

Summary
Hier presenteren we een protocol om enkele, SNARE-bemiddelde fusie gebeurtenissen tussen liposomen en ondersteund dubbellagen in microfluïdische kanalen met gepolariseerde TIRFM, met 'single molecule gevoeligheid en ~ 15 msec tijdsresolutie te detecteren. Lipide en oplosbare lading afgifte kan gelijktijdig worden gedetecteerd. Liposoom grootte, lipide diffusie, en fusie porie eigenschappen worden gemeten.
Abstract
In de alomtegenwoordige proces van membraanfusie de opening van een fusie porie stelt de eerste verbinding tussen twee voorheen afzonderlijke compartimenten. Tijdens neurotransmitter of afgifte van hormonen via exocytose, de fusie porie kan kortstondig openen en sluiten herhaaldelijk, het reguleren van de lading vrijgavekinetiek. Pore dynamiek ook de wijze van blaasje recycling vast te stellen; onomkeerbare nieuwe sluiting leidt tot voorbijgaande, "kiss-and-run" fusion, terwijl verwijding leidt tot een volledige fusie. Om beter te begrijpen welke factoren regeren porie dynamiek, ontwikkelden we een test om membraanfusie te controleren met behulp van gepolariseerd totale interne reflectie fluorescentie (TIRF) microscopie met 'single molecule gevoeligheid en ~ 15 msec tijdsresolutie in een biochemisch goed gedefinieerde in vitro systeem. Fusie van fluorescent gelabelde kleine unilamellaire blaasjes met v-SNARE eiwitten (v-SUV's) met een vlakke bilaag lager t-SNAREs, ondersteund op een zachte polymeer kussen (t-SBL, t-ondersteunde bilaag) Wordt bewaakt. De test maakt gebruik van microfluïdische stromingskanalen die zorgen voor een minimale steekproef consumptie, terwijl het leveren van een constante dichtheid van SUV's. Het benutten van de snelle verbetering van het signaal bij overdracht van lipide etiketten van de SUV naar de SBL tijdens de fusie, wordt kinetiek van lipiden kleurstofoverdracht gecontroleerd. De gevoeligheid van TIRF microscopie laat volgen enkele fluorescerende lipide labels, waaruit lipide diffusiviteit en SUV grootte kan worden afgeleid voor elke samensmelting. vrijmaking van kleurstof tijden lipide kan veel langer dan verwacht voor ongehinderde doorgang door permanent open poriën. Met een model dat veronderstelt vertraging van lipide afgifte wordt veroorzaakt door het trillen porie een porie "openheid", de fractie van tijd de poriën ook voor fusie blijft, kunnen worden geschat. Een oplosbaar marker kan worden ingekapseld in de SUV's voor de gelijktijdige monitoring van lipiden en oplosbare lading release. Dergelijke metingen geven een aantal poriën hersluiten na het verliezen van een gedeelte van het oplosbare lading.
Introduction
Membraan fusie is een universeel biologisch proces nodig is voor intracellulaire handel in vetten en eiwitten, secretie, de bevruchting, de ontwikkeling en omhuld virus binnenkomst in gastheerorganismen 1-3. Voor de meeste intracellulaire fusiereacties waaronder afgifte van hormonen en neurotransmitters via exocytose wordt de energie twee lipide bilagen smelten door vorming van een vier-helix bundel tussen cognate oplosbare N-ethylmaleimide-gevoelige factor attachment eiwitreceptor (SNARE) eiwitten, verankerd in het blaasje (v-SNARE) en het doelmembraan (t-SNARE) 4 resp. Synaptische vesikel exocytose is de meest strak gereguleerd fusiereactie en voordat één milliseconde na aankomst van een actiepotentiaal 1,4,5. De fusie porie, de initiële verbinding tussen de twee compartimenten fuseren, kunnen flikkeren open en meerdere malen gesloten voordat het opnieuw afsluiten of het uitbreiden van onomkeerbaar 5-7. De voormalige resultatenin voorbijgaande, "kiss & run" fusion, terwijl de laatste leidt tot volledige fusie. Factoren die het evenwicht tussen deze twee modi van fusie en mechanismen reguleren poriën flikkeren zijn niet goed begrepen 5,8.
SNARE eiwitten zijn nodig voor exocytose; synaptische blaasje fusie wordt na splitsing van SNAREs afgeschaft door neurotoxins 9. Bulk fusie-experimenten met kleine unilamellaire vesikels (SUV's) aangetoond dat SNAREs niet alleen vereist, maar ook voldoende membraanfusie 10 rijden. In deze massa assay SUV gereconstitueerd met v-SNAREs (v-SUV) werden gedoteerd met fluorescente fosfolipiden (N - (7-nitro-2-1,3-benzoxadiazol-4-yl) -phosphoethanolamine (NBD-PE) en ( N -. (lissamine rhodamine B sulfonyl) -phosphoethanolamine (LR-PE) en gemengd met ongelabeld blaasjes met t-SNAREs (t-SUV) Aanvankelijk was de fluorescentie van NBD-PE in v-SUV is geblust door Förster resonance energy transfer (FRET ) naar LR-PE. Zoals labeLED v-SUV fuseren met ongelabelde t-SUV, is de fluorofoor oppervlaktedichtheid in het nu gecombineerde membraan verminderd en de resulterende toename van NBD-PE fluorescentie meldt de mate van lipide mengen 10. Aangezien het grootste deel test is eenvoudig in te stellen en te analyseren, is het op grote schaal gebruikt om de mechanismen van SNARE gemedieerde fusie 10-14 bestuderen. Het heeft echter verscheidene beperkingen, zoals lage gevoeligheid en slechte tijdsresolutie. Belangrijker, als ensemble metingen is gemiddelde resultaten voor alle gebeurtenissen maken onderscheid tussen docking en fusion, alsmede opsporing van hemifusion tussenproducten moeilijk.
De afgelopen tien jaar verschillende groepen, waaronder de onze, zijn er nieuwe tests ontwikkeld om fusie gebeurtenissen bij de enkele blaasje level 15-27 volgen. Ha en collega's gebruikt v-SUV vastgebonden op een oppervlak en bewaakt hun fusie met gratis t-SUV 18,19. Lipide mengen werd gevolgd met behulp van FRET tussen twee lipide gebonden fluoroforen embedden in de v- en t-SUV's, respectievelijk, met behulp van totale interne reflectie fluorescentie (TIRF) microscopie 18. Later, Brunger het laboratorium gebruikt een enkele lipide-label soorten samen met een inhoud marker voor gelijktijdige detectie van lipiden en inhoud te mengen 20,28. Zowel de lipide en de inhoud merkers werden opgenomen in hoge zelfdovende concentraties; fusie met ongelabelde SUV resulteerde in fluorescentie dequenching 20,28.
Anderen hebben v-SUV gefuseerd met vlakke dubbellagen opgelost in t-SNAREs 15-17,21-27,29. De vlakke geometrie van het doel (t-SNARE bevattende) dubbellaag beter bootst de fysiologische fusieproces van kleine, zeer gebogen blaasjes met een platte plasmamembraan. De Steinem groep toegepast poriën spanning membranen opgelost in t-SNAREs, opgehangen boven een poreus siliciumnitride substraat en gedetecteerd fusion individuele v-SUV met behulp van confocale laser scanning microscopie 23. anderen fgebruikte v-SUV's naar vlakke dubbellagen opgelost in t-SNAREs, ondersteund op een glazen substraat 15-17,21,22,24-27,29. Het grote voordeel van het gebruik van ondersteunde bilagen (SBLs) dat TIRF microscopie kan worden gebruikt voor docking en fusiegebeurtenissen met uitstekende signaal-ruisverhouding en zonder interferentie van vrije v-SUV detecteren, hoewel behulp microfluïdische ook resolutie eenmalige gebeurtenis gebruikt standaard ver-field epifluorescentie microscopie 24.
Een belangrijk punt van zorg is de vraag of en hoe substraat-bilaag interacties invloed op ondersteunde bilaag kwaliteit en het fusieproces. Vroeg werk gebruik gemaakt van vlakke SBLs die direct werden ondersteund op een glazen of kwarts substraat 15-17. Deze SBLs werden door adsorptie, barsten, verspreiding en fusie van t-SUV membranen op het substraat. Het werd al snel besefte echter dat het weglaten van een belangrijke t-SNARE component, SNAP25, van SBLs op deze manier bereid resulteerde in v-SUV docking en fusion kinetiek indistinguishable van die verkregen met een totale t-SNAREs 17. Omdat SNAP25 absoluut voor fusie is nodig in vivo 30,31, werd de fysiologische relevantie van deze vroege pogingen in twijfel te trekken. Tamm de groep overwon deze uitdaging door het gebruik van beter gecontroleerd ondersteunde bilaag vorming 21. Vroeger Langmuir-Blodgett depositie voor het eiwit-vrij eerste folder van de SBL, gevolgd door de fusie van die monolaag met t-SUV's 21. Dit resulteerde in SNAP25-afhankelijke fusie.
Om potentiële artefacten geassocieerd met een dubbellaag direct afgezet op een glassubstraat zonder nader Langmuir-Blodgett methoden voorkomen Karatekin et al. Werd een zachte, gehydrateerde poly (ethyleenglycol) (PEG) kussen tussen de bilaag en het substraat 24. Deze modificatie resulteerde ook in SNAP25-afhankelijke fusie 24. Dubbellagen kussens op een zachte polymeerlaag was gekend om transmembraan beter behoudenmobiliteit en de functie 32-eiwit, en was gebruikt in fusion studies met virussen 33. Bovendien gePEGyleerd dubbellagen lijken wat vermogen om zichzelf te genezen behouden en zijn zeer robuust 34,35. Eerst wordt een fractie van commercieel verkrijgbare, lipide-gebonden PEG-ketens zijn opgenomen in de t-SUV membraan. Wanneer deze t-SUV burst en vormen een vlakke dubbellaag op een glazen substraat, een PEG borstel omvat zowel bladen van de vlakke bilaag. Omdat vorming vlakke dubbellaag wordt aangedreven door adhesie van het PEG-ketens rond t-SUV op het hydrofiele glasoppervlak, liposoom barsten en vlakke bilaag vorming betrekkelijk ongevoelig voor de lipidesamenstelling gebruikt. Wanneer echter grote hoeveelheden cholesterol zijn opgenomen, waardoor de cohesieve eigenschappen van de SUV, de SUV kan niet spontaan barsten. Als dit het geval is, kan osmotische shock of tweewaardige ionen worden toegepast om vlakke bilaag vorming 25 helpen.
Zoals hierboven vermeld, in dit apdering een PEG borstel omvat beide zijden van de vlakke, ondersteund dubbellaag. De borstel tegenover de microfluïdische stromingskanaal helpt om niet-specifieke hechting van inkomende v-SUV die ook meestal bedekt met een PEG-laag voorkomen. Vorming van V- en t-SNARE complexen begint vanaf de membraan-distale N-uiteinden en verloopt in etappes in de richting van het membraan-proximale domeinen 36. Voor de v-SUV om met de t-SBL, V- en t-SNARE N-termini nodig boven het PEG borstels, die lijkt op het geval onder de omstandigheden van de assay uitsteken. Borstelhoogte kan worden aangepast aan andere eiwitten dan SNAREs bestuderen door het variëren van de dichtheid van gepegyleerde lipiden en PEG ketenlengte 37,38. Een ander voordeel van de PEG borstels die de proximale oppervlakken van het fuseren van bilagen is dat zij bootsen de drukke omgeving van biologische membranen die zijn verpakt met 30.000-40.000 integrale membraaneiwitten per vierkante micron 39. Net als de PEG-ketens in deze test, de Repulsive eiwit laag die biologische membranen moeten opzij worden geduwd om te zorgen voor contact tussen de twee fosfolipiden dubbellagen voor fusie te komen.
Microfluïdische stromingskanalen worden in deze test, omdat ze unieke voordelen bieden. Ten eerste microfluïdische stroom kunnen meer uniforme afzetting van t-SUV te verspreiden en de zekering t-SBL vormen. Ten tweede, de kleine kanaal volume (<1 pl) minimaliseert monster consumptie. Ten derde, de kleine volumina vereist waarvan de hele experiment constant debiet te worden uitgevoerd. Stroom verwijdert zwak, vermoedelijk aspecifiek, gehecht v-SUV van SBL 16. Het heeft ook een constante dichtheid van v-SUV boven de t-SBL, het vereenvoudigen van kinetische analyse 17. Tenslotte worden gedokt blaasjes gemakkelijk te onderscheiden van vrije degenen door de stroming 25 uitgevoerd. Ten vierde, meerdere microkanalen kunnen worden gebruikt op dezelfde dekglaasje, elk probing een andere toestand. Dit maakt vergelijking van de voorwaarden During dezelfde experimentele run. Een soortgelijke aanpak is gebruikt door de van Oijen groep fusie tussen influenzavirus en kussens SBLs 33 bestuderen.
Bij TIRF microscopie, de exponentiële verval van het verdwijnende veld (met vervalconstante ~ 100 nm) fluorescentie excitatie beperkt tot die moleculen die in de directe nabijheid van het glas-buffer interface. Dit minimaliseert bijdrage van fluorescente moleculen die verder weg zijn, verhoogt de signaal-ruisverhouding en maakt enkel molecuul gevoeligheid belichtingsraam tijden van 10-40 msec. Het verdwijnende veld leidt ook tot een toename van signaal-fusie: de overdracht gemerkte lipiden uit de SUV in de SBL, bevinden zij zich gemiddeld sterker bekrachtigingsveld. Deze toename in fluorescentie wordt sterker grotere liposomen.
