

Summary
Här presenterar vi ett protokoll för att detektera enstaka, snara-medierad fusion händelser mellan liposomer och stöds dubbelskikt i mikroflödessystem kanaler med hjälp av polariserat TIRFM, med enda molekyl känslighet och ~ 15 ms tidsupplösning. Lipid och löslig lastfrigör kan detekteras samtidigt. Liposomstorlek, lipid diffusivitet, och fusion pore egenskaper mäts.
Abstract
I den allestädes närvarande processen för membranfusion öppnandet av en fusion por etablerar den första anslutningen mellan två tidigare separata fack. Under signalsubstans eller hormonfrisättning via exocytos, fusions por kan övergående öppna och stänga flera gånger, som reglerar lastfrisättningskinetik. Pore dynamik bestämmer också läget av vesikler återvinning, irreversibla återförslutning resulterar i övergående, "kiss-and-run" fusion leder medan utvidgning till full fusion. För att bättre förstå vilka faktorer styr pore dynamik, utvecklade vi en analys för att övervaka membranfusion med hjälp av polariserat total inre reflektion fluorescens (TIRF) mikroskopi med enda molekyl känslighet och ~ 15 ms tidsupplösning i ett biokemiskt väldefinierat in vitro-system. Fusion av fluorescerande små unilamellära vesiklar som innehåller v-SNARE proteiner (v-SUV) med en plan tvåskiktsmembran bärande t-snaror, uppburen på en mjuk polymer kudde (t-SBL, t-stöd tvåskiktsmembran), Övervakas. Analysen använder mikroflödessystem flödeskanaler som säkerställer minimal provförbrukningen samtidigt leverera en konstant densitet av stadsjeepar. Utnyttja den snabba signalförstärkning vid överföring av lipid etiketter från SUV till SBL under fusion, är kinetiken lipid färgöverförings övervakas. Känsligheten hos TIRF mikroskopi tillåter att spåra enskilda fluorescerande lipid etiketter, från vilken lipid diffusivitet och SUV storlek kan härledas för varje fusion händelse. Lipid färgfrisättningstider kan vara mycket längre än väntat för obehindrad passage genom permanent öppna porer. Med hjälp av en modell som förutsätter fördröjning av lipid frisättning beror på pore flimmer, en por "öppenhet", den del av tiden porerna förblir öppen under fusion, kan uppskattas. En löslig markör kan inkapslas i SUV för samtidig övervakning av lipid och löslig last release. Sådana mätningar indikerar vissa porer kan återförsluta efter att ha förlorat en del av den lösliga last.
Introduction
Membranfusion är en universell biologisk process som krävs för intracellulär handel med lipider och proteiner, utsöndring, gödsling, utveckling och insvept virus inträde i värdorganismer 1-3. För de flesta intracellulära fusionsreaktioner inklusive frisättning av hormoner och signalsubstanser via exocytos, är energin för att sammansmälta två lipiddubbelskikt tillhandahålls genom bildning av ett fyra-helix bundle mellan cognate löslig N-etylmaleimid-känslig faktor fastsättning proteinreceptor (SNARE) proteiner, förankrad i vesikeln (v-SNARE) och målmembranet (t-SNARE) 4, respektive. Synaptic vesikler exocytos är den mest hårt reglerad fusion reaktion och inträffar inom en millisekund efter ankomsten av en aktionspotential 1,4,5. Fusions por, den första anslutningen mellan de två fixerings facken, kan flimmer öppna och slutna flera gånger innan återförslutning eller expandera irreversibelt 5-7. De tidigare resultatövergående, "Kiss & run" fusion, medan den senare leder till full fusion. Faktorer som styr balansen mellan dessa två typer av fusion och mekanismer som reglerar por flimmer är inte väl förstått 5,8.
SNARE proteiner krävs för exocytos; synaptiska vesikelfusion avskaffas vid klyvning av snaror av neurotoxiner 9. Bulkfusionsexperiment med användning av små unilamellära vesiklar (SUVs) visade att snaror inte bara krävs, men också tillräcklig för att driva membranfusion 10. I detta bulk assay, SUVs rekonstitueras med v-snaror (v-SUV) till dopad med fluorescerande fosfolipider (N - (7-nitro-2-1,3-bensoxadiazol-4-yl) -phosphoethanolamine (NBD-PE) och ( N -. (lissaminrodamin B sulfonyl) -phosphoethanolamine (LR-PE) och blandades med omärkta vesiklar innehållande t-snaror (t-SUV) Initialt fluorescensen av NBD-PE i v-SUVs släckes genom Förster resonansenergiöverföring (FRET ) till LR-PE. som labELED v-SUV smälta samman med omärkta t-SUV, är fluoroforen ytdensitet i det nu kombinerade membranet reduceras och den resulterande ökningen i NBF-PE fluorescens rapporterar omfattningen av lipidblandning 10. Som bulk analysen är lätt att ställa upp och analysera, har det i stor utsträckning använts för att studera mekanismerna för SNARE-medierad sammansmältning 10-14. Det har dock flera begränsningar, såsom låg känslighet och dålig tidsupplösning. Viktigast som en ensemble mätning, det genomsnitt resultat under alla omständigheter gör diskriminering mellan dockning och fusion, samt detektering av hemifusion mellan svårt.
Under det senaste årtiondet flera grupper, inklusive vårt, har utvecklat nya analyser för att övervaka fusions evenemang på enstaka vesikler nivå 15-27. Ha och kollegor använde v-SUV bundna på en yta och övervakas deras fusion med gratis t-SUV 18,19. Lipidblandning övervakades med användning av FRET mellan ett par lipid-bundna fluoroforer embäddas in i v- och t-SUV, respektive, med hjälp av total inre reflektion fluorescens (TIRF) mikroskopi 18. Senare Brunger laboratorium använde en enda lipid-label art tillsammans med en innehåll markör för samtidig detektion av lipid och innehåll blanda 20,28. Både lipid och innehållet markörerna ingick vid höga, självläckande koncentrationer; fusion med omärkta SUVs resulterade i fluorescens dequenching 20,28.
Andra har smält v-SUV till plana bilayers ombildade med t-snaror 15-17,21-27,29. Den plana geometri målet (t-SNARE innehållande) biskikt bättre efterliknar den fysiologiska fusionsprocessen av små, mycket böjda vesiklar med platt plasmamembranet. Den Steinem grupp som användes pore överbryggande membran rekonstituerade med t-snaror, suspenderade genom ett poröst kiselnitrid substratet och detekteras fusion med enskilda v-SUVs med användning av konfokal laserscanningsmikroskopi 23. andra fbegagnade v-SUVs till plana biskikt rekonstituerade med t-snaror, uppburen på ett glassubstrat 15-17,21,22,24-27,29. Den stora fördelen med att använda stöds dubbla skikt (SBLs) är att TIRF mikroskopi kan användas för att detektera dockning och fusion händelser med utmärkt signal-brusförhållande och utan inblandning från fria v-SUV, även om användning av mikrofluidik ger också enstaka händelse upplösning med standard fjärrfälts epifluorescensmikroskopi 24.
En viktig fråga är om och hur substrat tvåskiktsmembran interaktioner påverkar stöds tvåskiktsmembran kvalitet och fusionsprocessen. Tidigt arbete utnyttjade plana SBLs som direkt stöd på ett glas eller kvartssubstrat 15-17. Dessa SBLs gjordes genom adsorption, spricker, spridning och blandning av t-SUV membran på substratet. Det var snart insåg dock att utelämna en viktig t-SNARE komponent, SNAP25 från SBLs framställda på detta sätt resulterade i v-SUV dockning och fusion kinetik indistinguisHable från de som erhållits genom att använda den fullständiga t-snaror 17. Eftersom SNAP25 är absolut nödvändig för fusion in vivo 30,31, var den fysiologiska relevansen av dessa tidiga försök ifrågasättas. Tamm grupp klarade denna utmaning genom att använda kontrolleras bättre stöd tvåskiktsmembran bildning 21. Det brukade Langmuir-Blodgett avsättning för proteinfritt första broschyr av SBL, följt av fusion av det monolager med t-SUV 21. Detta resulterade i SNAP25 beroende fusion.
För att undvika potentiella artefakter associerade med ett dubbelskikt direkt uppburen på ett glassubstrat utan behov av att använda Langmuir-Blodgett metoder, Karatekin et al. Infört en mjuk, hydratiserad poly (etylenglykol) (PEG) kudde mellan dubbelskiktet och substratet 24. Denna modifiering resulterade också i SNAP25 beroende fusion 24. Dubbelskikt vadderade på ett mjukt polymerskikt hade varit kända för att bättre bevara transprotein rörlighet och funktion 32, och hade använts i fusionsstudier med virus 33. Dessutom PEGylerade bilayers tycks behålla en del förmåga att självläka och är mycket robust 34,35. Först är en fraktion av kommersiellt tillgängliga, lipid-länkad PEG-kedjor som ingår i t-SUV membran. När dessa t-SUVs brista och bilda en plan tvåskiktsmembran på ett glassubstrat, omfattar en PEG borste båda broschyrer av den plana biskikt. Eftersom plana biskikt bildning drivs av vidhäftning av PEG-kedjor närliggande t-SUVs på den hydrofila glasytan, liposom sprängning och plana biskikt bildning är relativt okänsliga för lipidkompositionen användas. När emellertid stora mängder kolesterol ingår, ökar de kohesiva egenskaperna hos SUVs, SUVs kanske inte brista spontant. Om detta är fallet, kan osmotisk chock eller tvåvärda joner användas för att hjälpa till plana biskikt bildning 25.
Såsom nämnts ovan, i detta apgrepp en PEG borste täcker båda sidorna av den plana, uppburen dubbelskiktet. Borsten mot mikroflödesflödeskanalen hjälper till att förhindra ospecifik vidhäftning av inkommande v-SUV som också är oftast täckt med en PEG lager. Bildning av V- och t-SNARE-komplex utgår från membran distala N-terminaler och fortsätter i etapper mot membran-proximala domänerna 36. För v-SUV att interagera med t-SBL, V- och t-SNARE N-terminaler är nödvändigt att skjuta över PEG borstar, som verkar vara fallet i enlighet med villkoren i analysen. Borsthöjden kan anpassas för att studera andra än snaror proteiner genom att variera densiteten hos PEGylerade lipider och PEG kedjelängd 37,38. En annan fördel med de PEG-borstar som täcker de proximala ytorna av fixerings bilayers är att de efterliknar det trånga miljön i biologiska membran, vilka är packade med 30,000-40,000 integrerade membranproteiner per kvadratmikron 39. Precis som de PEG-kedjor i denna analys, den Repulsive proteinskikt som täcker biologiska membran måste skjutas åt sidan för att tillåta kontakt mellan de två fosfolipid bilayers för fusion skall ske.
Mikroflödesströmningskanaler används i denna analys, eftersom de erbjuder unika fördelar. Först gör mikroflödes flöde jämnare avsättning av T-SUV för att sprida och säkring för att bilda t-SBL. För det andra, (<1 l) minimerar den lilla kanalvolymen provförbrukning. För det tredje, de små volymer som behövs låta hela experimentet genomföras under konstant flöde. Flow avlägsnar svagt, förmodligen ospecifikt, vidhäftade v-SUV från SBL 16. Det har även en konstant densitet av v-SUV ovanför t-SBL, förenklar kinetisk analys 17. Slutligen dockade blåsor lätt skiljas från fri dem som bärs av flödet 25. Fjärde flera mikroflödessystem kanaler kan användas på samma täck varje sondering olika tillstånd. Detta möjliggör jämförelser av villkor During samma experimentkörning. Ett liknande tillvägagångssätt har använts av van Oijen för att studera fusion mellan influensavirus och vadde SBLs 33.
I TIRF mikroskopi, den exponentiella avklingningen av det evanescenta fältet (med en sönderfallskonstant ~ 100 nm) begränsar fluorescensexcitation till de molekyler som är i mycket nära anslutning för det glasbuffertgränssnittet. Detta minimerar bidraget från fluorescerande molekyler som ligger längre bort, ökar signal-brusförhållande, och tillåter enda molekyl känslighet med ram exponeringstider på 10 till 40 msek. Det evanescenta fältet leder också till en signalökning på fusion: som den märkta lipider överföring från SUV till SBL, de befinner sig, i genomsnitt, en starkare exciteringsfältet. Denna ökning i fluorescens är starkare för större liposomer.
