK_FRAPbatch.m - Matlab fil til batch-analysere fluorescens bedring efter fotoblegning. Denne fil kalder MATLAB underrutiner EK_getFRAPvals.m og EK_SoumpassisFit.m."_blank"> Klik her for at downloade denne fil.
EK_getFRAPvals.m -. Matlab fil, subrutine kaldes af EK_FRAPbatch.m Klik her for at downloade denne fil.
EK_SoumpassisFit.m -. Matlab fil, subrutine kaldes af EK_FRAPbatch.m Klik her for at downloade denne fil.
JN150923c1_FRAP_list.txt -. Et eksempel på en liste over filnavne at være batch-analyseret af de vigtigste MATLAB script EK_FRAPbatch.m Klik her for at downloade denne fil.
DetFølgende filer TIF stakke og de tilsvarende tekstfiler for fem eksempler på FRAP sekvenser, der kan analyseres ved at køre EK_FRAPbatch.m og vælge den JN150923c1_FRAP_list.txt filen, når du bliver bedt om at vælge en liste fil.
JN150923c1_F1_MMStack_Pos0.ome.tif -. FRAP billedstak Klik her for at downloade denne fil.
JN150923c1_F1_MMStack_Pos0_metadata.txt -. Svarende tekstfil med metadata Klik her for at downloade denne fil.
JN150923c1_F2_MMStack_Pos0.ome.tif -. FRAP billedstak Pllethed klik her for at downloade denne fil.
JN150923c1_F2_MMStack_Pos0_metadata.txt -. Svarende tekstfil med metadata Klik her for at downloade denne fil.
JN150923c1_F3_MMStack_Pos0.ome.tif -. FRAP billedstak Klik her for at downloade denne fil.
JN150923c1_F3_MMStack_Pos0_metadata.txt -. Svarende tekstfil med metadata Klik her for at downloade denne fil.
JN150923c1_F4_MMStack_Pos0.ome.tif - FRAP billedstak. Klik her for at downloade denne fil.
JN150923c1_F4_MMStack_Pos0_metadata.txt -. Svarende tekstfil med metadata Klik her for at downloade denne fil.
JN150923c1_F5_MMStack_Pos0.ome.tif -. FRAP billedstak Klik her for at downloade denne fil.
JN150923c1_F5_MMStack_Pos0_metadata.txt -. Svarende tekstfil med metadata Klik her for at downloade denne fil.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
En vellykket gennemførelse af SUV-SBL fusion assay beskrevet her afhænger kritisk på flere vigtige skridt, såsom funktionel rekonstitution af proteiner i liposomer, opnå god kvalitet SBLs, og vælge de rigtige imaging parametre til at opdage enkelte molekyler. Selv om det kan tage lidt tid og kræfter til at lykkes, når analysen gennemføres med succes, det giver et væld af oplysninger om fusionsprocessen ikke nogen anden in vitro-fusion assay diskuteret i Introduction. De satser, hvor SUV'er dock på og sikring med SBL, fordelingen af docking-til-fusion gange, lipid diffusivitet, størrelsen af hver sammensmelte liposom, og om lipid overgang til en given fusion begivenhed er diffusion eller pore-kontrollerede dåse bestemmes. Hvis frigivelse er meget langsommere end forventet fra diffusion, en model, der antager pore flimrende ligger til grund for retardering af lipid frigivelse udbytter, P 0, den del af tiden pore is åbent i fusion 27.