Als gepolariseerd licht wordt gebruikt om het verdwijnende veld genereren extra effecten bijdragen tot een verandering in fluorescentie bij transfer van etiketten van de SUV in de SBL. Sommige lipide kleurstoffen overgang dipool georiënteerd met een voorkeurs gemiddelde hoek ten opzichte van de dubbellaag waarin zij zijn ingebed. Hierdoor ontstaat een verschil in de hoeveelheid fluorescentie die door de fluoroforen wanneer zij in de SUV versus de SBL, aangezien het gepolariseerde bundel kleurstoffen anders prikkelen de twee membranen. In het eerste geval zal de excitatie balk interactie met overgang dipolen georiënteerde rond de sferische SUV, terwijl voor het laatste, zal dipool oriëntaties worden beperkt door de vlakke SBL geometrie. Wanneer bijvoorbeeld s-gepolariseerd invallend licht (gepolariseerd loodrecht op het vlak van inval) wordt gebruikt, excitatie efficiënter als de kleurstof in de SBL dan in SUV voor een lipide kleurstof transitiedipool evenwijdig aan het membraan 29,40 (zoals die van Dil of heb 41-43). Een SUV gedoteerd met zo'n fluorofoor verschijnt dimmen wanneer het dokken op de SBL (figuur 7, Representat ive resultaten). Als een fusie porie opent en verbindt de SUV en SBL membranen, fluorescerende probes diffunderen in de SBL en meer kans te worden enthousiast over de-s gepolariseerde verdwijnende veld 25,27,29. Bijgevolg het fluorescentiesignaal geïntegreerd rondom de fusieplaats sterk toeneemt tijdens de kleurstofoverdracht van de SUV in de SBL 27 (Figuur 3 en Figuur 7). Een bijkomende factor die bijdraagt aan veranderingen die gepaard fusie wordt dequenching van fluorescente labels zoals ze worden verdund wanneer overgebracht in de SBL signaal. De bijdrage van dequenching gewoonlijk klein in vergelijking met verdwijnende veld verval en polarisatie-effecten in de hier beschreven test, omdat slechts een kleine fractie (  ) Van de lipiden zijn gelabeld.
) Van de lipiden zijn gelabeld.
Het signaal stijging op fusie kan worden benut om fusie porie eigenschappen afleiden door de tijd te vergelijken,1 "src =" / files / ftp_upload / 54349 / 54349eq2.jpg "/>, die nodig is voor een lipide te ontsnappen via een porie die vrij doorlaatbaar voor lipiden aan de werkelijke release tijd,  . Als de twee tijdschalen vergelijkbaar zou worden geconcludeerd dat de poriën weer weinig weerstand tegen lipide flow. Indien de werkelijke lostijd aanzienlijk langer is dan de tijd voor diffusie beperkte release, zou een proces aangeven, zoals poriën flikkeren, vertragen lipide release. De diffusie beperkte release tijd,
. Als de twee tijdschalen vergelijkbaar zou worden geconcludeerd dat de poriën weer weinig weerstand tegen lipide flow. Indien de werkelijke lostijd aanzienlijk langer is dan de tijd voor diffusie beperkte release, zou een proces aangeven, zoals poriën flikkeren, vertragen lipide release. De diffusie beperkte release tijd,  , Afhankelijk van de grootte van de fuser en liposoom lipide-diffusie; haar ramingen vereist deze twee parameters te worden gekwantificeerd. De single molecule gevoeligheid van de test maakt het mogelijk lipide diffusie te meten door het bijhouden van meerdere afzonderlijke lipide fluoroforen na hun vrijlating in het SBL voor elke fusie event 26. De grootte van elke versmelten vesiclekan worden geschat 27 door het combineren van (i) de intensiteit van een enkele lipide kleurstof, (ii) de verandering in de totale fluorescentie rond docking site na alle fluoroforen op fusie worden overgebracht in de SBL, (iii) de bekende etikettering dichtheid van SUV lipiden, en (iv) de oppervlakte per lipide. Voor veel fusiegebeurtenissen werden de werkelijke lipide afgifte samengesteld worden veel langzamer dan verwacht door diffusie gecontroleerde afgifte 27 zijn, zoals eerder werd opgemerkt uitgaande uniforme SUV maat 44. Uitgaande vertraging van lipide afgifte wordt veroorzaakt door het trillen poriën, een kwantitatief model maakt schatting van "porie openheid", de fractie van tijd de poriën blijft ook voor fusie-27.
, Afhankelijk van de grootte van de fuser en liposoom lipide-diffusie; haar ramingen vereist deze twee parameters te worden gekwantificeerd. De single molecule gevoeligheid van de test maakt het mogelijk lipide diffusie te meten door het bijhouden van meerdere afzonderlijke lipide fluoroforen na hun vrijlating in het SBL voor elke fusie event 26. De grootte van elke versmelten vesiclekan worden geschat 27 door het combineren van (i) de intensiteit van een enkele lipide kleurstof, (ii) de verandering in de totale fluorescentie rond docking site na alle fluoroforen op fusie worden overgebracht in de SBL, (iii) de bekende etikettering dichtheid van SUV lipiden, en (iv) de oppervlakte per lipide. Voor veel fusiegebeurtenissen werden de werkelijke lipide afgifte samengesteld worden veel langzamer dan verwacht door diffusie gecontroleerde afgifte 27 zijn, zoals eerder werd opgemerkt uitgaande uniforme SUV maat 44. Uitgaande vertraging van lipide afgifte wordt veroorzaakt door het trillen poriën, een kwantitatief model maakt schatting van "porie openheid", de fractie van tijd de poriën blijft ook voor fusie-27.
Wanneer praktisch is belangrijk om fusie mechanismen waarbij zowel lipide en oplosbare inhoud labels testen. Bijvoorbeeld zou lipide afgifte vertraagd worden door andere dan de porie trillen werkwijzen, zoals restrictie lipide diffusie door de SNARE eiwitten die surround thij porie. Als dit het geval zou zijn, dan los van de inhoud zou release van lipide labels voorafgaan, op voorwaarde dat de porie is groot genoeg om doorgang van oplosbare probes mogelijk te maken. Een fundamentele fout in de benadering kan in de veronderstelling dat de overdracht van gemerkte lipiden SBL vindt plaats via een smalle fusie porie verbinden de SBL een vesicle die grotendeels zijn pre-fusie-vorm heeft behouden. Lipid overdracht in de SBL kan ook het gevolg zijn van de snelle uitzetting van de fusie porie met een gelijktijdige, extreem snelle ineenstorting van de SUV in de SBL membraan, zoals eerder gesuggereerd op basis van lipide vrijlating data alleen 29. Monitoring zowel lipide en inhoud kort tegelijk bleek dat veel poriën afgedicht na het loslaten alle lipide labels, maar bleven sommige van hun oplosbare lading 27. Dit geeft aan dat ten minste enkele liposomen niet samen in de SBL na fusie en dat het lipide kleurstofoverdracht in de SBL gebeurt door een fusie porie. Bovendien, lIPID en inhoud vrijlating vond plaats gelijktijdig 27, waardoor het onwaarschijnlijk is dat de vertraging van lipide vrijlating was te wijten aan hinder van lipide diffusie door de SNARE eiwitten rond de porie 45.
Een SUV-SBL fusie-protocol dat niet controleren oplosbaar inhoud vrijlating is eerder gepubliceerd door Karatekin en Rothman 25. Hier worden meer recente ontwikkelingen opgenomen, namelijk de gelijktijdige controle van lipiden en de inhoud vrij te geven en de schatting van SUV, lipide, en fusie porie eigenschappen 27. Het protocol begint met instructies voor het bereiden van de microfluïdische cellen, gemaakt door het binden van een poly (dimethylsiloxaan) (PDMS) elastomeer blok met groeven met een glas dekglaasje 25. Vervolgens wordt de bereiding van v-SUV met zowel lipide en inhoud markers toegelicht. Afdelingen 4 en 5 geven instructies voor de montage van de microfluïdische cellen, de vorming van de SBLs in situ en het controleren op gebreken en vloeibaarheid, de invoering van vSUV's in de stroom cellen en detectie van fusiegebeurtenissen. Hoofdstuk 6 bevat instructies voor data-analyse.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Bereiding van een PDMS blok naar de microfluïdische kanaal Formulier

Figuur 1. Microfabricage van stroomcel template en PDMS block bereiding. (A) Ontwerp van een vierkanaals stroomcel die past op een 24 x 60 mm glazen dekglaasje (onder). Zes identieke ontwerpen zijn ingericht om te passen op een 10 cm silicium wafer (boven). (B) Knip blok van ongeveer 5-8 mm dik PDMS op een perforator. (C) Het inbrengen van de slang in het gaatje met behulp van een pincet. Klik hier om een grotere versie van deze figuur te bekijken.
- Verkrijgen van een stroomcel template zoals in figuur 1A. Typische kanalen zijn 0,3-1 mm breed, 70-100 micrometer hoog en 1-2 cm lang. Fabriceren van een sjabloon onsing standaard fotolithografie technieken in een cleanroom faciliteit 25 of bestelling als de toegang cleanroom is niet beschikbaar. Alternatief machine een matrijs met grotere afmetingen van een geschikt materiaal.
Opmerking: Cleanroom personeel kan trainen en begeleiden onervaren gebruikers (toegang tot een cleanroom faciliteit is altijd beperkt tot gebruikers met de juiste opleiding) in het ontwerpen en bestellen van een masker, wafer schoonmaken, en fotolithografie. Zodra een sjabloon wordt verkregen, kan het buiten de cleanroom worden herhaaldelijk gebruikt, mits het in een gesloten schaal gehouden en zorg is genomen om stof uit te sluiten. - Maak een mengsel van ~ 100 ml PDMS (siliconenelastomeer base) en ~ 10 ml verknopingsmiddel (verharder) in een wegwerp plastic beker. Breek het topje van een wegwerp pipet om de viskeuze PDMS verwerken gemakkelijker. Weeg de PDMS in de plastic beker als pipetteren is niet nauwkeurig.
- Roer het mengsel goed. Verwijder de vele luchtbellen door ontgassen in een vacuüm exsiccator (ongeveer 20min). Als de beker onder vacuüm wordt gebracht, zal de bubbels aanvankelijk groeien in omvang, verhoging van het mengsel sterk. Controle van de onderdruk die wordt toegepast en zorg ervoor dat de beker is diep genoeg om een lekkage te voorkomen.
- Giet een grote daling van de ontgast PDMS mengsel in een glazen petrischaal (150 mm x 20 mm) en druk op de wafer op de PDMS met de sjabloon naar boven. Dit voorkomt luchtbellen gevangen onder de wafer. Opgesloten bellen zullen uitbreiden en kan de wafer kantelen wanneer de schotel wordt geplaatst in de oven. Giet ontgast PDMS op de mal bovenop de wafel totdat deze onder ongeveer 5-8 mm PMDS. Als voorzichtig een luchtbel optreedt verwijderen met een pipet tip.
- Bak de PMDS in een oven bij 60 ° C gedurende 3 uur. Zorg ervoor dat het gerecht is niveau.
- Gebruik een nieuw scalpel te snijden een PDMS blok met de gevormde kanaal structuren. De uitgesneden blok moet passen op een dekglaasje (zie stap 4.1.9).
- Schil de uitgesneden blok van het gerecht en plAce It op een stuk van schone aluminiumfolie.
Opmerking: Het verknoopte PMDS blokken kan enkele maanden worden bewaard. Op een enkele wafer, zijn er 6 reeksen stromingskanalen (figuur 1A), zodat 6 PDMS blokken kunnen in één PDMS molding. Na het snijden alle 6 van de PDMS blokken, schone losse stukjes PDMS, maar andere wijze de resterende PDMS niet verwijderen voor het storten en verknopen de volgende groep, die slechts ongeveer de helft van de PDMS voor de eerste partij vereist. Een goede template kan een paar jaar duren. - Gebruik een perforator te boren door de PDMS blok in één rechte beweging (Figuur 1B). Begin aan de kant van het kanaal groeven. Zorg ervoor dat u de gestanst stuk PDMS te verwijderen. Herhaal dit voor alle acht gaten van de vier kanalen design.
- Plaats de PDMS blok kanaal naar beneden op een nieuwe en kreukvrij stuk aluminiumfolie. Bewaar het blok tot enkele maanden in een droogkast, idealiter in een exsiccator.
- Duw de buis (0,25 mm ID, 0,76 mm OD) ongeveer een derde in het gaatje met een pincet (Figuur 1C). Snijd de buis schuin gemakkelijker inbrengen. Reactie buizen lang genoeg om de SUV reservoir en de spuitpomp bereikt, respectievelijk. Na het plaatsen van de gemonteerde chip op de microscoop, snijd de buizen opnieuw als ze te lang zijn.
- Om verbinding te maken met het spuiten van de pomp, snijd een stukje van de grotere silicone buis (0,51 mm ID, 2,1 mm OD) en steek de dunnere buis in één kant. Deze kant-en-klare blok PDMS kan worden gehouden gedurende enkele maanden.
2. Coverslip Cleaning
- Schone dekglaasjes volgens de in Karatekin en Rothman 25 beschreven protocol met een sterk oxiderende mengsel van zwavelzuur (H 2 SO 4) en waterstofperoxide (H 2 O 2). Voer deze procedure in een cleanroom, en de juiste veiligheidsmaatregelen te nemen. U kunt ook gebruik maken van een cleanroom gereinigd coverslip dat in de handel verkrijgbaar (zie Materials List).
3. Voorbereiding van v-SUVS die zowel Lipid en Content Labels

Figuur 2. Schematische voorstelling van SUV preparaat. Lipiden worden gemengd in een glazen buis (1) en het oplosmiddel wordt verdampt om een lipide film te vormen door draaiing van de buis in een waterbad (2). Resterende sporen oplosmiddel verwijderd onder hoog vacuüm (3). De lipidefilm gehydrateerd in reconstitutie buffer die detergens en eiwit terwijl gewerveld (4). Als inhoud kleurstof te kapselen wordt in deze stap en in de verdunningsstap (5). Verdunning van de concentratie detergens beneden de kritische micel concentratie leidt tot liposoomvorming. Wasmiddel wordt gedurende de nacht gedialyseerd weg (6). Voor t-SUV voor SBL vorming inbegrip NBD-PE (groen), zijn vesicles dreef in een dichtheidsgradiënt en verzameld ophet grensvlak tussen twee lagen (7a). Om v-SUV's te scheiden met ingekapselde inhoud marker van vrije kleurstof het monster wordt overreden een grootte-uitsluiting-kolom en verzameld in 0,5 ml fracties (7b). Klik hier om een grotere versie van deze figuur te bekijken.
- Bereid 4 L reconstitutie buffer, die 25 mM HEPES-KOH, 140 mM KCI, 100 uM EGTA en 1 mM DTT, pH 7,4. Met meeste buffer dialyse en 100 ml voor andere stappen.