Om polariserat ljus används för att generera det evanescenta fältet, ytterligare effekter bidra till förändringar i fluorescens vid tverför av etiketter från SUV i SBL. Vissa lipid-färgämnen har en övergångs dipol orienterad med en föredragen medel vinkel med avseende på dubbelskiktet i vilket de är inbäddade. Detta skapar en skillnad i mängden av fluorescens som emitteras av de fluoroforer när de är i SUV kontra SBL, eftersom den polariserade strålen att excitera färgämnen i de två membranen på olika sätt. För de förstnämnda kommer excitationsstrålen interagera med övergångs dipoler orienterade kring den sfäriska SUV, medan det senare kommer dipol riktlinjer begränsas av den plana SBL geometri. Till exempel, när s-polariserade infallande ljuset (polariserat vinkelrätt mot infallsplanet) används, är excitering mer effektiv när färgämnet är i SBL än i SUV för en lipid färgämne övergångs dipol orienterad parallellt med membranet 29,40 (såsom den av Dil eller gjorde 41-43). En SUV dopad med en sådan fluorofor visas dim när den dockar har laddats in i SBL (Figur 7, Representat ive Resultat). Som en fusion pore öppnas och förbinder de SUV och SBL membran, fluorescerande prober diffundera in i SBL och blir mer sannolikt att exciteras av s-polariserade evanescent fält 25,27,29. Följaktligen ökar fluorescenssignalen integreras runt fusionsstället kraftigt under färgöverföring från SUV till SBL 27 (Figur 3 och Figur 7). En ytterligare faktor som bidrar till signal förändringar som följer fusion dequenching av fluorescerande markörer som de späds när de överförs in i SBL. Bidraget från dequenching är vanligtvis små i jämförelse med evanescent fältröta och polariseringseffekter i analysen beskriven här, eftersom endast en liten fraktion (  ) Av lipiderna är märkta.
) Av lipiderna är märkta.
Signalökning vid fusion kan utnyttjas för att härleda fusion pore egenskaper genom att jämföra den tid,1 "src =" / filer / ftp_upload / 54349 / 54349eq2.jpg "/>, som krävs för en lipid fly genom en por som är fritt permeabel för lipider till frigöra tid själva,  . Om de två tidsskalor är jämförbara, skulle man dra slutsatsen att porerna uppvisar litet motstånd mot lipid flödet. Men om frigöra tid själva är betydligt längre än den tid för diffusion begränsad frisättning, skulle detta tyda på en process, såsom porer flimmer, fördröjer lipid release. Spridning begränsade frigöra tid,
. Om de två tidsskalor är jämförbara, skulle man dra slutsatsen att porerna uppvisar litet motstånd mot lipid flödet. Men om frigöra tid själva är betydligt längre än den tid för diffusion begränsad frisättning, skulle detta tyda på en process, såsom porer flimmer, fördröjer lipid release. Spridning begränsade frigöra tid,  , Beror på storleken av fixerings liposomer och lipid-diffusivitet; sin uppskattning kräver dessa två parametrar som skall kvantifieras. Den enda molekyl känsligheten för analysen tillåter lipid diffusivitet som skall mätas genom att spåra flera enda lipid fluoroforer efter frigivningen i SBL för varje fusion händelse 26. Storleken på varje fusing vesikelkan uppskattas 27 genom att kombinera (i) intensiteten av en enda lipid färgämne, (ii) förändring av det totala fluorescens runt en dockningsområdet efter det alla fluoroforer överförs till SBL upon fusion, (iii) den kända märkningsdensitet av SUV lipider, och (iv) området per lipid. För många fusion händelser, var de faktiska lipid frisättningstider visat sig vara mycket långsammare än väntat genom diffusion-kontrollerad frisättning 27, som noterades tidigare antagande enhetlig SUV storlek 44. Förutsatt fördröjning av lipid frisättning beror på pore flimmer, gör en kvantitativ modell uppskattning av "pore öppenhet", den del av tiden porerna förblir öppen under fusion 27.
, Beror på storleken av fixerings liposomer och lipid-diffusivitet; sin uppskattning kräver dessa två parametrar som skall kvantifieras. Den enda molekyl känsligheten för analysen tillåter lipid diffusivitet som skall mätas genom att spåra flera enda lipid fluoroforer efter frigivningen i SBL för varje fusion händelse 26. Storleken på varje fusing vesikelkan uppskattas 27 genom att kombinera (i) intensiteten av en enda lipid färgämne, (ii) förändring av det totala fluorescens runt en dockningsområdet efter det alla fluoroforer överförs till SBL upon fusion, (iii) den kända märkningsdensitet av SUV lipider, och (iv) området per lipid. För många fusion händelser, var de faktiska lipid frisättningstider visat sig vara mycket långsammare än väntat genom diffusion-kontrollerad frisättning 27, som noterades tidigare antagande enhetlig SUV storlek 44. Förutsatt fördröjning av lipid frisättning beror på pore flimmer, gör en kvantitativ modell uppskattning av "pore öppenhet", den del av tiden porerna förblir öppen under fusion 27.
Närhelst praktiskt, är det viktigt att testa fusionsmekanismer med användning av både lipid- och lösliga innehåll etiketter. Till exempel kan lipid frisättning fördröjas av andra än por flimmer processer, såsom begränsning av lipid diffusion av snaran proteiner som omger than pore. Om så vore fallet, släpp sedan av innehållet skulle föregå frigörande av lipid etiketter, förutsatt att por är tillräckligt stor för att tillåta passage av lösliga sonder. En mer grundläggande brist i den strategi skulle kunna vara i antagandet att överföringen av märkta lipider till SBL sker genom en smal fusion por ansluter SBL till en vesikel som till stor del har behållit sin pre-fusion form. Lipid överföring till SBL kan också vara resultatet av en snabb utvidgning av fusions por med en åtföljande, extremt snabb kollaps av SUV i SBL membran, som föreslog tidigare på lipid släppuppgifter enbart 29. Övervakning både lipid och innehåll släpper samtidigt visade det sig att många porer återförslutas efter att ha släppt alla sina fettetiketter, men behöll en del av deras lösliga last 27. Detta tyder på att åtminstone vissa liposomer inte kollapsar in i SBL efter smältning, och att lipiden färgämnesöverföring in i SBL sker genom en fusion por. Dessutom, lIPID och innehåll frisättning inträffade samtidigt 27, vilket gör det osannolikt att fördröjning av lipid frigör berodde på besvär av lipid diffusion av snaran proteiner omger por 45.
En SUV-SBL fusionsprotokoll som inte övervaka lösliga innehåll frisättning har tidigare publicerats av Karatekin och Rothman 25. Här är den senaste utvecklingen ingår, nämligen samtidig övervakning av lipid och innehåll släpper och uppskattning av SUV, lipid, och fusion pore egenskaper 27. Protokollet börjar med instruktioner för framställning av de mikroflödesceller, tillverkade genom bindning av en poly (dimetylsiloxan) (PDMS) elastomer segmentinnehållande räfflor med ett täckglas 25. Därefter framställning av v-SUV med både lipid och innehåll markörer förklaras. §§ 4 och 5 ger instruktioner för montering av mikroflödesceller, bildar SBLs in situ och kontroll av defekter och smidighet, införande VSUVS in flödescellerna och detektering av fusionshändelser. Avsnitt 6 innehåller instruktioner för dataanalys.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Framställning av en PDMS block för att bilda mikroflödessystem kanal

Figur 1. Mikroflödescell mall och PDMS-block förberedelse. (A) Konstruktion av en flödescell fyra kanaler som passar på en 24 x 60 mm täckglas (botten). Sex identiska mönster är anordnade att passa på en 10 cm kiselskiva (överst). (B) Skär ut block av cirka 5-8 mm tjocka PDMS på en hålslaget. (C) Insättning av slangen i det stansade hålet med en pincett. Klicka här för att se en större version av denna siffra.
- Erhålla en flödescell mall såsom en i figur 1A. Typiska kanaler är 0,3-1 mm breda, 70-100 um höga och 1-2 cm lång. Fabricera en mall ossing standard fotolitografiska tekniker i ett renrum anläggning 25 eller beställning om renrums tillgång inte är tillgänglig. Alternativt maskin en mall med större dimensioner från ett lämpligt material.
Obs: Renrums personal kan träna och vägleda oerfarna användare (tillgång till renrum anläggningen är alltid begränsad till användare med lämplig utbildning) i designen och beställning av en mask, rån rengöring, och fotolitografi. När en mall erhålls, kan den användas utanför renrummet upprepade gånger, förutsatt att den hålls i en stängd skål och försiktighet iakttas för att utesluta damm. - Förbered en blandning av ~ 100 ml PDMS (silikon elastomer bas) och ~ 10 ml av tvärbindare (härdare) i en engångsplastkopp. Bryt av spetsen av en engångspipett att hantera de viskösa PDMS lättare. Väg PDMS i plastmugg som pipettering är inte korrekt.
- Rör om blandningen väl. Avlägsna de många luftbubblor genom avgasning i en vakuumexsickator (ca 20min). Eftersom koppen placeras under vakuum, kommer bubblorna initialt växa i storlek, vilket ökar volymen av blandningen kraftigt. Styr vakuum som appliceras och se till att koppen är tillräckligt djup för att undvika spill.
- Häll en stor droppe av avgasade PDMS blandning i ett glas petriskål (150 mm x 20 mm) och tryck skivan på PDMS med mallen uppåt. Detta undviker bubblor att bli instängd under skivan. Instängda bubblor kommer att expandera och kan luta skivan när skålen placeras i ugnen. Nu hälla avgasade PDMS på mallen ovanpå skivan tills den är täckt med ca 5-8 mm PMDS. Om en luftbubbla uppstår försiktigt bort den med en pipettspets.
- Baka de PMDS i en ugn vid 60 ° C under 3 h. Se till att skålen är nivån.
- Använd en ny skalpellblad för att skära ut en PDMS-block innehållande de formade kanalstrukturer. Den utskurna blocket måste passa på en täck (se steg 4.1.9).
- Skala ut utskuren kvarter från skålen och pless den på en bit av ren aluminiumfolie.
Obs: De tvärbundna PMDS block kan hållas under några månader. På en enda skiva, det finns 6 uppsättningar av flödeskanaler (Figur 1A), så 6 PDMS block kan framställas i en enda PDMS gjutning. Efter att klippa ut alla sex av PDMS block, rena lösa bitar av PDMS, men annars inte bort de återstående PDMS innan du häller och tvärbindning nästa omgång, vilket kommer att kräva endast omkring hälften av PDMS som används för den första omgången. En bra mall kan pågå ett par år. - Använd en hålslag att borra genom PDMS block i en rak rörelse (Figur 1B). Börja på sidan av kanalspåren. Var noga med att ta bort utstansade bit PDMS. Upprepa detta för alla åtta hål av de fyra kanaldesignen.
- Placera PDMS blocket kanalsidan nedåt på en ny och skrynkelfri bit aluminiumfolie. Lagra blocket i upp till några månader i en torr låda, helst i en exsickator.
- Skjut slangen (0,25 mm ID, 0,76 mm OD) ca en tredjedel in i det stansade hålet med hjälp av en pincett (Figur 1C). Klipp slangen skev för att underlätta införandet. Lämnar rören tillräckligt länge för att nå SUV reservoaren och sprutpump, respektive. Efter att ha placerat den sammansatta chip på mikroskop, skär rören igen om de är för långa.
- Att ansluta till sprutorna i pumpen, skär en kort bit av större silikonslang (0,51 mm ID, 2,1 mm OD) och sätt det tunnare röret i en sida. Detta färdig att använda block av PDMS kan hållas under några månader.
2. Cover Rengöring
- Rena täckglas enligt protokollet beskrivet i Karatekin och Rothman 25 användning av en stark oxiderande blandning av svavelsyra (H 2 SO 4) och väteperoxid (H 2 O 2). Utför den här proceduren i ett renrum, och iaktta lämpliga försiktighetsåtgärder. Alternativt kan du använda ett renrum rengöras coverslip som är kommersiellt tillgängliga (se Material List).
3. Framställning av v-SUV innehållande både Lipid och innehåll etiketter

Figur 2. Schematisk av SUV beredning. Lipider blandas i ett glasrör (1) och lösningsmedlet avdunstas för att bilda en lipidfilm genom att rotera röret i ett vattenbad (2). Kvarvarande spår av lösningsmedel avlägsnades under högt vakuum (3). Lipidfilmen hydratiseras i beredning buffert innehållande tvättmedel och protein medan virvlas (4). Om innehållet färgämne ska kapslas in, ingår det i detta steg, liksom i utspädningssteget (5). Utspädning av koncentrationen för tvättmedlet under dess kritiska micellkoncentration leder till liposombildning. Tvättmedel dialyseras bort under natten (6). För t-SUV för SBL bildning inklusive NBD-PE (grön), är vesiklar flöt i en densitetsgradient och samlas pågränsytan mellan två skikt (7a). För att separera v-SUV med inkapslad innehåll markör från fritt färgämne provet körs över en storleksuteslutningskolonn och samlas i 0,5 ml fraktioner (7b). Klicka här för att se en större version av denna siffra.
- Förbereda 4 L av rekonstitution buffert, innehållande 25 mM HEPES-KOH, 140 mM KCl, 100 ^ M EGTA och 1 mM DTT, pH 7,4. Använda de flesta av bufferten för dialys och 100 ml för andra åtgärder.
Obs: Andra buffertar kan användas, men det är viktigt att hålla buffert osmolaritet konsekvent genom hela experimentet och välja förhållanden under vilka proteiner är funktionella. För framställning av v-SUV innehållande lipid etiketter endast och t-SUV, följa Karatekin och Rothman 25. - Som lipid lager lagras vid -20 ° C i kloroform eller kloroform: metanol (2: 1, v / v), låt flaskorna når rumstemperatur innan den öppnas för attundvika kondens.