Tidligere arbejde sondering enkelt SUV-SUV fusion 18-20,28 eller fusion af liposomer og virale partikler til SBLs 15-17,21,22,33,44 påberåbes FRET og selv-quenching mellem farvestof molekyler. I modsætning hertil vi i høj grad undgå farvestof dequenching at sonden kinetik lipid label overførsel til SBL, af to grunde. Første, FRET effektivitet er en yderst ulineær reporter af lipid frigivelse. Små ændringer i den inter-dye afstanden d resulterer i store ændringer i FRET effektivitet, men kun for et begrænset udvalg af D / R 0-værdier, hvor R 0 er Forster afstand. Dvs. en latenstid mellem den faktiske farvestof fortynding og en observeret stigning i dequenching signal kan forekomme på grund af den tid, der kræves for at falde til en værdi tæt på R 0. Tilsvarende er der ringe ændring i dequenching signalet, når farvestoffet tæthed aftager tilstrækkeligt ved lipid frigivelse, således at  1.4, selv om en betydelig mængde farvestof stadig i vesiklen. Disse egenskaber gør dequenching et godt værktøj til at opnå en stigning i signalet for at detektere fusionsbegivenheder, men komplicere kvantitativ kinetisk analyse af lipid frigørelse med høj tidsopløsning. I modsætning hertil er ændringer i intensitet på grund af dipol omlægning og flygtige felt virkninger som en fluorofor overførsler fra en SUV i SBL lineært relateret til den del af farvestoffet stadig tilbage i SUV. For det andet kan høje koncentrationer af lipid etiketter kræves for selv-quenching påvirke fusionsprocessen selv.
1.4, selv om en betydelig mængde farvestof stadig i vesiklen. Disse egenskaber gør dequenching et godt værktøj til at opnå en stigning i signalet for at detektere fusionsbegivenheder, men komplicere kvantitativ kinetisk analyse af lipid frigørelse med høj tidsopløsning. I modsætning hertil er ændringer i intensitet på grund af dipol omlægning og flygtige felt virkninger som en fluorofor overførsler fra en SUV i SBL lineært relateret til den del af farvestoffet stadig tilbage i SUV. For det andet kan høje koncentrationer af lipid etiketter kræves for selv-quenching påvirke fusionsprocessen selv.
For polariseret TIRF excitation, en hjemme-bygget setup som i figur 4 er omkostningseffektiv og giver fremragende kontrol over polarisationsegenskaber. Men et hjem bygget opsætning er ikke nødvendig, da i det mindste nogle kommercielle TIRF mikroskoper bruge polariseret excitation og off-the-shelftofarvede emission splitting moduler er tilgængelige. En kommerciel mikroskop, der anvender en polarisationsbevarende (PM) fiber til at koble polariserede excitationslys udgår fra laseren ind i mikroskopet blev med succes anvendt tidligere 25-27. På et sådant instrument, kan polarisationen drejes simpelthen ved at dreje PM fiber input til mikroskopets TIRF enhed. I modsætning hertil kan nogle andre kommercielle opsætninger anvende en polariserende stråledeler i TIRF enhed, hvilket gør polarisering justeringer mere vanskelig eller umulig. Desværre er polarisering egenskaber ofte ikke angivet for kommercielle systemer. Således er det afgørende for brugere, der ikke ønsker at konstruere en hjemmelavet indbygget system til at diskutere polarisering egenskaber, og at prøve en demo-enhed, før man investerer i en bestemt kommerciel mulighed.
Samtidig overvågning af lipid og opløselig last release indeholder yderligere oplysninger om fusionsprocessen. Mange porer synes at genlukke efter releAsing kun en brøkdel af den opløselige etiket indkapslet i en SUV 27. Normalt alle lipid etiketter er udgivet af den tid, pore genforsegler, så pore genlukning var ikke mistænkt tidligere fra overvågning lipid frigivelse alene. Faktisk, overvågning lipid frigivelse alene, havde Kiessling et al. 29 foreslået overførsel af mærkede lipider i SBL skete via ekstremt hurtige sammenbrud af hele SUV membran ind i SBL.
