

Summary
在这里,我们提出了一个协议来检测使用偏振TIRFM脂质体和微流体通道支持双层之间的单,SNARE介导的融合事件,与单分子敏感性和〜15毫秒的时间分辨率。脂质和可溶性货物放行可同时检测。脂质体大小,脂质扩散率,和融合孔性质被测量。
Abstract
在膜融合的泛在处理的融合孔的开口在两台以前分开的隔室之间的第一个连接。在通过胞外分泌神经递质或激素释放的融合孔可以瞬时打开和关闭多次,规范货物释放动力学。孔动力学也确定囊泡循环的模式;在短暂的“吻了就跑”融合不可逆转再加盖印章的结果,而扩张,从而达到完全融合。为了更好地了解哪些因素支配孔隙动力学,我们开发了一种测定中使用偏振全内反射荧光(TIRF)显微镜用单分子灵敏度和〜15毫秒的时间分辨率在体外系统中的生物化学良好定义并监控膜融合。融合的荧光标记的含V-SNARE蛋白(V-越野车)与平面双层轴承T-SNARE蛋白小单层囊泡,支持软聚合物垫(T-SBL,T-支持双层),被监视。该法采用微流体流动通道,以确保最少的样品消耗,同时提供越野车的密度不变。利用在熔化过程中从SUV脂质标签转移到SBL的快速信号增强,脂质染料转移的动力学进行监测。 TIRF显微镜的灵敏度可跟踪单个荧光脂质标签,从中脂质扩散和SUV的大小可以推断为每个融合事件。脂质染料释放时间可以更长比预期通过永久开放的毛孔畅通的通道。使用该假设脂质释放的相位差是由于孔隙闪烁,细孔“开放”,该时间孔隙保持熔融时打开的分数的模型,可以进行估计。可溶性标记可以在SUV的脂质和可溶性货物放行的同步监测进行封装。这种测量表明失去可溶性货物的一小部分经过一些孔隙可以重新密封。
Introduction
膜融合为脂质和蛋白质,分泌,施肥,发展的细胞内运输所需的通用生物过程,和包膜病毒进入宿主生物体1-3。对于大多数细胞内聚变反应包括通过胞吐的激素和神经递质的释放,融合2脂质双层的能量由形成同源可溶性N-乙基马来酰亚胺-敏感因子附着蛋白受体 (SNARE)的蛋白质之间的四螺旋束的提供,锚定在囊泡(v-SNARE)和分别的靶膜(叔SNARE)4,。突触小泡胞吐作用是最严格调控的聚变反应和动作电位1,4,5到货后一毫秒内发生。的融合孔,两个熔断隔间之间的初始连接,可以闪烁打开和再密封或不可逆5-7扩大之前关闭多次。前者结果在短暂的“亲吻与运行”的融合,而后者导致完全融合。理事融合这两种模式和调节孔径闪烁机制之间的平衡因素不能很好地理解5,8。
SNARE蛋白所需的胞吐作用;突触囊泡融合是由神经毒素9时SNARE蛋白裂解取消。使用小单层囊泡(SUV的)堆积聚变实验表明,斯耐尔不仅必需的,但也足以驱动膜融合10。在此批量测定中,重构用v-SNARE蛋白(V-SUV)的SUV被掺杂有荧光磷脂(N - (7-硝基2-1,3-苯并恶二唑-4-基)-phosphoethanolamine(NBD-PE)和( N - (丽丝胺罗丹明B磺酰基)-phosphoethanolamine(LR-PE)和含叔SNARE蛋白(T-SUV)最初的v型的SUV NBD-PE的荧光被荧光共振能量转移淬无标签的小泡混合(FRET ),以LR-PE作为实验室ELED v型越野车与未标记叔SUV的融合,在现在结合膜的荧光面密度降低,在NBD-PE的荧光所产生的增加报告脂质混合10的程度。象散法是容易设置和分析,它已被广泛用来研究SNARE介导的融合10-14的机制。然而,它有一些限制,如低灵敏度和时间分辨率差。最重要的是,作为一个集合进行测定,平均结果在使半融合中间体难以检测对接和融合区分,以及所有事件。
在过去的十年几组,包括我们,已经开发了新的分析方法来监控在单级囊泡融合15-27事件。厦和他的同事用拴在表面上的V-SUV和监控他们与免费的T SUV的融合18,19。脂质混合是一对脂相连的荧光团之间的FRET使用EM监视层状分别在v型和叔越野车,使用全内反射荧光(TIRF)显微镜18。后来,Brunger的实验室使用单一脂质标签的物种连同内容标记物的脂质和内容混合20,28同时检测。既脂质和内容标记被包括在高,自猝灭浓度;融合SUV的未标记导致荧光脱猝灭20,28。
其他人已经融合v型越野车与T-SNARE蛋白15-17,21-27,29重组平面双层。目标(含T-SNARE)双层更好地模拟与一台质膜小,高度弯曲的囊泡的生理融合过程中的平面形状。该斯泰纳姆组采用与T-SNARE蛋白重组孔跨越膜,悬浮在多孔氮化硅基板,并检测使用激光共聚焦显微镜23个人的V型SUV的融合。其他˚F使用V型越野车与T-SNARE蛋白重组平面双层,支持在玻璃基板上15-17,21,22,24-27,29。使用支持的双层(SBLs)的巨大优点在于,TIRF显微镜可以用于检测具有优良信噪比并且在不脱离自由V形的SUV干扰对接和融合事件,虽然使用微流体使用还提供了单事件分辨率标准的远场荧光显微镜24。
一个主要问题是底物的相互作用双层是否以及如何影响支持双层质量和融合的过程。早期工作取得使用该直接支持的玻璃或石英衬底上15-17平面SBLs的。这些SBLs被吸附制成,爆裂,扩散和在衬底上叔SUV膜融合。它很快实现,然而,省略了密钥t-SNARE组分,SNAP25,从以这种方式制备SBLs导致V形的SUV对接和融合动力学indistinguishable使用完整的T-17 SNARE蛋白获得的。因为SNAP25 在体内 30,31绝对必需的融合,这些早期的尝试生理相关投入的问题。塔姆的研究小组通过使用更好的控制支持双层形成21战胜了这一挑战。它使用朗缪尔-布洛杰特沉积为SBL的无蛋白质的第一小叶,接着用叔-越野车21,其单层融合。这导致SNAP25依赖性融合。
以避免与直接支撑在玻璃基板上,而不需要使用朗缪尔-布洛杰特法的双层相关的潜在假象,Karatekin 等人介绍了柔软,水合聚(乙二醇)(PEG)的双层和基板24之间的缓冲。该变形也导致SNAP25依赖性融合24。垫着柔软的聚合物层上双层一直被称为更好地保存跨膜蛋白质的流动性和功能32,并与病毒融合33研究中使用。此外,聚乙二醇化双层似乎保留一定的能力进行自我修复,而且非常坚固34,35。首先,商购,脂质连接PEG链的一小部分被包括在叔SUV膜。当这些叔SUV的突发,并形成在玻璃基板上的平面双层,一个PEG刷覆盖平面双层的两个小叶。因为平面双层形成通过叔SUV的周围到所述亲水玻璃表面的PEG链的粘附驱动,脂质体破裂和平面双层形成相对不敏感,以所使用的脂质组合物。然而,当包括大量的胆固醇的,增加了越野车的粘结性时,越野车可以不自发破裂。如果是这种情况,可以采用渗透休克或二价离子,以帮助平面双层形成25。
如在本AP如上所述,proach的PEG刷覆盖所述平面的两侧,支持双层。面对微流体流动通道的刷有助于防止其也通常覆盖有聚乙二醇层传入v型越野车的非特异性粘附。 v型和叔SNARE复合体的形成从膜远侧N-末端并且前进朝向膜近端域36阶段开始。为V形的SUV与叔SBL中,v型和叔SNARE N-末端需要突出的PEG刷,这似乎是在测定条件下,上述情况下进行交互。刷高度可适于通过改变PEG化脂质的密度和PEG链长度37,38学习比斯耐尔其它蛋白质。所述PEG刷子覆盖定影双层的近侧表面中的另一个好处是,它们模仿它们填充有每平方微米39 30,000-40,000整合膜蛋白生物膜的拥挤的环境。就像在此测定的PEG链,所述REPU覆盖生物膜lsive蛋白层需要被推开以允许两个磷脂双层之间的接触为发生融合。
微流体流动通道在该测定中使用的,因为它们提供独特的优点。首先,微流体流使T-的SUV更均匀的沉积扩散和融合形成了T-SBL。其次,小声道音量(<1微升)减少样品消耗。第三,所需的小体积允许下恒定流量进行的整个实验。流动弱除去,大概非特异性,从SBL 16附着v型越野车。它还维护V型的SUV的T-SBL以上,简化动力学分析17的密度不变。最后,停靠囊泡容易从由流25进行自由的区分。第四,多个微流体通道可以在相同的盖玻片一起使用,每个探测不同的条件。这使得杜里条件比较纳克相同的实验运行。类似的方法已被用于面包车Oijen组研究流感病毒和缓冲SBLs 33之间的融合。
在TIRF显微镜,渐逝场的指数衰减(以衰减常数〜100nm)的界限荧光激发到那些在玻璃 - 缓冲界面的非常接近的分子。这最小化是更远离荧光分子的贡献,增加信噪比,并且允许用10-40毫秒的帧的曝光时间单分子灵敏度。渐逝场也导致在融合的信号增加:从SUV标记的脂质转移到SBL,他们发现自己,平均而言,在更强的激励场。这增加了荧光较大的脂质体更强。
如果偏振光是用来产生渐逝场,附加效果有助于对T中的荧光变化从SUV到SBL标签的转让(BOT)。某些脂质染料具有与相对于它们所嵌入的双层的优选平均角度定向的跃迁偶极。这将在通过荧光团时,他们在SUV 兑 SBL发射的荧光的量的差异,由于偏振光分束将在两个膜激发染料不同。对于前者,激发光束将与周围的球形的SUV取向过渡偶极子相互作用,而对于后者,偶极取向将由扁平SBL几何局限。例如,当S偏振入射的光(偏振光垂直于入射平面)时,激发是更有效率的,当染料在SBL比在SUV脂质染料跃迁偶极方向平行于膜29,40一(如直接投资收益还是没41-43)。掺杂了这样的荧光团SUV似乎暗淡时,码头上SBL( 图7,Representat香港专业教育学院结果)。作为融合孔打开,并连接在SUV和SBL膜,荧光探针扩散到SBL和变得更容易被s偏振光渐逝场25,27,29激发。因此,周围的融合位点整合的荧光信号从所述的SUV染料转移到SBL 27( 图3和图7)期间急剧增加。有助于信号伴随融合脱猝灭的荧光标记的时转移到SBL因为它们稀释变化的另外一个因素。相比,在这里描述的测定消逝场衰减和极化效应脱猝灭的贡献通常是次要的,因为只有一小部分(  )脂质的标记。
)脂质的标记。
在融合信号增加可以利用通过比较时间来推断融合孔性质,1“SRC =”/文件/ ftp_upload / 54349 / 54349eq2.jpg“/>,对于脂质通过的孔是自由可渗透脂质实际释放时间逃逸要求 。如果两个时间尺度是可比,将它的结论是,孔呈现给脂质流动阻力小。然而,如果实际的释放时间是显著比时间为扩散限制释放更长的时间,这将表明一个过程中,如孔隙抖动,延迟脂质释放。扩散限制释放时间,