- Skölj ett glasrör med kloroform för att avlägsna spårmängder av detergent och blanda lipiderna vid det önskade förhållandet med en slutlig mängd av 1 | j, mol i en blandning av kloroform och metanol (2: 1, volym / volym). Använd endast glassprutor / rör för att hantera organiska lösningsmedel / lösta lipider.
Notera: De lipider som används här är POPC: SAPE: DOPS: Kolesterol: PEG2000-DOPE: DiD vid ett molförhållande av 57,4: 15: 12: 10: 4,6: 1 för v-SNARE innehållande vesiklar och POPC: SAPE: DOPS: Kolesterol : hjärna PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5) för t-SNARE SUVs. Se Material för mer information. Andra lipidkompositioner kan användas. - Indunsta lösningsmedlet under en försiktig ström av kväve, eller i en roterande indunstare. Doppa spetsen av röret i ett vattenbad (37 ° C) upphettas över den högsta smälttemperaturen för de lipider för att ge en homogen lipidfilm och för att undvika stora förändringar i temperatur medan avdunstning av lösningsmedlet. Börja vakuum vid omkring300 mbar tills lipidfilm bildas, sedan fortsätta under ~ 2 minuter vid högsta möjliga vakuum i rotationsindunstare.
- Ta bort glasröret och packa in den i aluminiumfolie för att undvika ljusexponering och avlägsna kvarvarande spår av lösningsmedel genom att placera röret i en torkapparat under högt vakuum under minst 2 timmar. Detta steg kan också köra över natten.
- Att rehydrera den torkade lipidfilmen, framställa en blandning av detergent (n-oktyl-β-D-glukopyranosid, OG), protein (v-snaror) och SRB i rekonstituering buffert (500 | j, l slutlig volym). Hålla slutkoncentration detergent ~ 2 gånger över den kritiska micellkoncentrationen (CMC, 20-25 mM för OG) och justera den slutliga volymen, så att tvättmedlet till lipid-förhållandet är> 10. Upplösa Sulforhodamine B (SRB) pulver i den proteintvättmedelslösning för att erhålla en slutlig koncentration av 10-50 mM SRB genom att försiktigt skaka lösningen. Justera osmolaritet rekonstitutionsbuffert när du lägger höga koncentrationer av SRB genom att minska samncentration av kaliumklorid i enlighet därmed.
Notera: Lipid-till-proteinförhållande (LP) för t-snaror är hög (LP ~ 20000, ~ 70 t-snaror per sq mikron 25.). Betydligt högre t-SNARE densiteter i SBL kan leda till inaktiva proteinaggregat med kraftigt minskad fusions priser 17,24. LP för v-snaror är mycket lägre (LP ~ 200, ~ 7000 v-snaror per ^ m 2), nära de v-SNARE densiteter som hittades i den synaptiska vesiklar 46,47. För att säkerställa proteinfunktion efter rekombinant uttryck och rening av v- och t-snaror det är bra att göra en enkel bulkfusionsanalys 48 innan du går igenom alla steg på den inre vesikelfusion analys. - Skaka försiktigt förvärmda glasrör innehållande den torkade lipidfilmen under tillsats av protein tvättmedel-SRB-lösning från steg 3,6. Försök att undvika att skapa bubblor. Fortsätt att skaka i 15 minuter vid 37 ° C.
- Späd detergent fem gånger genom tillsats av rekonstitutionbuffert innehållande SRB (2 ml, 2,5 ml slutlig volym) medan snabbt vortexa att undvika koncentrationsgradienter. Fortsätta att skaka i 15 min till 1 h vid 37 ° C.
Obs: Längre inkubation ökar protein beredning effektivitet. Den snabb utspädning minskar tvättmedels mängd som ligger under CMC och leder till bildning av små liposomer. - Dialysera vesikelsuspensionen först mot ~ 1 L av rekonstitution buffert under 1-2 timmar vid rumstemperatur och sedan mot 3 L av rekonstitution buffert över natten vid 4 ° C med 4 g polystyren adsorbent, med användning av ett 20.000 MWCO dialysrör eller kassett. Använd olika bägare för dialys av vesiklar med och utan SRB att förhindra korskontaminering.
- Jämvikt en gelfiltreringskolonn med beredning buffert. Kör vesikelsuspensionen genom kolonnen för att separera fritt SRB från v-SUVs med inkapslat SRB. Använd beredning buffert utan SRB som elueringsmedel gång hela provet hargick in i kolonnen. Samla v-SUV i 0,5 ml fraktioner.
- Kontrollera framgångsrik SRB inkapsling vid själv kylda koncentrationer genom att mäta SRB fluorescens före och efter tillsats av rengöringsmedel till en delmängd av blåsor 16. Med hjälp av en fluorescensspektrometer, excitera provet vid 550 nm och skanna emissions SRB mellan 570 nm och 630 nm. Förvänta sig en 4-8-faldig ökning i SRB fluorescens beroende på det inkapslade beloppet vid tvättmedels tillsats som membranet är solubiliseras och den frigjorda SRB utspädes.
- Karakterisera lipid och proteinåtervinning med användning av fluorescensspektroskopi 24 och SDS-PAGE-gelelektrofores, respektive. Använd en känslig färgningsmetod, särskilt för hög LP t-SUV-prover (se Materials List). Typiskt ca 50% av både lipid och protein ingång försvinner under framställningen resulterar i en LP nära det nominella värdet.
- Karakterisera SUV storlekar med dynamisk ljusspridning 24 eller elektron microscopy 48. Store SRB innehållande v-SUVs vid 4 ° C upp till ~ 3-4 dagar. Får ej frysas, som frysning och upptining bryter membranet och släpper inkapslad SRB.
4. SUV-SBL Fusion analys för att övervaka Lipid utsättningen först
- Bildandet av den bundna tvåskikts stöd inom mikroflödessystem flödeskanalerna
- Placera PDMS blocket (steg 1,11) under högt vakuum under minst 20 min före experimentet för att avlägsna lösta gaser. Detta minskar avsevärt risken för luftbubblor i mikroflödessystem kanaler under fusionsexperiment.
- Slå på mikroskop setup och värma scenen och provhållaren (fig 3 och 4) till den önskade temperaturen.
- Filtrera rekonstitution buffert som användes för utspädning av de vesiklar genom ett filter med 0,45 | j, m eller mindre porstorlek.
- Späd ~ 30 pl av NBD-PE märkt t-SUV eller proteinfritt (PF-SUV) kontroll liposomer (lagerLösningen 0,5-1 mM lipid) med ~ 60 pl buffert. Den slutliga koncentrationen är inte kritisk här.
- Degas denna blandning med användning av en 3 ml spruta. Tryck ut det mesta av luften ovanför provet medan du håller sprutan vertikalt. Täta (nålfri) spets med hjälp av paraffin film och skapa ett vakuum genom att dra ner kolven. Knackar du på sprutan för att påskynda avgasning av lösningen. Upprepa denna process några gånger tills inga fler bubblor uppstår när vakuum appliceras.
- Använda en hypodermisk nål med en diameter som är något större än det rör som är fäst vid PDMS-block för att stansa ett hål i locket på ett mikrocentrifugrör. Vara säker på att stansas ut bit plast inte är på insidan av röret eftersom det kan täppa till slangen som är ansluten till mikroflödeskanalinloppet senare.
- Fyll avgasade SUV lösningen i mikrocentrifugrör med ett hål i locket och placera den i hållaren på mikroskop scenen till jämvikt till den inställda temperaturen.
- pless en tidigare rengjorts täckglas (avsnitt 2) i en plasma renare och köra luftplasma under ca 5 min. Sätt plasmabehandlade täck (behandlade sidan uppåt) på toppen av några luddfria vävnader fungerar som en kudde.
- Ta bort aluminiumfolie från avgasade PDMS blocket och placera blocket på toppen av täck. Vid återanvändning av PDMS blocket sätta på och ta en bit tejp på kanalsidan för att rengöra den. Tryck ner PDMS blocket på täck med hjälp av en pincett för att få det att fästa, men tryck inte för hårt eftersom glaset kan gå sönder.
- Placera den monterade flödescell på mikroskop scenen och ansluta slangen till SUV reservoaren och sprutpump, respektive. Tejpa täckglas till scenen (figur 3B).
- Börja aspire SUVs vid 3 | j, l / min tills lösningen fyller kanalerna fullständigt (~ 2,25 mm / sek för 75 ^ m x 300 | im kanaltvärsnittet). När lösningar för alla kanaler börjar flytta upp the slangen på utgångssidan, minska flödet till 0,5 | j, l / min och inkubera under 30-45 min.
Notera: Tiden mellan plasma behandling av objektglas och som strömmar SUVs i kanalen bör inte överstiga 10 till 20 min som effekten av plasmabehandlingen är övergående. - Kontrollera kanalerna för eventuella läckor. Använd en 10-20x luft mål att observera NBD-PE fluorescens från NBD-PE innehållande T- eller PF-SUV. Excite NBF fluoroforer använder 488 nm laser. Läckor är svåra att upptäcka med hjälp av ljus fältbelysning.
- Sätt ett rör med avgasad beredning buffert i hållaren och låt temperaturen jämvikt som snabba förändringar i temperatur kan orsaka defekter i SBL. Stoppa flödet och vänta ~ 1 min för att säkerställa att flödet har stannat helt och inga luftbubblor kommer sugas in i röret innan du slår inloppsrören till den avgasade buffert.
- Skölj alla kanaler med avgasad buffert för att tvätta bort obundna SUV.
- Byt till en higher förstoring TIRF mål (60X, olja, NA 1,45-1,49) och kontrollera att biskiktet ser homogent och är fri från uppenbara storskaliga defekter som mörka fläckar eller lipid tubuli sträcker ut från SBL.
- Kontrollera fluiditeten av dubbelskiktet
- Om en särskild FRAP enhet inte är tillgänglig, eller om en FRAP sekvensen inte kan programmeras, sedan kvalitativt testmembran fluiditet på följande sätt.
- Stänga fältet membranet till en liten storlek (~ 40 | j, m i diameter) och justera 488 nm excitation ljusintensitet genom programvaran (20-80 ^ W, eller 15 till 60 nW / xm 2) att bleka NBD-PE fluorescens i den exponerade område signifikant, men inte helt. För en vätska biskikt, vid steady state, bör fluorescensintensiteten i mitten av den exponerade ytan vara lägre än vid kanterna, som intakta NBD-PE-molekyler in i exponerade området och sprida ett visst avstånd före blekning. I motsats, om ytan vidhäftande SUVs misslyckades att brista, eller av någon annan anledning stöds tvåskiktsmembran är inte vätska bör alla fluoroforer i det utsatta området bleka.
Obs: Laserintensitetsvärden i denna och senare steg ges som en grov utgångspunkt och bör optimeras för en given uppsättning av betingelser. - Stoppa belysning och starta den igen några minuter senare för att kontrollera resultaten av de stationära mätningar
- Stänga fältet membranet till en liten storlek (~ 40 | j, m i diameter) och justera 488 nm excitation ljusintensitet genom programvaran (20-80 ^ W, eller 15 till 60 nW / xm 2) att bleka NBD-PE fluorescens i den exponerade område signifikant, men inte helt. För en vätska biskikt, vid steady state, bör fluorescensintensiteten i mitten av den exponerade ytan vara lägre än vid kanterna, som intakta NBD-PE-molekyler in i exponerade området och sprida ett visst avstånd före blekning. I motsats, om ytan vidhäftande SUVs misslyckades att brista, eller av någon annan anledning stöds tvåskiktsmembran är inte vätska bör alla fluoroforer i det utsatta området bleka.
- Programmera en FRAP-sekvens för en mer kvantitativ mätning, om möjligt. Se kompletterande filer och motsvarande legend för mer information.
Notera: Ibland SUV kommer att följa på glaset täck, men misslyckas med att brista och bilda en vätska dubbelskiktet. Om detta inträffar, skölj kanalerna med avgasad beredning buffert innehållande 10 mM Mg 2 + för att hjälpa stöds tvåskiktsmembran bildning. Använd fluorescens blekning att bedöma tvåskiktsmembran fluiditet som i 4.2. När väl en vätske tvåskiktsmembran bildas, skölj med Mg2 + -fri reconstitution buffert.
- Om en särskild FRAP enhet inte är tillgänglig, eller om en FRAP sekvensen inte kan programmeras, sedan kvalitativt testmembran fluiditet på följande sätt.
- Införande av v-SUVs in i mikroflödessystem flödeskanaler
- Degas Reconstitution Buffer och använda det för att späda ut v-SUV stamlösning med en faktor av ca 10 mars - 10 maj beroende på v-SUV förrådskoncentration den. Sikta på en utspädning som resulterar i cirka 10-100 fusions händelser inom 60 sekunder i synfältet.
Obs: Alltför många fusioner ökar bakgrundsfluorescens (eftersom varje insättningar händelse LR-PE eller gjorde lipid etiketterna i SBL) och göra upptäckt och analys av fusionshändelser svårt. I kontrast, ett fusionsfrekvens som är för låga resulterar i dålig statistik eller kräver förvärv av många fler filmer. För en v-SUV koncentration av 0,1 mM lipid, börja med att späda 5 pl SUV lager i 995 pl beredning buffert och sedan späd 5-50 pl av detta i 950-995 pl beredning buffert. - Låt temperaturen jämvikt innan du stoppar flödet och INSERting inloppsröret i den utspädda v-SUV-lösning.