Opmerking: Andere buffers kunnen worden gebruikt, maar het is belangrijk om de buffer osmolariteit consistent in het experiment te houden en kies omstandigheden waarin de eiwitten functioneel. Voor het bereiden van v-SUV's met alleen lipide labels en t-SUV's, volg Karatekin en Rothman 25. - Lipide voorraden bij -20 ° C in chloroform en chloroform worden opgeslagen: methanol (2: 1, v / v), laat injectieflacons bereikt kamertemperatuur voorafgaand aanvoorkomen condensatie.
- Gespoeld glazen buis met chloroform om sporen van reinigingsmiddel te verwijderen en meng de lipiden in de gewenste verhouding met een uiteindelijke hoeveelheid van 1 pmol in een mengsel van chloroform en methanol (2: 1, v / v). Gebruik alleen glazen spuiten / buisjes aan organische oplosmiddelen / opgeloste lipiden te verwerken.
Opmerking: De lipiden die hier gebruikt zijn POPC: SAPE: DOPS: Cholesterol: PEG2000-DOPE: deed op een molaire verhouding van 57,4: 15: 12: 10: 4,6: 1 voor v-SNARE bevattende vesicles en POPC: SAPE: DOPS: Cholesterol : hersenen PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5) voor t-SNARE SUV. Zie Materiaal voor meer informatie. Andere lipide samenstellingen kunnen worden gebruikt. - Damp het oplosmiddel onder een zachte stroom van stikstof of in een rotatieverdamper. Dompel het uiteinde van de buis in een waterbad (37 ° C) boven de hoogste smelttemperatuur van de lipiden verwarmd tot een homogene lipidefilm opbrengst en grote temperatuurverschillen vermijden tijdens het verdampen van het oplosmiddel. Begin vacuüm bij ongeveer300 mbar totdat de lipide film wordt gevormd, vervolgens doorgaan ~ 2 minuten bij hoogste vacuüm in de roterende verdamper.
- Verwijder de glazen buis en wikkel het in aluminiumfolie om blootstelling aan licht te vermijden en verwijderen van resterende sporen oplosmiddel door het plaatsen van de buis in een exsiccator onder hoog vacuum gedurende ten minste 2 uur. Deze stap kan ook worden uitgevoerd 's nachts.
- De gedroogde lipidefilm rehydrateren, bereid een mengsel van detergens (n-octyl-β-D-glucopyranoside, OG), eiwit (v-SNAREs) en SRB in reconstitutie buffer (500 ui eindvolume). Houd de uiteindelijke detergensconcentratie ~ 2 maal de kritische micel concentratie (CMC, 20-25 mM voor OG) en stel het eindvolume zodat het detergens lipide verhouding> 10. Ontbinden Sulforhodamine B (SRB) poeder in het eiwit-detergens oplossing tot een eindconcentratie van 10-50 mM SRB verkrijgen door voorzichtig schudden van de oplossing. Stel de osmolariteit van de reconstitutie buffer bij het toevoegen van hoge concentraties SRB doordat de concentration van kaliumchloride dienovereenkomstig.
Opmerking: Lipid-to-eiwitverhouding (LP) voor t-SNAREs is hoog (LP ~ 20.000, ~ 70 t-SNAREs per vierkante micron. 25). Significant hoger t-SNARE dichtheden in de SBL kan leiden tot inactieve eiwit aggregaten met een significante vermindering van het fusie-tarieven 17,24. LP voor v-SNAREs veel lager (LP ~ 200, 7000 ~ v-SNAREs per um 2), nabij het v-SNARE dichtheden gevonden synaptische blaasjes 46,47. Om de eiwitfunctie na recombinante expressie en zuivering van v- en t-SNAREs waarborgen is het nuttig om een eenvoudige bulk fusie-assay 48 voeren alvorens alle stappen van de interne vesikel fusie assay. - Schud de voorverwarmde glazen buis met de gedroogde lipide film tijdens het toevoegen van het eiwit-detergens-SRB-oplossing van stap 3,6. Probeer te voorkomen dat het creëren van bubbels. Blijf schudden gedurende 15 minuten bij 37 ° C.
- Verdun het schoonmaakmiddel vervijfvoudigde door het toevoegen van reconstructiebuffer die SRB (voeg 2 ml, 2,5 ml uiteindelijk volume), terwijl snel vortexen concentratie gradiënten te vermijden. Blijf schudden gedurende 15 minuten tot 1 uur bij 37 ° C.
Let op: Langere incubatie toename eiwit reconstitutie efficiëntie. De snelle verdunning vermindert de concentratie detergens onder de CMC en leidt tot de vorming van kleine liposomen. - Dialyseer de vesicle suspensie eerst tegen ~ 1 L reconstitutie buffer gedurende 1-2 uur bij omgevingstemperatuur geroerd en vervolgens met 3 L reconstitutie buffer overnacht bij 4 ° C met 4 g polystyreen adsorptiemiddel, met een 20.000 MWCO dialysebuis of cassette. Gebruik verschillende bekers voor dialyse van vesicles met en zonder SRB om kruisbesmetting te voorkomen.
- Equilibreer een gelfiltratiekolom met reconstitutie buffer. Voer het vesicle suspensie door de kolom vrij SRB scheiden van de v-SUV met ingekapselde SRB. Gebruik reconstitutie buffer zonder SRB als elutiemiddel zodra het gehele monsterging de kolom. Verzamelen v-SUV in 0,5 ml fracties.
- Controleer succesvolle SRB inkapseling op zichzelf gedoofd concentraties door het meten van SRB fluorescentie vóór en na de toevoeging van het wasmiddel een hoeveelheid van 16 blaasjes. Met behulp van een fluorescentie spectrometer, prikkelen de steekproef bij 550 nm en scan de SRB-uitstoot tussen 570 nm en 630 nm. Verwacht een 4-8 voudige toename in fluorescentie SRB afhankelijk van de ingekapselde hoeveelheid wasmiddel na toevoeging als het membraan oplosbaar en bezit SRB verdund.
- Kenmerken de lipiden en eiwitten uit via fluorescentiespectroscopie 24 en SDS-PAGE gelelektroforese, respectievelijk. Gebruik een gevoelige kleuring methode, in het bijzonder voor de hoge LP t-SUV monsters (zie Materials List). Typisch ongeveer 50% van zowel lipide- en eiwit ingang verloren tijdens de bereiding waardoor een LP dicht bij de nominale waarde.
- Karakteriseren SUV maten met behulp van dynamische lichtverstrooiing 24 of elektron Micrpy 48. WINKEL SRB met v-SUV bij 4 ° C tot ~ 3-4 dagen. Niet in de vriezer, als het invriezen en ontdooien breekt het membraan en releases ingekapseld SRB.
4. SUV-SBL Fusion Analyse Monitor Lipid Alleen release
- Vorming van de tethered ondersteunde bilaag binnen de microfluïdische stromingskanalen
- Plaats de PDMS blok (stap 1.11) onder hoog vacuum gedurende ten minste 20 min voorafgaand aan het experiment om opgeloste gassen te verwijderen. Dit reduceert de kans op luchtbellen in de microkanalen in het fusie-experiment.
- Schakel de microscoop setup en verwarm het podium monsterhouder (figuren 3 en 4) op de gewenste temperatuur.
- Filter de reconstitutie buffer gebruikt om de vesicles te verdunnen door een filter met 0,45 urn of kleiner poriegrootte.
- Verdun ~ 30 ul van de NBD-PE gelabelde t-SUV's of eiwit-vrij (pf-SUV's) controle liposomen (voorraadoplossing 0,5-1 mM lipide) met ~ 60 pi buffer. De eindconcentratie is hier niet kritisch.
- Ontgas dit mengsel met behulp van een 3 ml spuit. Druk op het grootste deel van de lucht boven het monster terwijl de spuit verticaal. Sluit de tip (naald gratis) met behulp van paraffine film en creëren een vacuüm door te trekken naar beneden de zuiger. Tik de injectiespuitcilinder ontgassing van de oplossing te versnellen. Herhaal dit proces een paar keer totdat er geen luchtbellen meer voordoen wanneer vacuüm wordt toegepast.
- Gebruik een hypodermische naald met een diameter iets groter dan de buis die de PDMS blok is bevestigd aan een perforator op het dekken van een microcentrifugebuis. Zorg ervoor dat het doorgestoken stukje plastic is niet in de buis als het de slang later aangesloten op de microfluïdische kanaal inlaat kunnen verstoppen.
- Vul het ontgast SUV oplossing in de microcentrifugebuis met een gat in de dop en plaats deze in de houder op de microscoop podium in evenwicht tot de ingestelde temperatuur.
- place een vooraf gereinigd dekglaasje (deel 2) in een plasma schoner en lopen lucht plasma voor ongeveer 5 minuten. Zet de plasma behandelde dekglaasje (behandelde zijde naar boven) op een paar pluisvrije weefsels dienen als een kussen.
- Verwijder de aluminiumfolie van het ontgaste PDMS blok en zet het blok bovenop het dekglaasje. Bij hergebruik van de PDMS blok gezet op en maak een stukje plakband om het kanaal kant om het schoon te maken. Druk de PDMS te blokkeren op het dekglaasje met behulp van een pincet om het te plakken, maar niet te hard als het glas zou kunnen breken op.
- Plaats de gemonteerde doorstroomcel op de microscoop podium en sluit de slang aan de SUV reservoir en de spuitpomp respectievelijk. Tape het dekglaasje naar het podium (Figuur 3B).
- Begin opzuigen SUV bij 3 gl / min totdat de oplossing de kanalen opvult (~ 2,25 mm / sec voor 75 urn x 300 urn kanaaldoorsnede). Als oplossingen voor alle kanalen beginnen omhoog te bewegen the slang aan de uitgangszijde, verminderen stroom naar 0,5 pl / min en incubeer gedurende 30-45 min.
Opmerking: De tijd tussen plasma behandelen van het glaasje en stroomt de SUV in het kanaal niet meer dan 10-20 min het effect van de plasmabehandeling is voorbijgaand. - Controleer de kanalen voor een lek. Gebruik een 10-20x doelstelling lucht naar NBD-PE fluorescentie observeren vanuit het NBD-PE bevatten t- of pf-SUV's. Excite NBD fluoroforen met behulp van de 488 nm laser. Lekken zijn moeilijk op te sporen met behulp van helderveld verlichting.
- Zet een buis met ontgast reconstitutie buffer in de houder en laat de temperatuur in evenwicht komen zo snel veranderingen in temperatuur kan leiden tot defecten in het SBL. Stop de stroom en wacht ~ 1 min om te controleren of de doorstroming volledig is gestopt en er geen luchtbel zal worden opgezogen in de buis voordat u de inlaat buizen naar de ontgaste buffer.
- Spoel alle kanalen met ontgast buffer om ongebonden SUV's weg te wassen.
- Overschakelen naar een higher vergroting TIRF doelstelling (60X, olie, NA 1,45-1,49) en controleer of de bilaag lijkt homogeen en vrij is van alle voor de hand liggende grootschalige gebreken, zoals donkere vlekken of lipide buisjes uit dat zich uitstrekt van de SBL.
- Controleer de vloeibaarheid van de bilaag
- Als een specifieke FRAP eenheid niet beschikbaar is, of als een FRAP sequentie niet kan worden geprogrammeerd, dan kwalitatief testmembraan vloeibaarheid als volgt.
- Sluit het velddiafragma een kleine grootte (~ 40 urn diameter) en stel de 488 nm excitatie lichtintensiteit door de software (20-80 uW of 15-60 nW / um 2) de NBD-PE fluorescentie bleken de belichte gebied significant, maar niet volledig. Voor een fluïdum bilaag, bij steady state, de fluorescentie-intensiteit in het midden van het blootgestelde gebied moet lager dan aan de randen, als intacte NBD-PE moleculen Voer het blootgestelde gebied en diffunderen een zekere afstand voor het bleken. Wanneer daarentegen het oppervlak S gehechtUV's niet barsten of andere reden de ondersteunde bilaag niet vloeibaar moeten alle fluoroforen op het blootgestelde oppervlak bleken.
Opmerking: Laser intensiteitswaarden in dit en latere stappen zijn bedoeld als ruwe uitgangspunt en moeten worden geoptimaliseerd voor een gegeven stel omstandigheden. - Stop de verlichting en start opnieuw enkele minuten later de resultaten van de steady state metingen verifiëren
- Sluit het velddiafragma een kleine grootte (~ 40 urn diameter) en stel de 488 nm excitatie lichtintensiteit door de software (20-80 uW of 15-60 nW / um 2) de NBD-PE fluorescentie bleken de belichte gebied significant, maar niet volledig. Voor een fluïdum bilaag, bij steady state, de fluorescentie-intensiteit in het midden van het blootgestelde gebied moet lager dan aan de randen, als intacte NBD-PE moleculen Voer het blootgestelde gebied en diffunderen een zekere afstand voor het bleken. Wanneer daarentegen het oppervlak S gehechtUV's niet barsten of andere reden de ondersteunde bilaag niet vloeibaar moeten alle fluoroforen op het blootgestelde oppervlak bleken.
- Programmeer een FRAP sequentie voor een meer kwantitatieve meting, indien mogelijk. Zie aanvullende bestanden en de bijbehorende legenda voor details.
Let op: soms SUV zal hechten aan het glas dekglaasje, maar slagen er niet te barsten en vormen een vloeistof bilaag. Als dit gebeurt, spoel de kanalen met ontgast reconstitutie buffer die 10 mM Mg 2+ naar ondersteunde dubbellaagsformatie helpen. Gebruik fluorescentie bleken om tweelaags vloeibaarheid beoordelen als in 4.2. Zodra een vloeistof dubbellaag wordt gevormd, spoelen met Mg 2+ -vrij Reconstitution buffer.
- Als een specifieke FRAP eenheid niet beschikbaar is, of als een FRAP sequentie niet kan worden geprogrammeerd, dan kwalitatief testmembraan vloeibaarheid als volgt.
- Invoering v-SUV's in de microfluïdische stromingskanalen
- Ontgas reconstitutie buffer en hiermee de v-SUV voorraadoplossing met een factor van ongeveer 10 3 5 verdunnen tot 10, afhankelijk van de v-SUV voorraadconcentratie. Doel voor een verdunning die resulteert in ongeveer 10-100 fusiegebeurtenissen binnen 60 seconden in het gezichtsveld.