。如果两个时间尺度是可比,将它的结论是,孔呈现给脂质流动阻力小。然而,如果实际的释放时间是显著比时间为扩散限制释放更长的时间,这将表明一个过程中,如孔隙抖动,延迟脂质释放。扩散限制释放时间,  取决于定影脂质体和脂质扩散的大小;其估计需要这两个参数来量化。测定的单分子灵敏度允许脂质扩散到其释放到SBL为每个聚变事件26之后追踪几个单脂的荧光团测定。每一熔断囊泡的大小可通过将(i)的单脂质染料的强度,(ii)在所有荧光团在融合转移到SBL中围绕一个停靠站点的总荧光变化,(iii)该已知标记的SUV的密度来估计27脂质,和(iv)每脂质的区域。对于许多融合事件中,发现了实际脂质释放时间要慢得多比由扩散控制的释放27预期,如先前指出的假定均匀的SUV大小44。假设脂质释放迟缓是由于毛孔闪烁,定量模型允许“毛孔开放”的估计,时间的部分孔隙依然融合27期间开放。
取决于定影脂质体和脂质扩散的大小;其估计需要这两个参数来量化。测定的单分子灵敏度允许脂质扩散到其释放到SBL为每个聚变事件26之后追踪几个单脂的荧光团测定。每一熔断囊泡的大小可通过将(i)的单脂质染料的强度,(ii)在所有荧光团在融合转移到SBL中围绕一个停靠站点的总荧光变化,(iii)该已知标记的SUV的密度来估计27脂质,和(iv)每脂质的区域。对于许多融合事件中,发现了实际脂质释放时间要慢得多比由扩散控制的释放27预期,如先前指出的假定均匀的SUV大小44。假设脂质释放迟缓是由于毛孔闪烁,定量模型允许“毛孔开放”的估计,时间的部分孔隙依然融合27期间开放。
每当实际,以测试同时使用脂质和可溶性内容标签融合的机制是很重要的。例如,脂质释放可以通过比孔隙闪烁等工艺,如由SNARE蛋白是环绕吨脂质扩散的限制被延迟他孔隙。如果是这样的话,然后松开的内容将先于脂质标签释放,所提供的孔径是大到足以允许可溶性探针通道。在方法的一个更基本的缺陷可能是通过一个狭窄的融合孔的SBL连接到囊泡已经基本上保留了其融合前形状发生标记的脂质转移到SBL的假设。脂质转移到SBL也可以从融合孔与随之而来的,超极本的SUV进入SBL膜的迅速崩溃的快速散瞳造成的,正如上面所提到仅仅基于29脂质发布的数据。同时监测血脂和内容同时发布,发现许多气孔释放他们所有的脂质标签后重新密封,但保留了他们的一些可溶性的货物27。这表明,至少一些脂质体不塌陷到SBL融合之后,并且该脂质染料转移到SBL通过融合孔发生。此外,LIPID和内容,同时发布27发生,使得它不可能脂质释放迟缓,是由于受到周围孔45 SNARE蛋白阻碍脂质扩散的。
未监测溶于内容发布一款SUV-SBL融合协议之前由Karatekin和罗斯曼25公布。在这里,最近的事态发展都包括在内,脂质即同步监测和内容发布和SUV,脂质,并融合孔特性27的推定。该协议开始于制备微流体细胞,通过结合的聚(二甲基硅氧烷)制成(PDMS)弹性体包含块槽用玻璃盖玻片25的指令。接着,制备V-SUV的与两个脂质和内容标记进行说明。第4和5用于组装微流体细胞, 原位形成的SBLs并检查缺陷和流动性,引入VS的提供指令UVS到流动细胞和融合事件的检测。部分6提供用于数据分析的指令。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1.准备一个PDMS块,形成微流体通道

图1. 微细加工液流电池模板和PDMS块准备。(A)可以套到24×60毫米玻璃盖玻片(底部)四通道流动池的设计。六个相同的设计安排,以适应到10cm的硅晶片(上图)。 (B)剪下约5-8毫米厚的PDMS块打孔器。 (C)插入管插入用一把镊子的冲孔。 请点击此处查看该图的放大版本。
- 获得的流动池模板如一个在图1A中。典型的渠道是0.3-1毫米宽,70-100微米高,长1-2厘米。制作一个模板我们在洁净室设备25或为了ING标准光刻技术,如果洁净室访问不可用。可替代地,机器与由适当的材料更大的尺寸的模板。
注:洁净室的工作人员可以培养和引导雏用户口罩,晶圆清洗和光刻的设计和排序(进入洁净室设备总是受限于用户适当的培训)。一旦获得一个模板,它可以在洁净室外面反复使用,只要它保持在一个封闭的培养皿,因此必须注意排除灰尘。 - 在一次性塑料杯中准备约100ml的PDMS 10毫升交联剂(固化剂)的混合物(有机硅弹性体基峰)和〜。折断的一次性吸管的尖端更容易地处理粘性PDMS。权衡PDMS到塑料杯作为移液是不准确的。
- 搅拌混合好。在真空干燥器中除去由脱气许多气泡(大约20分钟)。作为杯放置在真空下,气泡最初将生长在大小,大大增加了混合物的体积。控制所施加的真空,确保杯子是深足,以避免泄漏。
- 倒入大降脱气PDMS混合成玻璃培养皿英寸(150毫米×20毫米),按片上用模板朝上PDMS。这避免了被捕获在晶片下方的气泡。截留气泡会展开,并在盘被放置在烤箱可倾斜晶片。现在倒脱气的PDMS上的晶片的顶部的模板,直到它被约5-8毫米PMDS的覆盖。如果轻轻发生气泡用枪头将其删除。
- 烘烤在60℃下在烘箱中的PMDS 3小时。确保菜的水平。
- 使用新的手术刀刀片切出含有成型通道结构的PDMS块。切出块需要适合到一个盖玻片(见步骤4.1.9)。
- 从盘剥出的切出块和PL王牌它放到一块干净的铝箔。
注意:交联PMDS块可保持几个月。在单个晶片上,有6台流动通道( 图1A),所以6的PDMS块可以在一个单一的PDMS成型来制造。切割出PDMS块,清洁片PDMS的松散的所有6个,但在其他浇筑和交联的下一批,这将需要大约只有一半用于首批PDMS前不要取出剩余的PDMS后。一个好的模板可以持续数年。 - 使用打孔器在一个直线运动( 图1B)通过与PDMS块钻。开始对信道槽的一侧。一定要去除冲压件的PDMS。重复此为四通道设计的所有八个孔。
- 放置在PDMS块信道的一面朝下到一个新的,无皱一张铝箔。存储长达几个月块在干燥箱中,理想地在干燥器中。
- 推管(0.25毫米内径,0.76毫米外径)的三分之一到使用一对镊子( 图1C)的冲孔。切倾斜,方便插入油管。离开管子足够长的时间,达到了SUV水库和注射泵,分别为。组装芯片上放置在显微镜后,再次下调管,如果他们是太长。
- 连接到泵的注射器,切一小段较大硅胶管(0.51毫米内径,2.1 mm外径)的,并插入较薄管插入一侧。 PDMS的这种准备使用的块可以保持为几个月。
2.清洁盖玻片
- 根据使用硫酸(H 2 SO 4)和过氧化氢 的强氧化混合物中Karatekin和罗斯曼25中描述的协议清洁的盖玻片(H 2 O 2)。在洁净室执行该过程,并遵守适当的安全预防措施。另外,使用洁净室清洗çoverslip即市售(见材料清单)。
3. V型的SUV制备含有脂质和内容标签

图2 示意图SUV制剂。脂类混合在玻璃管(1),并蒸发溶剂通过在水浴中旋转所述管,以形成脂质膜(2)。的溶剂剩余痕量高真空(3)下除去。脂质膜水合在重建缓冲液含有洗涤剂和蛋白而涡旋(4)。如果内容染料是被封装,它包括在此步骤以及在稀释步骤(5)。其临界胶束浓度低于洗涤剂浓度的稀释导致脂质体的形成。洗涤剂过夜透析远(6)。对于叔越野车为SBL形成包括NBD-PE(绿色),囊泡漂浮在密度梯度,并在收集两个层(7A)之间的接口。为了与无染料样品运行在一个尺寸排阻柱和0.5ml的部分(7B)收集封装的内容标记分隔V型的SUV。 请点击此处查看该图的放大版本。
- 制备4升重建缓冲液中,含有25mM HEPES-KOH,140mM的氯化钾,为100μMEGTA和1mM DTT,pH 7.4的。最常使用的缓冲液中透析和100ml对其他步骤。
注意:也可使用其它缓冲剂,但为了保持缓冲摩尔渗透压浓度在整个实验过程保持一致,并选择其中的蛋白质是功能性的条件下,它是重要的。对于仅准备含脂质的标签V-SUV和T-越野车,跟着Karatekin和罗斯曼25。 - 脂质库存储存在-20℃下在氯仿或氯仿:甲醇(2:1,V / V),让小瓶开口之前达到室温避免冷凝。
- 冲洗的玻璃管,用氯仿中的氯仿和甲醇的混合物用1微摩尔的最终量以除去洗涤剂的痕量和混合脂质在所需的比率(2:1,V / V)。仅使用玻璃注射器/管处理有机溶剂/溶解脂肪。
注:此处使用的脂质是POPC:SAPE:DOPS:胆固醇:PEG2000-DOPE:是否在57.4的摩尔比:15:12:10:4.6:1为含V-SNARE囊泡和POPC:SAPE:DOPS:胆固醇:脑PI(4,5)P2:PEG2000-DOPE:NBD-PE(54.9:15:12:10:3:4.6:0.5)为T-SNARE的SUV。看材质的详细信息。其他脂质组合物都可以使用。 - 蒸发溶剂在氮气缓流,或在旋转蒸发器。沉浸管的前端在上述脂质的最高熔融温度下加热,得到均匀的脂质膜,并避免在温度大的变化,而蒸发溶剂的水浴(37℃)。在各地启动真空300毫巴,直到形成脂质膜,然后继续在旋转蒸发器中为〜2分钟,在最高可能的真空。
- 取出玻璃管,敷在铝箔以避免曝光和通过在高真空下放置管在干燥器中至少2小时除去溶剂残留痕迹。这一步也可以在一夜之间运行。
- 再水合所述干燥的脂质膜,制备洗涤剂(正辛基β-D-吡喃葡萄糖苷,OG)在重建缓冲液(500微升最终体积)混合物,蛋白质(V-SNARE蛋白),和SRB。保持最终洗涤剂浓度〜2倍的临界胶束浓度(CMC,20-25毫对OG),并调整最终体积,使得洗涤剂与脂质比为> 10。溶解磺酰罗丹明B(SRB)粉末中的蛋白质 - 洗涤剂溶液轻轻摇动该溶液以获得10-50毫SRB的终浓度。通过减少CO添加高浓度的硫酸盐还原菌的时候调整重建缓冲液的渗透压氯化钾ncentration相应。
注:T-SNARE蛋白脂质与蛋白质的比例(LP)高(LP〜20000微米〜25每平方70吨-SNARE蛋白)。显著在SBL更高的T-SNARE密度可能导致活性蛋白聚集体显著降低融合率17,24。 LP为V-SNARE蛋白低得多(LP〜200,每次〜2微米的7000 V-SNARE蛋白),接近上突触小泡46,47中发现的V-SNARE密度。以确保重组表达和V-和叔SNARE蛋白的纯化后的蛋白质的功能是有用的前通过单囊泡融合法的所有步骤的将要执行一个简单的批量融合测定48。 - 轻轻摇动含有干燥的脂质膜,同时加入来自步骤3.6的蛋白质 - 洗涤剂SRB溶液的预热的玻璃管。尽量避免产生气泡。继续在37℃摇晃15分钟。
- 稀释,加入重建五倍洗涤剂含缓冲SRB(加2毫升,2.5 ml终体积),同时迅速的涡流,以避免浓度梯度。继续在37℃15分钟,摇到1小时。