- Justera TIRF vinkel och polarisationen enligt följande.
- Efter inställning av den önskade polarisationen genom att rotera excitationsstrålen, justera lutningen av spegeln kommenderade positionen för strålen via en stegmotor genom programvaran. Sakta flytta positionen för laserstrålen från mitten av mål på det bakre fokalplanet till en sida utanför centrum. Observera ljuset från målet vid den främre sidan komma ut med en ökande vinkel i förhållande till det mål axel som den positionen flyttas längre off-center.
- Notera motorposition när den utgående strålen först försvinner in i mål, det vill säga, när TIR först uppnås.
- Sakta hålla flytta strålen ställning ytterligare excentriskt under övervakning av fluorescens från ytan. Notera motorposition när ytan fluorescens försvinner när strålen har flyttat alltför långt utanför centrum.
- Välja en balk positionering n mellan de två gränser fastställts ovan. För bästa signal-brusförhållande och mer signalförstärkning vid fusion, välja ett grunt penetrationsdjup (balkläge närmare kanten av målet) som fortfarande ger jämn belysning av View.
Obs: Det är bäst att hålla samma TIR balkläge (samma penetrationsdjup) för alla experiment efter inställningar optimeras. Se till att vrida polarisationen inte leda till förändringar i balkläge.
- Flödes v-SUVs in i kanalen vid en flödeshastighet av 2 ul / min, vilket motsvarar en genomsnittlig linjär flödeshastighet på ~ 1,5 mm / sek för ett tvärsnitt av 75 ^ m x 300 | j, m. Byt till inställningarna excitation / emissions att övervaka lipidblandning (LR eller gjorde bara).
- Degas Reconstitution Buffer och använda det för att späda ut v-SUV stamlösning med en faktor av ca 10 mars - 10 maj beroende på v-SUV förrådskoncentration den. Sikta på en utspädning som resulterar i cirka 10-100 fusions händelser inom 60 sekunder i synfältet.
- Observation fusion mellan enskilda v-SUV och SBL
.jpg "/>
Figur 3. Den experimentella pTIRF setup. (A) Schematisk representation av en v-SUV och en t-SBL på ett glassubstrat. False-color TIRFM bilder av enkel SUV-SBL fusions händelse som visar lipid dockning (1) och frisättning av lipid färgämnet i SBL (2) följt av blekning och minskning av fluorescensintensitet (3). Den totala fluorescensintensiteten (summa av pixelvärdena i x 5,3 | im box 5,3 | j, m) signalen visas. (B) Den täckglas bunden till PDMS-block är tejpad på den uppvärmda scenen. Inloppsrör för mikroflödessystem kanaler tar ut prov från röret i provmetallhållare (höger) aspireras genom sprutpumpen (till vänster). Under pumpen är den dubbla enhet utsläpp. Klicka här för att se en större version av denna siffra.
oad / 54349 / 54349fig4.jpg "/>
Figur 4. Schematisk ritning av försöksuppställningen. Den evanescenta vågen skapas vid glasbuffert gränssnitt i mikroflödeskanalen. SBL bildas på glaset och v-SUV aspireras från provmetallhållaren (överst till höger) genom kanalen i sprutan (uppe till vänster). M, spegel; DM, dikroisk spegel; L, lins; F, filter; P, polarisationsfilter. Klicka här för att se en större version av denna siffra.
- Kontinuerligt excitera och övervaka v-SUV fluorescens med hjälp av en 561 nm (för LR-PE) eller 638 nm (för gjorde) laser beroende på fluoroforen införlivas i v-SUV. Som V-SUV når flödeskanalen och docka på och säkring med SBL börjar bakgrundsfluorescenssignalen att ackumuleras.
- Justera excitation laserintensiteten via programvaran kontinuerligt bleka bakgrundenfluorescens sådan som lätt kan observeras vid steady state nya dockning och fusion händelser.
Obs: Om blekning är för långsam, kommer bakgrundsfluorescens vara för hög. Om blekning är för snabb, kommer signaler från dockad SUV och från fluoroforer frigörs i SBL blekna snabbt förkorta fönstret under vilken fusion av dockade SUV kan upptäckas eller en fluorofor kan spåras. För LR-PE exciteras vid 561 nm, 2,5-7,5 mW makt att belysa en fim diameter cirkel 190 (100-250 nW / um 2) är ett rimligt värde för att starta. För 638 nm excitering av gjorde 0,8-1,6 mW makt över en diameter cirkel 190 fim (30-60 nW / um 2) kan användas för inledande tester. - Förvärva flera filmer på olika positioner i en given kanal. Kolla NBD-PE fluorescens för att verifiera att SBL inte har defekter på dessa positioner. Förvärva fullformat filmer på den högsta nivån (~ 50 bilder / sek) eller en beskuren region av intresse vid högre bildhastighet (upp till~ 100 Hz) under 60 sek.
- Flytta till en annan mikroflödessystem kanal och upprepa registreringarna under andra förhållanden. Inkludera negativa kontroller såsom ett proteinfritt SBL eller SUVs, tillsätt den lösliga cytoplasmiska domänen av v-SNARE VAMP2 (CDV) som en inhibitor eller behandla v-SUV med tetanus neurotoxin (30 min vid 37 ° C) i en eller flera av kanalerna på samma täckglas.
- Städa upp och PDMS återvinning
- I syfte att återanvända PDMS-block, skölj de mikrofluidiska kanaler med ~ 200 | il av 70% etanol vid en flödeshastighet av 5 | j, l / min. Slutligen, aspirera luft genom slangen och kanaler.
- Lossa PDMS blockera försiktigt från täck genom att klämma det något. Placera den på en ren bit aluminiumfolie. Förvara i en vakuumexsickator. Den kan återanvändas flera gånger.
- För en mer grundlig rengöring skölja kanalerna med tvättmedel eller natriumhydroxidlösning före 70% etanol. Alternativt, ta bort alla rör tillbaka m PDMS och Sonikera blocket för 30 minuter i isopropanol, före torkning och till ny slang.
5. SUV-SBL Fusion analys för att övervaka Lipid och innehåll Release samtidigt
- För tvåfärgade övervakning av samtidig lipid och lösliga innehåll släppa följer samma steg som i avsnitt 4, förutom användning liposomer märkta med både en lipid (gjorde) och ett lösligt innehåll (SRB) etikett, som beskrivs i avsnitt 3. SRB är inkapslat vid 10 mM och är initialt mycket själv släcktes.
- Använda den konfiguration som visas schematiskt i fig 4, hetsas SRB och gjorde fluorescens samtidigt med hjälp av 561 nm och 638 nm lasrar, respektive. En dikroisk spegel (640 nm) delar utsläpp i två strålar som går genom en kort (595/50 nm) och en lång (700/75 nm) våglängdsfilter för att detektera SRB och gjorde utsläpp respektive. De två emissionsstrålar projiceras sida vid sida på den EM-CCD-chip.
- Analys av FRAP uppgifter
- Använd MATLAB programmet tillhandahålls i den kompletterande information för att uppskatta lipid diffusionskoefficienten, D. Programmet läser en lista över OME-TIFF-filer 49, detekterar blekt område, tomter värdena medelpixel i blekt område som en funktion av tiden, och passar den resulterande återhämtningskurvan till en modell av Soumpasis 50 för att extrahera återhämtningstid,
 , Där w är radien hos den blekta cirkeln.
, Där w är radien hos den blekta cirkeln.
Obs: Ett MATLAB program tidigare 25 tillhandahålls för analys av FRAP filmer med Nikon ND2 filer. Det nuvarande programmet läser OME-TIFF-filer 49, eftersom de flesta filformat kan lätt omvandlas till OME-TIFF. En kvantitativ analys av FRAP uppgifter är enklaste och mest exakta vid blekning är ögonblicklig, den bleached område är en cirkel, och blekning under utläsning är försumbar. Även om dessa kriterier inte är strikt uppfyllda i de enkla FRAP mätningar som beskrivs här, kan erhållas en rimlig uppskattning av diffusionskoefficienten. För en mer exakt uppskattning, använder spårning av enstaka lipid färgämnen (avsnitt 6.3).
- Använd MATLAB programmet tillhandahålls i den kompletterande information för att uppskatta lipid diffusionskoefficienten, D. Programmet läser en lista över OME-TIFF-filer 49, detekterar blekt område, tomter värdena medelpixel i blekt område som en funktion av tiden, och passar den resulterande återhämtningskurvan till en modell av Soumpasis 50 för att extrahera återhämtningstid,
- Analys av dockningshastighet, fusion hastighet och dockning mot fusionsfördröjningstider
- Använd ImageJ för att öppna en film för att analysera och justera ljusstyrka och kontrast för att tydligt identifiera vesikler dockning och fixerings händelser. Starta SpeckleTrackerJ 26 plugin. Se Smith et al. 26 och onlinedokumentation för SpeckleTrackerJ för instruktioner för användning.
- Identifiera alla nya dockade SUV i SpeckleTrackerJ. För att skilja SUV som docka tätt från dem som studsar SBL införa en minimidocknings varaktighet några ramar. Spara spår, och upprepa för alla filmer.
Obs: För dockung takt, är allt som betyder något för att identifiera när en SUV dockad, så endast den första bildrutan där SUV dockad behöver registreras i dessa spår. Resten av spåren kommer inte att beaktas i analysen. Emellertid, auto-tracking varje SUV tills den bleker eller säkringar hjälper markera SUVs som redan har spårade. - Identifiera alla fixerings blåsor. För dessa bör spåren omfatta alla ramar från den första bildrutan en SUV dockad tills den första bildrutan i vilken fusion är framgår av en plötslig ökning av fluorescens av det spårade plats. Varaktigheten för dessa spår används för att beräkna de dockning-to-fusion fördröjningar. Spara alla spår. Upprepa för alla filmer.
- För parti analys av docknings eller fusionsdata sammanställa en lista över bana filer från de relevanta filmerna och köra MATLAB program som medföljer Karatekin och Rothman 25. Följ instruktionerna som tillhandahålls tillsammans med programmen.
Obs: Programmen rita cumulative dockning och fusion händelser som en funktion av tiden, baserat på uppgifter ur banan filer. Dockning och fusion priser beräknas från sluttningarna av dessa tomter. För fusionsuppgifter, är dockning mot fusion förseningar även beräknas och deras fördelning ritas som en överlevnads tomt, det vill säga sannolikheten att fusions ännu inte har inträffat en given fördröjning efter dockning.
- lipid diffusivitet
- För varje fusion händelse, spåra enskilda fluorescerande lipider när de blir urskiljas när de har diffunderat tillräckligt bort från fusionsstället (Figur 6). Använd SpeckleTrackerJ för spårning, och spara spåren för vidare analys med hjälp av MATLAB. Beroende på storleken av fixerings vesikeln, typiskt 3-30 enskilda fluoroforer kan spåras. Eftersom längre spår är önskvärda för beräkning av
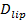 , Försöka spåra enskilda molekyler som längesom möjligt, manuell korrigering av banor om det behövs.
, Försöka spåra enskilda molekyler som längesom möjligt, manuell korrigering av banor om det behövs. - Beräkna medelkvadrat förskjutning (MSD) för ensamstående lipid markör banor som varar> 40-50 ramar (ca ~ 1,5 sek). Använda MSD att beräkna lipid diffusionskoefficienten, . Se et al. Smith 26 och et al. Stratton 27 för mer information.
- För varje fusion händelse, spåra enskilda fluorescerande lipider när de blir urskiljas när de har diffunderat tillräckligt bort från fusionsstället (Figur 6). Använd SpeckleTrackerJ för spårning, och spara spåren för vidare analys med hjälp av MATLAB. Beroende på storleken av fixerings vesikeln, typiskt 3-30 enskilda fluoroforer kan spåras. Eftersom längre spår är önskvärda för beräkning av
- Enda lipid färgintensitet, SUV-SBL intensitet reduktionsfaktor, och vesikelstorlek
- Mäta summan av pixelvärdena i en lipid markör i en 3 x 3 bildpunkter (0,8 ^ m x 0,8 ^ m) området runt markör för ~ 15 bilder före och efter markeringen blekmedel i ett enda steg. Subtrahera bakgrundsintensiteten som genomsnittet av de post-blekmedels ramar från pre-blekmedel intensitet snittas innan blekning för att erhålla intensiteten hos den spårade lipid markör. Upprepa mätningen för så många markörer som praktiskt för en given film.
- Rita fördelningen av enstaka lipid etikettintensitet,
 Och montera en Gauss att uppskatta medelvärdet.
Och montera en Gauss att uppskatta medelvärdet. - Beräkna fördröjningen mellan när fusion inträffat och när banan för en enda lipid markör släpptes i så fall slutade i ett enda steg blekning. Erhålla blekningstiden för en fluorofor i SBL,
 , Genom att plotta överlevande funktion av förseningarna och passar till en exponentiell.
, Genom att plotta överlevande funktion av förseningarna och passar till en exponentiell.
