

Introduction
أجسام مضادة قادرة على لطي وتعمل في بيئة الخلايا هي أدوات واعدة لكل من البحوث والتطبيقات العلاجية. لديهم القدرة على تعديل نشاط البروتين ملزمة لبروتين هدف داخل الخلايا لمنع تفاعلات البروتين البروتين، وتعطيل التفاعلات حمض البروتين النووي، أو منع الوصول إلى الركيزة الانزيمات 1-5.
على الرغم من أن الأجسام المضادة لديهم الكثير من الإمكانيات لتطبيقات الخلايا، هندسة منهم للطي السليم والذوبان في البيئة داخل الخلايا مع الحفاظ على القدرة على ربط لمستضد الهدف هو الصعبة. البيئة هيولي خفض يمنع تشكيل السندات ثاني كبريتيد المطلوبة عادة للطي استقرارا من الأجسام المضادة كامل طول وشظايا الأجسام المضادة، بما في ذلك سلسلة واحدة جزء متغير (scFv) الأجسام المضادة 6،7. وقد تم استخدام عدد من نهج التطور الموجه لهندسة الأجسام المضادة مع حالانتماءات IGH للهدف مستضدات 8-10. هذه الأساليب شيوعا في استخدام عرض فج، الخميرة عرض السطح، أو عرض سطح البكتيريا للكشف مكتبات كبيرة من الأجسام المضادة 11-13. وهذه الأساليب هي قوية وفعالة لتحديد الأجسام المضادة التي ترتبط الأهداف، ولكنها تعتمد على مسار إفرازية لنقل البروتينات التي سيتم عرضها 14-16. مسار إفرازية translocates البروتينات المطوية من السيتوبلازم الحد في تجويف الشبكة الإندوبلازمية في الخميرة أو في الجبلة المحيطية في البكتيريا. ثم طي البروتينات في ظل ظروف المؤكسدة ويتم عرض على سطح الخلية أو تعبئتها الى جزيئات فج للكشف عن تقارب 17،18 ملزمة. ونتيجة لذلك، فإن الأجسام المضادة المعزولة باستخدام هذه التقنيات لا أضعاف بالضرورة كذلك في السيتوبلازم، ويجب أن تكون في كثير من الأحيان هندسة الذوبان داخل الخلايا بشكل منفصل إذا كان سيتم استخدام الأجسام المضادة في التطبيقات داخل الخلايا.
لتحسينفعالية الأجسام المضادة الهندسية التي هي مطوية بشكل جيد في السيتوبلازم، ونحن ذكرت في وقت سابق نجاح MAD-TRAP (عرض الراسية غشاء للاعتراف أساس تات من البروتينات ربط)، وهي طريقة لفحص مكتبة الأجسام المضادة scFv باستخدام كولاي inner- عرض غشاء 19. البكتيرية عرض الداخلي غشاء يعتمد على المسار المزدوج أرجينين النبات (تات) لنقل الأجسام المضادة عرض، على النقيض من طرق العرض المشترك الأخرى التي تستخدم مسار إفرازية. يحتوي على مسار تات آلية لمراقبة الجودة التي تسمح فقط للذوبان والبروتينات مطوية بشكل صحيح ليتم نقلها من E. السيتوبلازم القولونية، عبر الغشاء الداخلي، وفي الجبلة المحيطية 20،21. ركائز Overexpressed تات (أي والبروتينات التي تستهدف مسار تات مع الانصهار N-محطة لالببتيد إشارة تات ssTorA) التي يتم طيها بشكل جيد في السيتوبلازم تشكل النبات طويل الأجل المتوسط مع N-محطة طن السيتوبلازم ومحطة سي في الجبلة المحيطية 19. وهذا يسمح عرض من ركائز تات مطوية بشكل صحيح، بما في ذلك أجزاء الأجسام المضادة، على وجه محيط بالجبلة من E. الغشاء الداخلي القولونية. بعد إزالة الغشاء الخارجي عن طريق الهضم الأنزيمي لتوليد spheroplasts، تتعرض الأجسام المضادة إلى الفضاء خارج الخلية (الشكل 1). وهذا يسمح ركائز تات للظهور في الغشاء الداخلي ليتم فرزهم للربط إلى هدف محدد. الأهم من ذلك، تسخير مسار تات للعرض على سطح الخلية يضمن فقط الأجسام المضادة في المكتبة التي هي مطوية بشكل جيد في السيتوبلازم سيتم التحقيق معهم لمدة ملزمة، مما يسمح الهندسة المتزامنة لتقارب وقابلة للطي داخل الخلايا ملزمة. في هذا البروتوكول، ونحن تصف كيفية عرض مكتبة scFv على E. الغشاء الداخلي القولونية، عموم مكتبة ضد المستضد الهدف، وأداء الشاشة الثانوية للتعرف على المقومات الواعدة في المكتبة. بينما نحن نركز البروتوكول الخاص scFvs، يمكن تطبيق الطريقة لهندسة أي البروتين الذي يتطلب للطي ملزمة والخلايا التطبيق.
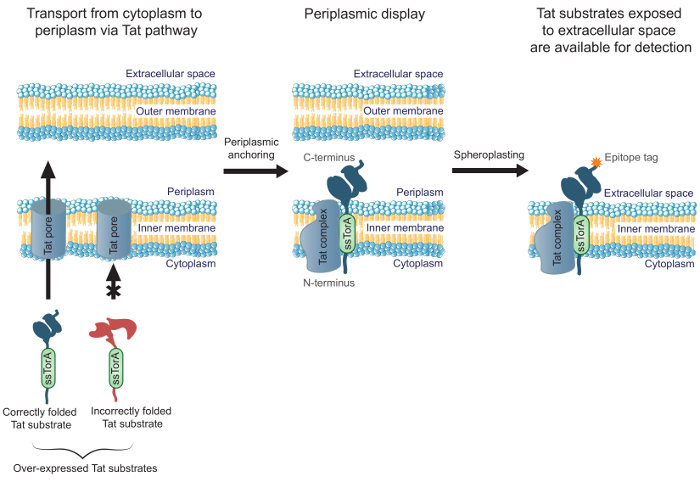
الشكل 1. تات عرض الداخلي غشاء. وفي E. القولونية، ويتم نقل الأجسام المضادة scFv التي يتم التعبير عنها كما انصهار لتسلسل إشارة ssTorA ومطوية في السيتوبلازم بشكل صحيح عبر الغشاء الداخلي. والنبات الأشكال الوسيطة، حيث ترتكز على scFvs في الغشاء الداخلي مع N-محطة في السيتوبلازم ومحطة سي في الجبلة المحيطية. وE. يتم هضم الغشاء الخارجي القولونية إنزيمي لتشكيل spheroplasts، وبالتالي تعريض الأجسام المضادة الراسية في الفضاء خارج الخلية وإتاحتها للكشف باستخدام الأجسام المضادة التي تربط إلى C-عضال العلامة حاتمة تنصهر على الضد المعروض.تحميل / 54583 / 54583fig1large.jpg "الهدف =" _ فارغة "> الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
Protocol
1. إعداد مكتبة scFv باعتباره فيوجن إلى ssTorA الإشارة تسلسل
- الحصول على مكتبة الحمض النووي الريبي منقوص الأكسجين (DNA) التي تحتوي على أنواع من الجينات scFv.
ملاحظة: يمكن أيضا إنشاء المكتبة باستخدام أي طريقة مناسبة لتوليد التنوع على الجينات scFv بأكمله أو المجالات المستهدفة 22 (على سبيل المثال، فإن التكامل الثالث تحديد المناطق، CDR3s). - إدراج مكتبة الحمض النووي في البلازميد pIMD (الشكل 2) باستخدام معيار أساليب الاستنساخ الجزيئي 23.
ملاحظة: هذا البلازميد يعبر عن scFvs كما التحام جيني لتسلسل إشارة ssTorA (N-محطة لscFv) وعلامة FLAG حاتمة (C-محطة لscFv). وقد وصفت تصميم البلازميد للعرض الغشاء الداخلي السابق 19. البلازميد pIMD متاح من المؤلفين.