Genlukning af fusion porer efter delvis frigivelse af indhold tidligere blev observeret i en liposom-tøjret dobbeltlag patch fusion assay. Rawle et al. 54 forberedte fritstående dobbeltlag patches, der blev forankret på et dækglas med ~ 8 nm lange DNA afstandsstykker. De indførte derefter liposomer hvis fusion til dobbeltlaget plaster blev drevet ved hybridisering af komplementære enkeltstrengede DNA-lipid-konjugater forankret til liposomet og patch membraner. 12% af alle hændelser var forbigående fusioner, hvilket resulterer i delvisial frigivelse af liposomindholdet. Liposom bursting begivenheder, set i nogle tidligere SUV-SBL fusion assays 44, var meget sjældne. Forfatterne foreslog ~ 8 nm pladsen under dobbeltlaget plaster, forhindrer dobbeltlag-substrat-interaktioner, kan have tilladt ikke-utætte overførsel af liposomindholdet til rummet under dobbeltlaget. Desuden blev diffusion af de frigivne opløselige etiketter ikke signifikant hindres i rummet mellem den plane dobbeltlag og glassubstratet. I assayet beskrevet her, er den plane dobbeltlag ligeledes holdes ~ 5 nm væk fra dækglasset ved tilstedeværelsen af poly (ethylenglycol) -lipid konjugater.
De supplerende oplysninger fra tofarvet overvågning af opløselige og lipid gods kommer med nogle kompromisser. Først, forberedelse af SUV'er med de to mærker er noget mere involveret end herunder lipid etiketter alene. For det andet er der en vis uundgåelig tab af signal-støj når de overvåger signalerne fra lipid etiketter ved than samme erhvervelse sats som i forsøg lipid release ensfarvede (15-20 ms pr ramme). Noget af dette tab skyldes det faktum, at gjorde, anvendes som et lipid etiket sammen med den opløselige SRB markering, er ikke så lyse som LR-PE anvendes i ensfarvede eksperimenter. Yderligere tab skyldes højere baggrund på grund af samtidig to-laser excitation og yderligere komponenter i den optiske vej (figur 4). Hidtil har den lavere signal-støj forhindret os pålidelig sporing af single DiD etiketter i tofarvede eksperimenter. Dette vil til gengæld forhindrede en grundig analyse for at udtrække alle de oplysninger, der kan være med LR-PE målinger ensfarvede. Denne udfordring kan overvindes i fremtiden ved at optimere akkvisitionsbetingelser og afprøve andre par af kompatible etiketter med forbedrede egenskaber.
Samtidig overvågning af opløselige og lipid last frigivelse kan også opdage hemifusion, en tilstand, hvor de proksimale foldere har smeltet, men den distalefoldere har ikke. Faktisk kan sådanne tofarvede eksperimenter giver den eneste afgørende vurdering af tilstedeværelsen af hemifusion i andre in vitro fusion assays 18,20. Dog kan hemifusion pålideligt påvises i ensfarvet SUV-SBL eksperimenter overvågning lipid frigivelse alene 22,44. I sådanne eksperimenter, omkring 1/2 den initiale lipid label intensiteten af en SUV forbliver på fusion stedet. Den resterende intensitet kan forsvinde, når de distale foldere sammensmelte nogen tid senere, hvis stedet endnu ikke er bleget.
Anvendelse af pegyleret lipider at indføre en blød pude mellem SBL og støtte glasset har været nøglen i at reproducere kravet SNAP25 af exocytose. Protokollen resulterer i en PEG børste dækker den side af SBL, som vender mod strømningskanalen samt. Dette har nogle fordele, såsom at reducere ikke-specifikke interaktioner mellem SUV'er og SBL, som forklaret i Introduction. Hvis proteiner, som interagerer med phospholipiderne iSBL introduceres i assayet vil imidlertid PEG børste præsentere en sterisk barriere mod sådanne interaktioner. Således til nogle anvendelser vil det være ønskeligt at inkludere PEG lag asymmetrisk, kun mellem SBL og dækglas. Den side af SBL vender strømningskanalen ville så kunne interagere med phospholipid-bindende proteiner, såsom synaptotagmin. Dette kan opnås ved hjælp af bifunktionelle PEG-kæder, som kan være kovalent forbundet med dækglasset i den ene ende og bærer en lipid anker i den anden. Langmuir-Blodgett fremgangsmåder kan derefter anvendes til at deponere et lipid monolag på PEG laget 55. Liposomer indført over denne hydrofobe overflade sikring med det, der danner et dobbeltlag 56. Det er håbet, at denne fremgangsmåde vil blive kombineret med mikrofluidik i fremtiden.