注:较长的潜伏期增加蛋白质的重建效率。迅速稀释而减小低于CMC的洗涤剂浓度,并导致小脂质体的形成。 - 第一透析囊泡悬浮针对〜1升重建缓冲液中,在环境温度1-2小时,然后对3升重建缓冲液中过夜,在4℃下与4克聚苯乙烯吸附剂,使用20000 MWCO透析管或盒。使用不同的烧杯与无SRB囊泡透析,防止交叉污染。
- 平衡与重建缓冲液的凝胶过滤柱。通过塔运行囊泡悬浮液以从V形的SUV包封SRB分离游离的SRB。使用重建缓冲液没有SRB作为洗脱液一旦整个样本有输入的列。收集0.5ml的分数v型的SUV。
- 之前和加入去污剂来泡16等分之后测量荧光SRB在验证自淬浓度成功SRB封装。使用荧光光谱仪,激发样品在550 nm和扫描570纳米和630纳米之间的SRB排放。作为膜是溶解和释放的SRB稀释期望在SRB荧光取决于在洗涤剂加入包封的量的4-8倍的增加。
- 表征使用荧光光谱24和SDS-PAGE凝胶电泳的脂类和蛋白质回收,分别。使用灵敏染色方法,尤其是对高LP叔SUV样品(见材料清单)。通常都脂类和蛋白质输入的约50%导致接近标称值的LP中的制备过程中被丢失。
- 表征使用动态光散射24或电子microsco尺寸SUVPY 48。在4℃下用含有V型越野车高达〜3-4天商店SRB。不冻结,因为冻融破坏细胞膜,释放封装SRB。
4. SUV-SBL融合化验,以监测血脂仅释放
- 微流体流动通道内的栓支撑双层的形成
- 放置在高真空下的PDMS块(步骤1.11),至少20分钟,在实验之前,以除去溶解的气体。这大大减少了在微流体通道的气泡的融合实验期间的风险。
- 打开显微镜安装和加热阶段和样本保持器( 图3和4),以所需的温度。
- 过滤用通过过滤器以0.45微米或更小的孔尺寸以稀释囊泡的重建缓冲液。
- 〜稀释的NBD-PE的30微升标记的T-SUV的或无蛋白(PF-越野车)控制脂质体(股票解决方案0.5-1毫米脂)与〜60微升缓冲。最终浓度这里不是关键的。
- 脱气使用3毫升注射器该混合物。按了大部分的空气样本的上方,而垂直持注射器。用石蜡膜密封(无针)尖端和创建拉下活塞的真空。点击针筒加速解决方案的脱气。直到施加真空时发生没有更多的气泡重复此过程数次。
- 用一皮下注射针,其直径比附着到PDMS块在微量离心管的盖冲一个孔油管稍大。确保塑料冲压出片不是在管内,因为它可能会堵塞连接到微流体通道入口以后的管路。
- 填充脱气的SUV溶液成具有在盖的孔的微量离心管中,并将其放置到显微镜舞台上的保持器平衡到设定的温度。
- PLACE在等离子体清洁器之前清洗盖玻片(第2部分),并运行空气等离子体约5分钟。把等离子体处理盖玻片(处理面朝上)在作为垫几无绒组织之上。
- 从脱气的PDMS块取下铝箔,并放置在该区块盖玻片顶部。重用当PDMS块装上和取下一块胶带到通道一边进行清洁。向下按PDMS块上用一把镊子,使其坚持盖玻片,但不要按太硬如玻璃可能打破。
- 放置装配流动池到显微镜阶段和管路连接到SUV贮存器和注射器泵,分别。大盘盖玻片到舞台( 图3B)。
- 开始在3微升/分钟抽吸越野车,直到溶液完全填充通道(〜2.25毫米/秒为75微米×300微米通道横截面)。当所有通道解决方案,开始动起来日在出口侧ë油管,减少流至0.5微升/分钟和孵育30-45分钟。
注意:等离子体处理载玻片和流动的越野车进入通道之间的时间应不超过10-20分钟的等离子体处理的效果是短暂的。 - 检查任何泄漏通道。使用10-20X空中目标从含叔或PF的SUV是NBD-PE观察NBD-PE荧光。激发荧光团NBD使用488纳米的激光。泄漏是很难用明视场照明检测。
- 把用脱气重建缓冲液的管子插入支架,让温度平衡的温度变化快可能导致SBL缺陷。停止流动,并等待约1分钟,以确保流动完全停止和无气泡将所述入口管切换到脱气之缓冲液前被吸入到管内。
- 冲洗脱气缓冲区中的所有渠道,洗去未结合的越野车。
- 切换到一个解释第1条- [R放大TIRF物镜(60X,油,NA 1.45-1.49),并验证双层看起来均匀,无任何明显的大规模的缺陷,如暗斑或从SBL延伸出来的小管脂质。
- 检查双层的流动性
- 如果一个专用的FRAP单元不可用,或者如果如下一个FRAP序列不能被编程,然后定性地测试膜的流动性。
- 关闭场光阑至小尺寸(〜直径40微米),并通过软件调整488nm的激发光的强度(20-80μW,或15-60纳瓦个/μm2)漂白的NBD-PE的荧光在曝光面积显著,但不完全。用于流体双层,在稳定状态下,在曝光区域的中部的荧光强度应比在边缘处较低,作为完整的NBD-PE分子进入曝光区和漂白前扩散一定的距离。相反,如果表面粘附小号UV的失败爆裂,或由于某些其他原因所支持的双层不是流体,在曝光区域中的所有荧光团应当漂白。
注意:在这个和后面的步骤的激光强度值给出一个粗略的起点,并应为一组给定的条件下进行优化。 - 停止照射和重新启动它,几分钟后,以验证稳态测量的结果
- 关闭场光阑至小尺寸(〜直径40微米),并通过软件调整488nm的激发光的强度(20-80μW,或15-60纳瓦个/μm2)漂白的NBD-PE的荧光在曝光面积显著,但不完全。用于流体双层,在稳定状态下,在曝光区域的中部的荧光强度应比在边缘处较低,作为完整的NBD-PE分子进入曝光区和漂白前扩散一定的距离。相反,如果表面粘附小号UV的失败爆裂,或由于某些其他原因所支持的双层不是流体,在曝光区域中的所有荧光团应当漂白。
- 程序更加定量测量FRAP序列,如果可能的话。见补充文件和细节相应的传奇。
注意:有时越野车将附着在玻璃盖玻片,但未能破裂,形成流体双层。如果发生这种情况,冲洗用含有10mM Mg 2+离子 ,以帮助支持双层形成脱气重建缓冲液的通道。使用荧光漂白来评估流动性双层为4.2。一旦流体双层形成,冲洗用Mg 2+ RECOnstitution缓冲区。
- 如果一个专用的FRAP单元不可用,或者如果如下一个FRAP序列不能被编程,然后定性地测试膜的流动性。
- 引入V形的SUV到微流体流动通道
- 脱气重建缓冲液,并用它来 通过约10 3倍的v型SUV原液稀释至10 5根据V形的SUV库存浓度。瞄准的稀释导致在约10-100融合事件中的视场60秒以内。
注意:太多的融合增加了背景荧光(因为每个事件的沉积LR-PE还是没脂质标签进入SBL),并融合事件的检测和分析困难。相反,融合率是统计差过低的结果或需要收购更多的电影。为0.1毫米的脂质的V形的SUV浓度,通过在995微升重建缓冲液稀释5微升的SUV库存开始,然后稀释5-50微升的此在950-995微升重建缓冲液。 - 让停止流动和inser前下平衡婷入口管插入稀释的V型SUV的解决方案。
- 调整TIRF角度和极化如下。
- 通过旋转激励光束设置所需偏振后,调整镜经由通过软件步进电机命令的光束的位置的倾斜。慢慢在后焦面移离目标的中心的激光束的位置向一侧偏心。从在前侧出现具有增加角相对于所述目标轴作为位置进一步移动偏离中心的目标观察的光。
- 注意电机位置时射出的光束第一次消失在目标, 即在首次实现TIR。
- 慢慢地保留进一步移动光束位置偏离中心,而从表面监测荧光。注意电机位置时,当梁太过分偏离中心的移动表面的荧光消失。
- 选择梁positio n中的两个极限之间上面确定。用于在融合最佳信噪比和多个信号增强,选择一个浅的穿透深度(光束位置更靠近物镜的边缘),该仍然提供了视场的均匀照明。
注意:最好是保持同样的TIR光束位置(同穿透深度),所有实验的设置进行了优化之后。确保旋转偏振不会导致在光束位置的变化。
- 流V-SUV的进入通道在2微升/分钟的流速,对应至1.5毫米/秒〜为75微米×300微米的横截面的平均线性流速。切换到激发/发射设置监控脂质混合(LR或仅所做的那样)。
- 脱气重建缓冲液,并用它来 通过约10 3倍的v型SUV原液稀释至10 5根据V形的SUV库存浓度。瞄准的稀释导致在约10-100融合事件中的视场60秒以内。
- 单V型越野车和SBL之间的融合观察
.JPG“/>
图 3. 实验pTIRF设置。(A)的示意图在玻璃基板上的V形的SUV和叔SBL的表示。单的SUV-SBL融合事件的假色TIRFM图像表示脂质对接(1)和脂质染料释放到SBL(2),随后的漂白和荧光强度的减少(3)。总荧光强度信号(在5.3微米×5.3微米的框的像素值的总和)被示出。 (B)中键合到PDMS块的盖玻片贴到加热阶段。对于微流体通道入口管由管金属样品架(右)通过注射泵(左)来吸取借鉴的样本。下方的泵是双发射单元。 请点击此处查看该图的放大版本。
OAD / 54349 / 54349fig4.jpg“/>
图4. 实验装置的示意图。在微流体通道中的玻璃-缓冲界面创建的渐逝波。的SBL形成在玻璃和v-SUV被从金属样品架(右上)通过通道进入注射器(左上)吸出。男,镜; DM,分色镜; L,镜头;楼滤波器; P,偏振器。 请点击此处查看该图的放大版本。
- 不断激发并根据V型的SUV纳入了荧光显示器采用561纳米(对于LR-PE)激光V-SUV荧光或638纳米(为DID)。为V的SUV与SBL到达流道和码头上和保险丝,背景荧光信号开始积累。
- 通过软件调整的激励激光强度连续地漂白背景荧光使得在稳定状态下新的对接和融合事件可以很容易地观察到。
注意:如果漂白太慢,背景荧光会太高。如果漂白太快,从停靠SUV和从释放到SBL荧光信号将迅速褪色,缩短其间停靠的SUV融合可以检测或荧光团可被跟踪的窗口。对于LR-PE在561 nm光激发,2.5-7.5毫瓦功率照射190微米直径的圆(100-250净重/微米2)是一个合理的值来启动。为638纳米的激发做了,在一个190微米直径的圆(30-60纳瓦个/μm2)0.8-1.6毫瓦的功率可用于初始测试。 - 获得在不同的位置几部电影在给定的信道。检查NBD-PE的荧光,以验证SBL没有在这些位置的缺陷。获得以最大速率(〜50帧/秒)或感兴趣的更高帧速率的裁剪区域(高达全帧的电影〜100赫兹)60秒。
- 移动到另一个微流体通道并重复录音的其他条件。包括诸如无蛋白质SBL或越野车,添加V-SNARE VAMP2(CDV)的可溶性胞质域作为抑制剂或在一个或多个治疗v型SUV与破伤风神经毒素(在37℃下30分钟)的阴性对照对同一盖玻片的通道。