الشكل 2. الداخلية غشاء عرض البلازميد (pIMD) خريطة (خطوات 1.2 خلال 1.3). يحتوي هذا البلازميد على PROMOTOR أمريكا اللاتينية والكاريبي، ColE1 أصل النسخ المتماثل، والجينة المقاومة الكلورامفينيكول. وتنصهر الجين scFv المدرج، على تسلسل إشارة ssTorA لاستهداف scFv إلى مسار تات وعلامة FLAG حاتمة، مع كل ثلاثة في نفس الإطار القراءة. يشار إلى مواقع الانزيم تقييد. لمكتبة إدراجها بين المواقع انزيم التقييد XbaI ونوتي، وحجم البلازميد هو 2219 بي بي زائد حجم scFv. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
- تحويل البلازميد التي تحتوي على مكتبة في MC4100 E. خلايا القولونية 23. تعافي ونمو هذا النوع البكتيري في المكتبة. أجهزة الطرد المركزي في 4000 × ز لمدة 15 دقيقة في RT لجمع الخلايا. إزالة طاف، و resuspend الخلايا التي تم جمعهافي 25٪ الجلسرين في لوريا، Bertani (LB) وسائل الاعلام. مخزن aliquots في -80 درجة مئوية لحين الحاجة إليها، أو الشروع في الخطوة 2.
ملاحظة: تم التحقق من بروتوكول مع خلايا MC4100، على الرغم من E. الآخرين ومن المتوقع أيضا سلالات بكتيريا لتكون متوافقة مع بروتوكول. Electroporation للهو الأسلوب المفضل للتحول، ويرجع ذلك إلى كفاءتها التحول عالية. يجب أن تتكون المكتبة عادة من 10 9 scFv المتغيرات في هذه المرحلة على الأقل، ويجب أن تحتوي كل قسامة ما يكفي من الخلايا بحيث يتم تغطية مكتبة 100 أضعاف.
2. التعبير عن المكتبة وإعداد Spheroplasts
- ذوبان الجليد قسامة واحدة من مكتبة البكتيرية (من الخطوة 1.3) في RT، وإضافة قسامة إلى قارورة تحتوي على 100 مل الإعلام LB مع 20 ميكروغرام / مل الكلورامفينيكول (سم). تنمو لمدة 3 ساعة على 37 درجة مئوية و 225 دورة في الدقيقة شاكر المحتضنة.
- بعد 3 ساعات، وإزالة قارورة من 37 درجة مئوية شاكر المحتضنة. السماح للتعبير عن مكتبة scFv لالمضي قدما O / N ل15 إلى 22 ساعة عند 20 درجة مئوية و 225 دورة في الدقيقة شاكر المحتضنة.
ملاحظة: ليس هناك حاجة إلى محفز عند استخدام البلازميد pIMD، والمروج هو المتسرب. لاحظ أن الخلايا MC4100 لا بإفراط على كاظمة لاك (ولم يتم العثور اسي على البلازميد).

الشكل 3. خلايا القولونية وspheroplasts. (A) E. خلايا القولونية هي اسطوانية الشكل. (B) بعد spheroplasting باستخدام EDTA والليزوزيم، الغشاء الخارجي للE. يتمزق خلايا القولونية، وspheroplasts الناتجة هم كروية الشكل. وقد تم الحصول على فرق تدخل النقيض (DIC) الصور المجهري باستخدام الهدف 100X على مجهر مقلوب. يرجى جلعق هنا لمشاهدة نسخة أكبر من هذا الرقم.
- إعداد spheroplasts المكتبة.
تتشكل Spheroplasts التي تمزق الغشاء الخارجي للE. ملاحظة: القولونية وتكون كروية الشكل (الشكل 3).- إعداد المخازن اللازمة.
ملاحظة: يجب أن تكون جميع مخازن العقيمة.- إعداد 1 × الفوسفات مخزنة المالحة (PBS، ودرجة الحموضة 7.4) عن طريق إذابة 8 ز كلوريد الصوديوم، 0.2 ز بوكل، 1.44 ز نا 2 هبو 4، و 0.24 ز KH 2 PO 4 في H المقطر 2 O إلى الحجم النهائي من 1000 مل. الحفاظ على الجليد.
- إعداد برنامج تلفزيوني مع 0.1٪ (ث / ت) البقري ألبومين المصل (BSA) عن طريق إذابة 0.2 غرام BSA إلى 200 مل 1 × برنامج تلفزيوني. الحفاظ على الجليد.
- إعداد المخزن المؤقت التجزئة (FB) عن طريق خلط 7.5 مل من العقيمة التي تمت تصفيتها 1 M السكروز، 1 مل من 1 عازلة M تريس (درجة الحموضة 8.0)، و 1.5 مل المقطر H 2 O. الحفاظ على الجليد.
- إعداد 1 ملم ثنائي أمين الإيثيلين رباعي حمض الخل (EDTA) وذلك بإضافة 30 ميكرولتر سو 0.5 M EDTA إلى 14.97 مل المقطر H 2 O.
- تحضير 0.5 M MgCl 2 عن طريق إذابة 4.76 ز MgCl 2 في 100 مل H 2 O. المقطر الحفاظ على الجليد.
- إزالة قارورة من شاكر، وقياس الكثافة الضوئية (OD) في 600 نانومتر باستخدام مقياس الطيف الضوئي لتحديد كثافة الخلية. حساب حجم ثقافة الناجم عن حاجة من هذا القبيل أن كل عينة لspheroplasting عدد 1 × 10 10 الخلايا.
ملاحظة: تقريب لOD 600 من 1 يشير إلى تركيز من 10 9 خلية / مل لE. القولونية يمكن استخدامها 24. - أجهزة الطرد المركزي في حجم يحسب للثقافة يتسبب في أنبوب 1.5 مل microcentrifuge في 12000 × ز في RT لمدة 5 دقائق. إعداد اثنين على الأقل من العينات في حال نشوء مسألة في إعداد العينات.
- إزالة طاف من الثقافات طرد وresuspend كل بيليه الخلية في 100 ميكرولتر من FB-الجليد الباردة. أجهزة الطرد المركزي في 12000 ×ز في RT لمدة 1 دقيقة، ثم إزالة طاف قبل pipetting. resuspend كل بيليه في 350 ميكرولتر من FB-المثلج تستكمل مع 3.5 ميكرولتر من 10 ملغ / مل الليزوزيم.
- ببطء دوامة كل أنبوب بينما تضيف، قطرة قطرة، 700 ميكرولتر من 1 ملم EDTA، ثم احتضان الأنابيب في RT لمدة 20 دقيقة في حين تناوب ببطء على محور دوار أنبوب لخلط العينات. إزالة الأنابيب من الدوار، وإضافة 50 ميكرولتر من الجليد الباردة 0.5 M MgCl 2 لكل أنبوب، واحتضان لهم على الجليد لمدة 10 دقيقة. أنابيب الطرد المركزي في 11000 × غرام عند 4 درجة مئوية لمدة 10 دقيقة.
- عزل بيليه الكوراء.
- استخدام micropipette مع طرف 1 مل لسحب ببطء جزء من بيليه. في حين عقد الأنبوب في زاوية مع افتتاح مباشرة فوق أنبوب 1.5 مل جديد، ورفع ببطء غيض ماصة للخروج من طاف وحرك بيليه في أنبوب جديد.
- إذا تم نقل حجم كبير من طاف لأنبوب جديد، إزالته من قبل pipetting. إذا كان بيليه ليس إينو شركةلاف لنقل، إعادة الطرد المركزي في 11000 × ز لمدة 2 دقيقة ومحاولة بيليه العزلة مرة أخرى.
- resuspend الكرية الكوراء في كل أنبوب في 1 مل من الجليد الباردة 1 × برنامج تلفزيوني. بالتناوب بين pipetting لوvortexing لببطء على حامل دوامة حتى يتم معلق بيليه تماما. لا تبقي عينات من الجليد لأكثر من 2 دقيقة في كل مرة، والعودة إلى الجليد لمدة 5 دقائق على الأقل قبل إزالة من الجليد مرة أخرى. الحفاظ على spheroplasts في 4 درجات مئوية (لمدة تصل إلى 2 أيام) حتى تستخدم لبالغسل في الخطوة 4.
- إعداد المخازن اللازمة.
3. شل الهدف مستضد على الخرز المغناطيسي
- Biotinylate المستضد الهدف في الجسم الحي خلال إنتاج المؤتلف في E. خلايا القولونية. بدلا من ذلك، استخدام الاقتران الكيميائية 25 أو المستضد الهدف الشراء التي تمت بالفعل البيروكسيديز، والشروع في الخطوة 3.2.