Opmerking: Te veel fusies verhoging van de achtergrond fluorescentie (aangezien elk event deposito's LR-PE of heb lipide etiketten in de SBL) en zorg detectie en analyse van fusie gebeurtenissen moeilijk. In tegenstelling tot een fusie tarief dat te laag is resulteert in een slechte statistieken of vereist de verwerving van veel meer films. Voor een v-SUV concentratie van 0,1 mM lipide, te beginnen door het verdunnen van 5 ul SUV leverbaar in 995 ul reconstitutie buffer en verdun vervolgens 5-50 ul van deze in 950-995 pi reconstitutie buffer. - Laat de temperatuur evenwicht voor het stoppen van de stroom en inserTing de inlaat buis in de verdunde v-SUV-oplossing.
- Pas de TIRF hoek en polarisatie als volgt.
- Nadat de gewenste polarisatie door rotatie van de excitatiebundel Pas de kanteling van de spiegel bevelen de positie van de bundel via een stappenmotor via de software. Verplaats de positie van de laserstraal van het centrum van het objectief aan de achterkant brandvlak opzij excentrisch. Observeren het licht van het doel aan de voorzijde komen met een toenemende hoek ten opzichte van de as als doel de positie verder wordt bewogen excentrisch.
- Let op de motorpositie wanneer de uittredende bundel eerste verdwijnt in het doel, dat wil zeggen, wanneer TIR eerder wordt behaald.
- houden langzaam verder bewegen van de bundel positie uit het midden, terwijl het bewaken van de fluorescentie van het oppervlak. Let op de motor positie wanneer het oppervlak fluorescentie verdwijnt wanneer de bundel te ver buiten het centrum is verhuisd.
- Kies een balk positio n tussen de twee bovengenoemde grenswaarden vastgesteld. Voor de beste signaal-ruisverhouding en signaalversterking bij fusie Kies een ondiepe penetratiediepte (straalpositie dichter bij de rand van het doel) die nog steeds zorgt voor een gelijkmatige verlichting van de Viewfield.
Opmerking: Het beste is om op dezelfde TIR balk positie (zelfde indringdiepte) voor alle experimenten te houden nadat de instellingen zijn geoptimaliseerd. Controleer het roteren van de polarisatie niet tot veranderingen van straalpositie.
- Flow-v SUV in het kanaal met een stroomsnelheid van 2 ul / min, overeenkomend met een gemiddelde lineaire stroomsnelheid van ~ 1,5 mm / sec tot een doorsnede van 75 urn x 300 um. Overschakelen naar de excitatie / emissie-instellingen om lipide mengen monitor (LR of alleen hebben).
- Ontgas reconstitutie buffer en hiermee de v-SUV voorraadoplossing met een factor van ongeveer 10 3 5 verdunnen tot 10, afhankelijk van de v-SUV voorraadconcentratie. Doel voor een verdunning die resulteert in ongeveer 10-100 fusiegebeurtenissen binnen 60 seconden in het gezichtsveld.
- Observeren fusie tussen enkel v-SUV's en de SBL
.jpg "/>
Figuur 3. De experimentele opstelling pTIRF. (A) Schematische weergave van een v-SUV en een t-SBL op een glassubstraat. Kunstmatige kleuren TIRFM afbeeldingen één SUV-SBL samensmelting tonen lipide docking (1) en afgifte van lipide kleurstof in de SBL (2) gevolgd door bleken en afname van de fluorescentie-intensiteit (3). De totale fluorescentie-intensiteit (som van de pixelwaarden in de 5.3 micrometer x 5,3 micrometer doos) signaal wordt getoond. (B) Het dekglaasje gebonden aan het PDMS blok geplakt op de verwarmde podium. Inlaatbuis voor microfluïdische kanalen monsters van de buis in metalen monsterhouder (rechts) opgezogen door de injectiepomp (links). Onder de pomp is de dubbele emissie unit. Klik hier om een grotere versie van deze figuur te bekijken.
oad / 54349 / 54349fig4.jpg "/>
Figuur 4. Schematische voorstelling van de experimentele opstelling. De uitdovende golf ontstaat op het glas-buffer interface in de microfluïde kanaal. De SBL is gevormd op het glas en v-SUV aangezogen uit de metalen monsterhouder (rechtsboven) door het kanaal in de spuit (linksboven). M, spiegel; DM, dichroïsche spiegel; L, lens; F, filter; P, polarisator. Klik hier om een grotere versie van deze figuur te bekijken.
- Voortdurend prikkelen en te bewaken v-SUV fluorescentie met behulp van een 561 nm (voor LR-PE) of 638 nm (voor deed) laser, afhankelijk van de fluorofoor opgenomen in v-SUV's. Als v-SUV's het stromingskanaal en dock op en de zekering te bereiken met de SBL, de achtergrond fluorescentie signaal begint op te stapelen.
- Stel de excitatie-intensiteit laser door de software continu bleken de achtergrondfluorescentie, zodanig dat bij steady state nieuwe docking en fusion gebeurtenissen kunnen gemakkelijk worden waargenomen.
Opmerking: Als het bleken te langzaam is, zal de achtergrond fluorescentie te hoog zijn. Als bleken te snel zullen signalen van aangemeerd SUV's en van fluoroforen vrijkomt in de SBL snel verdwijnen, verkorten het venster waarin fusie van aangemeerd SUV kan worden gedetecteerd of een fluorofoor kan worden gevolgd. LR-PE geëxciteerd bij 561 nm, 2,5-7,5 mW verlichten van een cirkel 190 urn diameter (100-250 nW / um 2) is een redelijke waarde te starten. Voor 638 nm excitatie van deed, 0,8-1,6 mW macht over een 190 micrometer cirkel met een diameter (30-60 nW / um 2) kan worden gebruikt voor de eerste tests. - Acquire meerdere films op verschillende posities in een bepaald kanaal. Controleer NBD-PE fluorescentie te controleren of de SBL geen defecten op deze posities heeft. Acquire full-frame films met de maximale snelheid (~ 50 frames / sec) of een bijgesneden gebied van belang bij hogere framesnelheid (tot~ 100 Hz) gedurende 60 sec.
- Ga naar een andere microfluïdische kanaal en herhaal de opnames voor andere aandoeningen. Onder negatieve controles zoals een eiwitvrij SBL of SUV, voeg de oplosbare cytoplasmatische domein van de v-SNARE VAMP2 (CDV) als een remmer of behandelt v-SUV met tetanus neurotoxine (30 min bij 37 ° C) in één of meer van de kanalen op dezelfde dekglaasje.
- Opruimen en PDMS recycling
- Om de PDMS blok hergebruiken, spoel de microkanalen met -200 pl 70% ethanol met een stroomsnelheid van 5 pl / min. Tot slot, zuigen lucht door de buizen en kanalen.
- Maak de PDMS blokkeren voorzichtig van het dekglaasje door lichtjes te knijpen. Plaats het op een schoon stuk aluminiumfolie. Bewaar in een vacuümexsiccator. Het kan meerdere keren worden hergebruikt.
- Voor een grondige reiniging spoel de kanalen met een detergent of natriumhydroxide oplossing vóór de 70% ethanol. Als alternatief, verwijder alle buizen weer m de ultrasone trillingen PDMS en het blok voor 30 minuten in isopropanol, voor het drogen en het invoegen van nieuwe leidingen.
5. SUV-SBL Fusion Analyse Monitor Lipid en Content kort tegelijk
- Voor tweekleurige monitoring van gelijktijdige lipiden en oplosbare inhoud vrij te geven, volgt u dezelfde stappen als in hoofdstuk 4, met uitzondering van het gebruik liposomen gemerkt met zowel een lipide (DID) en een oplosbare inhoud (SRB) label, zoals uitgelegd in hoofdstuk 3 SRB is ingekapseld bij 10 mM en aanvankelijk zeer zichzelf gedoofd.
- Met de configuratie schematisch weergegeven in figuur 4, wekken SRB en deed fluorescentie gelijktijdig gebruikt 561 nm en 638 nm lasers respectievelijk. Een dichroïsche spiegel (640 nm) splitst de emissie in twee bundels die worden uitgevoerd door middel van een korte (595/50 nm) en een lange (700/75 nm) golflengte filter om de SRB detecteren en deed uitstoot, respectievelijk. De twee emissie-bundels worden geprojecteerd side-by-side op de EM-CCD-chip.
- Analyse van de FRAP gegevens
- Gebruik de MATLAB-programma voorzien in de aanvullende informatie aan de lipide diffusiecoëfficiënt, D te schatten. Het programma leest een lijst van OME-TIFF bestanden 49, detecteert het gebleekte gebied plot de gemiddelde pixelwaarden in het gebleekte gebied als functie van de tijd, en past de resulterende herstelcurve een model van Soumpasis 50 om de hersteltijd te extraheren,
 , Waarbij w de straal van de cirkel gebleekt.
, Waarbij w de straal van de cirkel gebleekt.
Opmerking: Een MATLAB programma werd eerder 25 voor de analyse van FRAP films met behulp van Nikon ND2 bestanden voorzien. Het actuele programma leest OME-TIFF bestanden 49, omdat de meeste formaten gemakkelijk kan worden omgezet OME-TIFF. Een kwantitatieve analyse van FRAP gegevens eenvoudigste en meest nauwkeurig wanneer bleken instantaan de bleached regio is een cirkel, en bleken tijdens de uitlezing is te verwaarlozen. Hoewel deze criteria niet strikt in de eenvoudige FRAP metingen hier beschreven wordt voldaan, kan een redelijke schatting van de diffusiecoëfficiënt worden verkregen. Voor een meer nauwkeurige schatting, gebruik dan het volgen van enkele lipide kleurstoffen (paragraaf 6.3).
- Gebruik de MATLAB-programma voorzien in de aanvullende informatie aan de lipide diffusiecoëfficiënt, D te schatten. Het programma leest een lijst van OME-TIFF bestanden 49, detecteert het gebleekte gebied plot de gemiddelde pixelwaarden in het gebleekte gebied als functie van de tijd, en past de resulterende herstelcurve een model van Soumpasis 50 om de hersteltijd te extraheren,
- Analyse van docking rate, fusion rate en docking-to-fusion vertragingstijden
- Gebruik ImageJ om een film te openen om te analyseren en aan te passen helderheid en het contrast om duidelijk te identificeren docking en fuseren gebeurtenissen. Start de SpeckleTrackerJ 26 plugin. Zie Smith et al. 26 en online documentatie voor SpeckleTrackerJ voor instructies voor gebruik.
- Identificeer alle nieuw aangemeerd SUV in SpeckleTrackerJ. Om SUV's die nauw dock van degenen die stuiteren de SBL te onderscheiden, opleggen van een minimum docking duur van een paar frames. Sla de tracks, en herhaal voor alle films.
Opmerking: Voor de dockoning rate, het enige dat telt is om te bepalen wanneer een SUV gedokt, dus alleen het eerste frame waarin de SUV's gedokt moet worden opgenomen in deze tracks. De rest van de tracks zullen niet in aanmerking worden genomen in de analyse. Echter, auto-het bijhouden van elke SUV totdat het bleekt of zekeringen helpt merk SUV's die al zijn gevolgd. - Identificeer alle fusing blaasjes. Hiervoor moeten de sporen omvatten alle frames van het eerste frame een SUV aangemeerd tot het eerste frame waarin fusie duidelijk door een plotselinge toename van de fluorescentie van de vlek gevolgd. De duur van deze sporen worden gebruikt voor het docking-to-fusie vertragingen te berekenen. Sla alle tracks. Herhaal dit voor alle films.
- Voor batch analyse van docking of fusie van gegevens, een lijst opstellen van het traject bestanden uit de betreffende films en voer de MATLAB's die bij de Karatekin en Rothman 25. Aanwijzingen die samen worden voorzien van het programma.
Opmerking: De programma's plot van de cumulative docking en fusiegebeurtenissen als functie van de tijd, op basis van gegevens uit het traject bestanden. Docking en fusion tarieven zijn geschat op basis van de hellingen van deze percelen. Voor fusie gegevens wordt het docking-to-fusie vertragingen ook berekend en de verdeling uitgezet als survival plot, dwz de kans dat fusie nog niet heeft plaatsgevonden op een bepaalde vertraging na het koppelen.
- Lipid diffusiviteit
- Voor elke samensmelting, volgen enkele fluorescente lipiden mocht waarneembaar worden wanneer zij voldoende afstand van het fusie (figuur 6) zijn verspreid. Gebruik SpeckleTrackerJ voor het bijhouden, en sla de tracks voor verdere analyse met behulp van MATLAB. Afhankelijk van de grootte van de vesicle fusing, kenmerkend 3-30 enkele fluoroforen kunnen worden gevolgd. Omdat meer titels wenselijk voor het berekenen van
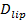 , Proberen om enkele moleculen zo lang bijhoudenmogelijk handmatig gecorrigeerd trajecten indien nodig.
, Proberen om enkele moleculen zo lang bijhoudenmogelijk handmatig gecorrigeerd trajecten indien nodig. - Bereken het gemiddelde kwadraat verplaatsing (MSD) voor enkele lipide marker trajecten die> 40-50 frames (ongeveer ~ 1,5 sec) duren. Gebruik de MSD aan de lipide diffusiecoëfficiënt berekenen, . Zie Smith et al. 26 en Stratton et al. 27 voor meer informatie.
- Voor elke samensmelting, volgen enkele fluorescente lipiden mocht waarneembaar worden wanneer zij voldoende afstand van het fusie (figuur 6) zijn verspreid. Gebruik SpeckleTrackerJ voor het bijhouden, en sla de tracks voor verdere analyse met behulp van MATLAB. Afhankelijk van de grootte van de vesicle fusing, kenmerkend 3-30 enkele fluoroforen kunnen worden gevolgd. Omdat meer titels wenselijk voor het berekenen van
- Single lipide kleurstof intensiteit, SUV-SBL intensiteit reductie factor, en blaasjes grootte
- Meet de som van de pixelwaarden van een lipide marker in een 3 x 3 pixels (0,8 um x 0,8 um) rondom de markering voor ~ 15 frames voor en na de markering bleekmiddelen in een enkele stap. Aftrekken van de achtergrondintensiteit gemiddeld over de post-bleekmiddel kaders van de pre-bleach gemiddelde intensiteit voor het bleken van de intensiteit van het gevolgde lipide marker verkrijgen. Herhaal de meting voor zoveel merkers als praktisch voor een bepaalde film.