- 清理和回收PDMS
- 为了重复使用的PDMS块,以5微升/分钟的流速冲洗以〜200微升70%乙醇的微流体通道。最后,通过管道和通道抽吸空气。
- 拆下PDMS从盖玻片稍微挤压它轻轻地阻止。将其放在一块干净的铝箔。储存在真空干燥器中。它可以重复使用多次。
- 要更彻底的清洁漂洗之前用70%的乙醇的洗涤剂或氢氧化钠溶液的通道。另外,删除所有管来回 m为PDMS和超声处理块在异丙醇30分钟,干燥,插入新的管道之前。
5. SUV-SBL融合化验,以监测血脂和内容同时发布
- 对于同时脂质和可溶性含量释放,遵循相同的步骤,在第4节,除标有两个脂质(DID)使用脂质体和可溶性含量(SRB)标签,在部分解释了双色监控3. SRB封装以10mM和最初高度自淬灭。
- 使用图4中示意性示出的结构中,激发的SRB并没有荧光同时使用561纳米和638纳米的激光,分别。分色镜(640纳米)分割排放到穿过短的(五十零分之五百九十五纳米)运行两个横梁和长(七十五分之七百nm)的波长滤波器检测SRB并没有排放量,分别为。两个发射束被投射侧由端上的EM-CCD芯片。
- FRAP数据分析
- 使用估计脂质扩散系数,D的补充资料中提供的MATLAB程序。程序读取的OME-TIFF文件49的列表,检测出漂白区,在漂白区作为时间的函数绘制的平均像素值,以及由Soumpasis 50适合的产生的恢复曲线模型以提取恢复时间,
 ,其中w是漂白圆的半径。
,其中w是漂白圆的半径。
注:MATLAB程序是为使用尼康ND2文件FRAP电影前面的分析提供了25。目前的程序读取OME TIFF文件49,因为大多数文件格式,可以很容易地转换为OME-TIFF。 FRAP数据的定量分析是最容易漂在最准确的是瞬间的bleacHED区域是一个圆,和漂白的读出过程中可以忽略不计。虽然这些标准还没有此处所描述的简单的FRAP测量严格满足,可以得到扩散系数的一个合理的估计。为了更准确的估计,使用单脂质染料(6.3节)的跟踪。
- 使用估计脂质扩散系数,D的补充资料中提供的MATLAB程序。程序读取的OME-TIFF文件49的列表,检测出漂白区,在漂白区作为时间的函数绘制的平均像素值,以及由Soumpasis 50适合的产生的恢复曲线模型以提取恢复时间,
- 对接率,融合率和对接至融合延迟时间的分析
- 使用ImageJ的开一部电影来分析和调整亮度和对比度,以清楚地识别囊泡对接和融合活动。启动SpeckleTrackerJ 26插件。见SpeckleTrackerJ Smith等 26和在线文档为使用说明。
- 识别SpeckleTrackerJ所有新的SUV停靠。为了区分,从那些弹开SBL紧密对接越野车,并处以几帧的最小时间对接。保存曲目,重复所有电影。
注:对于DOC王率,所有重要的是,以确定何时一个SUV停靠,所以只在该越野车停靠需要被记录在这些磁迹的第一帧。磁迹的剩余部分将不被考虑在分析中。但是,自动跟踪每个SUV,直到漂白或保险丝帮助那些已经被跟踪标志的SUV。 - 确定所有融合囊泡。对于这些,轨道应包括来自所述第一帧的SUV停靠直到其中融合通过在跟踪点的荧光的突然增加是明显的第一帧的所有帧。这些磁迹的持续时间被用于计算对接至融合延迟。保存所有曲目。重复所有电影。
- 对接或融合数据批量分析,编译从相关电影的轨迹文件的列表,并运行提供Karatekin和罗斯曼25 MATLAB程序。按照与该节目一起提供的说明。
注:程序绘制暨ulative对接和融合事件作为时间的函数的基础上,从轨迹文件中提取的信息。对接和融合率从这些图的斜率估计。融合数据,对接至融合延迟还计算及其分布绘制为生存曲线图, 即 ,还没有被由对接之后的给定延迟发生融合的概率。
- 脂质扩散
- 对于每个聚变事件,跟踪单个荧光脂质,因为它们变得可辨时,他们已扩散从融合位点( 图6)足够远。使用SpeckleTrackerJ跟踪,并保存利用MATLAB进一步分析的曲目。根据定影囊泡的大小,典型地3-30单荧光团可被跟踪。因为较长的轨道是期望的计算
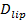 ,设法跟踪单个分子只要如可能的话,必要时可使用轨迹的手动校正。
,设法跟踪单个分子只要如可能的话,必要时可使用轨迹的手动校正。 - 计算均方为持续> 40-50帧(约1.5〜秒)单脂标记的轨迹位移(MSD)。使用MSD来计算脂质扩散系数, 。参见Smith 等人的 26和Stratton 等27的详细信息。
- 对于每个聚变事件,跟踪单个荧光脂质,因为它们变得可辨时,他们已扩散从融合位点( 图6)足够远。使用SpeckleTrackerJ跟踪,并保存利用MATLAB进一步分析的曲目。根据定影囊泡的大小,典型地3-30单荧光团可被跟踪。因为较长的轨道是期望的计算
- 单脂质染料力度,SUV-SBL强度降低的因素,囊泡大小
- 之前和在一个单一的步骤中的标记物的漂白剂后测量脂质标记的像素值的总和在3×3像素左右的标志物(0.8微米×0.8微米)区域〜15帧。减去背景强度平均从预漂白强度漂白后帧漂白以获得跟踪脂质标记物的强度之前平均。重复测量尽可能多的标记实用为给定的电影。
- 绘制单个脂质标签强度的分布,
 和适合估计平均高斯。
和适合估计平均高斯。 - 计算当融合发生时单一的脂质标志物在该事件发布的轨迹在单步的漂白结束之间的延迟。得到的漂白时间在SBL荧光团,
 ,通过绘制延迟的幸存者功能,拟合指数。
,通过绘制延迟的幸存者功能,拟合指数。
注意:由于偏光激发场夫妇更弱到在SUV的荧光团,一个SUV的漂白时间通常要慢得多27。 - 估计
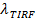 中,强度降低系数为脂质染料在SUV相对时,它是在SBL,以下力通27时(tp_upload / 54349 / 54349eq8.jpg“/> 在SUV单一染料)的强度。
中,强度降低系数为脂质染料在SUV相对时,它是在SBL,以下力通27时(tp_upload / 54349 / 54349eq8.jpg“/> 在SUV单一染料)的强度。
注意:如果漂白是可以忽略不计,将等于插接强度 (点(1)在图3A中 ,右面板),由所有荧光团后达到的总强度除以可应融合存入SBL(虚线标示
(点(1)在图3A中 ,右面板),由所有荧光团后达到的总强度除以可应融合存入SBL(虚线标示 在图3中)。然而,由于在SBL快速脱色, 通常没有达到和的精确估计需要的释放动力学拟合表达式27,其包括双方的UDE (参见6.4.3)和 。斯特拉顿等人提供了单独的表达式27孔限制和扩散限制释放动力学的案件。最佳拟合的情况下,从事件变化到事件(见6.5.1孔隙限定动力学的情况下)。
在图3中)。然而,由于在SBL快速脱色, 通常没有达到和的精确估计需要的释放动力学拟合表达式27,其包括双方的UDE (参见6.4.3)和 。斯特拉顿等人提供了单独的表达式27孔限制和扩散限制释放动力学的案件。最佳拟合的情况下,从事件变化到事件(见6.5.1孔隙限定动力学的情况下)。 - 计算泡区从27个事件
 ,其中是SUV对接力度, 是指单一脂质染料的强度在SBL, 为脂质染料的强度降低的因素,当是在SUV相对中,当它是在SBL,及通货膨胀1“SRC =”/文件/ ftp_upload / 54349 / 54349eq12.jpg“/>是脂质染料的已知面密度。
,其中是SUV对接力度, 是指单一脂质染料的强度在SBL, 为脂质染料的强度降低的因素,当是在SUV相对中,当它是在SBL,及通货膨胀1“SRC =”/文件/ ftp_upload / 54349 / 54349eq12.jpg“/>是脂质染料的已知面密度。
- 融合孔性能
- 假设释放孔的限制,适合血脂标签释放动力学至27
 ,其中
,其中 是停靠的SUV强度只是融合前和其它参数如前面所定义。利用价值 6.4.3获得固定的参数,并提取最合适的估计和
是停靠的SUV强度只是融合前和其它参数如前面所定义。利用价值 6.4.3获得固定的参数,并提取最合适的估计和 。
。 - 估计的时间部分的孔是开放的,P 0,脂质释放的假设相位差是由于孔隙闪烁27:60;
 =
=  ,其中A VES是囊泡区域(部分6.4.5),b为孔高度(通常取为〜15纳米),; R P≈3纳米的由扩散脂质标签所见,并且包括半有效孔隙半径双层厚度(〜2纳米), 是(以6.3计算)脂质扩散,并为从SUV被释放脂质进入SBL(从6.5.1)的时间。
,其中A VES是囊泡区域(部分6.4.5),b为孔高度(通常取为〜15纳米),; R P≈3纳米的由扩散脂质标签所见,并且包括半有效孔隙半径双层厚度(〜2纳米), 是(以6.3计算)脂质扩散,并为从SUV被释放脂质进入SBL(从6.5.1)的时间。 - 为了证实标称P 0> 1表示完全打开毛孔,P 0 = 1,适合强度的时间过程的方程斯特拉顿等人 27,预测动力学4永久开孔。这种配合应大于fittin更好克永久开口气孔在6.5.1中的表达。
- 假设释放孔的限制,适合血脂标签释放动力学至27
 ,其中w是漂白圆的半径。
,其中w是漂白圆的半径。  ,通过绘制延迟的幸存者功能,拟合指数。
,通过绘制延迟的幸存者功能,拟合指数。  (点(1)在图3A中 ,右面板),由所有荧光团后达到的总强度除以可应融合存入SBL(虚线标示
(点(1)在图3A中 ,右面板),由所有荧光团后达到的总强度除以可应融合存入SBL(虚线标示 在图3中)。然而,由于在SBL快速脱色,
在图3中)。然而,由于在SBL快速脱色,  ,其中
,其中 ,其中
,其中 是停靠的SUV强度只是融合前和其它参数如前面所定义。利用价值
是停靠的SUV强度只是融合前和其它参数如前面所定义。利用价值 =
=  ,其中A VES是囊泡区域(部分6.4.5),b为孔高度(通常取为〜15纳米),; R P≈3纳米的由扩散脂质标签所见,并且包括半有效孔隙半径双层厚度(〜2纳米),
,其中A VES是囊泡区域(部分6.4.5),b为孔高度(通常取为〜15纳米),; R P≈3纳米的由扩散脂质标签所见,并且包括半有效孔隙半径双层厚度(〜2纳米), Subscription Required. Please recommend JoVE to your librarian.