- إضافة 816 غرام bicine إلى 50 مل من الماء لجعل 10 × bicine العازلة. تمييع BUFفر إلى 1 × في المقطر H 2 O والحرارة إلى 50 درجة مئوية. إضافة 14.7 ملغ البيوتين إلى 12 مل من يسخن عازلة 1 × bicine لجعل الحل البيوتين الذي هو 5 ملم البيوتين في 10 ملي العازلة bicine. تخزين في -20 درجة مئوية لحين الحاجة إليها.
- التعبير عن وbiotinylate البروتين الهدف باستخدام البلازميد pAK400cb-BCCP 26، والذي يسمح إنتاج المستضد الهدف باعتباره الانصهار للبروتين البيوتين الكربوكسيل الناقل (BCCP).
ملاحظة: E. خلايا القولونية أصلا biotinylate BCCP، مما يلغي الحاجة إلى تنقية وbiotinylate البروتين المستهدفة قبل تجميد على حبات streptavidin المغلفة كيميائيا. الأصلي E. القولونية البيوتين يغاز البيرة غير كافية لbiotinylating البروتين الانصهار.- تنمو E. كولاي التي تحتوي على البلازميد biotinylation (مع المستضد الهدف إدراجها بوصفها الانصهار إلى N-محطة من BCCP) O / N ل15 إلى 18 ساعة في 5 مل من وسائل الاعلام LB تستكمل مع 20 ميكروغرام / مل سم عند 37 درجة مئوية في حين تهتز في 225 دورة في الدقيقة.
- قياس OD في 600 نانومتر باستخدام مقياس الطيف وحساب حجم الثقافة اللازمة (V إضافة) إلى ثقافة فرعية في التطوير التنظيمي بدءا من 0.05 في 25 مل الإعلام LB الطازجة مع 20 ميكروغرام / مل سم باستخدام المعادلة: V إضافة = (0.05 × 25 مل) / (OD 600-،05)، حيث OD 600 هو الكثافة البصرية للO / N الثقافة والخامس الإضافة هي حجم O / N الثقافة إضافة إلى LB. جديدة ثقافة فرعية وتنمو إلى OD من 0،5-0،8 في شاكر المحتضنة في 37 درجة مئوية و 225 دورة في الدقيقة.
- إضافة الآيزوبروبيل β-D-1-thiogalactopyranoside إلى تركيز النهائي من 100 ميكرومتر والبيوتين إلى تركيز النهائي من 5 ميكرومتر. حمل التعبير في شاكر المحتضنة لمدة 15 إلى 22 ساعة عند 20 درجة مئوية و 225 دورة في الدقيقة.
- البكتيريا الحصاد بواسطة الطرد المركزي في 4000 × ز عند 4 درجة مئوية لمدة 10 دقيقة. إزالة طاف. تخزين بيليه في -20 درجة مئوية لتصبح جاهزة للاستخدام.
- إضافة1 مل من المنظفات خلية تحلل في 0.2 غرام من بيليه الخلية. و resuspend من قبل pipetting وتناوب بلطف لمدة 20 دقيقة لليز الخلايا. بعد تحلل، الطرد المركزي في 16000 × ز و 4 درجات مئوية لمدة 20 دقيقة. ماصة للالمحللة للذوبان (طاف) في أنبوب 1.5 مل جديد.
- استخدام 3 كيلو دالتون العمود الوزن الجزيئي قطع لإزالة البيوتين غير منضم. ماصة للالمحللة في العمود، وأجهزة الطرد المركزي عند 20 درجة مئوية وفقا لتعليمات الشركة الصانعة. تغسل مع 1 × PBS حتى يتم تخفيفه البيوتين في المحللة 100 أضعاف وحجم المحللة غسلها يساوي حجم الأصلي للالمحللة. نقل المحللة إلى أنبوب جديد.
- شل المستضد الهدف المعقدة البيروكسيديز على حبات مغناطيسية streptavidin المغلفة.
- إعداد 1 × برنامج تلفزيوني و 1 × برنامج تلفزيوني مع 0.1٪ (ث / ت) BSA كما هو موضح في الخطوة 2.3.1.
- إعداد حبات مغناطيسية.
ملاحظة: هذا يتطلب استخدام رف الفصل المغناطيسي.- و resuspend streptavidin المغلفة حبات مغناطيسية في قارورة الأصلية. إما دوامة لمدة 30 ثانية على الأقل أو تدور لمدة 5 دقائق.
- نقل 7-10 × 10 9 حبات لأنبوب 1.5 مل.
ملاحظة: إن حجم المطلوب أن تعتمد على تركيز حبة المقدمة من قبل الشركة المصنعة. - وضع أنبوب يحتوي على حبات على الرف المغناطيس لمدة 2 دقيقة لجمع حبات على جانب الأنبوب. مع أنبوب لا يزال على المغناطيس، وإزالة بعناية طاف قبل pipetting دون تعطيل الخرز.
- أن يغسل، وإزالة أنبوب من المغناطيس، و resuspend حبات في 1 مل من 1 × برنامج تلفزيوني قبل pipetting دون توليد فقاعات. إعادة أنبوب إلى المغناطيس لمدة 2 دقيقة لجمع حبات، وإزالة بعناية طاف بواسطة pipetting. تكرار هذه العملية مرتين أخريين ليصبح المجموع ثلاثة يغسل. تأكد من عدم ترك أي السائل في الأنبوب بعد غسل النهائي.
- إضافة المحللة التي تحتوي على مستضد المعقدة البيروكسيديز إلى مكتب التحقيقات الفرنسى المغناطيسيس.
- إزالة أنبوب من المغناطيس و resuspend حبات في 1 مل من المحللة (من الخطوة 3.1.5). في احتضان RT لمدة 30 دقيقة في حين تناوب بلطف.
- وضع أنبوب على المغناطيس لمدة 3 دقائق لجمع حبات المغلفة المستضد. تغسل حبات مغلفة خمس مرات مع 1 × برنامج تلفزيوني مع 0.1٪ في جيش صرب البوسنة بنفس الطريقة كما هو موضح في خطوات 3.2.2.3 ل3.2.2.4. بعد غسل النهائي، resuspend والخرز في 1 × برنامج تلفزيوني مع 0.1٪ BSA تصل إلى نفس الحجم المستخدم في الخطوة 3.2.2.2.
- إذا كان المستضد الهدف يجمد مستقرة عند 4 درجات مئوية، وتخزين حبات المغلفة في 4 درجة مئوية لحين الحاجة إليها لبالغسل. خلاف ذلك، انتقل إلى الخطوة 4.
4. شاشة مكتبة scFv التي كتبها بالغسل ضد الهدف مستضد (الشكل 4)

الشكل 4. الطبخ (الخطوة 4). مستضد المغلفة الخرز المغناطيسي عالبريد حضنت مع spheroplasts التعبير عن المتغيرات مكتبة الأجسام المضادة. DNA البلازميد من spheroplasts محددة حبة يتم استرداد واستخدامها لتوليد sublibrary، الذي يتم فحص باستخدام شاشة ثانوية على أساس ELISA. المقابلة الخطوات بروتوكول لاحظت. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
- احتضان حبات مغلفة مع spheroplasts.
- استخدام الكوراء إلى حبة نسبة ما يقرب من 5: 1. إضافة 4 × 10 9 spheroplasts و 8 × 10 8 حبات لأنبوب 15 مل العقيمة.
ملاحظة: لنفترض أن أي خلايا فقدت أثناء عملية spheroplasting، وبالتالي فإن التركيز لا يزال 1 × 10 10 spheroplasts / مل. - إضافة 1 × برنامج تلفزيوني مع 0.1٪ BSA لتبرزي حجم الإجمالي إلى 4 مل. قسامة إلى أربعة أنابيب 1.5 مل مع 1 مل لكل منهما. احتضان ردود الفعل عند 4 درجة مئوية لمدة 5 ساعة في حين تناوب بلطف.
- استخدام الكوراء إلى حبة نسبة ما يقرب من 5: 1. إضافة 4 × 10 9 spheroplasts و 8 × 10 8 حبات لأنبوب 15 مل العقيمة.