- Zet de distributie van enkele lipide label intensiteiten,
 , En breng een Gauss naar het gemiddelde te schatten.
, En breng een Gauss naar het gemiddelde te schatten. - Bereken de vertraging tussen het moment fusie zich voordoet en de baan van een enkele lipide merker bezit in dat geval eindigde in een enkele stap bleken. Het verkrijgen van het bleken tijd voor een fluorofoor in de SBL,
 , Door het uitzetten van de overlevende functie van de vertragingen en passend bij een exponentiële.
, Door het uitzetten van de overlevende functie van de vertragingen en passend bij een exponentiële.
Let op: Omdat de gepolariseerde excitatie veld koppels meer zwak om de fluoroforen op een SUV, het bleken tijd van een SUV is meestal veel langzamer 27. - Schatting
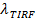 De intensiteit reductiefactor voor een lipide kleurstof wanneer de SUV opzichte wanneer het in de SBL na Stratton 27 (tp_upload / 54349 / 54349eq8.jpg "/> is de intensiteit van een kleurstof in de SUV).
De intensiteit reductiefactor voor een lipide kleurstof wanneer de SUV opzichte wanneer het in de SBL na Stratton 27 (tp_upload / 54349 / 54349eq8.jpg "/> is de intensiteit van een kleurstof in de SUV).
Let op: Als het bleken verwaarloosbaar waren, gelijk aan de gekoppelde intensiteit zou  (punt (1) in figuur 3A, rechter paneel), gedeeld door de totale intensiteit bereikt na alle fluoroforen zijn bij fusie gestort in het SBL (stippellijn gelabeld
(punt (1) in figuur 3A, rechter paneel), gedeeld door de totale intensiteit bereikt na alle fluoroforen zijn bij fusie gestort in het SBL (stippellijn gelabeld  in figuur 3). Vanwege de snelle bleking in de SBL, is meestal niet bereikt en een nauwkeurige schatting van vergt het aanbrengen van de release kinetiek om een uitdrukking die 27 incl udes beide (Zie 6.4.3) en . Aparte uitdrukkingen worden verschaft in Stratton et al 27 voor de gevallen van poriën beperkt en verspreiding beperkte release kinetiek.; de beste fit geval varieert van evenement naar evenement (zie 6.5.1 voor het geval van poriën beperkt kinetiek).
in figuur 3). Vanwege de snelle bleking in de SBL, is meestal niet bereikt en een nauwkeurige schatting van vergt het aanbrengen van de release kinetiek om een uitdrukking die 27 incl udes beide (Zie 6.4.3) en . Aparte uitdrukkingen worden verschaft in Stratton et al 27 voor de gevallen van poriën beperkt en verspreiding beperkte release kinetiek.; de beste fit geval varieert van evenement naar evenement (zie 6.5.1 voor het geval van poriën beperkt kinetiek). - Bereken het blaasje gebied voor afzonderlijke gebeurtenissen van 27
 waar is de aangemeerd SUV intensiteit, is de gemiddelde intensiteit van een enkele lipide kleurstof in de SBL, de intensiteit reductiefactor voor een lipide kleurstof wanneer het in de SUV opzichte wanneer het in de SBL enatie 1 "src =" / files / ftp_upload / 54349 / 54349eq12.jpg "/> is de bekende oppervlaktedichtheid van lipide kleurstoffen.
waar is de aangemeerd SUV intensiteit, is de gemiddelde intensiteit van een enkele lipide kleurstof in de SBL, de intensiteit reductiefactor voor een lipide kleurstof wanneer het in de SUV opzichte wanneer het in de SBL enatie 1 "src =" / files / ftp_upload / 54349 / 54349eq12.jpg "/> is de bekende oppervlaktedichtheid van lipide kleurstoffen.
- Fusion porie eigenschappen
- Ervan uitgaande dat release is porie beperkt is, passen de lipide label vrijgavekinetiek tot 27
 waar
waar  is aangemeerd SUV intensiteit vlak voor fusie en de overige parameters zijn zoals eerder gedefinieerd. Gebruik de waarde van verkregen in 6.4.3 als een vaste parameter, en haal de beste pasvorm ramingen voor en
is aangemeerd SUV intensiteit vlak voor fusie en de overige parameters zijn zoals eerder gedefinieerd. Gebruik de waarde van verkregen in 6.4.3 als een vaste parameter, en haal de beste pasvorm ramingen voor en  .
. - Schat de fractie van de tijd de porie open is, P 0, in de veronderstelling vertraging van lipide release is te wijten aan porie flikkeren 27:60;
 =
=  , Waarbij A ves is het blaasje gebied (sectie 6.4.5), b de poriën hoogte (meestal achterwege blijven ~ 15 nm), RP ≈ 3 nm is de effectieve poriestraal zoals gezien door de diffunderende lipide labels en omvat half de bilaag dikte (~ 2 nm), is de lipide diffusiviteit (berekend 6,3), en is de tijd voor de lipiden worden vrijgegeven uit de SUV in de SBL (van 6.5.1).
, Waarbij A ves is het blaasje gebied (sectie 6.4.5), b de poriën hoogte (meestal achterwege blijven ~ 15 nm), RP ≈ 3 nm is de effectieve poriestraal zoals gezien door de diffunderende lipide labels en omvat half de bilaag dikte (~ 2 nm), is de lipide diffusiviteit (berekend 6,3), en is de tijd voor de lipiden worden vrijgegeven uit de SUV in de SBL (van 6.5.1). - Om te bevestigen dat een nominale P 0> 1 geeft een volledig open poriën, P 0 = 1, past de intensiteit tijdsverloop aan vergelijking 4 van Stratton et al. 27, de voorspelde kinetiek voor een permanent open poriën. Deze fit moet beter zijn dan fittin zijng de expressie in 6.5.1 voor een permanent open poriën.
- Ervan uitgaande dat release is porie beperkt is, passen de lipide label vrijgavekinetiek tot 27
 , Waarbij w de straal van de cirkel gebleekt.
, Waarbij w de straal van de cirkel gebleekt.  , Door het uitzetten van de overlevende functie van de vertragingen en passend bij een exponentiële.
, Door het uitzetten van de overlevende functie van de vertragingen en passend bij een exponentiële.  (punt (1) in figuur 3A, rechter paneel), gedeeld door de totale intensiteit bereikt na alle fluoroforen zijn bij fusie gestort in het SBL (stippellijn gelabeld
(punt (1) in figuur 3A, rechter paneel), gedeeld door de totale intensiteit bereikt na alle fluoroforen zijn bij fusie gestort in het SBL (stippellijn gelabeld  in figuur 3). Vanwege de snelle bleking in de SBL,
in figuur 3). Vanwege de snelle bleking in de SBL,  waar
waar  waar
waar  is aangemeerd SUV intensiteit vlak voor fusie en de overige parameters zijn zoals eerder gedefinieerd. Gebruik de waarde van
is aangemeerd SUV intensiteit vlak voor fusie en de overige parameters zijn zoals eerder gedefinieerd. Gebruik de waarde van  =
=  , Waarbij A ves is het blaasje gebied (sectie 6.4.5), b de poriën hoogte (meestal achterwege blijven ~ 15 nm), RP ≈ 3 nm is de effectieve poriestraal zoals gezien door de diffunderende lipide labels en omvat half de bilaag dikte (~ 2 nm),
, Waarbij A ves is het blaasje gebied (sectie 6.4.5), b de poriën hoogte (meestal achterwege blijven ~ 15 nm), RP ≈ 3 nm is de effectieve poriestraal zoals gezien door de diffunderende lipide labels en omvat half de bilaag dikte (~ 2 nm), Subscription Required. Please recommend JoVE to your librarian.
Representative Results
SBL Kwaliteit
Het is cruciaal om de kwaliteit en de vloeibaarheid van de SBL vóór het fusie-experiment controleren. De fluorescentie onderaan glaszijde van een microfluïde kanaal moet uniform zijn, zonder duidelijke gebreken. Als een luchtbel passeert hoewel het kanaal, laat meestal zichtbare littekens op de SBL. Als er zulke grote schaal littekens / defecten, geen gebruik maken van dat kanaal. Soms SUV's kan hechten aan de ondergrond, maar niet te barsten en vormen een continue SBL. Als dat het geval is, kan de fluorescentie nog blijken zeer uniform, maar niet herstellen na bleken een klein gebied. De invoering van 10 mM Mg 2+ vaak helpt bij het oplossen van dit probleem. Andere middelen kunnen worden geïntroduceerd om te helpen het barsten van de SUV's, zoals andere divalente kationen 51, een polymeeroplossing 52 of osmotische shock 25.
Temperatuurveranderingen resulteren in veranderingen van SBL gebied, omdat de gemiddelde oppervlakte ingenomen door een lipide temperatuurafhankelijk. De temperatuur te verhogen of te verlagen met 5-10 ° C of meer kan leiden tot overmatige gebied gaan in buizen geëxtrudeerd uit het oppervlak of donker, SBL-vrijkomen die op het oppervlak, respectievelijk 53. Het is daarom belangrijk om temperatuurschommelingen tot een minimum te houden.
figuur 5 voor een SBL die relatief foutvrije en vocht was.

Figuur 5. FRAP om SBL vloeibaarheid te onderzoeken. Een opeenvolging van beelden met een verlichte gebied begrensd door de gesloten veld diafragma (diameter 49 pm) vóór (eerste frame) en na het bleken met een sterke lichtpuls. Het gebied wordt afgebeeld bij lage blootstelling licht om de fluorescentie herstel door diffusie van NBD volgen gelabelde lipiden. De grafiek toont de genormaliseerde fluorescentie als een functie van de tijd voor een aantal kanalen van verschillende dekglaasjes, waarbij de hoge mate van reproduceerbaarheid van de werkwijze. De lipidesamenstelling van de t-SBL is POPC: SAPE: DOPS: Cholesterol: hersenen PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5 mol% ) en LP = 20.000. Het bleekmiddel was 984,203 μm 3. De hersteltijd en de diffusiecoëfficiënt berekend uit een fit (zie 6.1) waren τ = 67,3 sec en D = 1,99 micrometer 2 / sec, respectievelijk. Klik hier om een grotere versie van deze figuur te bekijken.
Single-kleur Detectie van Docking en Fusion Events Met behulp van een Lipid Label
Handmatige telling van docking en fusion evenementen is een vervelende taak als 100-400 events per conditie zijn nodig voor goede statistieken. Omdat SUV's kan bewegen op de SBL voordat fuseren, is een tracking-software nodig. SpeckleTrackerJ 26 plugin voor ImageJ heeft functies die speciaal voor het bijhouden van SUV's voor de analyse van docking en fusion evenementen werden ontwikkeld. Vaak slechts een fractie van gedokt blaasjes uiteindelijk fusing;op het docking snelheid veel meer SUV's moeten worden bijgehouden dan die fuseren of een gedeelte van de film moet worden geanalyseerd totdat een voldoende aantal gebeurtenissen waargenomen gedurende een betrouwbare schatting te berekenen. Als de belangrijkste focus is de fusie te beoordelen en / of het docking-to-fusion vertragingen, alleen het fuseren van SUV's moeten worden geïdentificeerd en opgespoord.
Na om de kwaliteit van de SBL zijn SUV ingebracht in de kanalen en de fluorescentie wordt continu opgewekt onder gebruik van TIR s-gepolariseerd licht. Voor de eerste experimenten, kan de invalshoek van de TIR excitatiebundel moeten worden aangepast om af te stemmen de penetratiediepte van de uitdovende golf verkregen. Een vertegenwoordiger enkele SUV docking en fusion event is weergegeven in figuur 6A. Het blaasje aangemeerd tijdens het frame gemarkeerde D en fusie begon in frame F. Ongeveer 1,3 sec later (frame S) enkele lipiden werden waarneembaar als ze verspreid uit de buurt van de fusie zittene. De totale fluorescentie-intensiteit (som van pixelwaarden) in elk vak is uitgezet als functie van de tijd in figuur 6B, waarbij de frames waar docking en fusion opgetreden aangegeven. Het cumulatieve aantal fusiegebeurtenissen als functie van de tijd is uitgezet in Figuur 6C, vijf verschillende films onder toepassing van t-SNARE gereconstitueerd SBLs (t-SBL, LP = 20.000, ongeveer 70 t-SNAREs per sq. Micron). De fusie vanaf elke film werd berekend uit een lineaire aanpassing van de helling. Dit gemiddelde percentage fusion ± SEM wordt getoond als een staafdiagram in paneel D. De docking-to-fusie vertraging van het in figuur 6A, B, voorbeeld was 0,07 sec. De verdeling van vertragingen eveneens berekend voor een totaal van 158 gebeurtenissen wordt getoond in figuur 6D als nagelaten plot, dat wil zeggen, de waarschijnlijkheid dat een gekoppelde blaasje nog niet is gefuseerd met een bepaalde vertraging. Ongeveer 1/2 van deze blaasjes gefuseerd binnen 100 msec. Met het verhogen van het cholesterolgehalte, de distribution van docking-to-fusion vertragingen verkort, hoewel lipide diffusie vertraagt 27.
Het is belangrijk om controleproeven parallel. Op dezelfde dekglaasje waarvan de gegevens in Figuur 6B, D werden verkregen, werden controle omstandigheden getest in aangrenzende kanalen. In één kanaal heeft de SBL geen enkele t-SNAREs. Van de 5 films duren 60 seconden per stuk, waren slechts 7 fusie gebeurtenissen geregistreerd in vergelijking met 158 als de t-SNAREs werden opgenomen (figuur 6C). In een ander kanaal, de oplosbare cytoplasmatische domein van de v-SNARE VAMP2 (CDV) werd opgenomen. CDV bindt de t-SNAREs de SBL, waardoor binding van de volledige lengte VAMP2 die op de SUV. In vijf 60 sec films werden geen fusiegebeurtenissen gedetecteerd in de aanwezigheid van CDV (figuur 6C).