Representative Results
SBL质量
这是至关重要的事先验证SBL的质量和流动性,以融合实验。在一个微流体通道的底部,玻璃侧的荧光应均匀,而没有任何明显的缺陷。如果气泡虽然经过通道,它通常留下的SBL明显的疤痕。如果有如此大规模的疤痕/缺陷,不要使用该通道。有时越野车可能附着到衬底上,但未能突发和形成连续SBL。如果是这样的情况下,该荧光仍可能出现很均匀,但漂白小区域之后将不恢复。引入10毫镁离子通常有助于解决这一问题。可以引入其它试剂,帮助爆裂的越野车,如其它二价阳离子51,聚合物溶液52,或渗透压休克25。
温度的变化导致SBL区域的变化,因为由一个脂质所占的平均面积依赖于温度。由5-10℃以上提高温度或减小它会导致多余的区域进入从表面,或分别为53出现在表面上,避光,SBL-自由斑点挤出管。重要的是要保持的温度变化到最小是很重要的。
图5为SBL这是相对无缺陷的和流体。

图5. FRAP检查SBL流动性。通过展示前(第一帧)闭场光阑(直径49微米),并具有较强的光脉冲漂白后分隔照明区域的图像序列。该地区在低曝光成像,以遵循NBD扩散荧光标记恢复血脂。该图示出了归一化的荧光作为时间从不同盖玻片指示方法的再现性的高级别多个通道的功能。在T-SBL的脂质组成POPC:SAPE:DOPS:胆固醇:脑PI(4,5)P2:PEG2000-DOPE:NBD-PE(54.9:15:12:10:3:4.6:0.5%(摩尔) )和LP = 20,000。漂白面积为984.203μ米3。恢复时间和一个合适的(见6.1)计算的扩散系数分别为τ= 67.3秒,D = 1.99 平方微米/秒,分别为。 请点击此处查看该图的放大版本。
使用脂质标签对接与融合事件的单色检测
的对接和融合事件人工计数是一个繁琐的任务,都需要良好的统计每个条件100-400事件。由于越野车可以熔合之前就SBL移动,需要一个跟踪软件。 SpeckleTrackerJ 26插件ImageJ的拥有专门用于跟踪的SUV的对接和融合事件分析开发的功能。往往只停靠囊泡的一小部分最终融合;计算对接速率有更多的越野车需要比那些熔化或需要分析,直到一个可靠的估计被检测到的事件的足够数量的唯一的电影的部分进行跟踪。如果主要焦点是融合率和/或对接至融合延迟,只需要被识别和跟踪定影越野车。
确保SBL的质量后,越野车被引入到通道中,并使用S偏振光的荧光的TIR下连续激发。对于最初的实验中,TIR激发光束的入射角可能需要进行调整,以调整所产生的倏逝波的穿透深度。代表单一SUV的对接和融合过程如图6A。在帧期间停靠的囊泡标记D和融合帧F.约1.3秒后(帧S)开始,因为它们扩散从融合远坐单脂类成为可辨即在每个框中的总荧光强度(像素值的总和)被绘制为时间在图6B中的函数,以在该对接和融合发生所指示的帧。作为时间的函数在图6C中被绘制,对于使用叔SNARE五个不同的电影融合事件的累积数重构SBLs(叔SBL,LP = 20,000,每平方微米约70叔斯耐尔)。从每部电影的聚变速率由一个线性拟合的斜率计算。这个平均聚变速率±SEM被示为在面板D.条形图对接至融合延迟为在图6A,B所示的例子中为0.07秒。类似地,共158事件计算延迟的分布示于图6D作为幸存者情节, 即,一个对接囊泡尚未由给定延迟稠合的概率。在100毫秒内大约1/2这些囊泡融合的。随着胆固醇水平,所述DISTRIB对接到融合延迟ution缩短,即使脂质扩散减慢27。
并行运行控制实验是重要的。来自这在图6B中 ,D中的数据获得的相同盖玻片,控制条件在相邻信道进行测试。在一个通道中,SBL不包括任何T-SNARE蛋白。出5电影持久每60秒,只有7融合事件进行比较,以158记录时的t斯耐尔被列入( 图6C)。在另一个通道,V形SNARE VAMP2的(CDV)的可溶性胞质域被包括在内。 CDV结合在SBL的叔斯耐尔,防止全长VAMP2是在越野车的结合。在五个60秒的电影,在CDV( 图6C)的存在下没有检测到融合事件。

图6。 SUV-SBL对接和融合事件。(A)与T-SBL由pTIRF显微镜成像以下LR-PE的传播V型SUV融合活动图像序列。图片采取每18毫秒,只显示每隔一帧。 D和F标记的对接和融合的发作,分别。因为它们从融合位点和彼此的(S)扩散远离个人脂质标签成为识别。框架尺寸:16.7微米×16.7微米。 (B)的荧光强度作为时间(A)中示出的融合事件的功能。 (C)的融合事件的累计数作为时间标记与来自另一组的实验并v型越野车的功能。 (D)的归一化的聚变速率(平均值±SEM,SX微米2×PM SUV)-1,假定为2×10 4血脂每SUV和(E)的囊泡对接后的给定时间的生存概率。所有SB组成LS是POPC:SAPE:DOPS:胆固醇:脑PI(4,5)P2:PEG2000-DOPE:NBD-PE(54.9:15:12:10:3:4.6:0.5摩尔%)和LP = 20,000。 V-SUV的人组成的POPC:SAPE:DOPS:胆固醇:PEG2000-DOPE:LR-PE(57.4:15:12:10:4.6:1摩尔%)和LP = 200 A和B.对于CE,在V -SUV组成相似,只是没被使用LR-PE代替。 请点击此处查看该图的放大版本。
脂质扩散,囊泡大小和融合孔属性
测定的单分子灵敏度允许除了对接和融合率和对接至融合延迟分布的几个参数的提取。单,荧光标记的脂质变得明显,因为他们从融合部位( 图6A)弥漫开去。跟踪ŧHESE直接提供脂质扩散26, 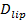 和漂白率在SBL 27荧光团的一个数字测量。在融合时转移到SBL标签(ⅰ)脱猝灭,由于它们的稀释:如在引言中提到,在SUV标签的荧光发射差异与在SBL,由于三个因素是在一般难以解开,(ⅱ)由于由TIR产生的倏逝场,和(iii)的荧光团过渡偶极子的取向相对于所述激发光束的偏振衰减较高的激发强度的SBL。因子(ⅱ)的大小取决于消逝场衰减长度和囊泡尺寸的相对尺寸和为每个聚变事件不同,由于在SUV尺寸的分散度。因子(ⅲ)取决于荧光团和使用的极化。而不是试图迪通过在周围融合前和所有荧光团后的SUV的区域进行比较的总的像素值直接sentangle这些效果,可以测量荧光变化的幅度已经沉积到SBL以下融合( 图3A中,(1)与( 2))。不像以前的工作29,44,该区域应选择足够大,使得没有荧光团将已通过融合后考虑的时间扩散离开它。融合后选择用于分析的30×30像素区域(8微米×8微米)到1.6秒通常是足够27。忽略漂白的时刻,比例
和漂白率在SBL 27荧光团的一个数字测量。在融合时转移到SBL标签(ⅰ)脱猝灭,由于它们的稀释:如在引言中提到,在SUV标签的荧光发射差异与在SBL,由于三个因素是在一般难以解开,(ⅱ)由于由TIR产生的倏逝场,和(iii)的荧光团过渡偶极子的取向相对于所述激发光束的偏振衰减较高的激发强度的SBL。因子(ⅱ)的大小取决于消逝场衰减长度和囊泡尺寸的相对尺寸和为每个聚变事件不同,由于在SUV尺寸的分散度。因子(ⅲ)取决于荧光团和使用的极化。而不是试图迪通过在周围融合前和所有荧光团后的SUV的区域进行比较的总的像素值直接sentangle这些效果,可以测量荧光变化的幅度已经沉积到SBL以下融合( 图3A中,(1)与( 2))。不像以前的工作29,44,该区域应选择足够大,使得没有荧光团将已通过融合后考虑的时间扩散离开它。融合后选择用于分析的30×30像素区域(8微米×8微米)到1.6秒通常是足够27。忽略漂白的时刻,比例 之前和之后的融合提供的荧光强度减少的因素,
之前和之后的融合提供的荧光强度减少的因素, 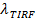 。囊泡大小可以定在SBL单个标签的荧光强度(估计尔斯/ ftp_upload / 54349 / 54349eq6.jpg“/>),该系数由它的强度会,如果它是在SUV(减少 ),在SUV标签的已知密度,并通过脂质所占的面积。在实践中,漂白通常是足够快的所有标记存入SBL的时候,有的已经漂白。出于这个原因,对于准确的估计,荧光强度降低的因素, 和脂质的释放时间,
。囊泡大小可以定在SBL单个标签的荧光强度(估计尔斯/ ftp_upload / 54349 / 54349eq6.jpg“/>),该系数由它的强度会,如果它是在SUV(减少 ),在SUV标签的已知密度,并通过脂质所占的面积。在实践中,漂白通常是足够快的所有标记存入SBL的时候,有的已经漂白。出于这个原因,对于准确的估计,荧光强度降低的因素, 和脂质的释放时间,  ,最好从一适合,考虑到漂白27的总的像素值的时间过程萃取。在SBL标签的漂白率独立地从单一的脂质染料跟踪估计,或者通过拟合指数为总强度分布,
,最好从一适合,考虑到漂白27的总的像素值的时间过程萃取。在SBL标签的漂白率独立地从单一的脂质染料跟踪估计,或者通过拟合指数为总强度分布,  ,在长时间( 例如,制度(3)在图3中)。知道定影囊泡的大小和脂质扩散允许的实际脂质释放时间比较, ,围绕囊泡的脂质的扩散时间
,在长时间( 例如,制度(3)在图3中)。知道定影囊泡的大小和脂质扩散允许的实际脂质释放时间比较, ,围绕囊泡的脂质的扩散时间 ,其中A VES是泡区, 是脂质扩散。如果两个时间尺度是可比,孔呈现给脂质释放小的阻力,并且释放是扩散限制。相反,如果
,其中A VES是泡区, 是脂质扩散。如果两个时间尺度是可比,孔呈现给脂质释放小的阻力,并且释放是扩散限制。相反,如果 ,则孔隙显著阻碍脂质通量。相位差的定量量度是“孔开放”,P 0,等于比率
,则孔隙显著阻碍脂质通量。相位差的定量量度是“孔开放”,P 0,等于比率 次Ø因素为了˚F团结与孔隙几何形状27。对于双状态,开/闭孔,P 0是时间在打开状态下的级分。对于一个闪烁孔,其大小连续变化的,P 0表示半径相对于全开半径的时间平均。
次Ø因素为了˚F团结与孔隙几何形状27。对于双状态,开/闭孔,P 0是时间在打开状态下的级分。对于一个闪烁孔,其大小连续变化的,P 0表示半径相对于全开半径的时间平均。
的信噪比必须跟踪单个脂质荧光团足够好的。这是更容易在单色实验取得,使用明亮的标记,如LR-PE。
脂质的同时检测和可溶性货物放行
代表性事件示于图7。在高浓度的用于封装的SRB荧光最初自淬灭。因此,大多数停靠V型的SUV只能由他们了DiD信号27检测。剧增中那样荧光标记的脂质混合,如单色脂质标签实验。脂质释放开始恰逢增加SRB荧光的发作。这一增长很可能是由于SRB脱猝灭的SRB分子逃脱了SUV,其余SRB是低于自猝灭浓度稀释到。有趣的是,SRB荧光达到稳定的平台。如果融合孔保持开放,人们会期望的SRB信号增加到由于脱猝灭最大值,随后下降到背景作为剩余SRB离开了SUV,或者如果在SUV折叠成SBL。稳定的高原最可能表示释放所有脂质的标签和可溶性货物的一小部分后重新密封所述孔。