Obs! Eftersom de polariserade excitation fält par svagare till fluoroforema på en SUV, är vanligtvis mycket långsammare 27 blekningstiden för en SUV. - Uppskatta
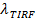 Intensiteten reduktionsfaktor för en lipid färgämne när SUV i förhållande till när det är i SBL efter Stratton 27 (tp_upload / 54349 / 54349eq8.jpg "/> är intensiteten hos en enda färgämne i SUV).
Intensiteten reduktionsfaktor för en lipid färgämne när SUV i förhållande till när det är i SBL efter Stratton 27 (tp_upload / 54349 / 54349eq8.jpg "/> är intensiteten hos en enda färgämne i SUV).
Obs: Om blekning var försumbar, skulle vara lika med den dockade intensitet  (punkt (1) i figur 3A, högra panelen), dividerat med det totala intensiteten uppnås efter alla fluoroforer sätts in SBL på fusion (streckad linje märkt
(punkt (1) i figur 3A, högra panelen), dividerat med det totala intensiteten uppnås efter alla fluoroforer sätts in SBL på fusion (streckad linje märkt  i figur 3). Men på grund av snabb blekning i SBL, är normalt inte uppnås och en noggrann uppskattning av kräver montering av frisättningskinetik till ett uttryck 27 som inkl udes både (Se 6.4.3) och . Separata uttryck finns i Stratton et al 27 för de fall då por-begränsade och diffusion begränsad frisättningskinetik. bästa passform fall varierar från fall till fall (se 6.5.1 för fallet med por begränsade kinetik).
i figur 3). Men på grund av snabb blekning i SBL, är normalt inte uppnås och en noggrann uppskattning av kräver montering av frisättningskinetik till ett uttryck 27 som inkl udes både (Se 6.4.3) och . Separata uttryck finns i Stratton et al 27 för de fall då por-begränsade och diffusion begränsad frisättningskinetik. bästa passform fall varierar från fall till fall (se 6.5.1 för fallet med por begränsade kinetik). - Beräkna vesikler området för enskilda händelser från 27
 , var är den dockade SUV intensitet, är medelintensiteten av en enda lipid färgämne i SBL, är intensiteten reduktionsfaktor för en lipid färgämne när det är i SUV i förhållande till när det är i SBL, ochation ett "src =" / filer / ftp_upload / 54349 / 54349eq12.jpg "/> är den kända ytdensitet av lipid färgämnen.
, var är den dockade SUV intensitet, är medelintensiteten av en enda lipid färgämne i SBL, är intensiteten reduktionsfaktor för en lipid färgämne när det är i SUV i förhållande till när det är i SBL, ochation ett "src =" / filer / ftp_upload / 54349 / 54349eq12.jpg "/> är den kända ytdensitet av lipid färgämnen.
- Fusion pore egenskaper
- Förutsatt release är por begränsad, passa lipid etikett frisättningskinetik till 27
 , var
, var  är den dockade SUV intensiteten strax före fusion och de andra parametrarna är såsom tidigare beskrivits. Använd värdet av erhållits i 6.4.3 som en fast parameter och extrahera de bästa passningsberäkningar för och
är den dockade SUV intensiteten strax före fusion och de andra parametrarna är såsom tidigare beskrivits. Använd värdet av erhållits i 6.4.3 som en fast parameter och extrahera de bästa passningsberäkningar för och  .
. - Uppskatta bråkdel av tiden pore är öppen, P 0, är antar fördröjning av lipid frisättning på grund av pore flimmer 27:60;
 =
=  , Där A ves är vesikler området (avsnitt 6.4.5), b poren höjd (vanligen anses vara ~ 15 nm), rp ≈ 3 nm är den effektiva porradien som ses av diffunderande lipid etiketter och inkluderar halv dubbelskiktet tjocklek (~ 2 nm), är lipid diffusiviteten (beräknat på 6,3), och är det dags för lipider för att frigöras från SUV till SBL (från 6.5.1).
, Där A ves är vesikler området (avsnitt 6.4.5), b poren höjd (vanligen anses vara ~ 15 nm), rp ≈ 3 nm är den effektiva porradien som ses av diffunderande lipid etiketter och inkluderar halv dubbelskiktet tjocklek (~ 2 nm), är lipid diffusiviteten (beräknat på 6,3), och är det dags för lipider för att frigöras från SUV till SBL (från 6.5.1). - För att bekräfta att en nominell P 0> 1 indikerar en helt öppen por P 0 = 1, passar intensiteten tidsförloppet att ekvation 4 av Stratton et al. 27, de förväntade kinetiken för en permanent öppen por. Detta passar bör vara bättre än fitting uttrycket i 6.5.1 för en permanent öppen por.
- Förutsatt release är por begränsad, passa lipid etikett frisättningskinetik till 27
 , Där w är radien hos den blekta cirkeln.
, Där w är radien hos den blekta cirkeln.  , Genom att plotta överlevande funktion av förseningarna och passar till en exponentiell.
, Genom att plotta överlevande funktion av förseningarna och passar till en exponentiell.  (punkt (1) i figur 3A, högra panelen), dividerat med det totala intensiteten uppnås efter alla fluoroforer sätts in SBL på fusion (streckad linje märkt
(punkt (1) i figur 3A, högra panelen), dividerat med det totala intensiteten uppnås efter alla fluoroforer sätts in SBL på fusion (streckad linje märkt  i figur 3). Men på grund av snabb blekning i SBL,
i figur 3). Men på grund av snabb blekning i SBL,  , var
, var  , var
, var  är den dockade SUV intensiteten strax före fusion och de andra parametrarna är såsom tidigare beskrivits. Använd värdet av
är den dockade SUV intensiteten strax före fusion och de andra parametrarna är såsom tidigare beskrivits. Använd värdet av  =
=  , Där A ves är vesikler området (avsnitt 6.4.5), b poren höjd (vanligen anses vara ~ 15 nm), rp ≈ 3 nm är den effektiva porradien som ses av diffunderande lipid etiketter och inkluderar halv dubbelskiktet tjocklek (~ 2 nm),
, Där A ves är vesikler området (avsnitt 6.4.5), b poren höjd (vanligen anses vara ~ 15 nm), rp ≈ 3 nm är den effektiva porradien som ses av diffunderande lipid etiketter och inkluderar halv dubbelskiktet tjocklek (~ 2 nm), Subscription Required. Please recommend JoVE to your librarian.
Representative Results
SBL Kvalitet
Det är mycket viktigt att kontrollera kvaliteten och fluiditet hos SBL före fusionsexperimentet. Fluorescensen vid botten, glassidan av en mikroflödessystem kanal bör vara enhetlig, utan några uppenbara defekter. Om en luftbubbla passerar även om kanalen, vanligtvis lämnar synliga ärr på SBL. Om det finns sådana storskaliga ärr / fel, inte använda den kanalen. Ibland SUV kan fastna på substratet men misslyckas med att brista och bilda en kontinuerlig SBL. Om så är fallet, kan fluorescens fortfarande verkar mycket jämn, men kommer inte att återhämta sig efter blekning ett litet område. Presentation 10 mM Mg 2+ ofta hjälper att lösa denna fråga. Andra medel kan införas för att hjälpa spricker SUVs, såsom andra tvåvärda katjoner 51, en polymerlösning 52, eller osmotisk chock 25.
temperaturförändringar leda till förändringar av SBL område eftersom medelytan upptas av en lipid beror på temperaturen. Öka temperaturen eller minska den med 5-10 ° C eller mer kan leda till kraftig område går till rör extruderade från ytan, eller mörka, SBL-fria fläckar som förekommer på ytan, respektive 53. Det är därför viktigt att hålla temperaturvariationerna till ett minimum.
fig 5 för en SBL som var relativt defektfritt och flytande.

Figur 5. FRAP att undersöka SBL fluiditet. En sekvens av bilder som visar ett belyst område som avgränsas av membranet stängda fält (diameter 49 nm) före (första bilden) och efter blekning med en stark ljuspuls. Området avbildas vid låg ljusexponering för att följa fluorescens återhämtning genom diffusion av NBF märkta lipider. Diagrammet visar den normaliserade fluorescens som en funktion av tiden för flera kanaler från olika täckglas som indikerar den höga nivån av metodens reproducerbarhet. Lipidkompositionen av t-SBL var POPC: SAPE: DOPS: Kolesterol: hjärna PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5 mol-% ) och LP = 20.000. Blekmedels området var 984.203 μm 3. Återhämtningstiden och diffusionskoefficienten beräknas från en passning (se 6.1) var τ = 67,3 sek och D = 1,99 um 2 / sek respektive. Klicka här för att se en större version av denna siffra.
Enfärgad Upptäckt av dockning och Fusion händelser med hjälp av en Lipid Label
Manuell räkning av docknings och fusionshändelser är en mödosam uppgift som 100-400 händelser per tillstånd behövs för bra statistik. Eftersom SUV kan röra sig på SBL innan sammansmältning behövs en mjukvara. SpeckleTrackerJ 26 plugin för ImageJ har funktioner som specifikt utvecklats för att spåra SUV för analys av docknings och fusionshändelser. Ofta bara en bråkdel av dockade vesiklar hamnar fixerings;att beräkna dockningshastigheten många fler SUVs behöver spåras än sådana som smälter eller endast en del av filmen måste analyseras tills ett tillräckligt antal händelser detekteras för en tillförlitlig uppskattning. Om huvudfokus är fusionshastigheten och / eller docknings-to-fusion förseningar endast fixerings SUV måste identifieras och spåras.
Efter att säkerställa kvaliteten på SBL, är SUV införs i kanalerna och deras fluorescens kontinuerligt upphetsad enligt TIR använder s-polariserat ljus. För de inledande experimenten, kan infallsvinkeln av TIR excitationsstrålen behöva justeras för att avstämma penetrationsdjupet för den resulterande evanescenta vågen. Ett representativt singel SUV dockning och fusion händelse visas i figur 6A. Vesikeln dockad under ramen märkt D och fusion startade i ram F. Om 1,3 sek senare (ram S) enskilda lipider blev skönjas när de sprids bort från fusions sittae. Den totala fluorescensintensiteten (summan av pixelvärden) i varje låda är avsatt såsom en funktion av tiden i figur 6B, med ramarna vid vilken dockning och fusion inträffade angivna. Det ackumulerade antalet fusionshändelser som en funktion av tiden plottas i Figur 6C, för fem olika filmer med hjälp av t-SNARE rekonstituerade SBLs (t-SBL, LP = 20.000, ca 70 t-snaror per sq. Mikron). Fusionshastigheten från varje film beräknades från en linjär anpassning till lutningen. Detta genomsnitt fusionshastighet ± SEM visas som ett stapeldiagram i panel D. Docknings till fusions fördröjning som visas i figur 6A, B, exempel var 0,07 sek. Fördelningen av förseningar beräknas på samma sätt för totalt 158 händelser visas i figur 6D som en överlevare tomt, det vill säga sannolikheten att en dockad vesikel ännu inte har smält av en viss fördröjning. Ca 1/2 av dessa vesiklar smält inom 100 ms. Med ökande kolesterolnivåer, det distribmepannor av dockning mot fusion förseningar förkortas, trots lipid diffusion saktar 27.
Det är viktigt att köra kontrollexperiment parallellt. På samma täck från vilken data i figur 6B, D förvärvades, var kontrollförhållanden testades i grannkanalerna. I en kanal, gjorde SBL inte omfatta några t-snaror. Av 5 filmer som varar 60 sekunder vardera, endast 7 fusionshändelser registreras jämfört med 158 när t-snaror ingick (Figur 6C). I en annan kanal, var den lösliga cytoplasmiska domänen av v-SNARE VAMP2 (CDV) ingår. CDV binder t-snaror på SBL, förhindrar bindning av fullängds VAMP2 som är på SUV. I fem 60 sec filmer ades inga fusionshändelser detekteras i närvaro av CDV (Figur 6C).

Figur 6. SUV-SBL dockning och fusion händelser. (A) Sekvens av bilder av en v-SUV fusion händelse med t-SBL avbildas av pTIRF mikroskopi efter spridning av LR-PE. Bilderna tas varje 18 ms, endast varannan bildruta visas. D och F markerar uppkomsten av dockning och fusion, respektive. Individuella lipid etiketter blir identifieras som de diffundera bort från fusionsstället och en annan (S). Ramdimensioner: 16,7 um x 16,7 um. (B) Fluorescensintensitet som en funktion av tid för fusions händelsen visas i (A). (C) Sammantagen antalet fusionshändelser som en funktion av tid för v-SUVs märkta med DiD från en annan uppsättning experiment. (D) den normaliserade fusionshastigheten (medelvärde ± SEM, sx ^ m 2 x pM SUV) -1, under antagande av 2 x 10 4 lipider per SUV och (E) den överlevnadssannolikhet för en given tid efter dockning av vesikeln. Kompositionen för alla SBLs var POPC: SAPE: DOPS: Kolesterol: hjärna PI (4,5) P2: PEG2000-DOPE: NBD-PE (54,9: 15: 12: 10: 3: 4,6: 0,5 mol%) och LP = 20.000. v-SUV bestod av POPC: Sape: DOPS: Cholesterol: PEG2000-DOPE: LR-PE (57,4: 15: 12: 10: 4,6: 1 mol%) och LP = 200 för A och B. För CE, v -SUV sammansättning var likartad, förutom gjorde var användes istället för LR-PE. klicka här för att se en större version av denna siffra.