Figuur 6. SUV-SBL docking en fusion evenementen. (A) Volgorde van beelden van een v-SUV fusie evenement met de t-SBL afgebeeld door pTIRF microscopie na de verspreiding van LR-PE. Beelden worden genomen om de 18 msec, slechts om de tweede frame wordt getoond. D en F markeren het begin van docking en fusion respectievelijk. Individuele lipide labels worden onderkend ze weg diffunderen van de fusieplaats en onderling (S). Frame afmetingen: 16,7 urn x 16,7 urn. (B) de fluorescentie-intensiteit als functie van de tijd voor de samensmelting getoond in (A). (C) Cumulatief aantal fusiegebeurtenissen als functie van de tijd voor v-SUV gelabeld met WIsT van een andere reeks experimenten. (D) de genormaliseerde fusie percentage (gemiddelde ± SEM, sx um x 2 pM SUV) -1, uitgaande van 2 x 10 4 lipiden per SUV en (E) de overlevingskans voor een bepaalde tijd na het koppelen van de vesicle. De samenstelling voor SBLs waren POPC: SAPE: DOPS: Cholesterol: hersenen PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5 mol%) en LP = 20.000. v-SUV's waren samengesteld uit POPC: SAPE: DOPS: Cholesterol: PEG2000-DOPE: LR-PE (57,4: 15: 12: 10: 4,6: 1 mol%) en LP = 200 voor A en B. Voor CE, de v -SUV samenstelling was vergelijkbaar, met uitzondering deed was gebruikt in plaats van LR-PE. klik hier om een grotere versie van deze figuur te bekijken.
Lipid diffusiviteit, vesiclegrootte en Fusion Pore Properties
De single-molecule gevoeligheid van de assay maakt winning van verschillende parameters naast de docking en fusie tarieven en de docking-to-fusie vertraging distributies. Single, fluorescent gelabelde lipiden worden waarneembaar als ze weg te diffunderen uit de fusie (figuur 6A). Tracking teze direct biedt lipide diffusie 26, 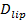 , En een digitale meting van het bleken tarief voor fluoroforen in de SBL 27. Zoals vermeld in de inleiding, de fluorescentie-emissie van de labels verschillen de SUV versus de SBL, als gevolg van drie factoren die algemeen moeilijk te ontwarren: (i) dequenching etiketten op fusie, door de oplossing wanneer overgebracht in de SBL (ii) een hogere excitatie-intensiteit in de SBL gevolg van het verval van het verdwijnende veld gegenereerd door TIR, en (iii) de oriëntatie van de fluorofoor transitiedipolen opzichte van de polarisatie van de activeringsbundel. De grootte van factor (ii) wordt bepaald door de relatieve afmetingen van het verdwijnende veld vervallengte en vesiclegrootte en verschilt per samensmelting door de dispersiteit SUV maten. Factor (iii) is afhankelijk van de fluorofoor en de polarisatie gebruikt. In plaats van te proberen te di sentangle deze effecten, kan de grootte van de fluorescentieverandering meten direct door de totale pixelwaarden in een gebied rond een SUV voor fusie en tenslotte fluoroforen zijn neergelegd in de volgende SBL fusie (figuur 3A (1) vs. ( 2)). In tegenstelling tot eerdere werk 29,44, moet de regio worden gekozen voldoende groot, zodat er geen fluoroforen zal gelaten hebben via diffusie voor de overwogen na fusie tijden. Het kiezen van een 30 x 30 pixelgebied (8 urn x 8 urn) voor analyse tot 1,6 sec na fusie doorgaans voldoende 27. Het negeren van het bleken voor het moment, de verhouding van
, En een digitale meting van het bleken tarief voor fluoroforen in de SBL 27. Zoals vermeld in de inleiding, de fluorescentie-emissie van de labels verschillen de SUV versus de SBL, als gevolg van drie factoren die algemeen moeilijk te ontwarren: (i) dequenching etiketten op fusie, door de oplossing wanneer overgebracht in de SBL (ii) een hogere excitatie-intensiteit in de SBL gevolg van het verval van het verdwijnende veld gegenereerd door TIR, en (iii) de oriëntatie van de fluorofoor transitiedipolen opzichte van de polarisatie van de activeringsbundel. De grootte van factor (ii) wordt bepaald door de relatieve afmetingen van het verdwijnende veld vervallengte en vesiclegrootte en verschilt per samensmelting door de dispersiteit SUV maten. Factor (iii) is afhankelijk van de fluorofoor en de polarisatie gebruikt. In plaats van te proberen te di sentangle deze effecten, kan de grootte van de fluorescentieverandering meten direct door de totale pixelwaarden in een gebied rond een SUV voor fusie en tenslotte fluoroforen zijn neergelegd in de volgende SBL fusie (figuur 3A (1) vs. ( 2)). In tegenstelling tot eerdere werk 29,44, moet de regio worden gekozen voldoende groot, zodat er geen fluoroforen zal gelaten hebben via diffusie voor de overwogen na fusie tijden. Het kiezen van een 30 x 30 pixelgebied (8 urn x 8 urn) voor analyse tot 1,6 sec na fusie doorgaans voldoende 27. Het negeren van het bleken voor het moment, de verhouding van  voor en na fusie geeft de fluorescentie-intensiteit reductiefactor
voor en na fusie geeft de fluorescentie-intensiteit reductiefactor 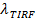 . De vesiclegrootte kan worden geschat aangezien de fluorescentie-intensiteit van een enkele label in de SBL (iles / ftp_upload / 54349 / 54349eq6.jpg "/>), de factor waarmee de intensiteit zou worden verlaagd als het ware in een SUV ( ), De bekende dichtheid van labels in de SUV, en het gebied bezet door een lipide. In de praktijk bleken is meestal snel genoeg dat tegen de tijd alle markeringen zijn aangebracht in de SBL, sommige reeds gebleekt. Daarom, om een nauwkeurige schatting, de fluorescentie-intensiteit reductiefactor En de lipide release tijd,
. De vesiclegrootte kan worden geschat aangezien de fluorescentie-intensiteit van een enkele label in de SBL (iles / ftp_upload / 54349 / 54349eq6.jpg "/>), de factor waarmee de intensiteit zou worden verlaagd als het ware in een SUV ( ), De bekende dichtheid van labels in de SUV, en het gebied bezet door een lipide. In de praktijk bleken is meestal snel genoeg dat tegen de tijd alle markeringen zijn aangebracht in de SBL, sommige reeds gebleekt. Daarom, om een nauwkeurige schatting, de fluorescentie-intensiteit reductiefactor En de lipide release tijd,  , Best geëxtraheerd uit een fit aan het tijdsverloop van de totale pixelwaarden die rekening houdt bleken 27. Het bleken tarief van labels in de SBL onafhankelijk geschat op basis van enkele lipide kleurstof tracking, of door het aanbrengen van een exponentiële aan de totale intensiteit profiel,
, Best geëxtraheerd uit een fit aan het tijdsverloop van de totale pixelwaarden die rekening houdt bleken 27. Het bleken tarief van labels in de SBL onafhankelijk geschat op basis van enkele lipide kleurstof tracking, of door het aanbrengen van een exponentiële aan de totale intensiteit profiel,  Bij lange tijden (bijvoorbeeld regime (3) in figuur 3). Het kennen van de grootte van consumeerbare vesikel en lipide diffusiviteit maakt vergelijking van de werkelijke lipide release tijd, , De diffusietijd van een lipide rond de vesicle
Bij lange tijden (bijvoorbeeld regime (3) in figuur 3). Het kennen van de grootte van consumeerbare vesikel en lipide diffusiviteit maakt vergelijking van de werkelijke lipide release tijd, , De diffusietijd van een lipide rond de vesicle  , Waarbij A ves is het blaasje gebied en is de lipide diffusiviteit. Als de twee tijdschalen vergelijkbaar, de poriën weer weinig weerstand tegen lipide afgifte en afgifte is diffusie beperkt. Als daarentegen
, Waarbij A ves is het blaasje gebied en is de lipide diffusiviteit. Als de twee tijdschalen vergelijkbaar, de poriën weer weinig weerstand tegen lipide afgifte en afgifte is diffusie beperkt. Als daarentegen  Dan de porie significant vertraagt lipide flux. Een kwantitatieve maat voor de vertraging is de "porie openheid", P 0, gelijk aan de verhouding
Dan de porie significant vertraagt lipide flux. Een kwantitatieve maat voor de vertraging is de "porie openheid", P 0, gelijk aan de verhouding  keren een factor of Om eenheid met betrekking tot de geometrie 27 porie. Voor een twee-staten, open / gesloten poriën, P 0 is de fractie van de tijd in de open toestand. Voor een flikkerende poriën waarvan de grootte varieert continu, p 0 is de tijd gemiddelde radius ten opzichte van de volledig geopende radius.
keren een factor of Om eenheid met betrekking tot de geometrie 27 porie. Voor een twee-staten, open / gesloten poriën, P 0 is de fractie van de tijd in de open toestand. Voor een flikkerende poriën waarvan de grootte varieert continu, p 0 is de tijd gemiddelde radius ten opzichte van de volledig geopende radius.
De signaal-ruisverhouding heeft, voldoende voor het volgen van één lipide fluoroforen zijn. Dit wordt gemakkelijker bereikt in één kleur experimenten, met behulp van een heldere merker zoals LR-PE.
Gelijktijdige detectie van Lipid en oplosbare Cargo release
Een representatief geval is getoond in figuur 7. Voor deze hoge concentraties voor inkapseling, de SRB fluorescentie aanvankelijk zichzelf gedoofd. Daarom zijn de meeste aangemeerd v-SUV's zijn alleen aantoonbaar door hun DiD signaal 27. Een scherpe stijgingin DiD fluorescentie markeert lipide mengen, zoals in één kleur lipide-label experimenten. Het begin van lipide afgifte viel samen met het begin van een toename van fluorescentie SRB. Deze toename was waarschijnlijk te wijten aan SRB dequenching de SRB moleculen ontsnapt aan de SUV en de resterende SRB werd verdund tot onder zelfdovende concentraties. Interessant is dat de SRB fluorescentie bereikte een stabiel plateau. Indien het fusie porie geopend blijven, zou men verwachten dat de SRB signaal stijgen tot een maximum wegens dequenching, gevolgd door een afname tot achtergrond als de resterende SRB verliet de SUV, of indien de SUV in de SBL ingestort. De stabiele plateau geeft waarschijnlijk de poriën opnieuw afgesloten na het loslaten alle lipide labels en een deel van de oplosbare lading.
In sommige gevallen werd het lipide vrijgavesignaal niet gepaard met detecteerbare signalen inhoud ontwijkt. Dit kan hetzij omdat de poriën te klein zijn om doorgang van SRB mogelijk waren, of eenhoe de SUV al de oplosbare lading had verloren. Om onderscheid te maken tussen deze twee mogelijkheden, kan bleken van gedokt SUV's worden geanalyseerd. Bleken van het lipide label WIsT vermindert de fluorescentiesignaal binnen enkele seconden, terwijl het bleken van de eerste self-afgeschrikt SRB leidt tot een toename van de SRB signaal als een fractie van de SRB zijn foto alternatieve productie en zelf-uitdoving wordt ontlast. Terwijl de meeste aangemeerd SUV's (48 van de 58 geanalyseerde evenementen) verschijnt de verwachte stijging van de SRB fluorescentie gedurende langere perioden van verlichting, heeft een kleine fractie (10/58) geen SRB signalen zien at all. Aldus kan een kleine fractie van de SUV gehalte aan oplosbare tijdens de periode tussen hun bereiding en toepassing in de experimenten verliezen.
In meer dan 80% van de gebeurtenissen (74 van 91), waarvoor zowel SRB en deed signalen kunnen worden gevolgd, de SRB fluorescentie toegenomen en bleef op een stabiel plateau 27. In ~ 20% van de tHese gevallen de resterende SRB fluorescentie werd plotseling vrijgelaten tot een paar seconden later. Dit kan ofwel door een heropening van de porie die snelle, volledige release binnen 1-2 frames (18-36 msec) of barsten van de SUV door geaccumuleerde foto-schade 16. Ongeacht de herkomst, tweede, snelle afgifte van SRB uitsluit dat het overblijvende SRB signaal na de eerste release kan het gevolg zijn van de SRB blijft ingevangen in de ruimte tussen de SBL en het glassubstraat na volledige release. In een eerdere studie waarin de SBL direct op glas werd ondersteund, met weinig ruimte tussen de twee 16 is een dergelijk mechanisme voorgesteld. Daarentegen moet de gepegyleerde lipiden gebruikt in deze assay 4-5 nm ruimte tussen de SBL en het glas 25 te brengen, zodat SRB weg van de fusieplaats diffunderen.

Figuur 7. Simultan exportgerichte bedrijven lipiden en content release. Fluorescentie signalen van lipiden (blauw) en de inhoud (rode) marker voor een SUV-SBL fusie event (voor details zie tekst). Snapshots van elk kanaal worden getoond aan de onderkant (4,0 micrometer breed). Cartoons (midden) illustreren de verschillende fasen. (1) SUV dokken, had fluorescentie is laag. (2) Fusion maakt overdracht van DiD in de SBL en deed fluorescentie toeneemt. SRB signaal verhoogt ook als zelf-uitdoving wordt verlicht door ontsnapping van SRB door de porie. (3) SRB signaal blijft stabiel, terwijl DiD diffunderen binnen de SBL wijst op een mogelijk opnieuw sluiten van de porie. (4) SRB signalen te verminderen zeer snel, als gevolg van een snelle volledige fusie of liposoom barsten. De samenstelling van v-SUV was POPC: SAPE: DOPS: PEG2000-DOPE: Heb (67: 15: 12: 5: 1 mol%), LP 200, terwijl de t-SBLs waren samengesteld uit POPC: SAPE: DOPS: hersenen PI (4,5) P2: PEG2000-DOPE: NBD-PE (64,5: 15: 12: 3: 5: 0,5 mol%) en LP = 20.000. = "_ Blank"> Klik hier om een grotere versie van deze figuur te bekijken.
FRAP_instructions.pdf -. Aanwijzingen voor het opzetten van een FRAP reeks Klik hier om dit bestand te downloaden.
ReadMe_FRAP.txt -. Tekstbestand beschrijving van de procedure om FRAP herstel van gegevens met behulp van de MATLAB-code die wordt verstrekt analyseren Klik hier om dit bestand te downloaden.
EK_FRAPbatch.m - MATLAB-bestand naar batch-analyse van fluorescentie herstel na fotobleken. Dit bestand noemt de MATLAB subroutines EK_getFRAPvals.m en EK_SoumpassisFit.m."_blank"> Klik hier om dit bestand te downloaden.