- العلاقات العامةepare وspheroplasts محددة حبة لتفاعل البلمرة المتسلسل (PCR).
- وضع أنابيب رد فعل بالغسل على مغناطيس لمدة 3 دقائق. إزالة طاف قبل pipetting، وغسل spheroplasts محددة حبة أربع مرات مع المثلج 1 × برنامج تلفزيوني مع 0.1٪ في جيش صرب البوسنة بنفس الطريقة كما هو موضح في خطوات 3.2.2.3 ل3.2.2.4. Resuspend وspheroplasts محددة حبة في كل أنبوب في 25 ميكرولتر من H 2 O. المقطر تخزين حبات عند درجة حرارة -20 درجة مئوية أو الشروع في الخطوة 4.3.
- أداء كامل البلازميد PCR على spheroplasts محددة حبة لتضخيم البلازميدات تحتوي على جينات scFvs محددة حبة.
- الحصول التمهيدي مع تسلسل التالي: 5'CCAACTCTTTTTCCGAAGGTAACTG3 "(التمهيدي إلى الأمام) و5'TAGCTCTTGATCCGGCAAACAAA3" (التمهيدي العكسي).
ملاحظة: هذه سيربط بين نهاية إلى نهاية على مسارات نقيض من البلازميد pIMD (الشكل 2)، وهي مصممة ليصلب لسمة مشتركة من pIMD، لذلك سوف يحدث التضخيم بغض النظر عنscFv تسلسل البديل. - الفوسفات الاشعال.
ملاحظة: دون الفسفرة، وإعادة ربط-لن يحدث. ويمكن أيضا أن يؤمر الاشعال مع 5'-الفسفرة، بدلا من استخدام هذه الطريقة الفسفرة في هذا البروتوكول.- في أنبوب 0.5 مل، واقامة رد فعل الفسفرة لالتمهيدي PCR إلى الأمام كما هو موضح في الجدول رقم 1. كرر هذه العملية لالتمهيدي عكسي.
- احتضان ردود الفعل عند 37 درجة مئوية لمدة 1 ساعة. ثم احتضان لهم عند 65 درجة مئوية لمدة 20 دقيقة لإلغاء تنشيط كيناز T4 عديد النوكليوتيد (PNK). تخزين الاشعال فسفرته في -20 درجة مئوية.
- أداء PCR.
- في أنبوب PCR، وإعداد تفاعل PCR كما هو موضح في الجدول رقم (2).
ملاحظة: ردود فعل متعددة يمكن أن يكون على استعداد لعائد أعلى. يمكن تخزين spheroplasts محددة حبة غير المستخدمة عند درجة حرارة -20 درجة مئوية. - حرارة تفاعلات PCR في 98 درجة مئوية لمدة 15 دقيقة في cycler الحرارية لضمان تحلل الكامل من spheroplasts. إزالة أنابيب من cycler الحرارية، وإضافة 0.5 ميكرولتر من بوليميريز عالي الدقة لكل منهما. العودة أنابيب إلى cycler الحرارية وتشغيل باستخدام برنامج مفصل في الجدول 3.
- تجميع المنتجات PCR حسب الاقتضاء. تخزين في درجة حرارة -20 درجة مئوية أو الشروع في الخطوة 4.4.
- في أنبوب PCR، وإعداد تفاعل PCR كما هو موضح في الجدول رقم (2).
- الحصول التمهيدي مع تسلسل التالي: 5'CCAACTCTTTTTCCGAAGGTAACTG3 "(التمهيدي إلى الأمام) و5'TAGCTCTTGATCCGGCAAACAAA3" (التمهيدي العكسي).
الجدول 1. PNK الفسفرة رد فعل (الخطوة 4.3.2.1).
| كاشف | الحجم (ميكرولتر) |
| H المقطر 2 O | 15 |
| 10X T4 DNA العازلة رد فعل يغاز | 2 |
| 100 ميكرومتر التمهيدي | 2 |
| T4 عديد النوكليوتيد كيناز (PNK) | 1 |
| كاشف | الحجم (ميكرولتر) |
| H المقطر 2 O | 28.5 |
| 5X عالية الدقة عازلة البلمرة | 10 |
| 10 ميكرومتر فسفرته التمهيدي إلى الأمام | 2.5 |
| 10 ميكرومتر فسفرته التمهيدي العكسي | 2.5 |
| 40 ملي مزيج dNTP (10 ملم كل dNTP) | 1 |
| spheroplasts محددة حبة | 5 |
الجدول 3. برنامج PCR (الخطوة 4.3.3.2).
| خطوة | درجة الحرارة (° C) | الوقت (دقيقة: ثانية) | عدد الدورات |
| بدل طبيعة الأولية | 98 | 00:30 | 1 |
| تفسد | 98 | 00:10 | 35 |
| الصلب | 69 | 00:30 | |
| تمديد | 72 | 00:30 في كيلوبايت | |
| تمديد النهائي | 72 | 06:00 | 1 |
| عقد | 12 | غير محدود | 1 |
- إعادة التعميم المنتجات PCR كاملة-البلازميد، واستخدام المنتج ligated لتحويل MC4100 E. خلايا القولونية.
- تنقية المنتج PCR عن طريق تشغيل تفاعل PCR على هلام الاغاروز 23، تلطيخ الحمض النووي في جل 23، واستخدامهلام عدة تنظيف لتنقية البلازميد خطي عن طريق اتباع التعليمات المرفقة من قبل الشركة المصنعة. قياس تركيز باستخدام معمل في 260 نانومتر. تخزين جزء تنقيته في -20 درجة مئوية لحين الحاجة إليها، أو الاستمرار في الخطوة 4.4.2.
- إعادة التعميم البلازميد من المنتج PCR.
- لمنع ربط الجزيئات من الناتج PCR، تنفيذ رد فعل الربط مع تركيز منخفض 27 من 1 نانوغرام / ميكرولتر من الناتج PCR. حساب حجم اللازمة لإعداد ربط رد فعل 800 ميكرولتر في هذا التركيز.
- إعداد رد فعل الربط على الجليد. في أنبوب، إضافة حجم المحسوبة في الخطوة 4.4.2.1 من الناتج PCR، 80 ميكرولتر من العازلة يغاز 10 × الحمض النووي، والمقطر H 2 O ما يصل إلى 800 ميكرولتر. إضافة 4 ميكرولتر من يغاز T4 الحمض النووي، والمكان على الفور أنابيب في 16 درجة مئوية في حمام مائي أو cycler الحرارية. احتضان عند 16 ° CO / N ل14 إلى 18 ساعة. تخزين ردود الفعل ربط أنجزت في -20 ° Cلحين الحاجة إليها، أو انتقل إلى الخطوة 4.4.3.
- ضع رد فعل ربط على كتلة الحرارة عند 65 درجة مئوية لمدة 15 دقيقة لتسخين-تعطيل يغاز الحمض النووي. ثم استخدام غشاء microdialysis أو تنظيف الحمض النووي عدة للتخلص من الملح الحمض النووي ligated. تخزين عند 20 درجة مئوية أو انتقل إلى الخطوة 4.4.4.
- استخدام كامل الحرارة المعطل، المملحة دي المنتج ربط لتحويل MC4100 E. خلايا القولونية 23. إعداد مخزون الجلسرين، كما هو موضح في الخطوة 1.3، من الخلايا التي تحتوي على sublibrary انتقادات الناتجة عن ذلك، وقسامات مخزن في -80 درجة مئوية.
- كرر الخطوة 4 في مجملها باستخدام قسامة من الخطوة 4.4.4 القيام بالغسل الثاني على sublibrary.
ملاحظة: بالغسل الثاني يساعد على إثراء للمكونات المكتبة التي تربط جيدا لالمستضد الهدف (19).