Lipid diffusivitet vesikelstorlek och Fusion Pore Properties
Den enda molekyl känsligheten för analysen tillåter utvinning av flera parametrar utöver docknings och fusions priser och dockning mot fusionsfördröjningsfördelningarna. Singel, fluorescerande lipider blir urskiljbara när de diffundera bort från fusionsstället (Figur 6A). spårning tessa direkt ger lipid diffusivitet 26, 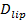 Och en digital mätning av blekningshastigheten för fluoroforer i SBL 27. Som nämnts i inledningen, fluorescensemissionen av etiketterna skiljer sig i SUV kontra i SBL, på grund av tre faktorer som är i allmänhet svårt att skilja: (i) dequenching av etiketter på fusion, på grund av deras utspädning när de överförs till SBL , (ii) en högre exciteringsintensitet i SBL på grund av sönderfallet av det evanescenta fält som alstras av TIR, och (iii) orienteringen av fluoroforen övergångs dipolerna med avseende på polarisationen hos excitationsstrålen. Storleken av faktor (ii) beror på de relativa storlekarna för det evanescenta fältet sönderfallslängd och vesikelstorlek och är olika för varje fusions händelse på grund av den dispersitet i SUV-storlekar. Faktor (iii) beror på fluoroforen och polarisation användas. Snarare än att försöka di sentangle dessa effekter, en kan mäta storleken på den fluorescensförändring direkt genom jämförelse av de totala värdena pixel i en region som omger en SUV före fusion och efter det att alla fluoroforer har deponerats i SBL efter fusion (figur 3A (1) mot ( 2)). Till skillnad från i tidigare arbeten 29,44, bör regionen väljas tillräckligt stor så att inga fluoroforer kommer att ha kvar den via diffusion för tiderna anses efter fusion. Att välja en 30 x 30 pixel region (8 pm x 8 pm) för analys upp till 1,6 sek efter fusion är vanligtvis tillräckligt 27. Ignorera blekning för tillfället, förhållandet mellan
Och en digital mätning av blekningshastigheten för fluoroforer i SBL 27. Som nämnts i inledningen, fluorescensemissionen av etiketterna skiljer sig i SUV kontra i SBL, på grund av tre faktorer som är i allmänhet svårt att skilja: (i) dequenching av etiketter på fusion, på grund av deras utspädning när de överförs till SBL , (ii) en högre exciteringsintensitet i SBL på grund av sönderfallet av det evanescenta fält som alstras av TIR, och (iii) orienteringen av fluoroforen övergångs dipolerna med avseende på polarisationen hos excitationsstrålen. Storleken av faktor (ii) beror på de relativa storlekarna för det evanescenta fältet sönderfallslängd och vesikelstorlek och är olika för varje fusions händelse på grund av den dispersitet i SUV-storlekar. Faktor (iii) beror på fluoroforen och polarisation användas. Snarare än att försöka di sentangle dessa effekter, en kan mäta storleken på den fluorescensförändring direkt genom jämförelse av de totala värdena pixel i en region som omger en SUV före fusion och efter det att alla fluoroforer har deponerats i SBL efter fusion (figur 3A (1) mot ( 2)). Till skillnad från i tidigare arbeten 29,44, bör regionen väljas tillräckligt stor så att inga fluoroforer kommer att ha kvar den via diffusion för tiderna anses efter fusion. Att välja en 30 x 30 pixel region (8 pm x 8 pm) för analys upp till 1,6 sek efter fusion är vanligtvis tillräckligt 27. Ignorera blekning för tillfället, förhållandet mellan  före och efter fusion ger fluorescensintensiteten reduktionsfaktor,
före och efter fusion ger fluorescensintensiteten reduktionsfaktor, 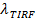 . Den vesikelstorlek kan uppskattas givet fluorescensintensiteten för en enda etikett i SBL (iles / ftp_upload / 54349 / 54349eq6.jpg "/>), den faktor med vilken dess intensitet skulle minska om det var i en SUV ( ), Den kända densiteten för etiketter i SUV, och området som upptas av en lipid. I praktiken är blekning vanligtvis snabb nog att när alla markörer sätts in SBL, några har redan blekt. Av denna anledning, för en korrekt uppskattning, fluorescensintensiteten reduktionsfaktor, Och tids lipid release,
. Den vesikelstorlek kan uppskattas givet fluorescensintensiteten för en enda etikett i SBL (iles / ftp_upload / 54349 / 54349eq6.jpg "/>), den faktor med vilken dess intensitet skulle minska om det var i en SUV ( ), Den kända densiteten för etiketter i SUV, och området som upptas av en lipid. I praktiken är blekning vanligtvis snabb nog att när alla markörer sätts in SBL, några har redan blekt. Av denna anledning, för en korrekt uppskattning, fluorescensintensiteten reduktionsfaktor, Och tids lipid release,  Är bäst utvinns ur en passning till tidsförloppet av den totala pixelvärden som beaktar blekning 27. Blekningstakten etiketter i SBL oberoende beräknas från enstaka lipid färgämne spårning, eller genom att montera en exponentiell till den totala intensiteten profil,
Är bäst utvinns ur en passning till tidsförloppet av den totala pixelvärden som beaktar blekning 27. Blekningstakten etiketter i SBL oberoende beräknas från enstaka lipid färgämne spårning, eller genom att montera en exponentiell till den totala intensiteten profil,  Vid långa tider (t.ex. regim (3) i Figur 3). Att veta storleken på en fixerings vesikel och lipid diffusivitet tillåter jämförelse av den faktiska lipid frigöra tid, , Till diffusionstiden av en lipid runt vesikeln
Vid långa tider (t.ex. regim (3) i Figur 3). Att veta storleken på en fixerings vesikel och lipid diffusivitet tillåter jämförelse av den faktiska lipid frigöra tid, , Till diffusionstiden av en lipid runt vesikeln  , Där A ves är vesikeln området och är lipiden diffusivitet. Om de två tidsskalor är jämförbara, presenterar por litet motstånd mot lipid release, och släpp är diffusion begränsad. Om däremot
, Där A ves är vesikeln området och är lipiden diffusivitet. Om de två tidsskalor är jämförbara, presenterar por litet motstånd mot lipid release, och släpp är diffusion begränsad. Om däremot  , Då por fördröjer lipid flödet avsevärt. Ett kvantitativt mått på retardationen är "pore öppenheten", P 0, lika med förhållandet
, Då por fördröjer lipid flödet avsevärt. Ett kvantitativt mått på retardationen är "pore öppenheten", P 0, lika med förhållandet  gånger en faktor of för enighet i samband med porgeometrin 27. För en två-stat, öppen / stängd pore, P 0 är den bråkdel av tiden i det öppna tillståndet. För en flimrande por vars storlek kontinuerligt varierar, P 0 representerar den genomsnittliga tids radie i förhållande till den helt öppna radie.
gånger en faktor of för enighet i samband med porgeometrin 27. För en två-stat, öppen / stängd pore, P 0 är den bråkdel av tiden i det öppna tillståndet. För en flimrande por vars storlek kontinuerligt varierar, P 0 representerar den genomsnittliga tids radie i förhållande till den helt öppna radie.
Den signal-brusförhållande måste vara tillräckligt bra för att spåra enskilda lipid fluoroforer. Detta är lättare att uppnå i enfärgade experiment med hjälp av en ljus markör som LR-PE.
Samtidig detektion av lipid och Löslig Cargo Release
Ett representativt händelsen visas i figur 7. Vid de höga koncentrationerna som användes för inkapsling, är SRB fluorescens initialt själv släcktes. Därför är de flesta dockad v-SUV är endast detekteras genom deras DiD signal 27. En kraftig ökningi DiD fluorescens markerar lipidblandning, som i enfärgade lipid-etikett experiment. Uppkomsten av lipid frisättning sammanföll med uppkomsten av en ökning av SRB fluorescens. Denna ökning var mest sannolikt på grund av SRB dequenching som de SRB molekylerna undgått SUV och den återstående SRB utspäddes till under självdämpande koncentrationer. Intressant nådde SRB fluorescens en stabil platå. Om fusions por var öppen, skulle man förvänta sig att SRB-signalen att öka till maximalt på grund av dequenching, följt av en minskning till bakgrund som den återstående SRB lämnade SUV, eller om SUV kollapsade i SBL. Den stabila platå troligen indikerar por återförslutas efter att ha släppt alla lipid etiketter och en bråkdel av den lösliga last.
I vissa fall var lipid frigör signalen inte åtföljs av några påvisbara innehåll släpper signaler. Detta kan vara antingen för att porerna var för små för att tillåta passage av SRB, eller någonhur SUV redan hade förlorat sin lösliga last. För att skilja mellan dessa två möjligheter, kan blekning av dockade SUV analyseras. Blekning av lipid etikett DiD minskar dess fluorescenssignal inom några sekunder, medan blekning av initialt själv kylda SRB resulterar i en ökning av SRB-signalen som en bråkdel av SRB är foto omvandlas till andra produkter och självsläckning är lättad. Medan de flesta dockade SUV (48 av 58 analyserade händelser) visade den förväntade ökningen av SRB fluorescens under längre perioder av belysning, hade en liten fraktion (10/58) inte visar några SRB signaler alls. Sålunda kan en liten del av de SUV förlorar sin lösliga innehåll under perioden mellan deras framställning och användning i experimenten.
I mer än 80% av händelser (74 av 91) som båda SRB och gjorde signaler kan följas, SRB fluorescens ökat och låg kvar på en stabil platå 27. I ~ 20% av tessa fall återstående SRB fluorescens plötsligt släpps upp till några sekunder senare. Detta kan antingen bero på att en återupptagandet av porer tillåter snabb, full frisättning inom 1-2 ramar (18-36 ms) eller en bristning av SUV på grund av ackumulerade foto skada 16. Oberoende av dess ursprung, detta andra, snabb frisättning av SRB utesluter möjligheten att rest SRB-signalen efter den initiala frisättningen kan bero på SRB förblir infångad i utrymmet mellan SBL och glassubstratet efter full release. En sådan mekanism föreslogs i en tidigare studie där SBL stöddes direkt på glas, med lite utrymme mellan de två 16. Däremot bör de PEGylerade lipider som användes i denna analys ge 4-5 nm av utrymmet mellan SBL och glaset 25, vilket gör att SRB att diffundera bort från fusionsstället.

Figur 7. Simultan Exportorienterade företag lipid och innehåll release. fluorescenssignaler från lipid (blå) och innehåll (röd) markör för en SUV-SBL fusion händelse (för detaljer se text). Ögonblicksbilder från varje kanal visas längst ner (4,0 fim bred). Tecknad film (mitten) visar de olika stegen. (1) SUV bryggor, gjorde fluorescens är låg. (2) Fusion tillåter överföring av DiD i SBL och gjorde fluorescensökningar. SRB-signal ökar också som självsläckning avlastas genom utsläpp av SRB genom poren. (3) SRB signalen förblir stabila medan gjorde molekyler diffundera inom SBL indikerar en möjlig återförslutning av porerna. (4) SRB signaler minskar mycket snabbt, på grund av snabb fullständig fusion eller liposomer spricker. Sammansättningen av v-SUVs var POPC: SAPE: DOPS: PEG2000-DOPE: DiD (67: 15: 12: 5: 1 mol%), LP 200, medan t-SBLs var sammansatta av POPC: SAPE: DOPS: hjärna PI (4,5) P 2: PEG2000-DOPE: NBD-PE (64,5: 15: 12: 3: 5: 0,5 mol-%) och LP = 20.000. = "_ Blank"> Klicka här för att se en större version av denna siffra.
FRAP_instructions.pdf -. Anvisningar för att upprätta en FRAP sekvens klicka god här för att ladda ner filen.
ReadMe_FRAP.txt -. Textfil som beskriver förfarandet för att analysera FRAP återhämtning data med hjälp av MatLab kod som tillhandahålls Klicka här för att ladda ner filen.
EK_FRAPbatch.m - MATLAB-filen till sats analysera fluorescens återhämtning efter fotoblekning. Denna fil anropar MATLAB subrutiner EK_getFRAPvals.m och EK_SoumpassisFit.m."_blank"> Klicka här för att ladda ner filen.
EK_getFRAPvals.m -. MATLAB fil, subrutin anropas av EK_FRAPbatch.m Klicka här för att ladda ner filen.
EK_SoumpassisFit.m -. MATLAB fil, subrutin anropas av EK_FRAPbatch.m Klicka här för att ladda ner filen.
JN150923c1_FRAP_list.txt -. Ett exempel på en lista över filnamn att vara satsvis analyseras av huvud MATLAB script EK_FRAPbatch.m Klicka här för att ladda ner filen.
DeFöljande filer TIF stackar och motsvarande textfiler för fem exempel på FRAP sekvenser som kan analyseras genom att köra EK_FRAPbatch.m och välja JN150923c1_FRAP_list.txt filen när du uppmanas att välja en fillista.
JN150923c1_F1_MMStack_Pos0.ome.tif -. FRAP bildstapel Klicka här för att ladda ner filen.
JN150923c1_F1_MMStack_Pos0_metadata.txt -. Motsvarande textfil med metadata klicka god här för att ladda ner filen.