Det forventes, at SUV-SBL assay præsenteret her, vil være nyttig i at udforske rollen af yderligere proteiner, der interagerer med kneb,såsom Munc18 / Sec1 familie, calcium sensor synaptotagmin-1, og complexin.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne erklærer, at de ikke har nogen konkurrerende finansielle interesser.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| Milli-Q (MQ) water | Millipore | ||
| KOH | J.T. Baker | 3040-05 | |
| Ethanol 190 Proof | Decon | ||
| Isopropanol | Fisher Chemical | A416P4 | |
| HEPES | AmericanBio | AB00892 | |
| Sodium Cholride (KCl) | AmericanBio | AB01915 | |
| Dithiothreitol | AmericanBio | AB00490 | |
| N-[2-hydroxyethyl] piperazine-N'-[2-ethanesulfonic acid] (HEPES) | AmericanBio | AB00892 | |
| EGTA | Acros Organics | 409911000 | |
| Buffers | |||
| HEPES-KOH buffer (pH 7.4) | 25 mM HEPES-KOH, 140 mM KCl, 100 μM EGTA, 1 mM DTT | ||
| Solvents | |||
| Chloroform | J.T. Baker | 9180-01 | in glass bottle, CAUTION, wear PPE |
| Methanol | J.T. Baker | 9070-03 | in glass bottle, CAUTION, wear PPE |
| Liposome preparation | |||
| Gastight Hamilton syringe | Hamilton | var. sizes | only use glass sringe with solents (Chlorophorm/ Methanon, 2:1, v/v) |
| Glass tubes Pyrex Vista 11 ml, 16x100 mm screw cap culture tube | Pyrex | 70825-16 | clean thoroughly, rinse with chloroform |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine, 16:0-18:1 PC (POPC) | Avanti Polar Lipids | 850457 | Lipids come dissolved in CHCl3 or as lyphilized powder in sealed vials. Aliquot upon opening. Store extra as dried lipid films under inert atmosphere at -20 °C. Keep stocks in CHCl3/MeOH (2:1, v/v) at -20 °C. let come to RT before opening |
| 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (sodium salt), 18:1 PS (DOPS) | Avanti Polar Lipids | 840035 | Same as above. |
| 1-stearoyl-2-arachidonoyl-sn-glycero-3-phosphoethanolamine, 18:0-20:4 PE (SAPE) | Avanti Polar Lipids | 850804 | Same as above. |
| L-α-phosphatidylinositol-4,5-bisphosphate (Brain, Porcine) (ammonium salt), Brain PI(4,5)P2 | Avanti Polar Lipids | 840046 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(7-nitro-2-1,3-benzoxadiazol-4-yl) (ammonium salt), 18:1 NBD PE | Avanti Polar Lipids | 810145 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene glycol)-2000] (ammonium salt), 18:1 PEG2000 PE | Avanti Polar Lipids | 880130 | Same as above. |
| cholesterol (ovine wool, >98%) | Avanti Polar Lipids | 700000 | Same as above. |
| DiD' oil; DiIC18(5) oil (1,1'-Dioctadecyl-3,3,3',3'-Tetramethylindodicarbocyanine Perchlorate) | Molecular Probes | D-307 | |
| Rotavapor R-210 | Buchi | R-210 | heat bath above Tm of lipids used |
| OG n-Octyl-β-D-Glucopyranoside | Affymetrix | 0311 | store at -20 °C, let come to RT before opening |
| Shaker - Eppendorf Thermomixer R | Eppendorf | ||
| Slide-A-Lyze Dialysis Cassettes, 20k MWCO, 3 ml | life technologies | 66003 | |
| Bio-Beads SM-2 Adsorbents | Bio-Rad | 1523920 | |
| OptiPrep Density Gradient Medium | Sigma-Aldrich | D1556 | |
| Ultracentrifugation tube, Thinwall, Ultra-Clear, 13.2 ml, 14 x 89 mm | Beckman Coulter | 41121703 | |