5. إجراء الشاشة الثانوية باستخدام المناعي الفحص الطريقة انزيم مرتبط إلى تحديد النسخ واعدة لمزيد من توصيف (الشكل 5) </ P>

يتم تلقيح الفحص بناء ELISA-الشكل 5. الثانوي (الخطوة 5). مكتبة (أ) المتغيرات من sublibrary التخصيب خلال بالغسل في الآبار الفردية من لوحة الثقافة للنمو والتعبير. وهي مغلفة (B) لوحة إليسا مع المستضد الهدف. يتم فحص (C) والمتغيرات المكتبة باستخدام الشاشة الثانوية أساس ELISA وصفها في البروتوكول. على تحليل البيانات التي تم الحصول عليها من الشاشة الثانوية، ويتم اختيار متغيرات المصالح وتتميز أبعد من ذلك. المقابلة الخطوات بروتوكول لاحظت. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
- ذوبان الجليد أنبوب واحد من sublibrary انتقادات (من الخطوة 4.4.4) ولوحة على لوحات أجار LB. لوحة عدة التخفيفاتبتركيزات منخفضة بما فيه الكفاية لضمان المستعمرات الفردية (على سبيل المثال، 10 2 - 10 6 التخفيفات أضعاف). احتضان لوحات لمدة 15 إلى 18 ساعة على 37 درجة مئوية. تخزين لوحات في 4 درجة مئوية أو الشروع في الخطوة 5.2.
- الثقافة ولحث على المستعمرات من sublibrary انتقادات. تنفيذ كافة الخطوات تحت ظروف معقمة. استخدام pipettor الأقنية لخطوات تشمل لوحات 96-جيدا.
- إضافة 200 ميكرولتر من LB مع 20 ميكروغرام / مل سم في كل بئر من لوحة ثقافة 96-جيدا ذهابا والقاع.
- اختيار مستعمرة الفردية من لوحة أجار مع طرف ماصة، ضع طرف في أول بئر من لوحة 96-جيدا، ويحرك برفق لتطعيم. استخدام معلومات جديدة لكل بئر. تطعيم مستعمرة واحدة في كل بئر. كعنصر تحكم، وتشمل السيطرة العقم واحدة على الأقل بشكل جيد مع أي مستعمرة تلقيح.
- كرر خطوات 5.2.1 و 5.2.2 لتطعيم عدة لوحات 96-جيدا.
- وضع لوحات 96-جيدا على شاكر صفيحة ميكروسكوبية 310 دورة في الدقيقة. في احتضان 376؛ مئوية لمدة 20 إلى 24 ساعة للتعبير عن scFvs.
- لكل لوحة الثقافة التي أعدت في الخطوة 5.2، طبقة واحدة 96-جيدا لوحة ELISA مع المستضد الهدف.
- تمييع النقي المستضد الهدف إلى التركيز المناسب (على سبيل المثال، 1 ميكروغرام / مل إلى 4 ميكروغرام / مل) في 1 × برنامج تلفزيوني لجعل حل الطلاء. جعل 5 مل من محلول طلاء لكل لوحة 96-جيدا.
ملاحظة: إن التركيز المناسب يعتمد على مستضد معين المستخدمة وقد تحتاج إلى تعديل. - إضافة 50 ميكرولتر من محلول طلاء على كل جانب من واضحة البوليسترين لوحة ELISA 96 جيد مرتفع ملزم. اضغط برفق لوحة على سطح الفوق لضمان كامل سطح كل بئر المغلفة. كرر لكل لوحة. احتضان لوحات عند 4 درجات CO / N.
- تمييع النقي المستضد الهدف إلى التركيز المناسب (على سبيل المثال، 1 ميكروغرام / مل إلى 4 ميكروغرام / مل) في 1 × برنامج تلفزيوني لجعل حل الطلاء. جعل 5 مل من محلول طلاء لكل لوحة 96-جيدا.
- تكرار المستعمرات من لوحات ثقافة 96-جيدا على لوحات أجار.
- وضع النسخ المتماثل البوليسترين عقيمة في آبار لوحة الثقافة لجمع كمية صغيرة من LIQUهوية شخصية. رفع بعناية المكرر ونقل إلى 15 سم LB لوحة أجار مثل أن كل النصائح ولمس لوحة. مرة واحدة وقد نقل السائل، ورفع النسخ المتماثل على التوالي. كرر لكل لوحة الثقافة.
- تسمية لوحة أجار مع التوجه الصحيح بحيث النتائج من الشاشة الثانوية في لوحة 96-جيدا يمكن أن تتطابق مع مستعمرة تكرارها الصحيحة على لوحة، وإذا كان المطلوب مزيد من التوصيف. ينمو بمعدل 37 درجة مئوية لمدة 15 إلى 18 ساعة، ومن ثم تخزينها في 4 درجات مئوية لحين الحاجة إليها.
- أداء الشاشة الثانوية ELISA.
- إعداد عرقلة الحل بجعل 2٪ (ث / ت) الحليب الجاف في 1 × برنامج تلفزيوني. تفريغ حل طلاء من لوحات ELISA. إضافة 100 ميكرولتر من عرقلة الحل إلى كل بئر. في احتضان RT لمدة 2 ساعة على الأقل، أو كتلة O / N عند 4 درجات مئوية.
- إعداد غسل العازلة بإضافة بوليسوربات 20 إلى تركيز النهائي من 0.05٪ في 1 × برنامج تلفزيوني. جعل 250 مل لكل لوحة ELISA.
- إضافة 20 ميكرولتر منالمركز خلية تحلل المنظفات لكل بئر من لوحة الثقافة ذهابا والقاع، واحتضان لوحة الثقافة على شاكر صفيحة ميكروسكوبية RT لمدة 15 إلى 20 دقيقة. بدء تحلل في الوقت نفسه إلى أن حجب لوحات ELISA اكتمال بحيث لا يمكن أن يؤديها تحلل وغسل الخطوة 5.5.4 بشكل متزامن.
- تفريغ حل عرقلة من لوحات ELISA. تغسل الصحون ELISA منع اربع مرات مع 200 ميكرولتر من غسل العازلة لكل بئر في غسل. إفراغ غسل العازلة من الآبار.
- نقل 50 ميكرولتر من كل بئر من لوحة خلية تحلل المقابلة بئر من لوحة ELISA، وذلك باستخدام معلومات جديدة لكل بئر. احتضان لوحة ELISA على RT لمدة 1-2 ساعة.
- إعداد الحل الضد للكشف عن scFvs ملزمة.
- استخدام البيروكسيديز الفجل (HRP) -conjugated الأجسام المضادة الأولية التي تربط لعلامة FLAG حاتمة تنصهر إلى scFvs المكتبة.
- تمييع الضد إلى التخفيف المناسبة لاستخدامها في ELISA (انظر المورد & #39؛ ق توصيات) في 2٪ (ث / ت) الحليب الجاف في 0.05٪ بوليسوربات 20 في 1 × برنامج تلفزيوني. إعداد 5 مل لكل لوحة.
- تغسل الصحون ELISA أربع مرات مع غسل العازلة كما هو موضح في الخطوة 5.5.4.
- إضافة 50 ميكرولتر من محلول الأجسام المضادة إلى كل بئر من لوحة ELISA. احتضان لمدة 1-2 ساعة على RT.
- إعداد الركيزة HRP عن طريق إذابة س -phenylenediamine هيدروكلوريد (العيادات الخارجية) أقراص في H المقطر 2 O في بروتوكول الشركة المصنعة مع تجنب ضوء. إعداد 20 مل لكل لوحة ELISA.
- إعداد 3 MH 2 SO 4 عن طريق تمييع H مركزة 2 SO 4 مع H المقطر 2 O عند الضرورة. إعداد 5 مل لكل لوحة ELISA.
تحذير: H 2 SO 4 هو حمض قوي. تأكد من ارتداء معدات الوقاية الشخصية المناسبة. - تغسل الصحون ELISA أربع مرات مع غسل العازلة، كما هو موضح في الخطوة 5.5.4.
- احتضان لوحات ELISA مع substra HRPالشركة المصرية للاتصالات.
- إضافة 200 ميكرولتر من الركيزة HRP إلى كل بئر. لتقليل التعرض للضوء، إضافة الركيزة لوحة ELISA واحد في وقت واحد، والتفاف مع رقائق الألومنيوم قبل الانتقال إلى لوحة القادمة. احتضان لوحات لمدة 30 إلى 60 دقيقة في RT في الظلام.
- بعد 30 دقيقة الأولى، والتحقق من لوحات لسواد الركيزة، واحتضان أطول إذا لزم الأمر لتصور وضع اللون.