JN150923c1_F2_MMStack_Pos0.ome.tif -. FRAP bildstapel Pllätta klicka här för att ladda ner filen.
JN150923c1_F2_MMStack_Pos0_metadata.txt -. Motsvarande textfil med metadata klicka god här för att ladda ner filen.
JN150923c1_F3_MMStack_Pos0.ome.tif -. FRAP bildstapel Klicka här för att ladda ner filen.
JN150923c1_F3_MMStack_Pos0_metadata.txt -. Motsvarande textfil med metadata klicka god här för att ladda ner filen.
JN150923c1_F4_MMStack_Pos0.ome.tif - FRAP bildstapel. Klicka här för att ladda ner filen.
JN150923c1_F4_MMStack_Pos0_metadata.txt -. Motsvarande textfil med metadata klicka god här för att ladda ner filen.
JN150923c1_F5_MMStack_Pos0.ome.tif -. FRAP bildstapel Klicka här för att ladda ner filen.
JN150923c1_F5_MMStack_Pos0_metadata.txt -. Motsvarande textfil med metadata klicka god här för att ladda ner filen.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Ett framgångsrikt genomförande av SUV-SBL fusion analys som beskrivs här beror kritiskt på flera viktiga steg, såsom funktionell beredning av proteiner i liposomer, få bra kvalitet SBLs, och välja rätt avbildningsparametrar för att detektera enstaka molekyler. Även om det kan ta lite tid och ansträngning för att lyckas, när analysen genomförs framgångsrikt, ger det en mängd information om fusionsprocess inte kan erhållas från någon annan in vitro fusionsanalys diskuteras i Introduktion. De kurser där SUV docka på och säkring med SBL, fördelningen av dockning mot fusion gånger, lipid diffusivitet, storleken på varje fusing liposomer, och om lipid övergång till en given fusion händelse är diffusion- eller por styrda burk vara beslutsam. Om release är mycket långsammare än väntat från diffusion, en modell som förutsätter por flimmer ligger bakom fördröjningen av lipid släpp avkastning, P 0, bråkdel av tiden porerna iär öppen under fusion 27.
Tidigare arbete sondering enda SUV-SUV fusion 18-20,28 eller fusion av liposomer och viruspartiklar till SBLs 15-17,21,22,33,44 åberopas FRET och självsläckning mellan färgämnesmolekyler. Däremot undviker vi i stort sett färg dequenching att sondera kinetik lipid etikett överföring till SBL, av två skäl. För det första är FRET effektiviteten en höggradigt icke-linjär reporter av lipid release. Små förändringar i inter-dye avståndet d resulterar i stora förändringar i FRET effektivitet, men bara för ett begränsat antal av D / R 0 värden, där R 0 är Forster avstånd. Det vill säga, en fördröjningstid mellan faktisk färgämne utspädning och en observerad ökning av dequenching signalen kan förekomma, på grund av den tid som krävs för att falla till ett värde nära R 0. Likaså finns det små förändringar i dequenching signalen när färgämnestätheten minskar tillräckligt på lipid frisättning så att  1,4, även om en betydande mängd av färgämne finns fortfarande kvar i vesikeln. Dessa egenskaper gör dequenching ett bra verktyg för att få en spik i signalen för att detektera fusionshändelser, men försvåra kvantitativ kinetisk analys av lipid frisättning med användning av hög tidsupplösning. Däremot är förändringarna i intensitet på grund av dipol omorientering och försvinnande fälteffekter som en fluorofor överföringar från en SUV i SBL linjärt relaterad till den del av färgen fortfarande finns kvar i SUV. För det andra kan höga koncentrationer av lipid etiketter som krävs för självsläckning påverka fusionsprocessen.
1,4, även om en betydande mängd av färgämne finns fortfarande kvar i vesikeln. Dessa egenskaper gör dequenching ett bra verktyg för att få en spik i signalen för att detektera fusionshändelser, men försvåra kvantitativ kinetisk analys av lipid frisättning med användning av hög tidsupplösning. Däremot är förändringarna i intensitet på grund av dipol omorientering och försvinnande fälteffekter som en fluorofor överföringar från en SUV i SBL linjärt relaterad till den del av färgen fortfarande finns kvar i SUV. För det andra kan höga koncentrationer av lipid etiketter som krävs för självsläckning påverka fusionsprocessen.
För polariserat TIRF excitation, ett hem-byggda installation såsom i fig 4 är kostnadseffektiv och ger utmärkt kontroll över polarisationsegenskaper. Men det är en hembyggda installationen inte nödvändigt, eftersom åtminstone en del kommersiella TIRF mikroskop använder polariserat excitation och off-the-shelftvåfärgad emissions delning moduler finns tillgängliga. En kommersiell mikroskop som använder en polarisationsbevarande (PM) fiber för att koppla den polariserat excitationsljus som kommer ut från lasern in i mikroskopet användes med framgång tidigare 25-27. På ett sådant instrument, kan polarisationen roteras helt enkelt genom att vrida fiberingång PM till mikroskopets TIRF enhet. Däremot kan vissa andra kommersiella inställningar anställa en polariserande stråldelare i TIRF enheten göra justeringar polarisering mer svårt eller omöjligt. Tyvärr är polarisering egenskaper ofta inte anges för kommersiella system. Således är det viktigt för användare som inte vill bygga en hembyggda system för att diskutera polariseringsegenskaper och prova en demo enhet innan du investerar i en viss kommersiell alternativ.
Samtidig övervakning av lipid och löslig last versionen innehåller ytterligare information om fusionsprocessen. Många porer verkar att återförsluta efter releAsing bara en bråkdel av den lösliga etikett inkapslas i ett SUV 27. Vanligtvis alla lipid etiketter frigörs vid den tidpunkt då por återförsluts, så por återförslutning inte misstänktes tidigare från övervaknings lipid frigör ensam. I själva verket, övervakning lipid frigör ensam, Kiessling et al. 29 hade föreslagit överföring av märkta lipider i SBL skett via extremt snabb kollaps av hela SUV membranet in i SBL.
Återförslutning av fusions porer efter partiell frigöring av innehållet har tidigare observerats i en liposom-bunden tvåskiktsmembran patch fusion analys. Rawle et al. 54 förberedda fristående dubbelskiktsplåster som bundna på ett täck av ~ 8 nm långa DNA-distanser. De introducerade därefter liposomer vars fusion till biskiktet patch drevs av hybridisering av komplementära enkelsträngade DNA-lipid-konjugat förankrade till liposomen och patch membran. 12% av alla händelser var övergående fusioner, vilket resulterar i en delial frisättning av liposomen innehåll. Liposom spricker händelser, sett i vissa tidigare SUV-SBL fusionsanalyser 44, var mycket sällsynt. Författarna föreslog ~ 8 nm utrymme nedan biskiktet lapp, förhindra biskikt-substratinteraktioner, kan ha tillåtit icke-läckande överföring av de liposom-innehållet till utrymmet nedanför dubbelskiktet. Dessutom var diffusion av de frigjorda lösliga etiketterna inte signifikant hindras i utrymmet mellan den plana biskikt och glassubstratet. I den analys som beskrivs här, är den plana biskikt liknande sätt hålls ~ 5 nm bort från täckglaset genom närvaron av poly (etylenglykol) -lipid konjugat.
Den ytterligare information som tillhandahålls av tvåfärgad övervakning av lösliga och lipid last kommer med några kompromisser. För det första är framställningen av SUV med två etiketter något mer engagerade än inklusive lipid etiketter ensam. För det andra, det finns en viss oundviklig förlust av signal-till-brus vid övervakning signaler från lipid etiketter vid than samma förvärvs takt som enfärgade lipid släpp experiment (15-20 ms per ram). En del av denna förlust beror på det faktum som gjorde, som används som en lipid etikett tillsammans med den lösliga SRB markör, är inte lika ljus som LR-PE som används i enfärgade experiment. Ytterligare förluster härrör från högre bakgrunden på grund av samtidig två-laser excitation och ytterligare komponenter i den optiska banan (Figur 4). Hittills förhindrat lägre signal-till-brus oss från tillförlitlig spårning av enstaka DiD etiketter i två färger experiment. Detta i sin tur förhindrade en grundlig analys för att extrahera all information som kan vara med LR-PE mätningar enfärgade. Denna utmaning kan övervinnas i framtiden genom att optimera förvärvsvillkor och testa andra par av kompatibla etiketter med förbättrade egenskaper.
Samtidig övervakning av lösliga och lipid last Meddelandet kan även upptäcka hemifusion, ett tillstånd i vilket de proximala broschyrer har smält, men den distalabroschyrer har inte. I själva verket kan sådana två-färg experiment ger den enda slutgiltig bedömning av förekomsten av hemifusion i andra in vitro fusions analyser 18,20. Däremot kan hemifusion tillförlitligt detekteras i enfärgade SUV-SBL experiment övervakning lipid frisättning enbart 22,44. I sådana experiment, fortfarande ca 1/2 den initiala lipid etikett intensiteten hos en SUV på fusions plats. Den kvarvarande intensitet kan försvinna när de distala flygblad smälta en tid senare, om platsen ännu inte har blekts.
Användning av PEGylerade lipider att införa en mjuk kudde mellan SBL och glas stöd har varit avgörande för att reproducera SNAP25 kravet på exocytos. Protokollresulterar i en PEG borsta som täcker den sida av SBL som vetter mot flödeskanalen också. Detta har vissa fördelar, såsom att minska icke-specifika interaktioner mellan SUV och SBL, som förklaras i Introduktion. Om proteiner som interagerar med fosfolipiderna iSBL införs i analysen kommer emellertid PEG borsten presentera en sterisk barriär mot sådana interaktioner. Således, för vissa tillämpningar, skulle det vara önskvärt att inkludera PEG skiktet asymmetriskt, bara mellan SBL och glastäckglas. Den sida av SBL som vetter mot flödeskanalen skulle då kunna interagera med fosfolipid-bindande proteiner, såsom synaptotagmin. Detta kan åstadkommas genom användning av bi-funktionella PEG-kedjor som kan vara kovalent kopplade till täckglas vid en ände och som uppbär en lipid ankare vid den andra. Langmuir-Blodgett-metoder kan sedan användas för att deponera en lipid monolager på PEG skiktet 55. Liposomer införs ovanför denna hydrofoba ytan säkring med det, som bildar ett dubbelskikt 56. Förhoppningen är att detta tillvägagångssätt kommer att kombineras med mikrofluidik i framtiden.