在某些情况下,脂质释放信号并没有伴随着任何可检测的内容释放信号。这可以是因为孔隙太小,以允许SRB的通道,或者一些如何SUV已经失去了它的可溶性的货物。这些区分两种可能性,越野车停靠漂白进行分析。脂质标签的漂白了DiD降低秒内其荧光信号,而在增加了SRB信号作为SRB的一小部分的最初自猝灭SRB结果漂白是光转换为其它的产品和自猝灭被解除。虽然大多数对接的SUV(48选自58分析事件)超过照度长时间显示在SRB荧光的预期增加,一小部分(58分之10)根本不显示任何SRB信号。因此,越野车的一小部分可以在其制备和用途在实验之间的期间失去其可溶物含量。
在80%以上的事件(74总分91),用于其中两个SRB并没有信号可遵循的SRB荧光增加并保持在一个稳定的平台27上 。在T〜20%HESE情况下,剩下的SRB荧光突然释放可达几秒钟后。这可能不是由于孔隙允许1-2帧(18-36毫秒)或SUV的破灭内快速,充分释放的重新开放,由于累积的光损伤16定。不论其来源,第二,SRB的快速释放排除了一种可能性,即初始发行后的剩余SRB信号可能是由于在SBL和充分释放后的玻璃基板之间的空间的SRB剩余的夹带。这样的机制被建议在早期研究,其中SBL于玻璃直接支撑,与两个16之间的小空间。与此相反,在该试验中所使用的聚乙二醇化的脂质应该提供的SBL和玻璃25之间的空间4-5纳米,使SRB漫射从融合位点远。

图7. SIMULTAN eous脂质和内容发布。从脂质(蓝色)和内容(红色)标记为SUV-SBL融合事件的荧光信号(详见正文)。来自每个信道的快照在底部(4.0微米宽)被示出。卡通(中)示出了不同的阶段。 (1)SUV码头,做过荧光低。 (2)Fusion允许转移了DiD进入SBL并没有荧光增加。自我淬火由SRB逃逸通过孔缓解SRB信号也随之增加。 (3)SRB信号保留在SBL内扩散稳定而了DiD分子表示孔隙的一个可能的再密封。 (4)SRB信号下降非常迅速,由于快速全面融合或脂质体破裂。 V型越野车的组成为POPC:SAPE:DOPS:PEG2000-DOPE:是否(67:15:12:5:1(摩尔)%),LP 200,而叔SBLs分别组成POPC的:SAPE:DOPS:脑PI (4,5)P2:PEG2000-DOPE:NBD-PE(64.5:15:12:3:5:0.5摩尔%)和LP = 20,000。 =“_空白”>点击此处查看该图的放大版本。
FRAP_instructions.pdf -说明建立FRAP序列请点击这里下载此文件。
ReadMe_FRAP.txt -文本文件的描述来分析使用所提供的MATLAB代码FRAP恢复数据的过程,请点击这里下载该文件。
EK_FRAPbatch.m - MATLAB文件批量分析荧光漂白后恢复。此文件调用子程序MATLAB和EK_getFRAPvals.m EK_SoumpassisFit.m。“_blank”>请点击这里下载该文件。
EK_getFRAPvals.m - MATLAB文件,子例程EK_FRAPbatch.m叫请点击这里下载此文件。
EK_SoumpassisFit.m - MATLAB文件,子例程EK_FRAPbatch.m叫请点击这里下载此文件。
JN150923c1_FRAP_list.txt -文件名 是批量分析了主要的MATLAB脚本EK_FRAPbatch.m列表的一个例子,请点击此处下载该文件。
该下面的文件是TIF栈以及可以通过运行EK_FRAPbatch.m并提示选择一个文件列表时选择JN150923c1_FRAP_list.txt文件进行分析FRAP序列的五个例子相应的文本文件。
JN150923c1_F1_MMStack_Pos0.ome.tif - FRAP图像堆栈请点击这里下载此文件。
JN150923c1_F1_MMStack_Pos0_metadata.txt -对应的文本文件的元数据,请点击这里下载该文件。
JN150923c1_F2_MMStack_Pos0.ome.tif - FRAP图像堆栈PL轻松点击即可下载该文件。
JN150923c1_F2_MMStack_Pos0_metadata.txt -对应的文本文件的元数据,请点击这里下载该文件。
JN150923c1_F3_MMStack_Pos0.ome.tif - FRAP图像堆栈请点击这里下载此文件。
JN150923c1_F3_MMStack_Pos0_metadata.txt -对应的文本文件的元数据,请点击这里下载该文件。
JN150923c1_F4_MMStack_Pos0.ome.tif - ˚FRAP图像堆栈。 请点击此处下载此文件。
JN150923c1_F4_MMStack_Pos0_metadata.txt -对应的文本文件的元数据,请点击这里下载该文件。
JN150923c1_F5_MMStack_Pos0.ome.tif - FRAP图像堆栈请点击这里下载此文件。
JN150923c1_F5_MMStack_Pos0_metadata.txt -对应的文本文件的元数据,请点击这里下载该文件。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
成功地实施这里所描述的SUV-SBL融合法的关键取决于几个关键步骤,例如蛋白质的功能性重组到脂质体中,获得良好的质量SBLs,并选择合适的摄像参数,以检测单个分子。虽然这可能需要一些时间和努力才能成功,一旦试验成功实施,它提供了无法从简介中讨论的任何其他体外实验融合了丰富的有关融合过程的信息。率在该越野车停靠到和保险丝与SBL,对接至融合时间的分布,脂类扩散率,每熔断脂质体的大小和脂质释放对于给定的融合事件是否是弥散或孔控制的罐来确定。如果释放慢得多比从扩散预期,假定孔闪烁模型underlies脂质释放产量的相位差,P 0,时间的分数的孔隙我聚变27期间开放。
以前的工作探测单一的SUV,SUV融合18-20,28或脂质体和病毒颗粒的融合15-17,21,22,33,44染料分子之间依靠FRET和自我淬火SBLs。相比之下,我们在很大程度上避免染料脱猝灭探测脂质标签转移动力学入SBL,有两个原因。首先,FRET效率是脂质释放的一个高度非线性的记者。小的变化的染料间距离d导致在FRET效率大的变化,但只有一个有限的范围D / R 0的值,其中,R 0是福斯特距离。即,实际的染料稀释,并在脱猝灭信号的观察到的增加,可能会出现,由于下降到接近至R 0值所需的时间之间的延迟时间。同样,在脱猝灭信号变化不大,一旦染料密度充分降低后的脂质释放,使得 1.4,即使染料显著量仍停留在囊泡。这些特性使得脱猝灭的好工具,以获得秒杀的信号,以检测融合事件,但使用高时间分辨率的脂质释放定量动力学分析复杂化。与此相反,在强度由于偶极再取向和渐逝场的效果从SUV到SBL荧光团转移的变化线性相关染料在SUV仍然保留的级分。第二,对于自猝灭需要高浓度的类脂的标签可能影响融合过程本身。
1.4,即使染料显著量仍停留在囊泡。这些特性使得脱猝灭的好工具,以获得秒杀的信号,以检测融合事件,但使用高时间分辨率的脂质释放定量动力学分析复杂化。与此相反,在强度由于偶极再取向和渐逝场的效果从SUV到SBL荧光团转移的变化线性相关染料在SUV仍然保留的级分。第二,对于自猝灭需要高浓度的类脂的标签可能影响融合过程本身。
对于偏光TIRF激励,一个自制的设置, 如图4的成本效益,并提供了偏振特性良好的控制。然而,一个自制的设置是没有必要的,因为至少有一些商业的TIRF显微镜使用偏振光激发和关闭的,现成的双色发射分光模块可用。使用的偏振波保持(PM)光纤耦合偏振光激发光从激光器进入显微镜新兴商业显微镜成功先前25-27使用。在这样一台仪器,极化可以简单地通过旋转光纤输入到显微镜的TIRF单元PM旋转。与此相反,一些其它商业设置可以采用在TIRF单元的偏振分束器,使得偏振调整更加困难或不可能。不幸的是,往往不用于商业系统指定的偏振特性。因此,它是不希望构建一个自制的系统来讨论偏振特性,并在特定的商业选择投资之前尝试演示单元谁用户的关键。
脂质和可溶性货物放行的同时监测提供了融合进程的进一步信息。许多毛孔似乎RELE后重新密封ASING仅可溶性标签的一小部分封装入一个SUV 27。通常所有的脂质标签由孔隙重新密封时间发布,所以毛孔再密封不是单从监控脂质释放先前怀疑。标记的脂质的确,单独监测血脂发布,基斯林等人 29曾建议转移到SBL通过整个SUV膜进入SBL以极快的速度发生塌方。
后的内容部分释放在脂质体 - 拴系的双层补丁融合测定先前观察到的熔化再密封气孔。罗尔等人由〜8纳米长的DNA衬垫拴到盖玻片的54准备独立双层补丁。然后,他们引入脂质体,其融合到双层补丁被锚定到脂质体和修补膜的互补单链DNA脂质缀合物杂交来驱动。所有事件的12%是瞬时融合,产生部分的脂质体内容物释放IAL。脂质体爆裂事件,在以前的一些SUV-SBL融合试验44所示,是非常罕见的。作者建议了〜8nm的空间的双层补丁的下方,防止双层底物相互作用,也可能允许的脂质体内容的非泄漏转移到双层下方的空间。另外,所释放的可溶性标签扩散不显著在平面双层与玻璃基板之间的空间位阻。在此处所描述的测定中,平面双层类似地由聚(乙二醇) - 脂质缀合物的存在下保持〜5纳米从盖玻片路程。
可溶性和脂质货物双色监控所提供的附加信息带有一些取舍。首先,准备与两个标签的SUV是有点比单独包括脂质标签更多地参与。第二,有一些不可避免的损失信号对噪声在t从脂质标签监测信号时他同时采集速率为单色脂质释放实验(每帧15-20毫秒)。这部分损失是由于做了事实,同时作为脂质标签与可溶性SRB标记,是不是在单色实验中使用LR-PE是光明的。额外的损失是由于更高的背景,由于同时双激光激发和在光路中的其他部件( 图4)。到目前为止,较低的信号 - 噪声从单一的可靠跟踪防止美国并在两色实验标签。这反过来又防止了深入的分析,以提取可以与单色的LR-PE测量的所有信息。这一挑战可能在将来通过优化获取条件和测试其他对兼容标签与改进的性质来克服。
同时监测可溶性和脂质货物释放也可以检测半融合,其中,所述近侧小叶已经融合的状态下,但远端小叶没有。实际上,这种双色实验可以提供半融合的体外融合实验18,20的存在下,在其他的唯一结论性的评估。然而,半融合能够可靠地在单色SUV-SBL实验监测血脂单独22,44释放检测。在这样的实验中,约1/2的SUV的初始脂质标记强度保持在融合点。当远端小叶以后熔化一定时间,如果该点还没有被漂白剩余强度可能会消失。
PEG化脂质的使用介绍SBL和玻璃之间支持一个软垫一直在重现胞外分泌的SNAP25要求的关键。在聚乙二醇的协议结果刷覆盖的面向流路以及所述SBL的一侧。这具有一些好处,例如减少的SUV和SBL之间的非特异性相互作用,如在引言说明。如果用的磷脂相互作用的蛋白质SBL引入测定中,然而,所述PEG刷子将提出对这种相互作用的立体屏障。因此,对于一些应用,这将是理想的不对称包括所述PEG层,只SBL和玻璃盖玻片之间。然后SBL的面向流路侧将能够与磷脂结合蛋白,例如突触结合蛋白相互作用。这可以通过使用可共价连接到盖玻片在一端双官能PEG链和携带在其他的脂质锚来实现。然后朗缪尔-布洛杰特法可用于沉积脂质单层上的PEG层55。脂质体上面与它该疏水表面保险丝引入,形成双层56。希望这种做法将在未来微流控相结合。