- إضافة 50 ميكرولتر من 3 MH 2 SO 4 إلى كل بئر لإرواء رد الفعل. باستخدام طرف مختلف لكل بئر، ومزيج الحل في الآبار من قبل pipetting بلطف صعودا وهبوطا دون مزبد. للحصول على تناسق ومنع التشبع، إضافة H 2 SO 4 بسرعة وبدقة لجميع لوحات ELISA قبل خلط الحل لكل لوحة.
- قياس الامتصاصية من الحل في الآبار من كل لوحة في 492 نانومتر باستخدام قارئ لوحة.
- تحليل البيانات الامتصاصية لتحديد scFv المتغيرات التي exhibانها واعدة إشارات ملزمة وتميز هذه scFvs واعدة. اختر scFvs التي تظهر إشارات الامتصاصية أعلى من إشارة الخلفية وأعلى من متوسط إشارة على كل لوحة.
ملاحظة: إن مستوى الامتصاصية أن تعتمد على خصائص المستضد والأجسام المضادة لمكافحة FLAG المستخدمة، جنبا إلى جنب مع قوة من المتغيرات scFv التي تم عزلها في الفرز.
Representative Results
البروتين للطي آلية لمراقبة الجودة داخل الخلايا من مسار تات من E. القولونية يحد النقل عبر غشاء الخلية الداخلي للبروتينات التي يتم طيها بشكل جيد في بيئة هيولي الحد. بواسطة overexpressing التحام من scFv لتسلسل إشارة ssTorA (تسلسل إشارة من البروتين طرة، الذي نقل بشكل طبيعي في مسار تات 20)، والمتوقفة النبات، مما أدى إلى عرض scFvs على الغشاء الداخلي 19. بعد انقطاع الأنزيمية للغشاء الخارجي، مصنوعة الأجسام المضادة عرض متاحة للفحص لنشاط ملزم المستضد. وقد أظهرت القدرة على الاستفادة من مسار تات للعرض scFv التي كارلسون وآخرون. 19 (الشكل 6). وتنصهر الأجسام المضادة scFv scFv13 وscFv13.R4 إما التسلسل ssTorA الأصلي أو ssTorA المحورة التي تفتقر الزوج بقايا أرجينين أرجينين معترف بها من قبلمسار تات. تم تصميم scFv13.R4 التي كتبها مارتينو وآخرون من scFv13 من خلال أربع جولات من التطور الموجه وكما هو معروف لأضعاف جيدا في السيتوبلازم 9. تم عرض هذه scFv على الغشاء الداخلي، ولكن فقط عندما يعبر عنه كنسبة الانصهار إلى الأصلي تسلسل إشارة ssTorA (الشكل 6). على العكس، هو scFv13 لا مطوية جيدا cytoplasmically 9، بحيث لا يتم عرضها بشكل جيد على الغشاء الداخلي، بغض النظر عن تسلسل إشارة إلى الذي تنصهر فيه. بالإضافة إلى ذلك، إذا تم أعرب عن scFvs في الخلايا التي تفتقر إلى البروتين TatC، عنصرا حيويا لآلية تات 20،28، لم يكن لوحظ العرض، تظهر حلقة وصل هامة بين العرض الداخلي غشاء ومسار تات. وهذه النتائج يثبت أن فقط البروتينات التي تحتوي على تات إشارة الببتيد وتكون صحيح الايدي السيتوبلازم يتم عرض على الداخلي الغشاء السماح النقل عن طريق تات طريق في وظيفة شاشة لداخل الخلايا فل دينغ.

تم تنفيذ الشكل 6. الكشف عن scFvs ظهور في الغشاء الداخلي. التدفق الخلوي تحليل للكشف عن عرض من سوء مطوية scFv13 ومطوية جيدا scFv13.R4 على الغشاء الداخلي. وتنصهر scFvs إلى ssTorA مواطن أو ssTorA (KK)، حيث تم تعديل الزوج الارجنتين-الارجنتين في تسلسل ssTorA إلى ليز-اليس. تم الكشف عن العلامات FLAG حاتمة C-محطة على scFvs مع ثيوسيانات فلوريسئين (FITC) -conjugated مكافحة FLAG الأجسام المضادة. تم اختبار الخلايا دون البروتين TatC (ΔtatC) وssTorA-scFv13 دون علامة FLAG والضوابط. M يشير إلى قيمة مضان متوسط. نقلا عن مرجع 19 بإذن. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
س: المحافظة على together.within الصفحات = "1"> عرض الداخلية غشاء يمكن عزل بنجاح الأجسام المضادة scFv مع مستويات عالية من تقارب لبروتين هدف ومستويات عالية من الذوبان حشوية. بالإضافة إلى ذلك، جولات لاحقة من التطور الموجه باستخدام شاشة الداخلي غشاء تحسين خصائص الأجسام المضادة 19. وللتدليل على ذلك، مكتبة PCR عرضة للخطأ استنادا scFv13، الذي لديه انخفاض مستوى ملزمة تقارب لβ غالاكتوزيداز، لانتقادات ضد المستضد الهدف β غالاكتوزيداز باستخدام الشاشة وبالغسل الطريقة الموصوفة في هذا البروتوكول. تم عزل scFv 1-4 بعد جولة واحدة من الطفرات وبالغسل، وعرضت تقارب ملزم العالي للبيتا غالاكتوزيداز من scFv13 (الشكل 7A) ومستوى أعلى من الذوبان حشوية (7B الشكل).
مكتبة جديدة، على أساس scFv 1-4، أحرز باستخدام عرضة للخطأ PCR، وبالغسل من هذه المكتبة الجيل الثاني ضدβ غالاكتوزيداز تم استخدام تعديل بروتوكول صفها. وقد تم بالغسل ضد β غالاكتوزيداز للجولة الثانية من تطور في وجود النقي، قابل للذوبان scFv 14 كمنافس لتحسين احتمالات لعزل الحيوانات المستنسخة مع تقارب أعلى من scFv 1-4. بعد هذه الجولة الثانية من الطفرات وبالغسل، scFv 2-1 و2-3 scFv تم عزل باستخدام الفحص الثانوي على أساس ELISA. هذه scFvs لا تعرض فقط تقارب ملزم العالي للβ غالاكتوزيداز من scFv13، ولكن أيضا عرضت ملزم أفضل من استنساخ من الدور الاول scFv 1-4. عرضت scFv 2-1 بيتا غالاكتوزيداز ملزمة مماثلة لتلك التي scFv13.R4 (7A الشكل). يظهر scFv 2-3 أيضا زيادة أخرى في الذوبان حشوية مقارنة scFv 14، وتسليط الضوء على الهندسة في وقت واحد من الذوبان وملزم المستضد. منذ يتم فحص تقارب والتعبير القابلة للذوبان في scFvs لفي وقت واحد، فمن الممكن أن scFv اختيار وزارة الدفاعالذوبان erate لكن بالعكس ملزم أو العكس عالية. على سبيل المثال، scFv 2-1 لديها أقل تعبير قابلة للذوبان من scFv 2-3، لكنه يسلك تقارب ملزم العالي للبيتا غالاكتوزيداز.