| Beckman SW41 Ti rotor | |||
| SuflorhodamineB | Molecular Probes | S-1307 | |
| Econo-Column Chromatography Columns, 2.5 × 10 cm | Bio-Rad | 7372512 | |
| Sepharose CL-4B | GE Healthcare | 17-0150-01 | |
| SYPRO Orange Protein Gel Stain | Molecular Probes | S-6650 | 5,000x Concentrate in DMSO |
| PDMS block | |||
| Sylgard 184 Silicone elastomer kit, PDMS | Dow Corning | 3097358-1004 | |
| Pyrex glass petri dish, 150 x 20 mm, complete with cover | Corning | 3160-152 | |
| Hole puncher - Reusable Biopsy Punch, 0.75 mm | World Precision Instruments | 504529 | |
| Manual Hole Punching Machine | SYNEO | MHPM-UNV | |
| Drill .035 x .026 x 1.5 304 SS TiN coated round punch | SYNEO | CR0350265N20R4 | drill diameter: 0.9 mm, punch bore size: 0.74 mm |
| Tygon Microbore tubing, 0.25 mm ID, 0.76 mm OD | Cole-Parmer | 06419-00 | 0.010" ID, 0.030" OD |
| Silicone Tubing (0.51 mm ID, 2.1 mm OD | Cole-Parmer | 95802-00 | 0.020" ID, 0.083" OD |
| Cover glass - cleanroom cleaned | |||
| Schott Nexterion cover slip glass D | Schott | 1472305 | |
| plasma cleaner | Harrick | PDC-32G | |
| pTIRF setup and accessories | |||
| IX81 microscope body | Olympus | IX81 | |
| EM CCD camera | Andor | ixon-ultra-897 | |
| Thermo Plate, heated microscope stage | Tokai Hit | MATS-U52RA26 | |
| 1 ml hamilton glass syringes (4x) | Hamilton | 81365 | |
| syringe pump | kd Scientific | KDS-230 | |
References
- Sudhof, T. C., Rothman, J. E. Membrane fusion: grappling with SNARE and SM proteins. Science. 323, 474-477 (2009).
- Wickner, W., Schekman, R. Membrane fusion. Nat Struct Mol Biol. 15, 658-664 (2008).
- Harrison, S. C. Viral membrane fusion. Nat Struct Mol Biol. 15, 690-698 (2008).
- Jahn, R., Scheller, R. H. SNAREs--engines for membrane fusion. Nat Rev Mol Cell Biol. 7, 631-643 (2006).
- Lindau, M., Alvarez de Toledo, G. The fusion pore. Biochim Biophys Acta. 1641, 167-173 (2003).
- Staal, R. G., Mosharov, E. V., Sulzer, D. Dopamine neurons release transmitter via a flickering fusion pore. Nat Neurosci. 7, 341-346 (2004).
- Wu, Z., et al. Nanodisc-cell fusion: Control of fusion pore nucleation and lifetimes by SNARE protein transmembrane domains. Sci. Rep. 6, 27287 (2016).
- Alabi, A. A., Tsien, R. W. Perspectives on kiss-and-run: role in exocytosis, endocytosis, and neurotransmission. Ann Rev Physiol. 75, 393-422 (2013).
- Rossetto, O., Pirazzini, M., Montecucco, C. Botulinum neurotoxins: genetic, structural and mechanistic insights. Nature Rev Microbiol. 12, 535-549 (2014).
- Weber, T., et al. SNAREpins: minimal machinery for membrane fusion. Cell. 92, 759-772 (1998).
- Nickel, W., et al. Content mixing and membrane integrity during membrane fusion driven by pairing of isolated v-SNAREs and t-SNAREs. Proc Natl Acad Sci U S A. 96, 12571-12576 (1999).
- McNew, J. A., et al. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature. 407, 153-159 (2000).
- Melia, T. J., You, D. Q., Tareste, D. C., Rothman, J. E. Lipidic antagonists to SNARE-mediated fusion. J Biol Chem. 281, 29597-29605 (2006).
- Hernandez, J. M., et al. Membrane fusion intermediates via directional and full assembly of the SNARE complex. Science. 336, 1581-1584 (2012).