可以预期的是,这里提出的SUV-SBL测定将在探索与SNARE蛋白相互作用的其他蛋白质的作用的帮助,如Munc18 / SEC1家族,钙传感器突触-1,和complexin。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者宣称,他们没有竞争的经济利益。
Materials
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| Milli-Q (MQ) water | Millipore | ||
| KOH | J.T. Baker | 3040-05 | |
| Ethanol 190 Proof | Decon | ||
| Isopropanol | Fisher Chemical | A416P4 | |
| HEPES | AmericanBio | AB00892 | |
| Sodium Cholride (KCl) | AmericanBio | AB01915 | |
| Dithiothreitol | AmericanBio | AB00490 | |
| N-[2-hydroxyethyl] piperazine-N'-[2-ethanesulfonic acid] (HEPES) | AmericanBio | AB00892 | |
| EGTA | Acros Organics | 409911000 | |
| Buffers | |||
| HEPES-KOH buffer (pH 7.4) | 25 mM HEPES-KOH, 140 mM KCl, 100 μM EGTA, 1 mM DTT | ||
| Solvents | |||
| Chloroform | J.T. Baker | 9180-01 | in glass bottle, CAUTION, wear PPE |
| Methanol | J.T. Baker | 9070-03 | in glass bottle, CAUTION, wear PPE |
| Liposome preparation | |||
| Gastight Hamilton syringe | Hamilton | var. sizes | only use glass sringe with solents (Chlorophorm/ Methanon, 2:1, v/v) |
| Glass tubes Pyrex Vista 11 ml, 16x100 mm screw cap culture tube | Pyrex | 70825-16 | clean thoroughly, rinse with chloroform |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine, 16:0-18:1 PC (POPC) | Avanti Polar Lipids | 850457 | Lipids come dissolved in CHCl3 or as lyphilized powder in sealed vials. Aliquot upon opening. Store extra as dried lipid films under inert atmosphere at -20 °C. Keep stocks in CHCl3/MeOH (2:1, v/v) at -20 °C. let come to RT before opening |
| 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (sodium salt), 18:1 PS (DOPS) | Avanti Polar Lipids | 840035 | Same as above. |
| 1-stearoyl-2-arachidonoyl-sn-glycero-3-phosphoethanolamine, 18:0-20:4 PE (SAPE) | Avanti Polar Lipids | 850804 | Same as above. |
| L-α-phosphatidylinositol-4,5-bisphosphate (Brain, Porcine) (ammonium salt), Brain PI(4,5)P2 | Avanti Polar Lipids | 840046 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(7-nitro-2-1,3-benzoxadiazol-4-yl) (ammonium salt), 18:1 NBD PE | Avanti Polar Lipids | 810145 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene glycol)-2000] (ammonium salt), 18:1 PEG2000 PE | Avanti Polar Lipids | 880130 | Same as above. |
| cholesterol (ovine wool, >98%) | Avanti Polar Lipids | 700000 | Same as above. |
| DiD' oil; DiIC18(5) oil (1,1'-Dioctadecyl-3,3,3',3'-Tetramethylindodicarbocyanine Perchlorate) | Molecular Probes | D-307 | |
| Rotavapor R-210 | Buchi | R-210 | heat bath above Tm of lipids used |
| OG n-Octyl-β-D-Glucopyranoside | Affymetrix | 0311 | store at -20 °C, let come to RT before opening |
| Shaker - Eppendorf Thermomixer R | Eppendorf | ||
| Slide-A-Lyze Dialysis Cassettes, 20k MWCO, 3 ml | life technologies | 66003 | |
| Bio-Beads SM-2 Adsorbents | Bio-Rad | 1523920 | |
| OptiPrep Density Gradient Medium | Sigma-Aldrich | D1556 | |
| Ultracentrifugation tube, Thinwall, Ultra-Clear, 13.2 ml, 14 x 89 mm | Beckman Coulter | 41121703 | |
| Beckman SW41 Ti rotor | |||
| SuflorhodamineB | Molecular Probes | S-1307 | |
| Econo-Column Chromatography Columns, 2.5 × 10 cm | Bio-Rad | 7372512 | |
| Sepharose CL-4B | GE Healthcare | 17-0150-01 | |
| SYPRO Orange Protein Gel Stain | Molecular Probes | S-6650 | 5,000x Concentrate in DMSO |
| PDMS block | |||
| Sylgard 184 Silicone elastomer kit, PDMS | Dow Corning | 3097358-1004 | |
| Pyrex glass petri dish, 150 x 20 mm, complete with cover | Corning | 3160-152 | |
| Hole puncher - Reusable Biopsy Punch, 0.75 mm | World Precision Instruments | 504529 | |
| Manual Hole Punching Machine | SYNEO | MHPM-UNV | |
| Drill .035 x .026 x 1.5 304 SS TiN coated round punch | SYNEO | CR0350265N20R4 | drill diameter: 0.9 mm, punch bore size: 0.74 mm |
| Tygon Microbore tubing, 0.25 mm ID, 0.76 mm OD | Cole-Parmer | 06419-00 | 0.010" ID, 0.030" OD |
| Silicone Tubing (0.51 mm ID, 2.1 mm OD | Cole-Parmer | 95802-00 | 0.020" ID, 0.083" OD |
| Cover glass - cleanroom cleaned | |||
| Schott Nexterion cover slip glass D | Schott | 1472305 | |
| plasma cleaner | Harrick | PDC-32G | |
| pTIRF setup and accessories | |||
| IX81 microscope body | Olympus | IX81 | |
| EM CCD camera | Andor | ixon-ultra-897 | |
| Thermo Plate, heated microscope stage | Tokai Hit | MATS-U52RA26 | |
| 1 ml hamilton glass syringes (4x) | Hamilton | 81365 | |
| syringe pump | kd Scientific | KDS-230 | |
References
- Sudhof, T. C., Rothman, J. E. Membrane fusion: grappling with SNARE and SM proteins. Science. 323, 474-477 (2009).