الرقم 7. الهدف ملزم والتعبير حشوية من scFv المتغيرات عزل تستخدم شاشة الداخلي غشاء. وأعرب عن (A) scFvs في سيتوبلازم E. خلايا القولونية (على سبيل المثال، من دون تسلسل إشارة ssTorA) مع hexahistidine (6 × العاهل) العلامة وتنقيته باستخدام حمض تدور الأعمدة والنيكل nitrilotriacetic. وقد تم قياس الربط من scFvs تنقيته لبيتا غالاكتوزيداز مع إليسا. تم تحميل scFvs تنقيته على لوحات ELISA المغلفة β غالاكتوزيداز، وتم الكشف عن scFvs ملزمة مع × العاهل الأجسام المضادة 6 المضادة. البيانات هي في المتوسط ستة مكررات، ويعرض شريط الخطأ خطأ المعياري للمتوسط.(ب) القابلة للذوبان كسور وغير قابلة للذوبان في الخلية لست] من خلايا معربا عن scFvs cytoplasmically تم تحليلها بواسطة لطخة غربية بحث مع و× العاهل الأجسام المضادة 6 المضادة. وقد استخدم تركيز البروتين الكلي لتطبيع تحميل العينات. طبع (A) وتكييفها (ب) من إشارة 19 بإذن. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
Discussion
هندسة الأجسام المضادة للنشاط حشوية مهمة صعبة نظرا للبيئة والحد من السيتوبلازم، مما يعوق تشكيل استقرار السندات كبريتيد 6،7. هذا يسبب الكثير من الأجسام المضادة ليكون غير نشط cytoplasmically إلا إذا صممت أنهم من أجل الاستقرار والقابلية للذوبان في السيتوبلازم، بالإضافة إلى كونها هندسيا لتقارب ملزمة. الأساليب الحالية لعرض فج، وعرض سطح البكتيرية، وطرق عرض الخميرة سطح كل استخدام مسار إفرازية 14-16 لعرض الأجسام المضادة هندسيا، ولكن هذه الأساليب ليس لديهم وسيلة لهندسة قابلة للطي داخل الخلايا. وتحسنت الأجسام المضادة هندسيا باستخدام العرض الداخلي غشاء الاستقرار حشوية والذوبان لمراقبة الجودة للطي لمسار تات يمنع النبات من الأجسام المضادة التي يتم سيئة مطوية وغير مستقرة في السيتوبلازم. هذه الطريقة يبسط عملية تكرارية من الأجسام المضادة الهندسة داخل الخلايا لتقارب لالثانية الذوبان، كما صممت خاصيتين في خطوة واحدة. وعلى الرغم من تصميم هذه الطريقة لهندسة الأجسام المضادة مع الذوبان في البيئة داخل الخلايا الحد، فإنه يمكن أيضا أن تطبق على الأجسام المضادة الهندسة على العمل في ظروف غير الحد، لأن البروتينات هندسيا باستخدام هذه الطريقة تحافظ قابلة للطي في البيئة المؤكسدة من الجبلة المحيطية.
على الرغم من أن هذه التقنية يبسط عملية هندسة الأجسام المضادة مع قابلية عالية والذوبان هيولي عالية، العديد من القيود هامة يجب مراعاتها عند استخدام هذا البروتوكول. عند تحليل الإشارات شاشة ELISA الثانوية لتحديد واعدة scFv المتغيرات، عتبة تمييز بين متغيرات يحتمل أن تكون مثيرة للاهتمام وتلك التي قد لا تظهر كافية ملزمة مستضد ليس من المرجح أن تكون واضحة حتى بعد عدة استنساخ تم التعرف على أكثر من ذلك. من المهم أن نبحث عن تحسن ملزمة على الأجسام المضادة الأم. ومع ذلك،إشارة عالية بشكل غير طبيعي يمكن أن يكون مؤشرا على الطمع 29 أو آثار التجميع 30، وهو تحد ليست وحيدة النهج عرض الفحص الداخلي غشاء. وجود قيود الواجب تذكره عند استخدام هذا البروتوكول هو عدم القدرة على استعادة spheroplasts بعد بالغسل، لأنها غير قابلة للحياة (بيانات غير منشورة). وهذا يستدعي التضخيم الحمض النووي والتحول خطوات لاسترداد البلازميدات ترميز الأجسام المضادة.
العديد من الخطوات الهامة للبروتوكول تمكن الهندسة المتزامنة للطي وملزمة من الأجسام المضادة. لفحص لتكون ناجحة، يجب أن أعرب مكتبة scFv يجري فرزهم كما انصهار للإشارة الببتيد ssTorA. بدون هذا التسلسل، لن يتم توجيه الأجسام المضادة لمسار تات وبالتالي لن يكون translocated إلى الجبلة المحيطية 19. بالإضافة إلى ذلك، لا بد من أن علامة حاتمة C-محطة وتنصهر إلى الأجسام المضادة للسماح للكشف عن الأجسام المضادة المعروضة في بنالمقايسات دينغ. ومن الواضح أن E. كولاي سلالة تستخدم للتعبير يجب أن يكون لديك ما يلزم تات الآلات ممر لscFvs، ولكن هذا ينطبق على E. يشيع استخدامها سلالات القولونية.
تعديلات على هذا البروتوكول هي الممكنة لتحسين قدرتها على عزل الأجسام المضادة مع الخصائص المطلوبة. قد يتم الانتهاء خطوة بالغسل مطروح قبل بالغسل ضد المستضد الهدف في استنزاف المكتبة scFv من المكونات غير المرغوب. وspheroplasts مكتبة يمكن حضنت مع الخرز المغناطيسي المغلفة مع BCCP وحدها أو المغلفة مع بروتين غير المرغوبة، وspheroplasts التي تربط لتلك الخرزات يمكن التخلص منها قبل الفرز وspheroplasts غير منضم المتبقية للربط إلى الهدف المنشود. كما جاء في نتائج التمثيلية، وهي طريقة لتحسين تقارب من scFv معزولة لتشمل منافس للذوبان في رد فعل بالغسل للتنافس مع scFvs ظهور في spheroplasts. لأن شركات القابلة للذوبانetitor هو البروتين النقي، يتم تضخيمه أي الحمض النووي من ذلك، سيتم استردادها تسلسل ذلك فقط من scFvs ظهور في spheroplasts في رد فعل PCR. بالإضافة إلى ذلك، يمكن أن يمتد هذا الأسلوب لهندسة أنواع أخرى من الأجسام المضادة أو البروتينات غير ملزم الأجسام المضادة.
كولاي عرض الداخلية غشاء هو منصة قوية لأجسام هندسية مع قابلية عالية ومستويات عالية من الذوبان داخل الخلايا. هذا الأسلوب هو مناسبة خاصة للهندسة فعالة من الأجسام المضادة مصممة للعمل في البيئة داخل الخلايا. ويجري بالفعل استكشاف هذه الأجسام المضادة الخلايا كما العلاجات المحتملة في عدد من المجالات، بما في ذلك أمراض الاعصاب والسرطان والالتهابات الفيروسية 31. هذه التقنية سوف تتيح استخدام أكثر انتشارا من الأجسام المضادة الخلايا كأدوات للبحث والطب في هذه المجالات وفي أي ميدان آخر حيث الدراسة والتعلم هو المطلوب الهدف البروتين في الموقع.