- Fix, M., et al. Imaging single membrane fusion events mediated by SNARE proteins. Proc Natl Acad Sci U S A. 101, 7311-7316 (2004).
- Bowen, M. E., Weninger, K., Brunger, A. T., Chu, S. Single molecule observation of liposome-bilayer fusion thermally induced by soluble N-ethyl maleimide sensitive-factor attachment protein receptors (SNAREs). Biophys J. 87, 3569-3584 (2004).
- Liu, T., Tucker, W. C., Bhalla, A., Chapman, E. R., Weisshaar, J. C. SNARE-driven, 25-millisecond vesicle fusion in vitro. Biophys J. 89, 2458-2472 (2005).
- Yoon, T. Y., Okumus, B., Zhang, F., Shin, Y. K., Ha, T. Multiple intermediates in SNARE-induced membrane fusion. Proc Natl Acad Sci U S A. 103, 19731-19736 (2006).
- Diao, J., et al. A single-vesicle content mixing assay for SNARE-mediated membrane fusion. Nat Commun. 1, 1-6 (2010).
- Kyoung, M., et al. In vitro system capable of differentiating fast Ca2+-triggered content mixing from lipid exchange for mechanistic studies of neurotransmitter release. Proc Natl Acad Sci U S A. 108, E304-E313 (2011).
- Domanska, M. K., Kiessling, V., Stein, A., Fasshauer, D., Tamm, L. K. Single vesicle millisecond fusion kinetics reveals number of SNARE complexes optimal for fast SNARE-mediated membrane fusion. J Biol Chem. 284, 32158-32166 (2009).
- Kreutzberger, A. J., Kiessling, V., Tamm, L. K. High Cholesterol Obviates a Prolonged Hemifusion Intermediate in Fast SNARE-Mediated Membrane Fusion. Biophys J. 109, 319-329 (2015).
- Schwenen, L. L., et al. Resolving single membrane fusion events on planar pore-spanning membranes. Sci Rep. 5, 12006 (2015).
- Karatekin, E., et al. A fast, single-vesicle fusion assay mimics physiological SNARE requirements. Proc Natl Acad Sci U S A. 107, 3517-3521 (2010).
- Karatekin, E., Rothman, J. E. Fusion of single proteoliposomes with planar, cushioned bilayers in microfluidic flow cells. Nat Protoc. 7, 903-920 (2012).
- Smith, M. B., et al. Interactive, computer-assisted tracking of speckle trajectories in fluorescence microscopy: application to actin polymerization and membrane fusion. Biophys J. 101, 1794-1804 (2011).
- Stratton, B. S., et al. Cholesterol Increases the Openness of SNARE-mediated Flickering Fusion Pores. Biophysical journal. 110, (2016).
- Diao, J., et al. Synaptic proteins promote calcium-triggered fast transition from point contact to full fusion. Elife. 1, e00109 (2012).
- Kiessling, V., Domanska, M. K., Tamm, L. K. Single SNARE-mediated vesicle fusion observed in vitro by polarized TIRFM. Biophys J. 99, 4047-4055 (2010).
- Blasi, J., et al. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature. 365, 160-163 (1993).
- Washbourne, P., et al. Genetic ablation of the t-SNARE SNAP-25 distinguishes mechanisms of neuroexocytosis. Nat Neurosci. 5, 19-26 (2002).
- Diaz, A. J., Albertorio, F., Daniel, S., Cremer, P. S. Double cushions preserve transmembrane protein mobility in supported bilayer systems. Langmuir. 24, 6820-6826 (2008).
- Floyd, D. L., Ragains, J. R., Skehel, J. J., Harrison, S. C., van Oijen, A. M. Single-particle kinetics of influenza virus membrane fusion. Proc Natl Acad Sci U S A. 105, 15382-15387 (2008).
- Albertorio, F., et al. Fluid and air-stable lipopolymer membranes for biosensor applications. Langmuir. 21, 7476-7482 (2005).