- Wickner, W., Schekman, R. Membrane fusion. Nat Struct Mol Biol. 15, 658-664 (2008).
- Harrison, S. C. Viral membrane fusion. Nat Struct Mol Biol. 15, 690-698 (2008).
- Jahn, R., Scheller, R. H. SNAREs--engines for membrane fusion. Nat Rev Mol Cell Biol. 7, 631-643 (2006).
- Lindau, M., Alvarez de Toledo, G. The fusion pore. Biochim Biophys Acta. 1641, 167-173 (2003).
- Staal, R. G., Mosharov, E. V., Sulzer, D. Dopamine neurons release transmitter via a flickering fusion pore. Nat Neurosci. 7, 341-346 (2004).
- Wu, Z., et al. Nanodisc-cell fusion: Control of fusion pore nucleation and lifetimes by SNARE protein transmembrane domains. Sci. Rep. 6, 27287 (2016).
- Alabi, A. A., Tsien, R. W. Perspectives on kiss-and-run: role in exocytosis, endocytosis, and neurotransmission. Ann Rev Physiol. 75, 393-422 (2013).
- Rossetto, O., Pirazzini, M., Montecucco, C. Botulinum neurotoxins: genetic, structural and mechanistic insights. Nature Rev Microbiol. 12, 535-549 (2014).
- Weber, T., et al. SNAREpins: minimal machinery for membrane fusion. Cell. 92, 759-772 (1998).
- Nickel, W., et al. Content mixing and membrane integrity during membrane fusion driven by pairing of isolated v-SNAREs and t-SNAREs. Proc Natl Acad Sci U S A. 96, 12571-12576 (1999).
- McNew, J. A., et al. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature. 407, 153-159 (2000).
- Melia, T. J., You, D. Q., Tareste, D. C., Rothman, J. E. Lipidic antagonists to SNARE-mediated fusion. J Biol Chem. 281, 29597-29605 (2006).
- Hernandez, J. M., et al. Membrane fusion intermediates via directional and full assembly of the SNARE complex. Science. 336, 1581-1584 (2012).
- Fix, M., et al. Imaging single membrane fusion events mediated by SNARE proteins. Proc Natl Acad Sci U S A. 101, 7311-7316 (2004).
- Bowen, M. E., Weninger, K., Brunger, A. T., Chu, S. Single molecule observation of liposome-bilayer fusion thermally induced by soluble N-ethyl maleimide sensitive-factor attachment protein receptors (SNAREs). Biophys J. 87, 3569-3584 (2004).
- Liu, T., Tucker, W. C., Bhalla, A., Chapman, E. R., Weisshaar, J. C. SNARE-driven, 25-millisecond vesicle fusion in vitro. Biophys J. 89, 2458-2472 (2005).
- Yoon, T. Y., Okumus, B., Zhang, F., Shin, Y. K., Ha, T. Multiple intermediates in SNARE-induced membrane fusion. Proc Natl Acad Sci U S A. 103, 19731-19736 (2006).
- Diao, J., et al. A single-vesicle content mixing assay for SNARE-mediated membrane fusion. Nat Commun. 1, 1-6 (2010).
- Kyoung, M., et al. In vitro system capable of differentiating fast Ca2+-triggered content mixing from lipid exchange for mechanistic studies of neurotransmitter release. Proc Natl Acad Sci U S A. 108, E304-E313 (2011).
- Domanska, M. K., Kiessling, V., Stein, A., Fasshauer, D., Tamm, L. K. Single vesicle millisecond fusion kinetics reveals number of SNARE complexes optimal for fast SNARE-mediated membrane fusion. J Biol Chem. 284, 32158-32166 (2009).
- Kreutzberger, A. J., Kiessling, V., Tamm, L. K. High Cholesterol Obviates a Prolonged Hemifusion Intermediate in Fast SNARE-Mediated Membrane Fusion. Biophys J. 109, 319-329 (2015).
- Schwenen, L. L., et al. Resolving single membrane fusion events on planar pore-spanning membranes. Sci Rep. 5, 12006 (2015).
- Karatekin, E., et al. A fast, single-vesicle fusion assay mimics physiological SNARE requirements. Proc Natl Acad Sci U S A. 107, 3517-3521 (2010).
- Karatekin, E., Rothman, J. E. Fusion of single proteoliposomes with planar, cushioned bilayers in microfluidic flow cells. Nat Protoc. 7, 903-920 (2012).
- Smith, M. B., et al. Interactive, computer-assisted tracking of speckle trajectories in fluorescence microscopy: application to actin polymerization and membrane fusion. Biophys J. 101, 1794-1804 (2011).
- Stratton, B. S., et al. Cholesterol Increases the Openness of SNARE-mediated Flickering Fusion Pores. Biophysical journal. 110, (2016).
- Diao, J., et al. Synaptic proteins promote calcium-triggered fast transition from point contact to full fusion. Elife. 1, e00109 (2012).
- Kiessling, V., Domanska, M. K., Tamm, L. K. Single SNARE-mediated vesicle fusion observed in vitro by polarized TIRFM. Biophys J. 99, 4047-4055 (2010).
- Blasi, J., et al. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature. 365, 160-163 (1993).
- Washbourne, P., et al. Genetic ablation of the t-SNARE SNAP-25 distinguishes mechanisms of neuroexocytosis. Nat Neurosci. 5, 19-26 (2002).
- Diaz, A. J., Albertorio, F., Daniel, S., Cremer, P. S. Double cushions preserve transmembrane protein mobility in supported bilayer systems. Langmuir. 24, 6820-6826 (2008).
- Floyd, D. L., Ragains, J. R., Skehel, J. J., Harrison, S. C., van Oijen, A. M. Single-particle kinetics of influenza virus membrane fusion. Proc Natl Acad Sci U S A. 105, 15382-15387 (2008).
- Albertorio, F., et al. Fluid and air-stable lipopolymer membranes for biosensor applications. Langmuir. 21, 7476-7482 (2005).
- Daniel, S., Albertorio, F., Cremer, P. S. Making lipid membranes rough, tough, and ready to hit the road. Mrs Bulletin. 31, 536-540 (2006).
- Gao, Y., et al. Single reconstituted neuronal SNARE complexes zipper in three distinct stages. Science. 337, 1340-1343 (2012).
- Kenworthy, A. K., Hristova, K., Needham, D., Mcintosh, T. J. Range and Magnitude of the Steric Pressure between Bilayers Containing Phospholipids with Covalently Attached Poly(Ethylene Glycol). Biophys J. 68, 1921-1936 (1995).
- Knoll, W., et al. Solid supported lipid membranes: New concepts for the biomimetic functionalization of solid surfaces. Biointerphases. 3, Fa125-Fa135 (2008).
- Quinn, P., Griffiths, G., Warren, G. Density of newly synthesized plasma membrane proteins in intracellular membranes II. Biochemical studies. J Cell Biol. 98, 2142-2147 (1984).
- Sund, S. E., Swanson, J. A., Axelrod, D. Cell membrane orientation visualized by polarized total internal reflection fluorescence. Biophys J. 77, 2266-2283 (1999).
- Johnson, D. S., Toledo-Crow, R., Mattheyses, A. L., Simon, S. M. Polarization-controlled TIRFM with focal drift and spatial field intensity correction. Biophys J. 106, 1008-1019 (2014).
- Anantharam, A., Onoa, B., Edwards, R. H., Holz, R. W., Axelrod, D. Localized topological changes of the plasma membrane upon exocytosis visualized by polarized TIRFM. J Cell Biol. 188, 415-428 (2010).
- Axelrod, D. Carbocyanine dye orientation in red cell membrane studied by microscopic fluorescence polarization. Biophys J. 26, 557-573 (1979).
- Wang, T., Smith, E. A., Chapman, E. R., Weisshaar, J. C. Lipid mixing and content release in single-vesicle, SNARE-driven fusion assay with 1-5 msec resolution. Biophys J. 96, 4122-4131 (2009).
- Chernomordik, L. V., Frolov, V. A., Leikina, E., Bronk, P., Zimmerberg, J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J Cell Biol. 140, 1369-1382 (1998).
- Takamori, S., et al. Molecular anatomy of a trafficking organelle. Cell. 127, 831-846 (2006).
- Wilhelm, B. G., et al. Composition of isolated synaptic boutons reveals the amounts of vesicle trafficking proteins. Science. 344, 1023-1028 (2014).
- Scott, B. L., et al. Liposome fusion assay to monitor intracellular membrane fusion machines. Methods Enzymol. 372, 274-300 (2003).
- Linkert, M., et al. Metadata matters: access to image data in the real world. J Cell Biol. 189, 777-782 (2010).
- Soumpasis, D. M. Theoretical analysis of fluorescence photobleaching recovery experiments. Biophys J. 41, 95-97 (1983).
- Ohki, S. A mechanism of divalent ion-induced phosphatidylserine membrane fusion. Biochim Biophys Acta. 689, 1-11 (1982).
- Berquand, A., et al. Two-step formation of streptavidin-supported lipid bilayers by PEG-triggered vesicle fusion. Fluorescence and atomic force microscopy characterization. Langmuir. 19, 1700-1707 (2003).
- Tamm, L. K., McConnell, H. M. Supported phospholipid bilayers. Biophys J. 47, 105-113 (1985).
- Rawle, R. J., van Lengerich, B., Chung, M., Bendix, P. M., Boxer, S. G. Vesicle fusion observed by content transfer across a tethered lipid bilayer. Biophys J. 101, L37-L39 (2011).
- Wagner, M. L., Tamm, L. K. Reconstituted syntaxin1a/SNAP25 interacts with negatively charged lipids as measured by lateral diffusion in planar supported bilayers. Biophys J. 81, 266-275 (2001).
- Kalb, E., Frey, S., Tamm, L. K. Formation of Supported Planar Bilayers by Fusion of Vesicles to Supported Phospholipid Monolayers. Biochimica Et Biophysica Acta. 1103, 307-316 (1992).