EK_getFRAPvals.m -. MATLAB-bestand, subroutine opgeroepen door EK_FRAPbatch.m Klik hier om dit bestand te downloaden.
EK_SoumpassisFit.m -. MATLAB-bestand, subroutine opgeroepen door EK_FRAPbatch.m Klik hier om dit bestand te downloaden.
JN150923c1_FRAP_list.txt -. Een voorbeeld van een lijst met bestandsnamen to-batch geanalyseerd door de belangrijkste MATLAB script EK_FRAPbatch.m zijn Klik hier om dit bestand te downloaden.
Devolgende bestanden TIF stacks en de bijbehorende tekstbestanden vijf voorbeelden van FRAP sequenties die kunnen worden geanalyseerd door het uitvoeren van EK_FRAPbatch.m en het kiezen van de JN150923c1_FRAP_list.txt bestand wanneer gevraagd om een lijst met bestanden te kiezen.
JN150923c1_F1_MMStack_Pos0.ome.tif -. FRAP afbeeldingsstapel Klik hier om dit bestand te downloaden.
JN150923c1_F1_MMStack_Pos0_metadata.txt -. Corresponderende tekstbestand met de metadata Klik hier om dit bestand te downloaden.
JN150923c1_F2_MMStack_Pos0.ome.tif -. FRAP afbeeldingsstapel Plverlichten klik hier om dit bestand te downloaden.
JN150923c1_F2_MMStack_Pos0_metadata.txt -. Corresponderende tekstbestand met de metadata Klik hier om dit bestand te downloaden.
JN150923c1_F3_MMStack_Pos0.ome.tif -. FRAP afbeeldingsstapel Klik hier om dit bestand te downloaden.
JN150923c1_F3_MMStack_Pos0_metadata.txt -. Corresponderende tekstbestand met de metadata Klik hier om dit bestand te downloaden.
JN150923c1_F4_MMStack_Pos0.ome.tif - FRAP afbeeldingsstapel. Klik hier om dit bestand te downloaden.
JN150923c1_F4_MMStack_Pos0_metadata.txt -. Corresponderende tekstbestand met de metadata Klik hier om dit bestand te downloaden.
JN150923c1_F5_MMStack_Pos0.ome.tif -. FRAP afbeeldingsstapel Klik hier om dit bestand te downloaden.
JN150923c1_F5_MMStack_Pos0_metadata.txt -. Corresponderende tekstbestand met de metadata Klik hier om dit bestand te downloaden.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Succesvolle implementatie van de SUV-SBL fusie test hier beschreven is afhankelijk kritisch over een aantal belangrijke stappen, zoals functionele reconstructie van eiwitten in liposomen, het verkrijgen van goede kwaliteit SBLs, en het kiezen van de juiste beeldvorming parameters om enkele moleculen te detecteren. Hoewel het wat tijd en moeite kunnen nemen om te slagen, wanneer de test succesvol is geïmplementeerd, biedt het een schat aan informatie over het fusieproces niet beschikbaar zijn van een andere in vitro fusie test besproken in de Inleiding. De snelheid waarmee SUV dock op en zekering met SBL, de verdeling van docking-to-fusie tijden lipide diffusiviteit, de grootte van elke liposoom fuseren en of lipiden bericht voor een bepaalde samensmelting is diffusie of porie gecontroleerde can wees vastberaden. Als release is veel langzamer dan verwacht van diffusie, een model dat porie flikkeren veronderstelt ten grondslag ligt aan de vertraging van de opbrengsten lipide release, P 0, de fractie van de tijd de porie iis geopend tijdens fusion 27.
Vorige werk indringende enkele SUV-SUV fusie 18-20,28 of fusie van de liposomen en virale deeltjes SBLs 15-17,21,22,33,44 vertrouwden op FRET en zelf-uitdoving tussen kleurstofmoleculen. Daarentegen we grotendeels voorkomen kleurstof dequenching tot kinetiek van lipide etiketoverdrachtspositie sonde in de SBL, om twee redenen. Ten eerste FRET efficiëntie is een sterk niet-lineaire reporter lipide release. Kleine veranderingen in de onderlinge afstand d kleurstof leiden tot grote veranderingen in FRET efficiëntie, maar slechts voor een beperkt aantal d / R 0-waarden, waarbij R 0 is de Forster afstand. Dat wil zeggen een vertraging tussen de werkelijke kleurstof verdunning en een waargenomen toename van het dequenching signaal optreden, als gevolg van de tijd die dalen tot een waarde dichtbij 0 R. Ook is er weinig verandering in de dequenching signaal zodra de kleurstofdichtheid afneemt voldoende op lipide afgifte zodat  1,4, zelfs wanneer een aanzienlijke hoeveelheid kleurstof nog steeds in het blaasje. Deze eigenschappen maken dequenching een goed hulpmiddel om een piek te verkrijgen in het signaal voor fusie gebeurtenissen op te sporen, maar compliceren kwantitatieve kinetische analyse van lipide vrijkomen met behulp van hoge tijdsresolutie. Daarentegen worden veranderingen in intensiteit vanwege dipool heroriëntering en verdwijnende veld effecten als fluorofoor overdrachten van een SUV in de SBL lineair gerelateerd aan de fractie van de kleurstof nog in de SUV. Ten tweede kunnen hoge concentraties lipiden labels vereist voor zelf-uitdoving het fusieproces zelf beïnvloeden.
1,4, zelfs wanneer een aanzienlijke hoeveelheid kleurstof nog steeds in het blaasje. Deze eigenschappen maken dequenching een goed hulpmiddel om een piek te verkrijgen in het signaal voor fusie gebeurtenissen op te sporen, maar compliceren kwantitatieve kinetische analyse van lipide vrijkomen met behulp van hoge tijdsresolutie. Daarentegen worden veranderingen in intensiteit vanwege dipool heroriëntering en verdwijnende veld effecten als fluorofoor overdrachten van een SUV in de SBL lineair gerelateerd aan de fractie van de kleurstof nog in de SUV. Ten tweede kunnen hoge concentraties lipiden labels vereist voor zelf-uitdoving het fusieproces zelf beïnvloeden.
Bipolaire TIRF excitatie een zelfgebouwde opstelling als in figuur 4 is kosteneffectief en biedt een uitstekende controle over de polarisatie-eigenschappen. Echter, een zelfgebouwde setup is niet noodzakelijk, ten minste sommige commerciële TIRF microscopen gepolariseerde excitatie en off-the-shelftwee kleuren emissie splitsing modules beschikbaar. Een commerciële microscoop die een polarisatie handhaven (PM) vezels gebruikt voor het koppelen van het gepolariseerd excitatielicht die uit de laser in de microscoop werd eerder met succes 25-27 toegepast. Op een dergelijk instrument kan de polarisatie eenvoudig worden geroteerd door het roteren van de PM vezel ingevoerd in de microscoop TIRF apparaat. Daarentegen kunnen andere commerciële instellingen een polariserende bundelsplitser gebruik in de TIRF eenheid, waardoor polarisatie aanpassingen moeilijker of onmogelijk. Helaas zijn polarisatie-eigenschappen vaak niet gespecificeerd voor commerciële systemen. Het is dus van cruciaal belang voor gebruikers die niet willen een zelfgebouwde systeem om polarisatie-eigenschappen te bespreken en om een demo-eenheid te proberen alvorens te investeren in een bepaalde commerciële optie te construeren.
Gelijktijdige bewaking van lipiden en oplosbare lading vrijgave geeft nadere informatie over het fusieproces. Veel poriën lijken opnieuw te verzegelen na releAsing slechts een fractie van de oplosbare label ingekapseld in een SUV 27. Meestal zijn alle lipide etiketten worden vrijgegeven op het moment dat de porie reseals, dus poriën opnieuw afdichten werd niet eerder verdacht van het toezicht op lipide-versie alleen. Sterker nog, het toezicht op lipide vrijgave alleen, had Kiessling et al. 29 voorgesteld overdracht van gelabelde lipiden in de SBL heeft plaatsgevonden via een extreem snelle ineenstorting van het hele SUV membraan in de SBL.
Nieuwe sluiting van fusie poriën na de gedeeltelijke vrijgave van de inhoud die eerder werd waargenomen in een liposoom-tethered bilaag patch fusion-test. Rawle et al. 54 bereid vrijstaande tweelaags patches die op een dekglaasje werden vastgebonden door ~ 8 nm lang DNA spacers. Vervolgens leidt liposomen waarvan de fusie aan de bilaag patch gedreven door hybridisatie van complementaire enkelstrengs DNA-lipide conjugaten verankerd aan het liposoom en patch membranen. 12% van alle gebeurtenissen werden voorbijgaand fusies, die gedeeltelijk samenhangtial vrijkomen van de inhoud liposoom. Liposoom barsten gebeurtenissen, gezien in een aantal eerdere SUV-SBL fusie assays 44, waren zeer zeldzaam. De auteurs gesuggereerd the ~ 8 nm ruimte onder de bilaag patch, voorkomen bilaag-substraat interacties kunnen toegestaan niet-lekkende overbrengen van de inhoud liposoom om de ruimte onder de bilaag. Bovendien, werd diffusie van de vrijgemaakte oplosbare labels niet significant belemmerd in de ruimte tussen de vlakke bilaag en het glassubstraat. In de hier beschreven test, is de vlakke dubbellaag eveneens bewaard ~ 5 nm van het dekglaasje door de aanwezigheid van poly (ethyleenglycol) -lipid conjugaten.
De door tweekleurige monitoring van oplosbaar en lipide lading aanvullende informatie geleverd met een aantal trade-offs. Ten eerste, de voorbereiding van SUV's met de twee labels is iets meer betrokken dan met inbegrip van lipide-labels alleen. Ten tweede, is er enige onvermijdelijke verlies van signaal-ruis bij het monitoren signalen van het lipide labels bij thij dezelfde overname tempo als in één kleur lipide-afgifte-experimenten (15-20 msec per frame). Sommige van deze schade te wijten aan het feit dat was gebruikt als een lipide label met de oplosbare SRB marker, is niet zo helder als LR-PE gebruikt eenkleurige experimenten. Extra verliezen gevolg van de hogere achtergrond door gelijktijdig twee excitatie-laser en andere componenten in het optische pad (figuur 4). Tot dusver zijn de lagere signaal-ruis ons verhinderd betrouwbaar volgen van enkele DiD labels in twee kleuren experimenten. Hierdoor voorkwam een grondige analyse van alle informatie die met een kleur LR-PE metingen te extraheren. Deze uitdaging kan in de toekomst worden overwonnen door het optimaliseren van overname voorwaarden en het testen van andere paren van compatibele etiketten met verbeterde eigenschappen.
Gelijktijdige controle van oplosbare en lipide lading vrijlating kan ook hemifusion, een toestand waarin de proximale folders versmolten detecteren, maar de distalefolders niet. Inderdaad kunnen dergelijke tweekleurige experimenten de enige definitieve beoordeling van de aanwezigheid van hemifusion in andere in vitro assays 18,20 fusie verschaffen. Toch kan hemifusion betrouwbaar worden gedetecteerd in één kleur SUV-SBL experimenten bewaking lipide alleen 22,44 release. In dergelijke experimenten ongeveer 1/2 van de oorspronkelijke lipide label intensiteit van een SUV blijft het fusie plaats. De resterende intensiteit kan verdwijnen wanneer het distale bladen fuseren enige tijd later, wanneer de vlek nog niet gebleekt.
Het gebruik van gePEGyleerde lipiden in te voeren een zacht kussen tussen de SBL en de glazen steun heeft sleutel in het reproduceren van de SNAP25 eis van exocytose geweest. Het protocol levert een PEG borstel die de zijkant van de SBL dat het stromingskanaal gezichten ook. Dit heeft een aantal voordelen, zoals het verminderen van niet-specifieke interacties tussen SUV en SBL, zoals uiteengezet in de inleiding. Als eiwitten die interageren met de fosfolipiden van deSBL worden in de assay, maar de PEG borstel een sterische barrière tegen dergelijke interacties presenteren. Zo sommige toepassingen is het wenselijk om de PEG laag asymmetrisch omvatten zijn alleen tussen de SBL en glazen dekglaasje. De zijkant van de SBL tegenover het stromingskanaal zou dan in staat zijn met fosfolipide bindende eiwitten, zoals synaptotagmin. Dit kan worden bereikt door gebruik bifunctioneel PEG ketens die covalent kan worden gekoppeld aan het dekglaasje aan één einde en met een lipide anker aan het andere. Langmuir-Blodgett werkwijzen kunnen vervolgens worden gebruikt om een lipide monolaag op het PEG-laag 55 deponeren. Liposomen boven dit hydrofoob oppervlak zekering introduceerde, de vorming van een tweelaags 56. Gehoopt wordt dat deze aanpak zal worden gecombineerd met microfluidics in de toekomst.