Materials
| Name | Company | Catalog Number | Comments |
| scFv library | Varies | A suitable scFv library should be obtained from a commercial or academic source. | |
| MC4100 E. coli cells | Coli Genetic Stock Center | 6152 | Cells need to be chemically competent or electrocompetent, depending on the selected transformation method. |
| Glycerol | Fisher Scientific | BP229-4 | |
| Difco dehydrated culture media LB Broth, Miller (Luria-Bertani) | BD | 244610 | |
| Chloramphenicol (Cm) | Fisher Scientific | BP904-100 | |
| Sodium chloride (NaCl) | Fisher Scientific | BP358-1 | |
| Potassium chloride (KCl) | Fisher Scientific | BP366-500 | |
| Sodium phosphate, dibasic (Na2HPO4) | Fisher Scientific | BP332-500 | |
| Potassium phosphate, monobasic (KH2PO4) | Fisher Scientific | BP362-500 | |
| Bovine serum albumin (BSA) | Fisher Scientific | BP9706-100 | |
| Sucrose | Fisher Scientific | BP220-1 | |
| Tris base | Fisher Scientific | BP1521 | |
| Ethylenediaminetetraacetic acid (EDTA), 0.5 M | Fisher Scientific | BP2482-500 | |
| Magnesium chloride (MgCl2) | Fisher Scientific | BP214-500 | |
| Lysozyme | Sigma Aldrich | L3790-10X1ML | |
| Vortex mixer | VWR | 97043-564 | |
| Bicine | Fisher Scientific | BP2646100 | |
| D-Biotin | Fisher Scientific | BP232-1 | |
| Isopropyl β-D-1-thiogalactopyranoside | Fisher Scientific | BP1755-1 | |
| BugBuster Master Mix (cell lysis detergent) | EMD Millipore | 71456 | |
| Vivaspin 2 MWCO, 3,000 daltons | GE Healthcare Sciences | 28932240 | |
| Target antigen | Varies | N/A | Purified target antigen may be purchased or produced/purified. |
| Dynabeads MyOne Streptavidin T1 | Invitrogen | 65601 | |
| Dynamag-2 magnet | Invitrogen | 12321D | |
| Tube rotator | VWR | 13916-822 | |
| PCR primers | IDT | N/A | Primer sequences are as described in the protocol. |
| 10x T4 DNA ligase reaction buffer | New England BioLabs | B0202S | |
| T4 Polynucelotide kinase (PNK) | New England BioLabs | M0201S | Make sure the T4 ligase buffer used in the primer phosphorylation reaction contains 1 mM ATP. |
| 5x Phusion HF buffer pack | New England BioLabs | B0518S | |
| Deoxynucleotide (dNTP) solution mix, 10 mM each dNTP | New England BioLabs | N0447L | |
| Phusion DNA polymerase | New England BioLabs | M0530S | Other high-fidelity polymerases may be used as an alternative, but the annealing temperature in Table 3 must be adjusted. |
| C1000 Touch thermal cycler with dual 48/48 fast reaction module | Bio-Rad | 185-1148 | |
| Agarose | Promega | V3121 | |
| SYBR Safe DNA gel stain | Invitrogen | S33102 | |
| Wizard SV gel and PCR clean-up system | Promega | A9281 | |
| T4 DNA ligase | New England BioLabs | M0202S | |
| Microdialysis membrane filter | EMD Millipore | VSWP04700 | |
| Agar | BD | 214030 | |
| 96-well polystyrene round-bottom cell culture plates | VWR | 10062-902 | |
| Costar general polystyrene assay plate lids | Corning | 3931 | |
| Microtitre plate shaker | VWR | 12620-926 | |
| Costar 96 well EIA/RIA Easy Wash clear flat bottom polystyrene high bind microplate | Corning | 3369 | |
| Bel-blotter polycarbonate 96-well replicating tool | Bel-Art Products | 378760002 | |
| Instant nonfat dry milk | Quality Biological | A614-1000 | |
| Tween 20 (polysorbate 20) | Fisher Scientific | BP337-500 | |
| PopCulture reagent (concentrated cell lysis detergent) | EMD Millipore | 71092-3 | |
| Monoclonal ANTI-FLAG M2-Peroxidase(HRP) antibody produced in mouse | Sigma Aldrich | A8592 | |
| SigmaFast OPD | Sigma Aldrich | P9187-50SET | |
| Sulfuric acid (H2SO4), 10 N solution | Fisher Scientific | SA200-1 | |
| Reynolds Wrap aluminum foil | VWR | 89079-075 | |
| BioTek Epoch microplate spectrophotometer | Fisher Scientific | 11120570 |
References
- Biocca, S., Pierandrei-Amaldi, P., Campioni, N., Cattaneo, A. Intracellular immunization with cytosolic recombinant antibodies. Biotechnology (NY). 12 (4), 396-399 (1994).
- Chen, S. Y., Bagley, J., Marasco, W. A. Intracellular antibodies as a new class of therapeutic molecules for gene-therapy. Hum. Gene Ther. 5 (5), 595-601 (2008).
- Gargano, N., Biocca, S., Bradbury, A., Cattaneo, A. Human recombinant antibody fragments neutralizing human immunodeficiency virus type 1 reverse transcriptase provide an experimental basis for the structural classification of the DNA polymerase family. J Virol. 70 (11), 7706-7712 (1996).
- Mhashilkar, A. M., et al. Inhibition of HIV-1 Tat-mediated LTR transactivation and HIV-1 infection by anti-Tat single chain intrabodies. Embo J. 14 (7), 1542-1551 (1995).
- Strube, R. W., Chen, S. Y. Characterization of anti-cyclin E single-chain Fv antibodies and intrabodies in breast cancer cells: enhanced intracellular stability of novel sFv-F-c intrabodies. J. Immunol. Meth. 263 (1-2), 149-197 (2002).
- Mössner, E., Koch, H., Plückthun, A. Fast selection of antibodies without antigen purification: adaptation of the protein fragment complementation assay to select antigen-antibody pairs. J. Mol. Biol. 308 (2), 115-122 (2001).
- Wörn, A., et al. Correlation between in vitro stability and in vivo performance of anti-GCN4 intrabodies as cytoplasmic inhibitors. J Biol Chem. 275 (4), 2795-2803 (2000).
- Knappik, A., Plückthun, A. Engineered turns of a recombinant antibody improve its in vivo folding. Protein Eng. 8 (1), 81-89 (1995).
- Martineau, P., Jones, P., Winter, G. Expression of an antibody fragment at high levels in the bacterial cytoplasm. J Mol Biol. 280 (1), 117-127 (1998).
- Steipe, B., Schiller, B., Plückthun, A., Steinbacher, S. Sequence statistics reliably predict stabilizing mutations in a protein domain. J. Mol. Biol. 240 (3), 188-192 (1994).
- Daugherty, P. S. Protein engineering with bacterial display. Curr Opin Struct Biol. 17 (4), 474-480 (2007).
- Lener, M., et al. Diverting a protein from its cellular location by intracellular antibodies. Eur. J. Biochem. 267 (4), 1196-1205 (2000).
- Lynch, S. M., Zhou, C., Messer, A. An scFv intrabody against the nonamyloid component of α-synuclein reduces intracellular aggregation and toxicity. J. Mol. Biol. 377 (1), 136-147 (2008).
- Gai, S. A., Wittrup, K. D. Yeast surface display for protein engineering and characterization. Curr. Opin. Struc. Biol. 17 (4), 467-473 (2007).
- Kieke, M. C., et al. Selection of functional T cell receptor mutants from a yeast surface-display library. Proc. Natl. Acad. Sci. USA. 96 (10), 5651-5656 (1999).
- Steiner, D., Forrer, P., Stumpp, M. T., Pluckthun, A. Signal sequences directing cotranslational translocation expand the range of proteins amenable to phage display. Nat. Biotechnol. 24, 823-831 (2006).
- Pugsley, A. P. The complete general secretory pathway in gram-negative bacteria. Microbiol. Rev. 57 (1), 50-108 (1993).
- Rapoza, M. P., Webster, R. E. The filamentous bacteriophage assembly proteins require the bacterial SecA protein for correct localization to the membrane. J. Bacteriol. 175 (6), 1856-1859 (1993).
- Karlsson, A. J., et al. Engineering antibody fitness and function using membrane-anchored display of correctly folded proteins. J. Molec. Biol. 416 (1), 94-107 (2012).
- DeLisa, M. P., Tullman, D., Georgiou, G. Folding quality control in the export of proteins by the bacterial twin-arginine translocation pathway. Proc Natl Acad Sci U S A. 100 (10), 6115-6120 (2003).
- Fisher, A. C., Kim, W., DeLisa, M. P. Genetic selection for protein solubility enabled by the folding quality control feature of the twin-arginine translocation pathway. Protein Sci. 15 (3), 449-458 (2006).
- Maynard, J., Georgiou, G. Antibody engineering. Annu Rev Biomed Eng. 2, 339-376 (2000).
- Green, M. R., Sambrook, J. Molecular Cloning: A Laboratory Manual. 1, Fourth, Cold Spring Harbor Laboratory Press. (2012).
- Milo, R., Jorgensen, P., Moran, U., Weber, G., Springer, M. BioNumbers--the database of key numbers in molecular and cell biology. Nucleic Acids Res. 38, D750-D753 (2010).
- Hermanson, G. T. Bioconjugate Techniques. , Third, Elsevier/Academic Press. (2013).
- Tayapiwatana, C., Chotpadiwetkul, R., Kasinrerk, W. A novel approach using streptavidin magnetic bead-sorted in vivo biotinylated survivin for monoclonal antibody production. J Immunol Methods. 317 (1-2), 1-11 (2006).
- Zhu, G., Song, L., Lippard, S. J. Visualizing inhibition of nucleosome mobility and transcription by cisplatin-DNA interstrand crosslinks in live mammalian cells. Cancer Res. 73 (14), 4451-4460 (2013).
- Bogsch, E. G., et al. An essential component of a novel bacterial protein export system with homologues in plastids and mitochondria. J. Biol. Chem. 273, 18003-18006 (1998).
- Julian, M. C., et al. Co-evolution of affinity and stability of grafted amyloid-motif domain antibodies. Protein Eng. Des. Sel. 28 (10), 339-350 (2015).
- Garber, K. Bispecific antibodies rise again. Nat. Rev. Drug. Discov. 13 (11), 799-801 (2014).
- Marschall, A. L., Dübel, S., Böldicke, T. Specific in vivo knockdown of protein function by intrabodies. MAbs. 7 (6), 1010-1035 (2015).