Det förväntas att SUV-SBL-analysen presenteras här kommer att vara till hjälp för att utforska rollen av ytterligare proteiner som interagerar med snaror,såsom Munc18 / Sec1 familjen, kalciumsensor synaptotagmin-1, och complexin.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna förklarar att de inte har några konkurrerande ekonomiska intressen.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| Milli-Q (MQ) water | Millipore | ||
| KOH | J.T. Baker | 3040-05 | |
| Ethanol 190 Proof | Decon | ||
| Isopropanol | Fisher Chemical | A416P4 | |
| HEPES | AmericanBio | AB00892 | |
| Sodium Cholride (KCl) | AmericanBio | AB01915 | |
| Dithiothreitol | AmericanBio | AB00490 | |
| N-[2-hydroxyethyl] piperazine-N'-[2-ethanesulfonic acid] (HEPES) | AmericanBio | AB00892 | |
| EGTA | Acros Organics | 409911000 | |
| Buffers | |||
| HEPES-KOH buffer (pH 7.4) | 25 mM HEPES-KOH, 140 mM KCl, 100 μM EGTA, 1 mM DTT | ||
| Solvents | |||
| Chloroform | J.T. Baker | 9180-01 | in glass bottle, CAUTION, wear PPE |
| Methanol | J.T. Baker | 9070-03 | in glass bottle, CAUTION, wear PPE |
| Liposome preparation | |||
| Gastight Hamilton syringe | Hamilton | var. sizes | only use glass sringe with solents (Chlorophorm/ Methanon, 2:1, v/v) |
| Glass tubes Pyrex Vista 11 ml, 16x100 mm screw cap culture tube | Pyrex | 70825-16 | clean thoroughly, rinse with chloroform |
| 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine, 16:0-18:1 PC (POPC) | Avanti Polar Lipids | 850457 | Lipids come dissolved in CHCl3 or as lyphilized powder in sealed vials. Aliquot upon opening. Store extra as dried lipid films under inert atmosphere at -20 °C. Keep stocks in CHCl3/MeOH (2:1, v/v) at -20 °C. let come to RT before opening |
| 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (sodium salt), 18:1 PS (DOPS) | Avanti Polar Lipids | 840035 | Same as above. |
| 1-stearoyl-2-arachidonoyl-sn-glycero-3-phosphoethanolamine, 18:0-20:4 PE (SAPE) | Avanti Polar Lipids | 850804 | Same as above. |
| L-α-phosphatidylinositol-4,5-bisphosphate (Brain, Porcine) (ammonium salt), Brain PI(4,5)P2 | Avanti Polar Lipids | 840046 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(7-nitro-2-1,3-benzoxadiazol-4-yl) (ammonium salt), 18:1 NBD PE | Avanti Polar Lipids | 810145 | Same as above. |
| 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene glycol)-2000] (ammonium salt), 18:1 PEG2000 PE | Avanti Polar Lipids | 880130 | Same as above. |
| cholesterol (ovine wool, >98%) | Avanti Polar Lipids | 700000 | Same as above. |
| DiD' oil; DiIC18(5) oil (1,1'-Dioctadecyl-3,3,3',3'-Tetramethylindodicarbocyanine Perchlorate) | Molecular Probes | D-307 | |
| Rotavapor R-210 | Buchi | R-210 | heat bath above Tm of lipids used |
| OG n-Octyl-β-D-Glucopyranoside | Affymetrix | 0311 | store at -20 °C, let come to RT before opening |
| Shaker - Eppendorf Thermomixer R | Eppendorf | ||
| Slide-A-Lyze Dialysis Cassettes, 20k MWCO, 3 ml | life technologies | 66003 | |
| Bio-Beads SM-2 Adsorbents | Bio-Rad | 1523920 | |
| OptiPrep Density Gradient Medium | Sigma-Aldrich | D1556 | |
| Ultracentrifugation tube, Thinwall, Ultra-Clear, 13.2 ml, 14 x 89 mm | Beckman Coulter | 41121703 | |
| Beckman SW41 Ti rotor | |||
| SuflorhodamineB | Molecular Probes | S-1307 | |
| Econo-Column Chromatography Columns, 2.5 × 10 cm | Bio-Rad | 7372512 | |
| Sepharose CL-4B | GE Healthcare | 17-0150-01 | |
| SYPRO Orange Protein Gel Stain | Molecular Probes | S-6650 | 5,000x Concentrate in DMSO |
| PDMS block | |||
| Sylgard 184 Silicone elastomer kit, PDMS | Dow Corning | 3097358-1004 | |
| Pyrex glass petri dish, 150 x 20 mm, complete with cover | Corning | 3160-152 | |
| Hole puncher - Reusable Biopsy Punch, 0.75 mm | World Precision Instruments | 504529 | |
| Manual Hole Punching Machine | SYNEO | MHPM-UNV | |
| Drill .035 x .026 x 1.5 304 SS TiN coated round punch | SYNEO | CR0350265N20R4 | drill diameter: 0.9 mm, punch bore size: 0.74 mm |
| Tygon Microbore tubing, 0.25 mm ID, 0.76 mm OD | Cole-Parmer | 06419-00 | 0.010" ID, 0.030" OD |
| Silicone Tubing (0.51 mm ID, 2.1 mm OD | Cole-Parmer | 95802-00 | 0.020" ID, 0.083" OD |
| Cover glass - cleanroom cleaned | |||
| Schott Nexterion cover slip glass D | Schott | 1472305 | |
| plasma cleaner | Harrick | PDC-32G | |
| pTIRF setup and accessories | |||
| IX81 microscope body | Olympus | IX81 | |
| EM CCD camera | Andor | ixon-ultra-897 | |
| Thermo Plate, heated microscope stage | Tokai Hit | MATS-U52RA26 | |
| 1 ml hamilton glass syringes (4x) | Hamilton | 81365 | |
| syringe pump | kd Scientific | KDS-230 | |
References
- Sudhof, T. C., Rothman, J. E. Membrane fusion: grappling with SNARE and SM proteins. Science. 323, 474-477 (2009).
- Wickner, W., Schekman, R. Membrane fusion. Nat Struct Mol Biol. 15, 658-664 (2008).
- Harrison, S. C. Viral membrane fusion. Nat Struct Mol Biol. 15, 690-698 (2008).
- Jahn, R., Scheller, R. H. SNAREs--engines for membrane fusion. Nat Rev Mol Cell Biol. 7, 631-643 (2006).
- Lindau, M., Alvarez de Toledo, G. The fusion pore. Biochim Biophys Acta. 1641, 167-173 (2003).
- Staal, R. G., Mosharov, E. V., Sulzer, D. Dopamine neurons release transmitter via a flickering fusion pore. Nat Neurosci. 7, 341-346 (2004).
- Wu, Z., et al. Nanodisc-cell fusion: Control of fusion pore nucleation and lifetimes by SNARE protein transmembrane domains. Sci. Rep. 6, 27287 (2016).
- Alabi, A. A., Tsien, R. W. Perspectives on kiss-and-run: role in exocytosis, endocytosis, and neurotransmission. Ann Rev Physiol. 75, 393-422 (2013).
- Rossetto, O., Pirazzini, M., Montecucco, C. Botulinum neurotoxins: genetic, structural and mechanistic insights. Nature Rev Microbiol. 12, 535-549 (2014).
- Weber, T., et al. SNAREpins: minimal machinery for membrane fusion. Cell. 92, 759-772 (1998).
- Nickel, W., et al. Content mixing and membrane integrity during membrane fusion driven by pairing of isolated v-SNAREs and t-SNAREs. Proc Natl Acad Sci U S A. 96, 12571-12576 (1999).
- McNew, J. A., et al. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature. 407, 153-159 (2000).
- Melia, T. J., You, D. Q., Tareste, D. C., Rothman, J. E. Lipidic antagonists to SNARE-mediated fusion. J Biol Chem. 281, 29597-29605 (2006).
- Hernandez, J. M., et al. Membrane fusion intermediates via directional and full assembly of the SNARE complex. Science. 336, 1581-1584 (2012).
- Fix, M., et al. Imaging single membrane fusion events mediated by SNARE proteins. Proc Natl Acad Sci U S A. 101, 7311-7316 (2004).
- Bowen, M. E., Weninger, K., Brunger, A. T., Chu, S. Single molecule observation of liposome-bilayer fusion thermally induced by soluble N-ethyl maleimide sensitive-factor attachment protein receptors (SNAREs). Biophys J. 87, 3569-3584 (2004).
- Liu, T., Tucker, W. C., Bhalla, A., Chapman, E. R., Weisshaar, J. C. SNARE-driven, 25-millisecond vesicle fusion in vitro. Biophys J. 89, 2458-2472 (2005).
- Yoon, T. Y., Okumus, B., Zhang, F., Shin, Y. K., Ha, T. Multiple intermediates in SNARE-induced membrane fusion. Proc Natl Acad Sci U S A. 103, 19731-19736 (2006).
- Diao, J., et al. A single-vesicle content mixing assay for SNARE-mediated membrane fusion. Nat Commun. 1, 1-6 (2010).
- Kyoung, M., et al. In vitro system capable of differentiating fast Ca2+-triggered content mixing from lipid exchange for mechanistic studies of neurotransmitter release. Proc Natl Acad Sci U S A. 108, E304-E313 (2011).
- Domanska, M. K., Kiessling, V., Stein, A., Fasshauer, D., Tamm, L. K. Single vesicle millisecond fusion kinetics reveals number of SNARE complexes optimal for fast SNARE-mediated membrane fusion. J Biol Chem. 284, 32158-32166 (2009).
- Kreutzberger, A. J., Kiessling, V., Tamm, L. K. High Cholesterol Obviates a Prolonged Hemifusion Intermediate in Fast SNARE-Mediated Membrane Fusion. Biophys J. 109, 319-329 (2015).
- Schwenen, L. L., et al. Resolving single membrane fusion events on planar pore-spanning membranes. Sci Rep. 5, 12006 (2015).
- Karatekin, E., et al. A fast, single-vesicle fusion assay mimics physiological SNARE requirements. Proc Natl Acad Sci U S A. 107, 3517-3521 (2010).
- Karatekin, E., Rothman, J. E. Fusion of single proteoliposomes with planar, cushioned bilayers in microfluidic flow cells. Nat Protoc. 7, 903-920 (2012).
- Smith, M. B., et al. Interactive, computer-assisted tracking of speckle trajectories in fluorescence microscopy: application to actin polymerization and membrane fusion. Biophys J. 101, 1794-1804 (2011).
- Stratton, B. S., et al. Cholesterol Increases the Openness of SNARE-mediated Flickering Fusion Pores. Biophysical journal. 110, (2016).
- Diao, J., et al. Synaptic proteins promote calcium-triggered fast transition from point contact to full fusion. Elife. 1, e00109 (2012).
- Kiessling, V., Domanska, M. K., Tamm, L. K. Single SNARE-mediated vesicle fusion observed in vitro by polarized TIRFM. Biophys J. 99, 4047-4055 (2010).
- Blasi, J., et al. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature. 365, 160-163 (1993).
- Washbourne, P., et al. Genetic ablation of the t-SNARE SNAP-25 distinguishes mechanisms of neuroexocytosis. Nat Neurosci. 5, 19-26 (2002).
- Diaz, A. J., Albertorio, F., Daniel, S., Cremer, P. S. Double cushions preserve transmembrane protein mobility in supported bilayer systems. Langmuir. 24, 6820-6826 (2008).
- Floyd, D. L., Ragains, J. R., Skehel, J. J., Harrison, S. C., van Oijen, A. M. Single-particle kinetics of influenza virus membrane fusion. Proc Natl Acad Sci U S A. 105, 15382-15387 (2008).
- Albertorio, F., et al. Fluid and air-stable lipopolymer membranes for biosensor applications. Langmuir. 21, 7476-7482 (2005).
- Daniel, S., Albertorio, F., Cremer, P. S. Making lipid membranes rough, tough, and ready to hit the road. Mrs Bulletin. 31, 536-540 (2006).
- Gao, Y., et al. Single reconstituted neuronal SNARE complexes zipper in three distinct stages. Science. 337, 1340-1343 (2012).
- Kenworthy, A. K., Hristova, K., Needham, D., Mcintosh, T. J. Range and Magnitude of the Steric Pressure between Bilayers Containing Phospholipids with Covalently Attached Poly(Ethylene Glycol). Biophys J. 68, 1921-1936 (1995).
- Knoll, W., et al. Solid supported lipid membranes: New concepts for the biomimetic functionalization of solid surfaces. Biointerphases. 3, Fa125-Fa135 (2008).
- Quinn, P., Griffiths, G., Warren, G. Density of newly synthesized plasma membrane proteins in intracellular membranes II. Biochemical studies. J Cell Biol. 98, 2142-2147 (1984).
- Sund, S. E., Swanson, J. A., Axelrod, D. Cell membrane orientation visualized by polarized total internal reflection fluorescence. Biophys J. 77, 2266-2283 (1999).
- Johnson, D. S., Toledo-Crow, R., Mattheyses, A. L., Simon, S. M. Polarization-controlled TIRFM with focal drift and spatial field intensity correction. Biophys J. 106, 1008-1019 (2014).
- Anantharam, A., Onoa, B., Edwards, R. H., Holz, R. W., Axelrod, D. Localized topological changes of the plasma membrane upon exocytosis visualized by polarized TIRFM. J Cell Biol. 188, 415-428 (2010).
- Axelrod, D. Carbocyanine dye orientation in red cell membrane studied by microscopic fluorescence polarization. Biophys J. 26, 557-573 (1979).
- Wang, T., Smith, E. A., Chapman, E. R., Weisshaar, J. C. Lipid mixing and content release in single-vesicle, SNARE-driven fusion assay with 1-5 msec resolution. Biophys J. 96, 4122-4131 (2009).
- Chernomordik, L. V., Frolov, V. A., Leikina, E., Bronk, P., Zimmerberg, J. The pathway of membrane fusion catalyzed by influenza hemagglutinin: restriction of lipids, hemifusion, and lipidic fusion pore formation. J Cell Biol. 140, 1369-1382 (1998).
- Takamori, S., et al. Molecular anatomy of a trafficking organelle. Cell. 127, 831-846 (2006).
- Wilhelm, B. G., et al. Composition of isolated synaptic boutons reveals the amounts of vesicle trafficking proteins. Science. 344, 1023-1028 (2014).
- Scott, B. L., et al. Liposome fusion assay to monitor intracellular membrane fusion machines. Methods Enzymol. 372, 274-300 (2003).
- Linkert, M., et al. Metadata matters: access to image data in the real world. J Cell Biol. 189, 777-782 (2010).
- Soumpasis, D. M. Theoretical analysis of fluorescence photobleaching recovery experiments. Biophys J. 41, 95-97 (1983).
- Ohki, S. A mechanism of divalent ion-induced phosphatidylserine membrane fusion. Biochim Biophys Acta. 689, 1-11 (1982).
- Berquand, A., et al. Two-step formation of streptavidin-supported lipid bilayers by PEG-triggered vesicle fusion. Fluorescence and atomic force microscopy characterization. Langmuir. 19, 1700-1707 (2003).
- Tamm, L. K., McConnell, H. M. Supported phospholipid bilayers. Biophys J. 47, 105-113 (1985).
- Rawle, R. J., van Lengerich, B., Chung, M., Bendix, P. M., Boxer, S. G. Vesicle fusion observed by content transfer across a tethered lipid bilayer. Biophys J. 101, L37-L39 (2011).
- Wagner, M. L., Tamm, L. K. Reconstituted syntaxin1a/SNAP25 interacts with negatively charged lipids as measured by lateral diffusion in planar supported bilayers. Biophys J. 81, 266-275 (2001).
- Kalb, E., Frey, S., Tamm, L. K. Formation of Supported Planar Bilayers by Fusion of Vesicles to Supported Phospholipid Monolayers. Biochimica Et Biophysica Acta. 1103, 307-316 (1992).