Verwacht wordt dat de SUV-SBL test hier gepresenteerde behulpzaam onderzoeken de rol van additionele eiwitten die interageren met SNAREs zal zijn,zoals de familie Munc18 / Sec1, calcium sensor synaptotagmin-1 en complexin.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs verklaren dat ze geen concurrerende financiële belangen.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| Milli-Q (MQ) water | Millipore | ||
| KOH | J.T. Baker | 3040-05 | |
| Ethanol 190 Proof | Decon | ||
| Isopropanol | Fisher Chemical | A416P4 | |
| HEPES | AmericanBio | AB00892 | |
| Sodium Cholride (KCl) | AmericanBio | AB01915 | |
| Dithiothreitol | AmericanBio | AB00490 | |
| N-[2-hydroxyethyl] piperazine-N'-[2-ethanesulfonic acid] (HEPES) | AmericanBio | AB00892 | |
| EGTA | Acros Organics | 409911000 | |
| Buffers | |||
| HEPES-KOH buffer (pH 7.4) | 25 mM HEPES-KOH, 140 mM KCl, 100 μM EGTA, 1 mM DTT | ||
| Solvents | |||
| Chloroform | J.T. Baker | 9180-01 | in glass bottle, CAUTION, wear PPE |
| Methanol | J.T. Baker | 9070-03 | in glass bottle, CAUTION, wear PPE |
| Liposome preparation | |||
| Gastight Hamilton syringe | Hamilton | var. sizes | only use glass sringe with solents (Chlorophorm/ Methanon, 2:1, v/v) |
| Glass tubes Pyrex Vista 11 ml, 16x100 mm screw cap culture tube | Pyrex | 70825-16 | clean thoroughly, rinse with chloroform |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine, 16:0-18:1 PC (POPC) | Avanti Polar Lipids | 850457 | Lipids come dissolved in CHCl3 or as lyphilized powder in sealed vials. Aliquot upon opening. Store extra as dried lipid films under inert atmosphere at -20 °C. Keep stocks in CHCl3/MeOH (2:1, v/v) at -20 °C. let come to RT before opening |
| 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (sodium salt), 18:1 PS (DOPS) | Avanti Polar Lipids | 840035 | Same as above. |
| 1-stearoyl-2-arachidonoyl-sn-glycero-3-phosphoethanolamine, 18:0-20:4 PE (SAPE) | Avanti Polar Lipids | 850804 | Same as above. |
| L-α-phosphatidylinositol-4,5-bisphosphate (Brain, Porcine) (ammonium salt), Brain PI(4,5)P2 | Avanti Polar Lipids | 840046 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(7-nitro-2-1,3-benzoxadiazol-4-yl) (ammonium salt), 18:1 NBD PE | Avanti Polar Lipids | 810145 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene glycol)-2000] (ammonium salt), 18:1 PEG2000 PE | Avanti Polar Lipids | 880130 | Same as above. |
| cholesterol (ovine wool, >98%) | Avanti Polar Lipids | 700000 | Same as above. |
| DiD' oil; DiIC18(5) oil (1,1'-Dioctadecyl-3,3,3',3'-Tetramethylindodicarbocyanine Perchlorate) | Molecular Probes | D-307 | |
| Rotavapor R-210 | Buchi | R-210 | heat bath above Tm of lipids used |
| OG n-Octyl-β-D-Glucopyranoside | Affymetrix | 0311 | store at -20 °C, let come to RT before opening |
| Shaker - Eppendorf Thermomixer R | Eppendorf | ||
| Slide-A-Lyze Dialysis Cassettes, 20k MWCO, 3 ml | life technologies | 66003 | |
| Bio-Beads SM-2 Adsorbents | Bio-Rad | 1523920 | |
| OptiPrep Density Gradient Medium | Sigma-Aldrich | D1556 | |
| Ultracentrifugation tube, Thinwall, Ultra-Clear, 13.2 ml, 14 x 89 mm | Beckman Coulter | 41121703 | |
| Beckman SW41 Ti rotor | |||
| SuflorhodamineB | Molecular Probes | S-1307 | |
| Econo-Column Chromatography Columns, 2.5 × 10 cm | Bio-Rad | 7372512 | |
| Sepharose CL-4B | GE Healthcare | 17-0150-01 | |
| SYPRO Orange Protein Gel Stain | Molecular Probes | S-6650 | 5,000x Concentrate in DMSO |
| PDMS block | |||
| Sylgard 184 Silicone elastomer kit, PDMS | Dow Corning | 3097358-1004 | |
| Pyrex glass petri dish, 150 x 20 mm, complete with cover | Corning | 3160-152 | |
| Hole puncher - Reusable Biopsy Punch, 0.75 mm | World Precision Instruments | 504529 | |
| Manual Hole Punching Machine | SYNEO | MHPM-UNV | |
| Drill .035 x .026 x 1.5 304 SS TiN coated round punch | SYNEO | CR0350265N20R4 | drill diameter: 0.9 mm, punch bore size: 0.74 mm |
| Tygon Microbore tubing, 0.25 mm ID, 0.76 mm OD | Cole-Parmer | 06419-00 | 0.010" ID, 0.030" OD |
| Silicone Tubing (0.51 mm ID, 2.1 mm OD | Cole-Parmer | 95802-00 | 0.020" ID, 0.083" OD |
| Cover glass - cleanroom cleaned | |||
| Schott Nexterion cover slip glass D | Schott | 1472305 | |
| plasma cleaner | Harrick | PDC-32G | |
| pTIRF setup and accessories | |||
| IX81 microscope body | Olympus | IX81 | |
| EM CCD camera | Andor | ixon-ultra-897 | |
| Thermo Plate, heated microscope stage | Tokai Hit | MATS-U52RA26 | |
| 1 ml hamilton glass syringes (4x) | Hamilton | 81365 | |
| syringe pump | kd Scientific | KDS-230 | |
References
- Sudhof, T. C., Rothman, J. E. Membrane fusion: grappling with SNARE and SM proteins. Science. 323, 474-477 (2009).
- Wickner, W., Schekman, R. Membrane fusion. Nat Struct Mol Biol. 15, 658-664 (2008).
- Harrison, S. C. Viral membrane fusion. Nat Struct Mol Biol. 15, 690-698 (2008).
- Jahn, R., Scheller, R. H. SNAREs--engines for membrane fusion. Nat Rev Mol Cell Biol. 7, 631-643 (2006).
- Lindau, M., Alvarez de Toledo, G. The fusion pore. Biochim Biophys Acta. 1641, 167-173 (2003).
- Staal, R. G., Mosharov, E. V., Sulzer, D. Dopamine neurons release transmitter via a flickering fusion pore. Nat Neurosci. 7, 341-346 (2004).
- Wu, Z., et al. Nanodisc-cell fusion: Control of fusion pore nucleation and lifetimes by SNARE protein transmembrane domains. Sci. Rep. 6, 27287 (2016).
- Alabi, A. A., Tsien, R. W. Perspectives on kiss-and-run: role in exocytosis, endocytosis, and neurotransmission. Ann Rev Physiol. 75, 393-422 (2013).
- Rossetto, O., Pirazzini, M., Montecucco, C. Botulinum neurotoxins: genetic, structural and mechanistic insights. Nature Rev Microbiol. 12, 535-549 (2014).
- Weber, T., et al. SNAREpins: minimal machinery for membrane fusion. Cell. 92, 759-772 (1998).
- Nickel, W., et al. Content mixing and membrane integrity during membrane fusion driven by pairing of isolated v-SNAREs and t-SNAREs. Proc Natl Acad Sci U S A. 96, 12571-12576 (1999).
- McNew, J. A., et al. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature. 407, 153-159 (2000).
- Melia, T. J., You, D. Q., Tareste, D. C., Rothman, J. E. Lipidic antagonists to SNARE-mediated fusion. J Biol Chem. 281, 29597-29605 (2006).
- Hernandez, J. M., et al. Membrane fusion intermediates via directional and full assembly of the SNARE complex. Science. 336, 1581-1584 (2012).
- Fix, M., et al. Imaging single membrane fusion events mediated by SNARE proteins. Proc Natl Acad Sci U S A. 101, 7311-7316 (2004).
- Bowen, M. E., Weninger, K., Brunger, A. T., Chu, S. Single molecule observation of liposome-bilayer fusion thermally induced by soluble N-ethyl maleimide sensitive-factor attachment protein receptors (SNAREs). Biophys J. 87, 3569-3584 (2004).
- Liu, T., Tucker, W. C., Bhalla, A., Chapman, E. R., Weisshaar, J. C. SNARE-driven, 25-millisecond vesicle fusion in vitro. Biophys J. 89, 2458-2472 (2005).
- Yoon, T. Y., Okumus, B., Zhang, F., Shin, Y. K., Ha, T. Multiple intermediates in SNARE-induced membrane fusion. Proc Natl Acad Sci U S A. 103, 19731-19736 (2006).
- Diao, J., et al. A single-vesicle content mixing assay for SNARE-mediated membrane fusion. Nat Commun. 1, 1-6 (2010).
- Kyoung, M., et al. In vitro system capable of differentiating fast Ca2+-triggered content mixing from lipid exchange for mechanistic studies of neurotransmitter release. Proc Natl Acad Sci U S A. 108, E304-E313 (2011).
- Domanska, M. K., Kiessling, V., Stein, A., Fasshauer, D., Tamm, L. K. Single vesicle millisecond fusion kinetics reveals number of SNARE complexes optimal for fast SNARE-mediated membrane fusion. J Biol Chem. 284, 32158-32166 (2009).
- Kreutzberger, A. J., Kiessling, V., Tamm, L. K. High Cholesterol Obviates a Prolonged Hemifusion Intermediate in Fast SNARE-Mediated Membrane Fusion. Biophys J. 109, 319-329 (2015).
- Schwenen, L. L., et al. Resolving single membrane fusion events on planar pore-spanning membranes. Sci Rep. 5, 12006 (2015).
- Karatekin, E., et al. A fast, single-vesicle fusion assay mimics physiological SNARE requirements. Proc Natl Acad Sci U S A. 107, 3517-3521 (2010).
- Karatekin, E., Rothman, J. E. Fusion of single proteoliposomes with planar, cushioned bilayers in microfluidic flow cells. Nat Protoc. 7, 903-920 (2012).
- Smith, M. B., et al. Interactive, computer-assisted tracking of speckle trajectories in fluorescence microscopy: application to actin polymerization and membrane fusion. Biophys J. 101, 1794-1804 (2011).
- Stratton, B. S., et al. Cholesterol Increases the Openness of SNARE-mediated Flickering Fusion Pores. Biophysical journal. 110, (2016).
- Diao, J., et al. Synaptic proteins promote calcium-triggered fast transition from point contact to full fusion. Elife. 1, e00109 (2012).
- Kiessling, V., Domanska, M. K., Tamm, L. K. Single SNARE-mediated vesicle fusion observed in vitro by polarized TIRFM. Biophys J. 99, 4047-4055 (2010).
- Blasi, J., et al. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature. 365, 160-163 (1993).
- Washbourne, P., et al. Genetic ablation of the t-SNARE SNAP-25 distinguishes mechanisms of neuroexocytosis. Nat Neurosci. 5, 19-26 (2002).
- Diaz, A. J., Albertorio, F., Daniel, S., Cremer, P. S. Double cushions preserve transmembrane protein mobility in supported bilayer systems. Langmuir. 24, 6820-6826 (2008).
- Floyd, D. L., Ragains, J. R., Skehel, J. J., Harrison, S. C., van Oijen, A. M. Single-particle kinetics of influenza virus membrane fusion. Proc Natl Acad Sci U S A. 105, 15382-15387 (2008).
- Albertorio, F., et al. Fluid and air-stable lipopolymer membranes for biosensor applications. Langmuir. 21, 7476-7482 (2005).
- Daniel, S., Albertorio, F., Cremer, P. S. Making lipid membranes rough, tough, and ready to hit the road. Mrs Bulletin. 31, 536-540 (2006).
- Gao, Y., et al. Single reconstituted neuronal SNARE complexes zipper in three distinct stages. Science. 337, 1340-1343 (2012).
- Kenworthy, A. K., Hristova, K., Needham, D., Mcintosh, T. J. Range and Magnitude of the Steric Pressure between Bilayers Containing Phospholipids with Covalently Attached Poly(Ethylene Glycol). Biophys J. 68, 1921-1936 (1995).
- Knoll, W., et al. Solid supported lipid membranes: New concepts for the biomimetic functionalization of solid surfaces. Biointerphases. 3, Fa125-Fa135 (2008).
- Quinn, P., Griffiths, G., Warren, G. Density of newly synthesized plasma membrane proteins in intracellular membranes II. Biochemical studies. J Cell Biol. 98, 2142-2147 (1984).
- Sund, S. E., Swanson, J. A., Axelrod, D. Cell membrane orientation visualized by polarized total internal reflection fluorescence. Biophys J. 77, 2266-2283 (1999).
- Johnson, D. S., Toledo-Crow, R., Mattheyses, A. L., Simon, S. M. Polarization-controlled TIRFM with focal drift and spatial field intensity correction. Biophys J. 106, 1008-1019 (2014).
- Anantharam, A., Onoa, B., Edwards, R. H., Holz, R. W., Axelrod, D. Localized topological changes of the plasma membrane upon exocytosis visualized by polarized TIRFM. J Cell Biol. 188, 415-428 (2010).
- Axelrod, D. Carbocyanine dye orientation in red cell membrane studied by microscopic fluorescence polarization. Biophys J. 26, 557-573 (1979).
- Wang, T., Smith, E. A., Chapman, E. R., Weisshaar, J. C. Lipid mixing and content release in single-vesicle, SNARE-driven fusion assay with 1-5 msec resolution. Biophys J. 96, 4122-4131 (2009).
- Chernomordik, L. V., Frolov, V. A., Leikina, E., Bronk, P., Zimmerberg, J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J Cell Biol. 140, 1369-1382 (1998).
- Takamori, S., et al. Molecular anatomy of a trafficking organelle. Cell. 127, 831-846 (2006).
- Wilhelm, B. G., et al. Composition of isolated synaptic boutons reveals the amounts of vesicle trafficking proteins. Science. 344, 1023-1028 (2014).
- Scott, B. L., et al. Liposome fusion assay to monitor intracellular membrane fusion machines. Methods Enzymol. 372, 274-300 (2003).
- Linkert, M., et al. Metadata matters: access to image data in the real world. J Cell Biol. 189, 777-782 (2010).
- Soumpasis, D. M. Theoretical analysis of fluorescence photobleaching recovery experiments. Biophys J. 41, 95-97 (1983).
- Ohki, S. A mechanism of divalent ion-induced phosphatidylserine membrane fusion. Biochim Biophys Acta. 689, 1-11 (1982).
- Berquand, A., et al. Two-step formation of streptavidin-supported lipid bilayers by PEG-triggered vesicle fusion. Fluorescence and atomic force microscopy characterization. Langmuir. 19, 1700-1707 (2003).
- Tamm, L. K., McConnell, H. M. Supported phospholipid bilayers. Biophys J. 47, 105-113 (1985).
- Rawle, R. J., van Lengerich, B., Chung, M., Bendix, P. M., Boxer, S. G. Vesicle fusion observed by content transfer across a tethered lipid bilayer. Biophys J. 101, L37-L39 (2011).
- Wagner, M. L., Tamm, L. K. Reconstituted syntaxin1a/SNAP25 interacts with negatively charged lipids as measured by lateral diffusion in planar supported bilayers. Biophys J. 81, 266-275 (2001).
- Kalb, E., Frey, S., Tamm, L. K. Formation of Supported Planar Bilayers by Fusion of Vesicles to Supported Phospholipid Monolayers. Biochimica Et Biophysica Acta. 1103, 307-316 (1992).