- Daniel, S., Albertorio, F., Cremer, P. S. Making lipid membranes rough, tough, and ready to hit the road. Mrs Bulletin. 31, 536-540 (2006).
- Gao, Y., et al. Single reconstituted neuronal SNARE complexes zipper in three distinct stages. Science. 337, 1340-1343 (2012).
- Kenworthy, A. K., Hristova, K., Needham, D., Mcintosh, T. J. Range and Magnitude of the Steric Pressure between Bilayers Containing Phospholipids with Covalently Attached Poly(Ethylene Glycol). Biophys J. 68, 1921-1936 (1995).
- Knoll, W., et al. Solid supported lipid membranes: New concepts for the biomimetic functionalization of solid surfaces. Biointerphases. 3, Fa125-Fa135 (2008).
- Quinn, P., Griffiths, G., Warren, G. Density of newly synthesized plasma membrane proteins in intracellular membranes II. Biochemical studies. J Cell Biol. 98, 2142-2147 (1984).
- Sund, S. E., Swanson, J. A., Axelrod, D. Cell membrane orientation visualized by polarized total internal reflection fluorescence. Biophys J. 77, 2266-2283 (1999).
- Johnson, D. S., Toledo-Crow, R., Mattheyses, A. L., Simon, S. M. Polarization-controlled TIRFM with focal drift and spatial field intensity correction. Biophys J. 106, 1008-1019 (2014).
- Anantharam, A., Onoa, B., Edwards, R. H., Holz, R. W., Axelrod, D. Localized topological changes of the plasma membrane upon exocytosis visualized by polarized TIRFM. J Cell Biol. 188, 415-428 (2010).
- Axelrod, D. Carbocyanine dye orientation in red cell membrane studied by microscopic fluorescence polarization. Biophys J. 26, 557-573 (1979).
- Wang, T., Smith, E. A., Chapman, E. R., Weisshaar, J. C. Lipid mixing and content release in single-vesicle, SNARE-driven fusion assay with 1-5 msec resolution. Biophys J. 96, 4122-4131 (2009).
- Chernomordik, L. V., Frolov, V. A., Leikina, E., Bronk, P., Zimmerberg, J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J Cell Biol. 140, 1369-1382 (1998).
- Takamori, S., et al. Molecular anatomy of a trafficking organelle. Cell. 127, 831-846 (2006).
- Wilhelm, B. G., et al. Composition of isolated synaptic boutons reveals the amounts of vesicle trafficking proteins. Science. 344, 1023-1028 (2014).
- Scott, B. L., et al. Liposome fusion assay to monitor intracellular membrane fusion machines. Methods Enzymol. 372, 274-300 (2003).
- Linkert, M., et al. Metadata matters: access to image data in the real world. J Cell Biol. 189, 777-782 (2010).
- Soumpasis, D. M. Theoretical analysis of fluorescence photobleaching recovery experiments. Biophys J. 41, 95-97 (1983).
- Ohki, S. A mechanism of divalent ion-induced phosphatidylserine membrane fusion. Biochim Biophys Acta. 689, 1-11 (1982).
- Berquand, A., et al. Two-step formation of streptavidin-supported lipid bilayers by PEG-triggered vesicle fusion. Fluorescence and atomic force microscopy characterization. Langmuir. 19, 1700-1707 (2003).
- Tamm, L. K., McConnell, H. M. Supported phospholipid bilayers. Biophys J. 47, 105-113 (1985).
- Rawle, R. J., van Lengerich, B., Chung, M., Bendix, P. M., Boxer, S. G. Vesicle fusion observed by content transfer across a tethered lipid bilayer. Biophys J. 101, L37-L39 (2011).
- Wagner, M. L., Tamm, L. K. Reconstituted syntaxin1a/SNAP25 interacts with negatively charged lipids as measured by lateral diffusion in planar supported bilayers. Biophys J. 81, 266-275 (2001).
- Kalb, E., Frey, S., Tamm, L. K. Formation of Supported Planar Bilayers by Fusion of Vesicles to Supported Phospholipid Monolayers. Biochimica Et Biophysica Acta. 1103, 307-316 (1992).
